Introduction
Mechanical ventilation (MV) is an important therapy
used to assist critically ill patients with moderate or severe
ARDS. However, mechanical ventilation can induce or aggravate lung
injury, which is referred to as ventilator-induced lung injury
(VILI) (1). Increased infiltration
of inflammatory cells, alveolar edema, and barrier dysfunction due
to excessive mechanical stresses generated during mechanical
ventilation have been recognized as the major pathophysiologic
events of VILI (2). A growing body
of evidence suggests that VILI is essentially an excessive,
uncontrolled inflammatory response in the lung (3,4).
Despite the existence of lung protective ventilator strategies for
improving the ventilation procedures, the morbidity and mortality
of VILI have remained high (5).
Thus, the investigation of the precise molecular mechanism of VILI
is urgently required.
Although the mechanism responsible for initiating
VILI is not completely understood, it is believed that inflammatory
responses are key factors (6).
Studies (7,8) have revealed that disturbance in
intracellular ion homeostasis is a major event eliciting both the
inflammatory responses and production of inflammatory mediators,
such as IL-1β and TNF-α. Some studies (9,10)
have suggested that a Ca2+ influx mediates the
augmentation of many different inflammation and autoimmune
diseases. Notably, Ca2+ influx has been proposed to
perform an essential function in the development of VILI (11); in contrast, the inhibition of
cation channels protects against VILI (12). These studies imply that
Ca2+ mobilization plays a critical role in the
progression of VILI.
Calcineurin is a calcium-activated protein
phosphatase and an essential nodal factor in the regulation of
cellular functions (13). An
increase in the levels of intracellular Ca2+ activates
calcineurin. Activated calcineurin dephosphorylates the nuclear
factor of activated T cells (NFAT), which in turn modulates the
transcription of target genes (14). In recent years, various
inflammatory cytokines and adhesion molecules have been identified
as NFAT regulatory targets (15).
These molecules include VCAM-1, ICAM-1, IL-6, IL-8 and MCP-1. The
inhibition of NFAT was revealed to be an effective method for
reducing the multiple inflammatory cytokines induced by TNF-α in
human retinal microvascular endothelial cells (16), thus further highlighting NFAT
signaling as a potential anti-inflammatory target. Recent studies
(17,18) have revealed that NFATc3 is a key
molecular regulator of sepsis-induced lung injury. Notably, NFATc4
has been suggested to be a pivotal regulatory event in endothelial
cell inflammation (15), which is
an early step in the development of lung injury. Therefore, the
calcineurin/NFATc4 signaling pathway has also received considerable
attention due to its fundamental role in mediating lung
injuries.
However, whether calcineurin/NFATc4 signaling is
involved in the development of VILI remains unknown. Thus, it was
determined whether the calcineurin/NFATc4 signaling pathway
contributes to VILI.
Materials and methods
Animals
Adult male Wistar rats (weighing 200–230 g) were
obtained from the Comparative Medicine Center of YangZhou
University. All rats were housed in air-filtered,
temperature-controlled units with access to chow and water. All
experimental operations were performed with approval by and in
accordance with the guidelines set by the Animal Ethics Committee
of YangZhou University. The rats were anesthetized by
intraperitoneal injection with ketamine hydrochloride (50 mg/kg)
and xylazine (10 mg/kg).
In the first series of experiments, the rats were
randomly divided into the following three groups: The nonventilated
control group (Control, n=8); the ventilated with a low tidal
volume (6 ml/kg ventilation) for 6-h group (LV, n=8); and the
ventilated with a high tidal volume (30 ml/kg ventilation) for 6-h
group (HV, n=8).
In the second series of experiments, the rats were
randomly divided into the following four groups: the nonventilated
control group (Control, n=8); the pretreated with cyclosporine A
group (CsA, n=8); the ventilated with a high tidal volume (30 ml/kg
ventilation) for 6-h group (HV, n=8); and the HV-treated with
cyclosporine A group (HV + cyclosporine A, n=8).
The CsA-treated rats were administered cyclosporine
A (Abcam) (7 mg/kg), which is a selective inhibitor of calcineurin,
by intraperitoneal injection. The HV + cyclosporine A-treated rats
were intraperitoneally administered/pretreated with cyclosporine A
(10 mg/kg) 1 h before MV. Upon completion of the experiments, rats
were anesthetized with i.p. ketamine hydrochloride (50 mg/kg) and
xylazine (10 mg/kg) and sacrificed by exsanguinations from their
inferior vena cava.
Induction of VILI
The rats underwent tracheotomies and were intubated
with an 18-G angiocatheter. Then, the rats were connected to a
volume-limited rodent ventilator (DW3000; Zhenghua Biologic
Apparatus Facilities Co., Ltd.). The rats were mechanically
ventilated for 6 h using either a low tidal volume (VT=6 ml/kg) or
a high tidal volume (VT=30 ml/kg) with a positive end-expiratory
pressure of 0 cm H2O, a respiratory rate of 40
breaths/min and an FIO2 of 21% (Fig. 1).
Assessment of lung histology
The lung tissues were fixed and stained with
hematoxylin and eosin (H&E) as previously described (5). Two investigators blinded to group
assignments analyzed the samples and determined levels of lung
injury according to the semiquantitative scoring outlined below.
All lung fields at an ×20 magnification were examined for each
sample. Assessment of histological lung injury was performed
according to combined assessments of alveolar septal thickening,
inflammatory infiltration, microatelectasis or alveolar
overdistension, and microhemorrhage. Neutrophils were counted in
the alveolar airspaces, alveolar walls, and interstitium. The
injury scores were as follows: 0, absent with normal appearance; 1,
slight injury (<30% lung section); 2, intermediate injury
(30–60% lung section); and 3, severe injury (>60% lung
section).
Measurement of protein
concentrations
The bronchoalveolar lavage fluid (BALF) was
harvested by injecting and retrieving sterile PBS three times
through the tracheal intubation. The BALF (4 ml) was centrifuged at
4°C and 300 × g for 10 min. The total protein concentrations in the
BALF were determined using a bicinchoninic acid (BCA) protein assay
kit (Beyotime Institute of Biotechnology) according to the
manufacturer's instructions.
Wet-to-dry lung weight ratio
The right upper lobe of the lungs was weighed to
measure the wet weight. The dry weight was determined after heating
the lungs at 80°C for 48 h.
Myeloperoxidase (MPO) activity
bioassay
MPO can be used as an index of lung neutrophil
infiltration. The MPO activity in the lung tissues was assayed by
using an MPO assay kit (Jiancheng Bioengineering Institute of
Nanjing). Briefly, the lung tissues were homogenized and
centrifuged at 12,000 × g and 4°C for 10 min. An MPO activity assay
was performed at 450 nm to calculate the MPO activity.
RT-PCR
The total RNA in the lung tissue was harvested using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA
was synthesized using qRT SuperMix (Vazyme, JiangSu, China).
RT-qPCR was performed with AceQ SYBR-Green Master Mix (Vazyme). The
following primers were used for PCR analysis: ICAM-1 forward,
5′-GTGATGCTCAGGTATCCATCCA-3′ and reverse,
5′-CACAGTTCTCAAAGCACAGCG-3′; VCAM-1 forward,
5′-TTGGGAGCCTCAACGGTACT-3′ and reverse,
5′-GCAATCGTTTTGTATTCAGGGGA-3′; TNF-α forward,
5′-GGAACACGTCGTGGGATAATG-3′ and reverse,
5′-GGCAGACTTTGGATGCTTCTT-3′; IL-8 forward,
5′-AGCAGTCCCAACTAAGCAGTA-3′ and reverse,
5′-CAGCCAGTAGAGGATGCTGA-3′; and β-actin forward,
5′-GAGTCCTACGACATCATCGCT-3′ and reverse,
5′-CGTCCGACATAGTTTGGGAAA-3′. The cycle threshold (CT) method was
used to analyze the data in each experimental group (19).
Calcineurin activity analysis
The lung tissues of the rats were weighed and
homogenized in RIPA lysis buffer (P0013B, Beyotime Institute of
Biotechnology) for 30 min; then, the total protein levels were
measured using a BCA assay. The protein lysates were used
immediately for the CaN activity assay; specifically, the
Calcineurin Activity Assay Kit (Nanjing Jiancheng Bioengineering
Institute) was used.
Preparation of cytoplasmic and nuclear
extracts
The isolation of the cytoplasmic and nuclear
extracts was performed using a cytoplasmic and nuclear protein
extraction kit (cat. no. R0050; Solarbio Life Sciences). Briefly,
50 mg lung tissues were weighed, homogenized and centrifuged in
ice-cold PBS at 500 × g for 3 min. Then, the sediments were
dissolved in 200 µl of cytoplasmic extraction reagent by vortexing,
and centrifuged at 12,000 × g for 10 min at 4°C. The supernatant
was harvested as the cytoplasmic fraction. The sediments were
re-dissolved in 100 µl of nuclear extraction buffer by vortexing
for 15 sec and incubated on ice for 10 min, and then centrifuged
for 10 min at 16,000 × g at 4°C. Then, the supernatant was
harvested as the nuclear fraction. The concentrations of each
extract were assessed using a BCA protein assay kit (Beyotime
Institute of Biotechnology).
Western blotting
Western blotting was performed as previously
described (20). Equal amounts (20
µl) of sample buffer were loaded onto a 10% SDS-PAGE gel. The
protein was electrotransferred to polyvinylidene difluoride (PVDF)
membranes. Then, the membranes were incubated with the following
primary antibodies at 4°C for 12 h: Anti-inhibitors against NFATc4
(ab99431, 1:1,000; Abcam), VCAM-1 (ab134047, 1:1,000; Abcam),
ICAM-1 (10020-1-AP, 1:1,000; Proteintech), IL-8 (ab7747, 1:1,000;
Abcam), TNF-α (17590-1-AP, 1:1,000; Proteintech), Lamin B (AF1408,
1:1,000; Beyotime Institute of Biotechnology) and β-actin
(20536-1-AP 1:1,000; Proteintech). The membranes were incubated at
room temperature for 1 h with a peroxidase-conjugated secondary
antibody (1:2,000; Beyotime Institute of Biotechnology). The
protein bands were visualized using a ChemiDoc imaging system
(Bio-Rad Laboratories, Inc.), and the intensity was quantified
using ImageJ 1.8.0 (National Institutes of Health).
Statistical analysis
The data are presented as the mean ± standard
deviation (SD). The statistical analysis was performed via one-way
analysis of variance with Bonferroni multiple comparison tests
using GraphPad Prism V.5.01 (GraphPad, Inc.). The data were
considered to be statistically significant when the P-value was
<0.05.
Results
MV induces lung injury and
inflammatory responses
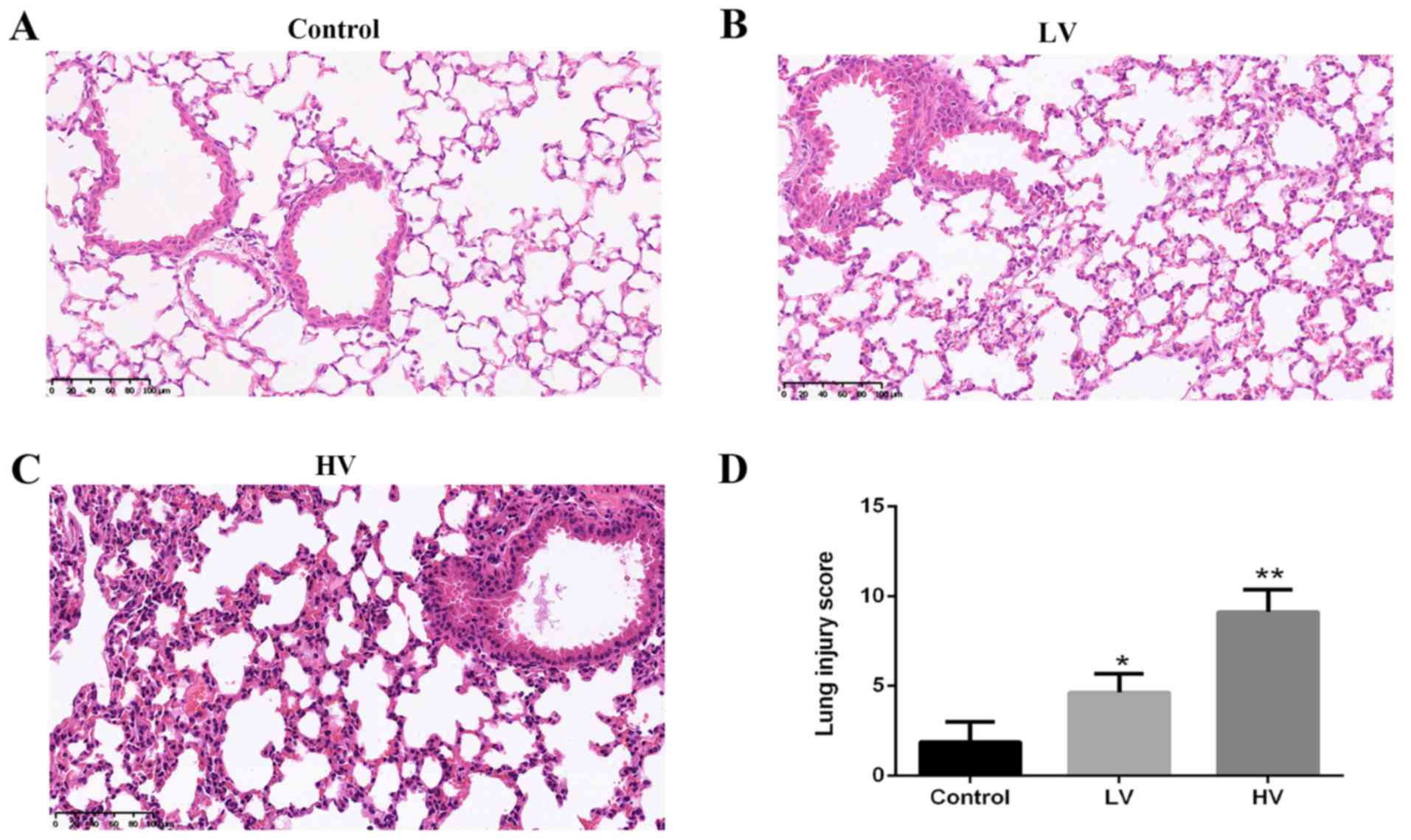
A histologic examination was performed to evaluate
the lung tissue damage induced by MV. Consistent with previous
studies, the results revealed severe lung injury characterized by
neutrophil accumulation and edema disruption in the lung tissues in
the HV group. The histological injury score in the HV group was
markedly higher than that in the control group (P<0.01)
(Fig. 2A-D).
To further clarify the effect of MV in lung injury,
the changes in the wet/dry ratios and the total protein
concentrations were investigated. The results demonstrated that MV
induced a significant increase in the wet/dry ratios (P<0.01)
(Fig. 3A) and total protein
concentrations of BALF (P<0.01) (Fig. 3B).
 | Figure 3.MV induces edema and an inflammatory
response in the lungs of rats. After the rats were treated with MV
(6 ml/kg or 20 ml/kg), pulmonary edema was determined by (A) the
wet/dry weight ratio and (B) the protein levels in BALF. (C) Lung
MPO activity was also assessed. The mRNA levels of (D) ICAM-1, (E)
VCAM-1, (F) TNF-α and (G) IL-8 were assessed. (H) The protein
expression of VCAM-1, ICAM-1, TNF-α and IL-8 were assessed. Blots
are representative of at least three independent experiments. (I)
Semi-quantification of immunoblots shown in (H). The data are
presented as the means ± SD (n=8). *P<0.05, **P<0.01,
compared with the Control group. MV, mechanical ventilation; LV,
ventilated with a low tidal volume; HV, ventilated with a high
tidal volume. |
To further confirm the effects of MV in the
inflammatory reactions, the expression levels of adhesion molecules
and proinflammatory cytokines and MPO activity were examined.
Significant regulation of ICAM-1, VCAM-1, TNF-α and IL-8 in the HV
group rats was observed (P<0.01) (Fig. 3D-I). Similarly, it was revealed
that MV promoted neutrophil infiltration into the lung tissues,
which was reflected by the increased expression of lung MPO
activity (Fig. 3C). These results
provided indirect evidence of the involvement of the inflammatory
response.
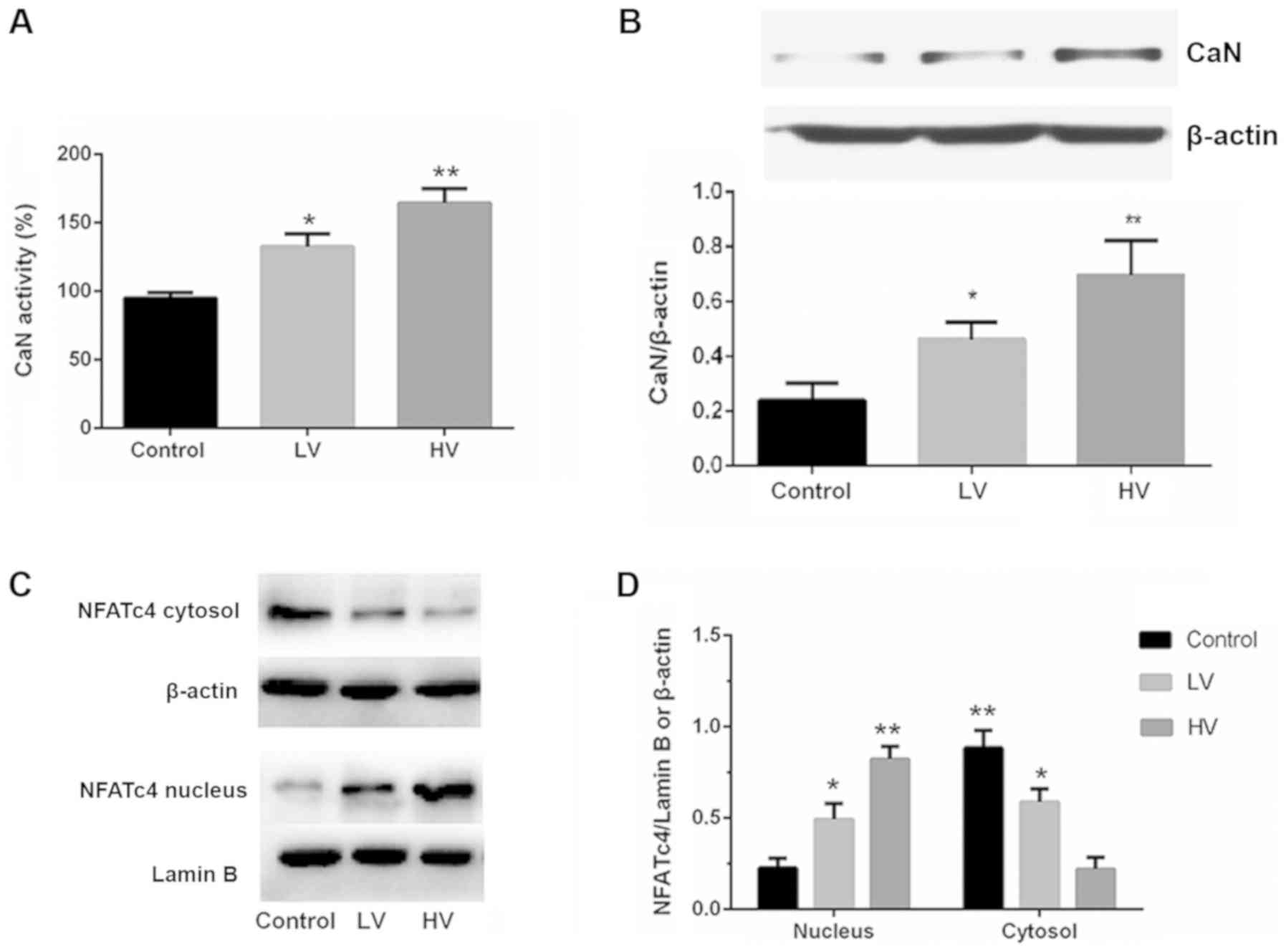
MV activates the calcineurin/NFAT4
signaling pathway in the lungs
Calcineurin is a calcium-activated protein
phosphatase. Activated calcineurin dephosphorylates NFATs, which
then translocate to the cell nucleus to regulate the transcription
of NFAT-responsive genes. To investigate whether the
calcineurin/NFATc4 pathway influences VILI, the activation of the
calcineurin/NFATc4 signaling pathway was examined. Fig. 4A and B demonstrated that the
activity and protein expression of CaN were significantly increased
after MV (P<0.01).
Subsequently, the effects of MV on the
phosphorylation and distribution of NFATc4 were examined. As
revealed in Fig. 4C and D, the
levels of the nuclear translocation of NFATc4 were increased, and
the levels of the cytoplasmic phosphorylation and translocation of
NFATc4 were also decreased. These results indicate that MV exposure
could activate lung calcineurin/NFAT4 signaling.
VILI is suppressed by
calcineurin/NFATc4 signaling antagonists
To further confirm the role of calcineurin/NFATc4
pathway activation in VILI, lung injury after VILI in the presence
of calcineurin antagonists was examined. The treatment with the
calcineurin antagonists significantly reduced the VILI pathological
changes (P<0.05), the wet/dry ratios (P<0.05), the total
protein concentrations in BALF and MPO activity (P<0.05)
(Fig. 5A-E). In contrast, the lung
injury in the naive rats was not affected by the treatment with
CsA), indicating that CsA improved VILI but did not affect normal
lung tissue. These results indicated that the calcineurin/NFATc4
pathway plays an essential role in VILI.
Inhibition of calcineurin/NFATc4
signaling suppresses the inflammatory response of MV
Since inflammatory responses play an important role
in VILI, whether the administration of calcineurin inhibitors could
reverse the increase in the expression levels of adhesion molecules
and proinflammatory cytokines was investigated. The results
demonstrated that the pretreatment with CsA prevented the increases
in ICAM-1, VCAM-1, TNF-α and IL-8 levels (P<0.05) (Fig. 6A-F). These results further
indicated that the calcineurin/NFATc4 pathway may be involved in
the inflammatory responses of VILI.
Discussion
The present study revealed that calcineurin/NFATc4
signaling plays a critical role in the development of VILI. The
principle findings are as follows: i) MV caused the activation of
the calcineurin/NFATc4 pathway, which was associated with the
degree of lung injury in the rats; ii) calcineurin/NFATc4 signaling
activation in the lungs led to increased levels of adhesion
molecules and proinflammatory cytokines; iii) the inhibition of the
calcineurin/NFATc4 signaling pathway attenuated VILI and the
release of pro-inflammatory cytokines. Collectively, the present
results highlighted the critical role of the calcineurin/NFATc4
signaling pathway in VILI.
Although essential in critically ill patients,
mechanical ventilation often induces or aggravates lung injury,
which is termed VILI (21).
Accumulating studies (22,23) have revealed that VILI is mainly
characterized by increases in pulmonary vascular permeability,
influxes of inflammatory cells and various cytokines. These
inflammatory cells could produce inflammatory cytokines.
Furthermore, these cytokines could facilitate the infiltration of
inflammatory cells (24).
Therefore, the inflammatory response is critical for the
development of VILI. However, the mechanisms of VILI remain
unclear. In the present study, it was demonstrated that the
calcineurin/NFAT4 signaling pathway was activated during VILI. A
high-VT MV for 6 h activated both calcineurin and NFATc4;
conversely, the blockage of the calcineurin/NFATc4 signaling
pathway with cyclosporine A attenuated VILI. These findings provide
strong evidence that the calcineurin/NFATc4 signaling pathway may
be among the most important signaling pathways activated by MV and,
thus, is involved in the development of VILI.
Calcineurin is a calcium-activated protein
phosphatase that has been implicated in various inflammatory
diseases (25). Activated
calcineurin dephosphorylates the nuclear factor of activated T
cells (NFAT), which, in turn, modulates the transcription of target
genes associated with cellular proliferation, differentiation,
inflammation and angiogenesis (14). The NFAT protein family consists of
the following five members: NFATc1, NFATc2, NFATc3, NFATc4 and
NFAT5 [tonicity-responsive enhancer-binding protein (TonEBP)]
(26). NFATc3 has been revealed to
play an important role in the activation of macrophages during
sepsis (17,18). Collectively, these experimental
studies indicated that the calcineurin/NFATc4 signaling pathway
plays a fundamental role in mediating inflammation in lung injury.
The present study extended the understanding of the important role
of calcineurin/NFATc4 signaling in lung injury by demonstrating
that the calcineurin/NFATc4 pathway contributes to VILI.
In the present study, it was observed that MV
induced the calcineurin/NFATc4-dependent production of adhesion
molecules and pro-inflammatory cytokines. These adhesion molecules
and proinflammatory cytokines are known to serve as an important
regulatory event to deteriorate VILI (27). On the basis of the present
findings, it was speculated that the mechanical stresses generated
during mechanical ventilation can activate the calcineurin/NFATc4
signaling pathway, resulting in increases in adhesion molecules and
proinflammatory cytokines. However, whether these effects are
direct or indirect is unclear.
In conclusion, the present results indicated that
VILI depends on the activation of the calcineurin/NFATc4 pathway,
which then leads to increases in the release of adhesion molecules
and proinflammatory cytokines responsible for inducing the
activation of inflammation in the lungs. Since the present study
assessing the mechanism of VILI mainly focused on the
calcineurin/NFATc4 pathway, the role of other members of the NFAT
family has not been established. Therefore, other mechanisms may be
involved in VILI.
Acknowledgements
Not applicable.
Funding
The present study were supported by The National
Natural Science Fund (grant no. 81401626) and Research projects of
Subei People's Hospital (grant no. yzucms201714).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DS designed the study and the experiments. ML and XF
performed the majority of the experiments. ML wrote the manuscript.
XF and XL analyzed the data. YZ, YX, XM and XY helped with
experiments, especially in vitro cell cultures and western blot
analysis.
Ethics approval and consent to
participate
All experimental operations were performed with
approval by and in accordance with the guidelines set by the Animal
Ethics Committee of YangZhou University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Slutsky AS and Ranieri VM:
Ventilator-induced lung injury. N Engl J Med. 369:2126–2136. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cressoni M, Gotti M, Chiurazzi C, Massari
D, Algieri I, Amini M, Cammaroto A, Brioni M, Montaruli C, Nikolla
K, et al: Mechanical power and development of ventilator-induced
lung injury. Anesthesiology. 124:1100–1108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yildiz C, Palaniyar N, Otulakowski G, Khan
MA, Post M, Kuebler WM, Tanswell K, Belcastro R, Masood A,
Engelberts D and Kavanagh BP: Mechanical ventilation induces
neutrophil extracellular trap formation. Anesthesiology.
122:864–875. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fang XZ, Huang TF, Wang CJ, Ge YL, Lin SY,
Zhang Y and Gao J: Preconditioning of physiological cyclic stretch
attenuated HMGB1 expression in pathologically mechanical
stretch-activated A549 cells and ventilator-induced lung injury
rats through inhibition of IL-6/STAT3/SOCS3. Int Immunopharmacol.
31:66–73. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Park SY, Kim HJ, Yoo KH, Park YB, Kim SW,
Lee SJ, Kim EK, Kim JH, Kim YH, Moon JY, et al: The efficacy and
safety of prone positioning in adults patients with acute
respiratory distress syndrome: A meta-analysis of randomized
controlled trials. J Thorac Dis. 7:356–367. 2015.PubMed/NCBI
|
|
6
|
Wang T, Gross C, Desai AA, Zemskov E, Wu
X, Garcia AN, Jacobson JR, Yuan JX, Garcia JG, Black SM, et al:
Endothelial cell signaling and ventilator-induced lung injury:
Molecular mechanisms, genomic analyses, and therapeutic targets. Am
J Physiol Lung Cell Mol Physiol. 312:L452–L476. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang T, Lang X, Xu C, Wang X, Gong T, Yang
Y, Cui J, Bai L, Wang J, Jiang W and Zhou R: CLICs-dependent
chloride efflux is an essential and proximal upstream event for
NLRP3 inflammasome activation. Nat Commun. 8:2022017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schappe MS, Szteyn K, Stremska ME, Mendu
SK, Downs TK, Seegren PV, Mahoney MA, Dixit S, Krupa JK, Stipes EJ,
et al: Chanzyme TRPM7 mediates the Ca2+ influx essential
for lipopolysaccharide-induced toll-like receptor 4 endocytosis and
macrophage activation. Immunity. 48:59–74.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu MC, Yu CL, Chen HC, Yu HC, Huang HB and
Lai NS: Aberrant T cell expression of Ca2+ influx-regulated miRNAs
in patients with systemic lupus erythematosus promotes lupus
pathogenesis. Rheumatology. 54:343–348. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guo Y, Yang X, He J, Liu J, Yang S and
Dong H: Important roles of the Ca2+-sensing receptor in
vascular health and disease. Life Sci. 209:217–227. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Michalick L, Erfinanda L, Weichelt U, van
der Giet M, Liedtke W and Kuebler WM: Transient receptor potential
vanilloid 4 and serum glucocorticoid-regulated kinase 1 are
critical mediators of lung injury in overventilated mice in vivo.
Anesthesiology. 126:300–311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pairet N, Mang S, Fois G, Keck M, Kühnbach
M, Gindele J, Frick M, Dietl P and Lamb DJ: TRPV4 inhibition
attenuates stretch-induced inflammatory cellular responses and lung
barrier dysfunction during mechanical ventilation. PLoS One.
13:e01960552018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shibasaki F, Hallin U and Uchino H:
Calcineurin as a multifunctional regulator. J Biochem. 131:1–15.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li S, Pan Y, Ke R, Xie X, Zhai C, Shi W,
Wang J, Yan X, Chai L, Wang Q, et al: Inhibition of
phosphodiesterase-5 suppresses calcineurin/NFAT-mediated TRPC6
expression in pulmonary artery smooth muscle cells. Sci Rep.
7:60882017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu BX, Yuan JN, Zhang FR, Liu YY, Zhang
TT, Li K, Lv XF, Zhou JG, Huang LY, Shang JY and Liang SJ:
Inhibition of Orai1-mediated Ca2+ entry limits
endothelial cell inflammation by suppressing calcineurin-NFATc4
signaling pathway. Biochem Biophys Res Commun. 495:1864–1870. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Savage SR, Bretz CA and Penn JS: RNA-Seq
reveals a role for NFAT-signaling in human retinal microvascular
endothelial cells treated with TNFα. PLoS One. 10:e01169412015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Karpurapu M, Lee YG, Qian Z, Wen J,
Ballinger MN, Rusu L, Chung S, Deng J, Qian F, Reader BF, et al:
Inhibition of nuclear factor of activated T cells (NFAT) c3
activation attenuates acute lung injury and pulmonary edema in
murine models of sepsis. Oncotarget. 9:10606–10620. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ranjan R, Deng J, Chung S, Lee YG, Park
GY, Xiao L, Joo M, Christman JW and Karpurapu M: The transcription
factor nuclear factor of activated T cells c3 modulates the
function of macrophages in sepsis. J Innate Immun. 6:754–764. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Q, Ge YL, Li M, Fang XZ, Yuan YP, Liang
L and Huang SQ: miR-127 contributes to ventilator-induced lung
injury. Mol Med Rep. 16:4119–4126. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang R, Pan Y, Fanelli V, Wu S, Luo AA,
Islam D, Han B, Mao P, Ghazarian M, Zeng W, et al: Mechanical
stress and the induction of lung fibrosis via the midkine signaling
pathway. Am J Respir Crit Care Med. 192:315–323. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iwaki M, Ito S, Morioka M, Iwata S,
Numaguchi Y, Ishii M, Kondo M, Kume H, Naruse K, Sokabe M and
Hasegawa Y: Mechanical stretch enhances IL-8 production in
pulmonary microvascular endothelial cells. Biochem Biophys Res
Commun. 389:531–536. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lin JY, Jing R, Lin F, Ge WY, Dai HJ and
Pan L: High tidal volume induces mitochondria damage and releases
mitochondrial DNA to aggravate the ventilator-induced lung injury.
Front Immunol. 9:14772018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Curley GF, Laffey JG, Zhang H and Slutsky
AS: Biotrauma and ventilator induced lung injury: Clinical
implications. Chest. 150:1109–1117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ni C, Li Z, Qian M, Zhou Y, Wang J and Guo
X: Isoflurane induced cognitive impairment in aged rats through
hippocampal calcineurin/NFAT signaling. Biochem Biophys Res Commun.
460:889–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miyakawa H, Woo SK, Dahl SC, Handler JS
and Kwon HM: Tonicity-responsive enhancer binding protein, a
rel-like protein that stimulates transcription in response to
hypertonicity. Proc Natl Acad Sci USA. 96:2538–2542. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Imanaka H, Shimaoka M, Matsuura N,
Nishimura M, Ohta N and Kiyono H: Ventilator-induced lung injury is
associated with neutrophil infiltration, macrophage activation, and
TGF-beta 1 mRNA upregulation in rat lungs. Anesth Analg.
92:428–436. 2001. View Article : Google Scholar : PubMed/NCBI
|