Introduction
The association of the consumption of a high-fat
diet (HFD) or obesity promoting diet with metabolic disorders, such
as obesity, diabetes, hypertension and cardiovascular diseases, has
long been recognized (1). A large
number of studies have suggested that an HFD also affects the
severity of tumorigenesis in the gut, particularly in colon cancer,
independent of obesity (2).
However, little is known regarding changes in the intestinal
epithelium in response to HFD ingestion, both in the small
intestine and colon.
The intestinal tract is the first point of contact
of ingested food, and thus disturbances in gut homeostasis may
represent the first point in a chain of events resulting in
diseases associated with HFD (3).
An increasing number of studies have demonstrated that changes in
diet are manifested quickly as changes to the composition of the
gut microbiota (4,5), whose composition is associated with a
variety of diseases (6,7). Diet, gut microbiota and host
interactions that occur at intestinal epithelial cells (IECs) serve
pivotal roles in shaping the microbial ecology (8). Indeed, IECs constitute a barrier
surface that separates the host from the external environment and
conserves the host-microbial interactions and tissue homeostasis
(9). Therefore, IECs are crucial
mediators that maintain the host-commensal microbial relationship
and immune cell homeostasis in the intestine. Therefore, the aim of
the present study was to determine the effects of an HFD on the
function of IECs.
Weight gain accompanied by an increased tendency for
central fat distribution is more common among women than in men,
particularly in middle-aged individuals, with an age of
approximately 45–64 years. Bjerregaard et al (10) reported that being overweight at the
age of 7 years was associated with an increased risk of developing
type 2 diabetes as an adult only if the individual continued to be
overweight until puberty or at a later age. Therefore, weight gain
in middle-aged individuals is more harmful and more closely
associated with cardiovascular diseases and type 2 diabetes
(10). Previously, HFD models were
induced in mice with an age of approximately 2–3 months, and thus
the effects of aging on disease progression have rarely been taken
into consideration.
Therefore, in the present study, middle-aged female
mice (12-month-old) were fed an HFD for a period of 14 weeks to
investigate how HFD influenced the gut pathophysiology, as well as
obesity-associated metabolic dysfunction and disorders. The results
revealed that HFD increased the intestinal stem cell (ISC) counts
and crypt function in the small intestine and colon, and
compromised the epithelial barrier function of the colon. These
findings may be helpful in understanding how an HFD influences the
intestinal epithelium in maintaining tissue homeostasis and
suggested the importance of careful consideration of dietary
habits.
Materials and methods
Animal studies
A total of 14 female C57BL/6J mice were purchased
from the Model Animal Research Center of Nanjing University
(Nanjing, China). At 12 months of age and at an average weight of
32.0 g, the mice were randomly assigned to the regular diet (n=6)
or HFD (n=8) group and provided their respective diet for 14 weeks.
The HFD consisted of 60% calories as fat, 20% as carbohydrate and
20% as protein. Water was freely available at all times. Mice were
housed at 23±1°C with an average humidity of 60±1% and a 12-h
light/dark cycle. The body weight and food intake of animals were
measured weekly. At the end of the feeding period, mice were
anesthetized with intraperitoneal injection of sodium pentobarbital
at a dose of 50–90 mg/kg of body weight and then sacrificed by
cervical dislocation, followed by additional removal of the heart
to ensure death. The experimental protocols of the present study
were approved by the Animal Care and Use Committee of Nanjing
Medical University (Nanjing, China), and conducted in accordance
with the guidelines of this committee.
Oral glucose tolerance test (oGTT) and
insulin tolerance test (ITT)
For oGTT, mice were fasted overnight (14–18 h) and
then given a glucose load (25% stock solution in saline) of 2 g per
kg of body weight by oral administration. For ITT, intraperitoneal
injection of an insulin bolus of 4 IU per kg of body weight was
performed. Blood samples were collected from the tail vein at 0,
15, 30, 60 and 120 min after administration of glucose or insulin.
Plasma glucose concentration was measured using an Accu-Chek Aviva
system (Roche Diagnostics).
Cell staining, immunohistochemical and
immunofluorescence assays
Mice were weighed and euthanized, and then the small
intestine and colon were removed. Next, the lengths of the small
intestine (from the pylori to the ileocecal junction) and the colon
(from the cecum to the rectum) were measured. From the small
intestine, the initial 5 cm from the pylorus associated with the
pancreas was discarded in order to avoid autolytic changes. The
following 1.5 cm corresponding to the portion of proximal jejunum,
which represents the majority of the small intestine, was
collected. For the colon, 1 cm at the middle of the colon was
dissected for histological analysis. The small intestine and colon
samples were fixed in Carnoy's solution composed of ethanol and
glacial acetic acid (3:1) at room temperature for 30 min and stored
overnight at 4°C. The fixed intestine was cut transversely into
three (small intestine) or two (colon) 0.5 cm segments and embedded
in paraffin. Subsequently, paraffin-embedded tissues were sliced
into 6-µm sections for cross-sectional histological analysis.
Tissue sections were deparaffinized with xylene and rehydrated in a
descending alcohol series of 100, 95, 80, 70 and 50%. For
heat-induced antigen retrieval, the sections were incubated twice
in antigen retrieval buffer (0.1 M sodium citrate, 0.1 M citric
acid, pH 6.0) for 10 min at 90–100°C. These initial processing
steps were the same for all the staining procedures described below
and all staining procedures were performed on the same samples, but
different sections (3–5 sections used for each staining
technique).
Hematoxylin and eosin (H&E) staining was
performed for morphological analyses using standard histological
protocols (11). Briefly, tissue
sections were stained in hematoxylin solution for 8 min at room
temperature and counterstained with eosin solution for 30–60 sec at
room temperature. Stained sections were observed under a light
microscope at ×40 magnification. The villus length was measured
from the base to the top of the villi, and the crypt depth was
measured between the crypt-villus junction and the base of the
crypt.
Goblet cell quantification was performed using the
histochemical Periodic Acid-Schiff (PAS) staining kit (Baso Biotech
Co., Ltd.) according to the manufacturer's protocol. Briefly,
tissue sections were sequentially stained with Alcian blue solution
(pH 2.5) for 10 min, Periodic acid solution for 10 min, Schiff
reagent for 20 min and Mayer's hematoxylin solution for 5 min, all
at room temperature. Stained cells were observed using a light
microscope at ×20 magnification.
For immunohistochemical analysis, following the
initial processing steps, sections were blocked with 7.5% goat
serum (Boster Biological Technology) for 30 min at room
temperature. Subsequently, sections were incubated overnight at 4°C
with the following primary antibodies: Anti-Ki67 (1:200; cat. no.
GB13030-2; Wuhan Servicebio Technology Co., Inc.), anti-p21 Cip 1
(1:200; cat. no. DF6423; Affinity Biosciences) and anti-p53 (1:200;
cat. no. AF0879; Affinity Biosciences). Following the primary
antibody incubation, the slides were incubated with a horseradish
peroxidase-conjugated secondary antibody (1:300; cat. no. ZB-2301;
ZSGB-BIO; OriGene Technologies, Inc.) at 37°C for 20 min. Sections
were subsequently stained with 3′,3′-diaminobenzidine (DAB; 0.05%
DAB, 0.015% H2O2, 0.01 M PBS, pH 7.2) at room
temperature for 1–3 min and observed under a light microscope at
×40 magnification.
For immunofluorescence staining, the blocking
process was the same as described above for immunohistochemical
analysis. Tissue sections were incubated overnight at 4°C with the
following primary antibodies: Anti-zona occludens 1 (ZO-1) tight
junction protein (1:200; cat. no. ab216880; Abcam), anti-Claudin-2
(1:200; cat. no. ab53032; Abcam) and anti-Lgr5/GPR49 (1:200; cat.
no. ab NBP1-28904; Novus Biologicals, LLC) antibody. Following the
primary antibody incubation, the sections were subsequently
incubated with Cy™3-conjugated anti-rabbit immunoglobulin G heavy
and light chain secondary antibody (1:200; cat. no. 111-165-003;
Jackson Immuno Research Laboratories, Inc.) at room temperature for
1.5–2 h. The nucleus was counterstained using DAPI (1:1,000; cat.
no. D1306; Thermo Fisher Scientific, Inc.) for 5 min at room
temperature and the sections were mounted using
VECTASHIELD® antifade mounting medium (Vector
Laboratories, Inc.; Maravai LifeSciences) and a coverslip. The
fluorescence was observed under a fluorescent microscope at ×40
magnification.
TUNEL
A TUNEL assay (DeadEnd™ Fluorometric TUNEL System;
Promega Corporation) was conducted to measure the rate of apoptosis
according to the manufacturer's protocol. In addition to the
fixation and deparaffinization steps described above, tissue
sections were subsequently fixed with 4% formaldehyde in PBS for 15
min at room temperature. Sections were then permeabilized with 20
µg/ml Proteinase K solution for 10 min at room temperature and
fixed for a second time with 4% formaldehyde in PBS for 5 min at
room temperature. Next, 100 µl equilibration buffer was added to
the sections for 10 min at room temperature and sections were
subsequently incubated with TdT reaction mix for 60 min at 37°C in
the dark to label nuclear DNA fragmentation. Sections were immersed
in 2X SSC for 15 min at room temperature to stop the reaction. Cell
nuclei were counterstained with DAPI (1:1,000; cat. no. D1306;
Thermo Fisher Scientific, Inc.) for 5 min at room temperature and
mounted with VECTASHIELD® antifade mounting medium
(Vector Laboratories, Inc.; Maravai LifeSciences). TUNEL-positive
cells were observed under a fluorescent microscope at ×20
magnification and the number of crypts and crypt-villus units were
counted.
Dissection of interscapular brown
adipose tissue (iBAT), epididymal white adipose tissue (eWAT) and
inguinal white adipose tissue (iWAT)
Following euthanasia of the mice and removal of the
skin, iWAT in the inguinal triangular area, iBAT pads, which are
located near the shoulder blades, and eWAT pads, which are attached
to the gonads, were detached, weighed and snap frozen in liquid
nitrogen.
Isolation and culture of primary
intestinal crypts
Small intestines and colons were removed and opened
longitudinally. Next, the tissues were washed 4–5 times with
ice-cold PBS until they were clean, the villi of the small
intestine were scraped off, and the small intestine and colons were
sectioned into 5-mm sections (12,13).
Tissue fragments were subsequently transferred to Hank's balanced
salt solution without calcium and magnesium, containing 0.5 mM
dithiothreitol and 8 mM EDTA, and incubated on ice for 30–60 min.
Intestinal crypts were mechanically released from the lamina
propria by vigorous pipetting. The supernatants containing the
crypts were filtered through a 70-µm mesh and centrifuged at 100 ×
g for 5 min at 4°C. Isolated crypts were counted and embedded in
Matrigel (Corning, Inc.) on ice at ~10 crypts per µl. Matrigel
beads occupying the center of the well were constructed using 20 µl
of Matrigel to form a solid dome-like structure in a 24-well plate,
and were subsequently overlaid with 500 µl Advanced DMEM/F12 with
HEPES (Gibco; Thermo Fisher Scientific, Inc.) containing 1X N-2
Supplement (Thermo Fisher Scientific, Inc.), B27 1× (Thermo Fisher
Scientific, Inc.), 1X GlutaMAX™ Supplement (Gibco; Thermo Fisher
Scientific, Inc.), 1 µM N-acetyl-l-cysteine (Sigma-Aldrich; Merck
KGaA), 100 U/ml penicillin, 100 µg/ml streptomycin (Gibco; Thermo
Fisher Scientific, Inc.), 50 ng/ml EGF (PeproTech, Inc.), 1 µg/ml
R-spondin-1 (PeproTech, Inc.) and 100 ng/ml Noggin (PeproTech,
Inc.) (14–16). Colonic crypts were cultured with an
additional 100 ng/ml Wnt-3a (PeproTech, Inc.) (17). Primary intestinal crypts were
incubated in a fully humidified culture chamber with 5%
CO2 at 37°C. The culture medium was changed every 48 h,
and the organoid-forming efficiency was calculated on days 4 and
6.
Organoid immunofluorescence
staining
Organoids embedded in Matrigel were fixed with 2%
paraformaldehyde and 1% glutaraldehyde for 30 min at room
temperature, and the fixed Matrigel was then placed in 20% sucrose
overnight at 4°C. Subsequently, the Matrigel domes were collected
into O.C.T. compound (Sakura Finetek USA, Inc.), snap frozen and
cryosectioned into 10-µm sections. The sections were then incubated
for 15-min in 0.2 M glycine solution at room temperature to quench
the aldehyde groups. In order to permeabilize the slices, they were
placed in 0.15% Triton X-100 for 15 min at room temperature. Next,
the slices were incubated with an anti-lysozyme antibody (1:200;
Abcam; cat. no. ab108508) overnight at 4°C, followed by secondary
Cy™3-conjugated goat anti-rabbit IgG H&L antibody, as described
earlier. Organoids were counterstained with DAPI for 5 min at room
temperature and visualized using an epi-fluorescence microscope.
Images were obtained at ×40 or ×100 magnification.
Fluorescence-activated cell sorting
(FACs)
Intestinal crypts and villi were isolated by EDTA
chelation buffer treatment, as described earlier. The cell
suspensions, composed of crypts and villi in the small intestine,
which houses a single cell layer of epithelium from the submucosa,
were dissociated into single cells using TrypLE Express (Gibco;
Thermo Fisher Scientific, Inc.), and the cells were passed through
40-µm filters. Cells were fixed in 1X intracellular fixation buffer
(cat. no. 00-8222; eBioscience; Thermo Fisher Scientific, Inc.) for
30 min at 4°C and subsequently washed in 1X permeabilization buffer
(cat. no. 00-8333; eBioscience; Thermo Fisher Scientific, Inc.) for
10 min at 4°C. Cells were blocked at 4°C for 30 min in 0.5% BSA
(cat. no. 4240GR100; neoFroxx)/PBS blocking solution and
subsequently incubated with antibodies against surface markers at
4°C for 30 min: Allophycocyanin (APC)-conjugated Lgr5/GPR49 (0.5
µg/106 cells; R&D Systems, Inc.; cat. no.
FAB82401A), phycoerythrin (PE)-conjugated CD24 (1:200; BD
Biosciences; cat. no. 553262), PE-conjugated mucin 2 (Muc2; 1:200;
Santa Cruz Biotechnology, Inc.; cat. no. sc-515032), and
FITC-conjugated CD326 (EpCAM) antibodies (0.125 µg/106
cells; eBioscience; Thermo Fisher Scientific, Inc.; cat. no.
14-5791-81). Apoptosis was detected using propidium iodide (cat.
no. BMS500PI; eBioscience; Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. A BD FACSAria II SORP
cell sorter (BD Biosciences) was used to isolate different types of
cells: ISCs were isolated as Lgr5hiEpcam+
cells, progenitor cells were isolated as
Lgr5lowEpcam+, Paneth cells were isolated as
CD24hiSidescatter hiEpcam+ and
goblet cells were isolated as Muc2hiEpcam+
cells (18). Data analysis was
performed using FlowJo software (version 10; Tree Star, Inc.). All
RT-qPCR experiments were repeated ≥2 times.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from eWAT and intestinal
tissues using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol, and
the RNA concentration was measured at 260 and 280 nm using a
NanoDrop spectrophotometer (Thermo Fisher Scientific, Inc.). Total
RNA samples (2 µg) were then reverse transcribed into cDNA using
200 U M-MLV Reverse Transcriptase (Promega Corporation) in the
presence of 0.5 mM deoxynucleotide triphosphate, 25 U RNase
inhibitor and 10 mM random hexamer primers (Promega Corporation),
with a final volume of 25 µl. Samples were incubated at 37°C for 1
h, 95°C for 5 min and then placed in an ice water mixture for 5
min. qPCR was subsequently performed in triplicate using
SYBR® Green Real-Time PCR Master mix (Roche Diagnostics)
at a final volume of 25 µl, using an ABI 7000 StepOnePlus Real-Time
PCR system (Thermo Fisher Scientific, Inc.). The PCR thermal
cycling conditions were as follows: 10 min at 95°C; 40 cycles of 15
sec at 95°C and 60 sec at 60°C. The temperature was increased from
60°C to 95°C, and the PCR melting curve was plotted at every 1°C
following the amplification step. The PCR primers were designed
using Primer 5 software (Premier Biosoft) and are presented in
Table SI. Expression levels were
normalized to β-actin. The mean value of triplicate analyses for
each sample was calculated and expressed as the quantification
cycle threshold (Cq). Gene expression was then calculated as the
ΔCq, defined as the difference between the Cq value of the sample
and the Cq value of β-actin, which was used as an internal
reference. Finally, the relative expression levels were determined
using the comparative 2−ΔΔCq method (19).
Statistical analysis
All the data were analyzed with Microsoft Excel
(version 16.16.13; Microsoft Corporation) and GraphPad Prism
(version 6.0c; GraphPad Software, Inc.) software and the data was
presented as the mean ± SD. RT-qPCR experiments were repeated ≥2
times, whereas 3 mice/group were analyzed in the FACS assay, which
was repeated once. For histological analysis, including TUNEL,
30–40 crypt-villus units in the small intestine/section and 30–40
crypts in the colon/section were observed and assessed in a single
experiment. For organoid assays, 5–6 wells/group with 2–3 different
mice were analyzed in a single experiment. Statistical significance
was calculated using Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Characteristics of the HFD-fed
mice
In the present study, 12-month old C57BL/6J female
mice were fed an HFD for 14 weeks to establish an HFD-induced obese
mouse model (Fig. 1A). Metabolic
assessments were performed on the mice, which indicated that they
developed metabolic abnormalities. Mice fed an HFD had a
significantly increased body weight (Fig. 1B) after 14 weeks of the HFD
(P<0.01), while they were glucose intolerant (Fig. 1C) and insulin resistant (Fig. 1D) compared with the mice fed a
normal chow diet. In addition, the fat weight of the iBAT
(P<0.05), eWAT (P<0.05) and iWAT (P<0.05) increased
significantly in the HFD mice (Fig. 1E
and F). Furthermore, the mRNA expression levels of the
inflammatory factors F4/80 (P<0.05) and monocyte chemoattractant
protein-1 (MCP-1; P<0.001) were found to be upregulated in the
eWAT of the HFD mice (Fig. 1G and
H). These results demonstrated that certain physiological
characteristics were altered in the HFD mice and thus, an obese
mouse model was successfully generated.
 | Figure 1.Characteristics of middle-aged
(12-month-old) female C57BL/6 mice fed an HFD for 14-weeks. (A)
Representative appearance of Con and HFD mice. (B) Body weights
were determined weekly. (C) oGTT and (D) ITT results. (E)
Representative images of iBAT, eWAT and iWAT. (F) Tissue weight of
iBAT, eWAT and iWAT from the two mouse groups. mRNA expression
levels of (G) F4/80 and (H) MCP-1 inflammation-associated genes in
eWAT. Data are presented as the mean ± standard deviation.
*P<0.05, **P<0.01 and ***P<0.001, vs. Con group. HFD,
high-fat diet; Con, control; oGTT, oral glucose tolerance test;
ITT, insulin tolerance test; iBAT, interscapular brown adipose
tissue; eWAT, epididymal white adipose tissue; iWAT, inguinal white
adipose tissue. |
HFD alters the intestinal
morphology
In order to elucidate the effect of HFD on
intestinal function, the overall appearance of the intestine was
first evaluated. The length of the small intestine was not
significantly altered in the HFD mice (Fig. 2A and B). H&E staining was used
to analyze the intestinal tissue and to quantify the length of the
intestinal villi and the depth of intestinal crypts. In the small
intestine, the results revealed that the length of villi in the HFD
group was significantly shorter compared with that in the control
group (P<0.05; Fig. 2C and D),
although the crypt depth was not evidently altered (Fig. 2E). In the colon, the length was
significantly reduced (P<0.05; Fig.
2F and G) and the depth of the crypts was significantly shorter
in the HFD mice compared with the control group (P<0.05;
Fig. 2H and I). These data suggest
that the intestinal morphology was evidently altered in mice fed an
HFD, and thus the intestinal function may also be altered by this
diet.
 | Figure 2.HFD alters the intestinal morphology.
Mice were fed an HFD for 14-weeks, and then the small intestines
and colons were removed. (A) Representative images (scale bar, 2
cm), and (B) quantified length of the small intestine. (C)
Representative H&E staining images of the crypt and villi in
the jejunum (scale bar, 50 µm; magnification, ×40). (D) Villus
length and (E) crypt depth in the small intestine. (F)
Representative images of the colon of mice in the two groups (scale
bar, 2 cm), and (G) quantified colon length. (H) H&E staining
images of colonic crypt (scale bar, 50 µm; magnification, ×40), and
(I) quantified colonic crypt depth. Data are presented as the mean
± standard deviation. *P<0.05 vs. Con group. HFD, high-fat diet;
Con, control; H&E, hematoxylin and eosin. |
HFD increases the ISC count and alters
crypt function in the small intestine
To determine the effects of HFD on ISCs, flow
cytometry was performed to count Lgr5-high ISCs and Lgr5-low
progenitor cells in the small intestine. Mice fed an HFD had a
significantly increased number of ISCs (P<0.05) and progenitor
cells (P<0.0001) in the epithelium of the small intestine when
compared with the control group (Fig.
3A-C). The mRNA expression of Lgr5 in small intestine
epithelial tissues was also measured, and it was revealed that the
HFD increased Lgr5 mRNA levels compared with the control group
(P<0.05; Fig. 3D).
As the ISC counts increased in the HFD mice, the
effect of HFD on regeneration was also assessed. Using an in
vitro approach, the potential of intestinal crypts to form
multipotent, self-renewing intestinal organoids was investigated.
For this, small intestinal crypts were isolated from the control
and HFD mice, and were cultured in Matrigel to develop organoid
colonies (Fig. 3E). Compared with
the control group, the proportion of small intestinal crypts that
grew into intestinal organoids was significantly higher in the
crypts obtained from the HFD mice compared with the control mice
after 4 days of culture (P<0.05; Fig. 3F). The number of crypt-domains per
organoid at day 4 were also increased in the crypts obtained from
the HFD mice (P<0.05; Fig. 3G).
At day 6, the results also demonstrated that the proportion of
small intestinal crypts growing into intestinal organoids and the
crypt-domains per organoid were both increased in the crypts
obtained from the HFD mice (P<0.05; Fig. 3H and I). Taken together, an HFD
resulted in an increase in ISC counts, and HFD-derived crypts were
more likely to form mini-intestine organoids in a 3D culture.
HFD increases the ISC count but does
not affect crypt function in the colon
In the colon, flow cytometry revealed that the
proportion of Lgr5-high ISCs increased in the HFD mice (P<0.05;
Fig. 4A and B), whereas the
percentage of Lgr5-low progenitor cells remained the same (Fig. 4A and C). Using immunofluorescence
staining, it was demonstrated that the expression of Lgr5 in colon
epithelial tissues was upregulated in the HFD mice (P<0.05;
Fig. 4D-F). It was also observed
that the mRNA levels of Lgr5 (P<0.01) in the colonic epithelial
tissues of HFD mice were significantly upregulated compared with
those in control mice (Fig. 4G).
Furthermore, immunohistochemical staining for Ki67, an ISC marker,
indicated that the expression of Ki67 was upregulated in the HFD
mice (P<0.05; Fig. 4H and I),
while a similar change was observed in the mRNA level of Ki67
(P<0.01; Fig. 4J).
 | Figure 4.An HFD increases ISC count in the
colon. Colon crypts were isolated, and epithelial cell suspensions
were dissociated into single cells. (A) Representative flow
cytometry analysis showing Lgr5 expression in the CD326 cell
population obtained from the colon. (B) ISCs defined as
Lgr5high, and (C) progenitor cells defined as
Lgr5low in the entire colon. (D) Representative images
of immunofluorescence staining for Lgr5, seen as red fluorescence,
with DAPI used as a counterstain (scale bar, 50 µm; magnification,
×40). (E) Percentage of Lgr5+ cells among the crypt
cells, and (F) the number of Lgr5+ cells per a crypt.
(G) mRNA expression of Lgr5. (H) Representative images of
immunohistochemical staining for Ki67 (scale bar, 100 µm;
magnification, ×100), and (I) quantified Ki67+ cells as
a percentage of the crypt cells. (J) mRNA expression of Ki67 in the
colon. (K) Colon crypts were isolated from Con and HFD mice and
cultured in Matrigel to develop organoid colonies. Representative
images of colonic organoids at days 0, 4 and 6 are displayed (scale
bar, 100 µm; magnification, ×20). Organoids per crypt at (L) day 4
and (M) day 6 are shown. Data are presented as the mean ± standard
deviation. *P<0.05 and **P<0.01, vs. Con group. ISC,
intestinal stem cell; HFD, high-fat diet; Con, control. |
Intestinal organoids derived from colon crypts tend
to remain spherical and undifferentiated due to the addition of
Wnt3a to the culture medium, which is essential for maintaining
ISCs (20). In the present study,
an HFD did not affect the growth of colonic crypts into organoids
by day 4 (Fig. 4K and L) or day 6
(Fig. 4K and M). These data
support the notion that an HFD only increased the count of ISCs in
the colon but had no effect on crypt function.
Barrier function of the small
intestine is not affected by HFD
To determine whether the barrier function of the
small intestine was affected by HFD, flow cytometry analysis was
used to analyze the proportion and number of IECs, particularly
goblet and Paneth cells. The results revealed that there were no
marked changes in the proportion of goblet cells in the HFD mice
(Fig. 5A and B). No significant
difference in the number of goblet cells in single crypts,
determined by PAS staining, were observed in the HFD group compared
with the control group (Fig.
5C-E). Similarly, the proportion of Paneth cells was not
affected by the HFD (Fig. 5F and
G), and the number of Paneth cells in the small intestinal
crypt derived from organoids did not differ significantly between
the HFD and control mice (Fig. 5H and
I).
 | Figure 5.Barrier function of the small
intestine was not affected by HFD. Goblet and Paneth cell numbers,
and cell apoptosis were analyzed to determine the barrier function
of the small intestine. (A) Representative flow cytometry analysis
of Muc2 expression in CD326 cell populations. (B) Percentage of
goblet cells in the small intestine. (C) Ileal sections were
stained for goblet cells using PAS (scale bar, 500 µm;
magnification, ×20). (D) Percentage of area stained as PAS-positive
among the total villi-crypts sections. (E) Quantification of the
number of goblet cells per villus-crypt unit. (F) Representative
flow cytometry analysis showing CD24 expression in CD326 cell
populations. (G) Percentage of Paneth cells in small intestinal
crypts. (H) Crypt-derived organoids of Paneth cells, including
lysozyme-positive cells, assessed using immunofluorescence (blue,
DAPI; red, lysozyme; scale bar, 200 µm; magnification, ×40). (I)
Quantification of Paneth cells in small intestinal crypt-derived
organoids. (J) mRNA expression levels of Muc2 in the small
intestine of the Con and HFD mice. HFD, high-fat diet; Con,
control; PAS, Periodic Acid-Schiff; Muc2, mucin 2. |
It is known that the secretion of highly
glycosylated mucins into the intestinal lumen by goblet cells
creates the first line of defense against microbial encroachment
(21). In the present study, the
mRNA expression levels of Muc2 in the small intestine tissues did
not significantly differ in the HFD group compared with the control
mice, as determined by RT-qPCR (Fig.
5J).
The rate of apoptosis of IECs was then determined by
conducting a TUNEL assay. The results demonstrated no significant
difference in the apoptosis rate between the HFD and control mice
(Fig. 6A-C). Furthermore,
immunohistochemical staining for P21 (Fig. 6D-F) and P53 (Fig. 6G-I), which are apoptosis-marker
genes, was not significantly altered by the HFD. Additionally,
consistent results were obtained for the mRNA expression levels of
P21 and P53 (Fig. 6J and K). These
results suggested that the small intestine barrier between the host
and the internal environment was not affected by the HFD.
HFD compromises the intestinal barrier
function
To determine the effect of an HFD on the barrier
function of the colon, flow cytometry analysis was also used to
analyze the proportion and number of goblet cells in the colon. The
results revealed that the proportion of goblet cells amongst
colonic epithelial cells was significantly lower in the HFD group
compared with that in the control group (P<0.01; Fig. 7A and B). Using PAS staining, it was
demonstrated that the number of goblet cells was also decreased
(P<0.05; Fig. 7C-E). Muc2 is
the most abundant mucin produced by goblet cells and serves an
essential role in the organization of the intestinal mucous layers
at the epithelial surface of the colon (22). Therefore, the mRNA expression
levels of Muc2 were assessed, and it was observed that Muc2 levels
were significantly decreased in the HFD mice compared with the
control mice (P<0.001; Fig.
7F). These data suggest that an HFD results in damage to the
IEC secretory function.
 | Figure 7.An HFD compromises the barrier
function of the colon. Goblet cell number, tight junctions and cell
apoptosis were analyzed to determine the barrier function of the
colon. (A) Representative flow cytometry analysis, showing Muc2
expression among the CD326 cell populations. (B) Proportion of
goblet cells. (C) Histochemical PAS staining of goblet cells (scale
bar, 100 µm; magnification, ×20), (D) percentage of area stained as
PAS-positive among the colonic crypts, and (E) quantification of
the average number of goblet cells per crypt. (F) mRNA expression
of Muc2 in the colon. (G) Representative images of
immunofluorescence staining for ZO-1 seen as red fluorescence, with
DAPI used as the counterstain (scale bar, 50 µm; magnification,
×40), and (H) quantification of red fluorescence. (I)
Representative images of immunofluorescence staining for Claudin-2
seen as red fluorescence, with DAPI used as the counterstain (scale
bar, 100 µm; magnification, ×40), and (J) quantification of red
fluorescence. (K) mRNA expression of Claudin-2 in the colon.
*P<0.05, **P<0.01 and ***P<0.001, vs. Con group. HFD,
high-fat diet; Con, control; PAS, Periodic Acid-Schiff; Muc2, mucin
2. |
Below the mucous layers, IECs form a continuous
physical barrier (21,23). Tight junctions connect adjacent
IECs and are associated with intestinal permeability (24). Disruption of barrier function is
characterized by defects in epithelial tight-junction-associated
proteins, such as ZO-1 and Claudin-2 (25). In the current study, PAS staining
indicated that the expression of ZO-1 was downregulated in the HFD
mice compared with the controls (P<0.05; Fig. 7G and H). However, the expression of
Claudin-2 was significantly upregulated in the HFD mice (P<0.05;
Fig. 7I-K).
The rate of apoptosis in colonic epithelial cells
may also influence the function of the colonic barrier. Therefore,
the apoptotic rate of these cells was measured. The results
revealed that the rate of apoptosis was increased in the HFD mice
(P<0.01; Fig. 8A and B). In
addition, immunohistochemical staining for P21 (Fig. 8C and D) and P53 (Fig. 8E and F) demonstrated increasing
expression of these genes in the HFD mice (P<0.05). The mRNA
expression levels of P21 (P<0.05) and P53 (P=0.0518) were also
increased in the HFD mice, compared with the control mice (Fig. 8G and H). These findings suggest
that an HFD results in increased apoptosis of IECs of the
colon.
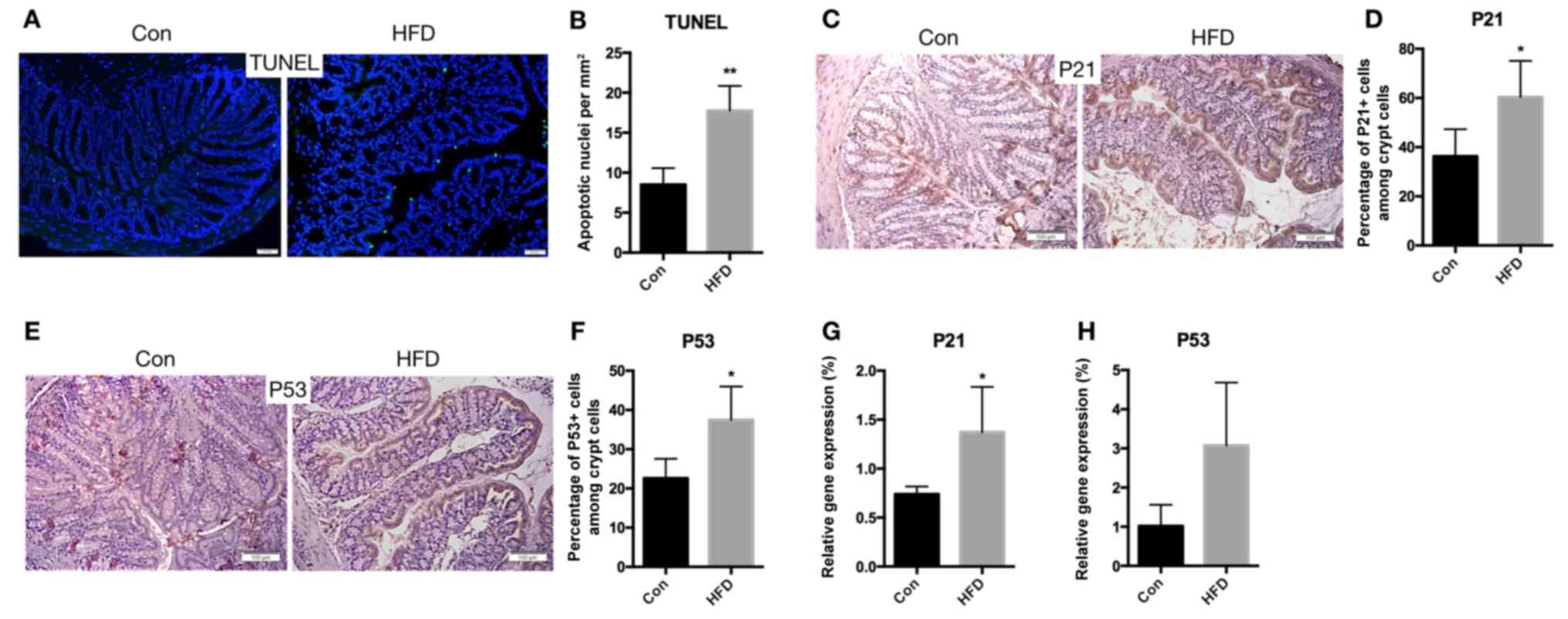 | Figure 8.Increased apoptosis in the colon of
HFD-fed mice. (A) Colon sections were stained for apoptotic cells
using TUNEL assay (scale bar, 50 µm; magnification, ×40), and (B)
the quantified number of apoptotic cells is shown. (C)
Immunohistochemical staining for P21 expression in the colon (scale
bars, 100 µm; magnification, ×40), and (D) quantified percentage of
P21-positive crypt cells. (E) Representative images of
immunohistochemical staining for P53 (scale bars, 100 µm;
magnification, ×40), and (F) quantified percentage of P53-positive
crypt cells. (G) P21 and (H) P53 mRNA expression levels in the
colon. Data are presented as the mean ± standard deviation.
*P<0.05 and **P<0.01, vs. Con group. HFD, high-fat diet; Con,
control. |
Discussion
As the intestine consumes a considerable amount of
energy, modulating the intestinal volume and cellular architecture
is an important adaptation to fluctuations in nutrient availability
(26,27). Several studies have reported that
excess or reduced calorie consumption have profound implications on
the physiology of the intestine. For instance, reduced calorie
intake resulted in reduced overall mass of the small intestine in
murine models, while increased calorie intake reversed these
changes (28). Furthermore,
overfeeding or consumption of an HFD stimulates proliferation of
the intestinal epithelium (29).
However, studies on the effects of an HFD on the morphology of the
intestine, particularly the small intestine, are contradictory.
Beyaz et al (18) reported
that the small intestines of HFD-fed mice were shorter in length,
and that an HFD also resulted in a mild reduction in villi length
and an increase in crypt depth. However, other studies observed
that the duodenal and jejunal crypt-villus height increased
significantly in HFD-fed or db/db mice (29,30).
These inconsistent results may be due to the differences in the
animal models, such as the high-fat feeding time and age of the
mice. In the present study, the villus length of small intestine,
as well as the total length and crypt depth of the colon, decreased
in the HFD-fed group. These morphological changes suggest an effect
of HFD on the intestinal function.
An increasing number of epidemiological studies have
indicated that obesity is associated with an elevated risk of
several different types of cancer, particularly of gastrointestinal
tract cancer (31–33). One of the primary drivers of the
contribution of diet to cancer incidence is through alterations to
the biology of ISCs. A previous study demonstrated that different
dietary regimens and calorie restriction increased the stem cell
numbers and regulated ISC function (34). By contrast, in the present study,
an HFD increased the ISC counts, induced self-renewal and increased
features associated with stemness (organoid-initiating capacity).
Similar results were obtained in the study by Beyaz et al
(18), in that an HFD enhanced the
stemness and tumorigenicity of intestinal progenitor cells,
suggesting that an HFD enhances regeneration. A recent study
revealed that ISCs in females have greater proliferative ability as
compared with those in males (35). The results of the present study
indicated that female mice fed an HFD exhibited increased ISC
counts, while crypts obtained from mice fed an HFD were more likely
to form mini-intestine organoids in a 3D culture, which may
partially account for the increase in intestinal tumors in obese
individuals.
Tight junctions are structures in-between IECs and a
key element of a tight leak-proof barrier (36,37).
The expression of junctional proteins in the intestine is tightly
regulated, and the complex pattern is associated with the specific
functions of a particular intestinal region and localization
(38). The Claudin family of
proteins is more complex than other tight junction proteins, and a
total of 24 distinct Claudin family gene members have been
identified in humans to date (39). Claudins exhibit distinct tissue-,
cell- and developmental stage-specific expression patterns
(40,41). Claudin-2 is restricted to
undifferentiated crypt cells and is undetectable in the normal
colon, but is upregulated along an inflamed crypt epithelium
(42,43). This may explain the increase in
Claudin-2 expression that was observed in mice fed an HFD in the
present study, as compared with the control mice.
In conclusion, the data of the present study
suggested that an HFD results in changes to the regulation of the
stemness of ISCs and to the epithelial barrier function of the
colon. Thus, dietary intervention may be an effective strategy for
reducing the risk of colon cancer and other systemic diseases, such
as cardiovascular diseases and metabolic syndrome. Further research
should focus on understanding the mechanisms through which
nutrients impact other tissues via the intestine.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant. no. 91649122)
and the National Natural Science Youth Foundation of China (grant.
no. 81600688).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
GD and JY conceived and designed the study. FD, YX,
WD and YL performed the experiments. FX and YS analyzed the data.
JY wrote the manuscript.
Ethics approval and consent to
participate
The experimental protocols of the present study were
approved by the Animal Care and Use Committee of Nanjing Medical
University (Nanjing, China), and conducted in accordance with the
guidelines of this committee.
Patient consent for publication
Not applicable.
Competing of interests
The authors declare that they have no competing
interests.
References
|
1
|
Nagao M, Asai A, Sugihara H and Oikawa S:
Fat intake and the development of type 2 diabetes. Endocr J.
62:561–572. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schulz MD, Atay C, Heringer J, Romrig FK,
Schwitalla S, Aydin B, Ziegler PK, Varga J, Reindl W, Pommerenke C,
et al: High-fat-diet-mediated dysbiosis promotes intestinal
carcinogenesis independently of obesity. Nature. 514:508–512. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bleau C, Karelis AD, St-Pierre DH and
Lamontagne L: Crosstalk between intestinal microbiota, adipose
tissue and skeletal muscle as an early event in systemic low-grade
inflammation and the development of obesity and diabetes. Diabetes
Metab Res Rev. 31:545–561. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
David LA, Maurice CF, Carmody RN,
Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y,
Fischbach MA, et al: Diet rapidly and reproducibly alters the human
gut microbiome. Nature. 505:559–563. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu GD, Chen J, Hoffmann C, Bittinger K,
Chen YY, Keilbaugh SA, Bewtra M, Knights D, Walters WA, Knight R,
et al: Linking long-term dietary patterns with gut microbial
enterotypes. Science. 334:105–108. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Backhed F, Ley RE, Sonnenburg JL, Peterson
DA and Gordon JI: Host-bacterial mutualism in the human intestine.
Science. 307:1915–1920. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qin N, Yang F, Li A, Prifti E, Chen Y,
Shao L, Guo J, Le Chatelier E, Yao J, Wu L, et al: Alterations of
the human gut microbiome in liver cirrhosis. Nature. 513:59–64.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Carmody RN, Gerber GK, Luevano JM Jr,
Gatti DM, Somes L, Svenson KL and Turnbaugh PJ: Diet dominates host
genotype in shaping the murine gut microbiota. Cell Host Microbe.
17:72–84. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kurashima Y, Goto Y and Kiyono H: Mucosal
innate immune cells regulate both gut homeostasis and intestinal
inflammation. Eur J Immunol. 43:3108–3115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bjerregaard LG, Jensen BW, Angquist L,
Osler M, Sorensen TIA and Baker JL: Change in overweight from
childhood to early adulthood and risk of type 2 diabetes. N Engl J
Med. 378:1302–1312. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feldman AT and Wolfe D: Tissue processing
and hematoxylin and eosin staining. Methods Mol Biol. 1180:31–43.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dame MK and Varani J: Human colon tissue
in organ culture. Replacing animal models. Davies J: John Wiley
& Sons, Inc.; Hoboken, NJ: pp. 69–80. 2012, View Article : Google Scholar
|
|
13
|
Sato T and Clevers H: Primary mouse small
intestinal epithelial cell cultures. Methods Mol Biol. 945:319–328.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sato T, Vries RG, Snippert HJ, van de
Wetering M, Barker N, Stange DE, van Es JH, Abo A, Kujala P, Peters
PJ and Clevers H: Single Lgr5 stem cells build crypt-villus
structures in vitro without a mesenchymal niche. Nature.
459:262–265. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Date S and Sato T: Mini-gut organoids:
Reconstitution of the stem cell niche. Annu Rev Cell Dev Biol.
31:269–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Koo BK, Sasselli V and Clevers H:
Retroviral gene expression control in primary organoid cultures.
Curr Protoc Stem Cell Biol. 27:Unit 5A.6. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sato T, van Es JH, Snippert HJ, Stange DE,
Vries RG, van den Born M, Barker N, Shroyer NF, van de Wetering M
and Clevers H: Paneth cells constitute the niche for Lgr5 stem
cells in intestinal crypts. Nature. 469:415–418. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Beyaz S, Mana MD, Roper J, Kedrin D,
Saadatpour A, Hong SJ, Bauer-Rowe KE, Xifaras ME, Akkad A, Arias E,
et al: High-fat diet enhances stemness and tumorigenicity of
intestinal progenitors. Nature. 531:53–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Merker SR, Weitz J and Stange DE:
Gastrointestinal organoids: How they gut it out. Dev Biol.
420:239–250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peterson LW and Artis D: Intestinal
epithelial cells: Regulators of barrier function and immune
homeostasis. Nat Rev Immunol. 14:141–153. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Johansson ME, Phillipson M, Petersson J,
Velcich A, Holm L and Hansson GC: The inner of the two Muc2
mucin-dependent mucus layers in colon is devoid of bacteria. Proc
Natl Acad Sci USA. 105:15064–15069. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vancamelbeke M and Vermeire S: The
intestinal barrier: A fundamental role in health and disease.
Expert Rev Gastroenterol Hepatol. 11:821–834. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ahmad R, Sorrell MF, Batra SK, Dhawan P
and Singh AB: Gut permeability and mucosal inflammation: Bad, good
or context dependent. Mucosal Immunol. 10:307–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Assimakopoulos SF, Papageorgiou I and
Charonis A: Enterocytes' tight junctions: From molecules to
diseases. World J Gastrointest Pathophysiol. 2:123–137. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Matheson PJ, Wilson MA and Garrison RN:
Regulation of intestinal blood flow. J Surg Res. 93:182–196. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Mihaylova MM, Sabatini DM and Yilmaz OH:
Dietary and metabolic control of stem cell function in physiology
and cancer. Cell Stem Cell. 14:292–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dunel-Erb S, Chevalier C, Laurent P, Bach
A, Decrock F and Le Maho Y: Restoration of the jejunal mucosa in
rats refed after prolonged fasting. Comp Biochem Physiol A Mol
Integr Physiol. 129:933–947. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mao J, Hu X, Xiao Y, Yang C, Ding Y, Hou
N, Wang J, Cheng H and Zhang X: Overnutrition stimulates intestinal
epithelium proliferation through β-catenin signaling in obese mice.
Diabetes. 62:3736–3746. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Baldassano S, Amato A, Cappello F, Rappa F
and Mule F: Glucagon-like peptide-2 and mouse intestinal adaptation
to a high-fat diet. J Endocrinol. 217:11–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Basen-Engquist K and Chang M: Obesity and
cancer risk: Recent review and evidence. Curr Oncol Rep. 13:71–76.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Calle EE and Kaaks R: Overweight, obesity
and cancer: Epidemiological evidence and proposed mechanisms. Nat
Rev Cancer. 4:579–591. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Berger NA: Obesity and cancer
pathogenesis. Ann N Y Acad Sci. 1311:57–76. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yilmaz OH, Katajisto P, Lamming DW,
Gültekin Y, Bauer-Rowe KE, Sengupta S, Birsoy K, Dursun A, Yilmaz
VO, Selig M, et al: mTORC1 in the Paneth cell niche couples
intestinal stem-cell function to calorie intake. Nature.
486:490–495. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou W, Davis EA, Li K, Nowak RA and
Dailey MJ: Sex differences influence intestinal epithelial stem
cell proliferation independent of obesity. Physiol Rep.
6:e137462018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suzuki T: Regulation of intestinal
epithelial permeability by tight junctions. Cell Mol Life Sci.
70:631–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Menard S, Cerf-Bensussan N and Heyman M:
Multiple facets of intestinal permeability and epithelial handling
of dietary antigens. Mucosal Immunol. 3:247–259. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Laukoetter MG, Bruewer M and Nusrat A:
Regulation of the intestinal epithelial barrier by the apical
junctional complex. Curr Opin Gastroenterol. 22:85–89. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tsukita S, Furuse M and Itoh M:
Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol.
2:285–293. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ebnet K, Suzuki A, Ohno S and Vestweber D:
Junctional adhesion molecules (JAMs): More molecules with dual
functions? J Cell Sci. 117:19–29. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bazzoni G: The JAM family of junctional
adhesion molecules. Curr Opin Cell Biol. 15:525–530. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Heller F, Florian P, Bojarski C, Richter
J, Christ M, Hillenbrand B, Mankertz J, Gitter AH, Bürgel N, Fromm
M, et al: Interleukin-13 is the key effector Th2 cytokine in
ulcerative colitis that affects epithelial tight junctions,
apoptosis, and cell restitution. Gastroenterology. 129:550–564.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu Z, Ding L, Lu Q and Chen YH: Claudins
in intestines: Distribution and functional significance in health
and diseases. Tissue Barriers. 1:e249782013. View Article : Google Scholar : PubMed/NCBI
|
















