Introduction
Glioblastoma (GBM) represents 15% of all brain
tumors (1). In the majority of
cases, the length of survival following diagnosis is 12–15 months,
with fewer than 3–5% of patients surviving for >5 years
(2). In general, surgery after
chemotherapy and radiation therapy are commonly used for GBM
treatment (3). However, the cancer
usually recurs, as there is no clear way to prevent the disease
(4).
Improved understanding of the mechanisms underlying
GBM at molecular and structural levels is valuable for clinical
treatment (5). Bioinformatics may
be effectively used to analyze GBM microarray data in order to
provide a theoretical reference for further exploration of
tumorigenic mechanisms and aid in searching for potential target
genes (6). Based on bioinformatic
studies, certain differentially expressed genes (DEGs), including
p21 (RAC1)-activated kinase 7 (PAK7), were identified as potential
therapeutic targets for glioma (7). As the longest member of the PAK
family, PAK7 (renamed from PAK5) contains a total of eight
protein-coding exons (8). PAK7,
initially cloned as a brain-specific kinase, was hypothesized to be
involved in neurite growth in neuronal cells (9,10).
The downregulation of PAK7 expression was shown to inhibit glioma
cell migration and invasion through competing endogenous RNA
(ceRNA) networks such as the PAK7-early growth response 1-matrix
metalloproteinase 2 signaling pathway (11). The levels of PAK7 are regulated by
microRNAs (miRNAs/miRs) such as miR-129 and are associated with
long non-coding RNAs (lncRNAs) in glioma cell progression (12). However, the potential molecular
mechanisms underlying PAK7 activity in GBM remain unclear.
In the present study, gene expression data from GBM
were downloaded and reanalyzed using bioinformatic tools. The DEGs,
including differentially expressed lncRNAs (DElncRNAs) and
differentially expressed mRNAs (DEMs) between GBM samples and
normal samples were investigated and subjected to functional and
pathway enrichment analyses. PAK7-associated miRNAs were
investigated and a PAK7-associated ceRNA network was constructed.
Furthermore, The Cancer Genome Atlas (TCGA) survival analysis was
performed to confirm the effect of PAK7 expression on GBM. The
study aimed to reveal the potential molecular mechanisms underling
PAK7 expression in GBM and identify candidate prognostic biomarkers
for GBM.
Materials and methods
Microarray data
The RNA-seq profile data and clinical data of
TCGA-GBM were downloaded from the University of California Santa
Cruz (UCSC) database (http://xena.ucsc.edu) based on the platform of
Illumina HiSeq 2000 RNA Sequencing (Illumina, Inc.). A total of 144
tumor samples (GBM group) and five normal brain samples (control
group) were enrolled in the present study.
Exploration of lncRNAs and mRNAs by
re-annotation
An exon was defined as lncRNA or mRNA if its
starting and ending positions (including the positive and negative
chain) in the current RNAseq data were in accordance with the
chromosome location annotation of lncRNA or mRNA (hg19, V25) in
GENCODE database (https://www.gencodegenes.org/) (13), respectively.
Investigation of DEGs
Genes with low expression (expression value was 0 in
>50% samples) were removed. Linear regression and empirical
Bayesian methods (14,15) in Limma package of R (version:
3.40.2) (16) were used to explore
DElncRNAs and DEMs. Multiple correction on the P-value of each DEG
was performed using the Benjamini-Hochberg method (17) to obtain the adjusted P value. An
adjusted P<0.05 and log|fold change| >1.5 were selected as
the thresholds for DEG screening. A volcano plot was constructed
based on the different expression values of DEMs and DElncRNAs.
Non-metric multidimensional scaling (NMDS) (18) was performed on DElncRNAs and DEMs
using the vegan (version 2.5–2; http://cran.r-project.org/web/packages/vegan/) package
(19) in R, followed by the
construction of an NMDS plot. Analysis of similarities (20) was performed for sample similarity
analysis. The distance between two points was defined as
Bray-Curtis, and displacement tests were carried out. The threshold
was set as P<0.05.
Gene set enrichment analysis (GSEA)
for PAK7
Pearson's correlation coefficients for all mRNA-PAK7
pairs were calculated and pathway analysis (21) was performed on these mRNA-PAK7
pairs (according to the descending order of the correlation
coefficient) via GSEA in the clusterProfiler package of R (version:
3.12.0) (22). A value of
P<0.05 was considered as the threshold for current analysis.
miRNA prediction and co-expression
investigation
Pearson's correlation coefficients for all
DElncRNA-PAK7 and DEM-PAK7 relations were calculated and
correlation tests were performed. DElncRNAs and DEMs with |r|
>0.6 and P<0.05 were considered as key genes that were
significantly associated with PAK7. The correlation between lncRNAs
and mRNAs was calculated with the methods mentioned above. miRNA
predictions for DEMs that positively correlated with mRNAs
(including PAK7) was performed using miRWalk2.0 software (23,24)
and combined with the prediction results from six databases,
including miRWalk, miRanda (http://www.microrna.org/) (25), miRDB (http://www.mirdb.org/) (26), miRMap (http://www.mirmap.ezlab.org/) (27), RNA22 (https://www.rna-seqblog.com/rna22) (28) and TargetScan 7.1 (http://www.targetscan.org/vert_71/) (29). PAK7-DEMs-miRNAs that appeared in at
least five databases were selected for further investigation.
Based on the miRNA-lncRNA relations in both starBase
(30) (version 2.0; http://starbase.sysu.edu.cn/) and InCeDB (31) (http://gyanxet-beta.com/lncedb/) databases, the
DElncRNAs that were positively related with PAK7 were predicted,
and PAK7-DElncRNA-miRNA associations were investigated. The RNAs
with same miRNA-binding site were defined as ceRNAs. Based on the
PAK7-DEM-miRNA and PAK7-DElncRNA-miRNA interactions, the
lncRNA-mRNA relationships regulated by the same miRNA were
investigated. miR2Disease database (http://watson.compbio.iupui.edu:8080/miR2Disease/index.jsp)
was used for miRNA investigation (search keyword: ‘Glioblastoma
multiforme’) followed by lncRNA-mRNA association analysis. The
miRNA-lncRNA-mRNA regulation network was constructed using
lncRNA-mRNA and DEM-DElncRNA relationships obtained. Finally,
lncRNAs and mRNAs regulated by the same miRNA (PAK7 was also
regulated by this miRNA) were screened as key ceRNAs for PAK7.
Survival analysis
To reveal the prognostic value of the hub genes in
patients with GBM, survival analysis, including overall survival
(OS) and OS status, was performed. The samples in the data were
divided into a high expression group (up group) and low expression
group (down group) according to the mean and median values of all
lncRNAs/mRNAs in the ceRNA network. The survival package (version
2.41–3; http://cran.r-project.org/web/views/Survival.html) in
R was used for the log-rank test (32) in two groups. P<0.05 was
considered statistically significant. Associations between
lncRNAs/mRNAs and GBM prognosis were analyzed. The Kaplan-Meier
(KM) (33) survival curve was
constructed based on these selected lncRNAs/mRNAs.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
A total of 8 tumor specimens were collected from
patients with primary GBM (33–79 years old; 3 female and 5 male)
who underwent neurosurgery at Fourth Affiliated Hospital of Harbin
Medical University from February 2017 to July 2018, and 8 normal
brain specimen samples (44–83 years; 4 female and 4 male) were
obtained from patients who underwent cerebral contusion at Fourth
Affiliated Hospital of Harbin Medical University from April 2017 to
May 2018. Patients with a family history of cancer, and patients
with chronic heart, lung, liver and kidney diseases were excluded.
The study was approved by the Human Ethics Committee of Fourth
Affiliated Hospital of Harbin Medical University and informed
consent was obtained from all patients. RT-qPCR analysis was
performed to detect the expression levels of several DElncRNAs
(LINC00599) and DEMs [PAK7, PNMA family member 5 (PNMA5), and
somatostatin receptor 1 (SSTR1)], as well as miR-185-5p. Briefly,
total RNA was extracted from eight GBM samples and eight normal
brain tissue samples using TRIzol® reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). cDNA was synthesized using
PrimeScript™ RT Master Mix (Perfect Real Time) (Takara
Biotechnology, Co., Ltd.). The amplification was performed SYBR
Premix Ex Taq kit (cat. no. DRR041A; Takara Bio, Inc.) under the
following conditions on an ABI ViiA 7 Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.): 50°C for 3 min, 95°C
for 3 min, then 25 cycles of 95°C for 10 sec and 60°C for 30 sec,
followed by dissociation curve analysis (60°C-95°C; 0.5°C
increments for 10 sec). The primers used are shown in Table I. Relative expression levels were
normalized to GAPDH and calculated with the 2−ΔΔCq
method (34), and each sample was
assayed in triplicate.
 | Table I.Primers used for RT-qPCR. |
Table I.
Primers used for RT-qPCR.
| Primer | Primer sequence
(5′→3′) |
|---|
| PAK7-hF |
GTAGTAGTTCCCCAGCGTGC |
| PAK7-hR |
ACATGGTCTCAGAGGTCCAG |
| PNMA5-hF |
TGCCCAGTCACATACCAGGAA |
| PNMA5-hR |
CATACTTCGGCCCTCATCTTTC |
| SSTR1-hF |
TGGCTATCCATTCCATTTGACC |
| SSTR1-hR |
AGGACTGCATTGCTTGTCAGG |
| LINC00599-hF |
CTAAACCCCTTGCCCCACAA |
| LINC00599-hR |
AGGTTTTACAGGAGGGCAGC |
|
hsa-miR-185-5p-R |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAGGA |
|
hsa-miR-185-5p-F |
GGCTGGAGAGAAAGGCAGT |
| GAPDH-hF |
TGACAACTTTGGTATCGTGGAAGG |
| GAPDH-hR |
AGGCAGGGATGATGTTCTGGAGAG |
| U6-hF |
CTCGCTTCGGCAGCACA |
| U6-hR |
AACGCTTCACGAATTTGCGT |
| human-U6-RT |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAATATG |
| Downstream
universal primer |
GTGCAGGGTCCGAGGT |
Data are presented as the mean ± SD, and statistical
analysis was performed using GraphPad Prism 5.0 (GraphPad Software,
Inc.). The difference between two groups was analyzed with a
t-test, and a P<0.05 was considered to indicate a statistically
significant difference.
Results
Investigation of DElncRNAs and
DEMs
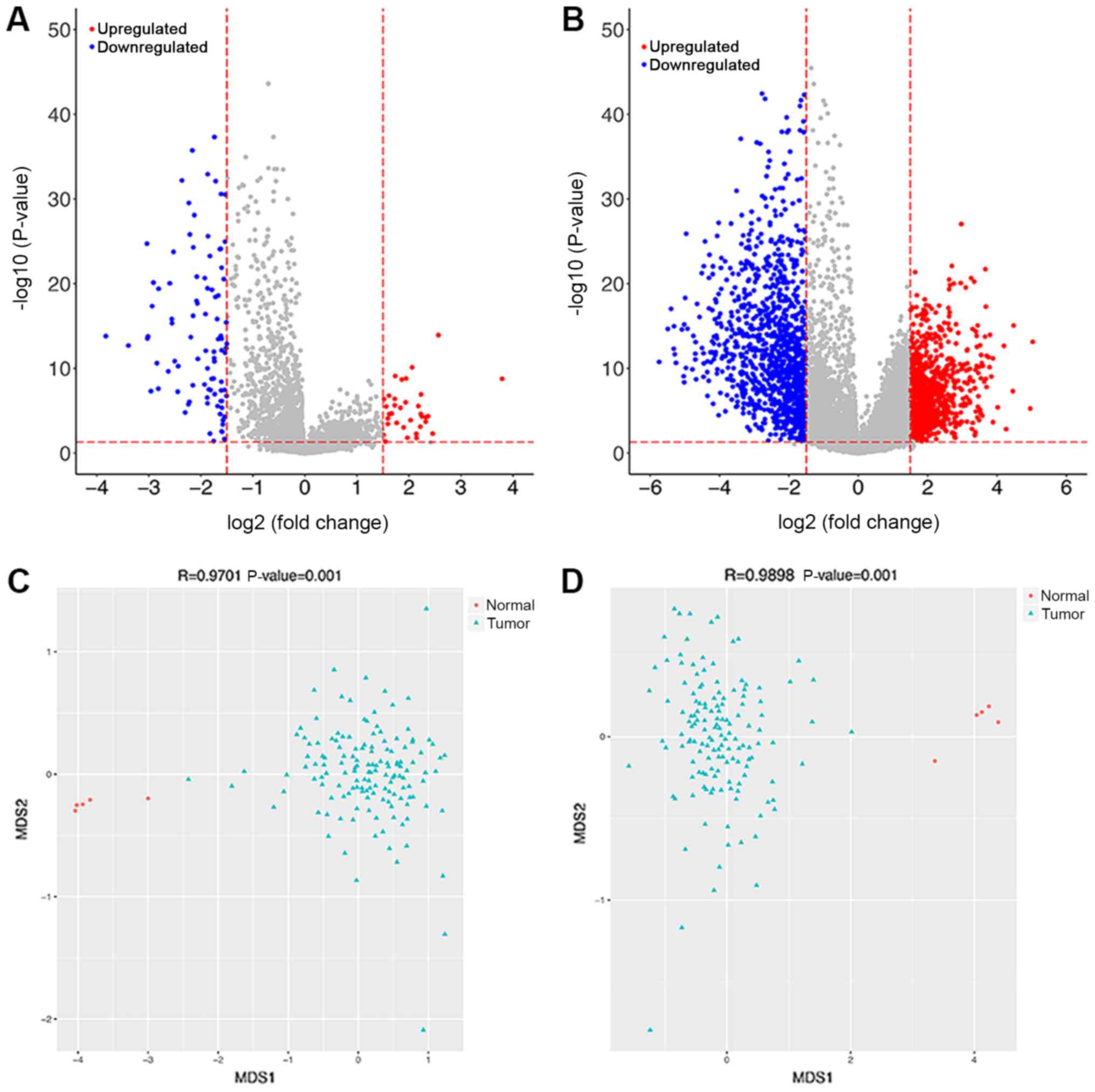
A total of 34 upregulated and 92 downregulated
DElncRNAs were revealed between two groups. A total of 920
upregulated DEMs and 1,142 downregulated DEMs were observed. The
volcanic diagrams for DElncRNAs and DEMs are shown in Fig. 1A and B. The expression values of
DElncRNAs and DEMs were used to construct NMDS plots for these
genes (Fig. 1C and D). The results
showed that the green points (samples in GBM group) and red points
(samples in control group) were distinctly separated, indicative of
clear separation of samples based on the DEMs and DElncRNAs.
Pathway enrichment analysis
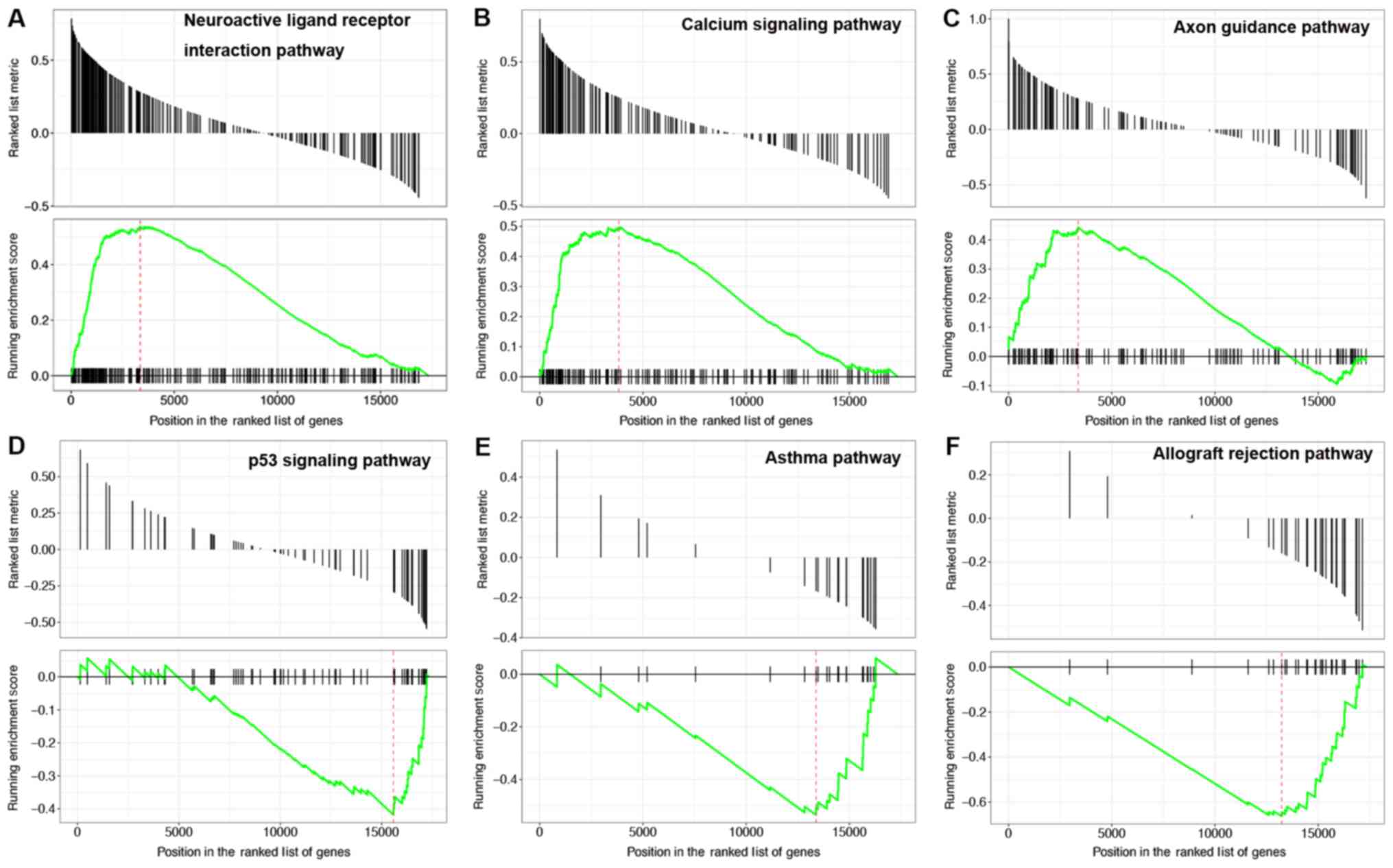
GSEA pathway analysis was performed on the mRNAs in
mRNA-PAK7 pairs. The results showed that the upregulated mRNAs were
mainly enriched in 14 pathways, including neuroactive ligand
receptor interactions, calcium signaling pathways and axon guidance
(Fig. 2A-C). The downregulated
mRNAs were mainly enriched in 35 pathways, including p53 signaling
pathways, asthma and allograft rejection (Fig. 2D-F). According to the P-value, the
detailed information for the top six upregulated and downregulated
pathways is shown in Table II.
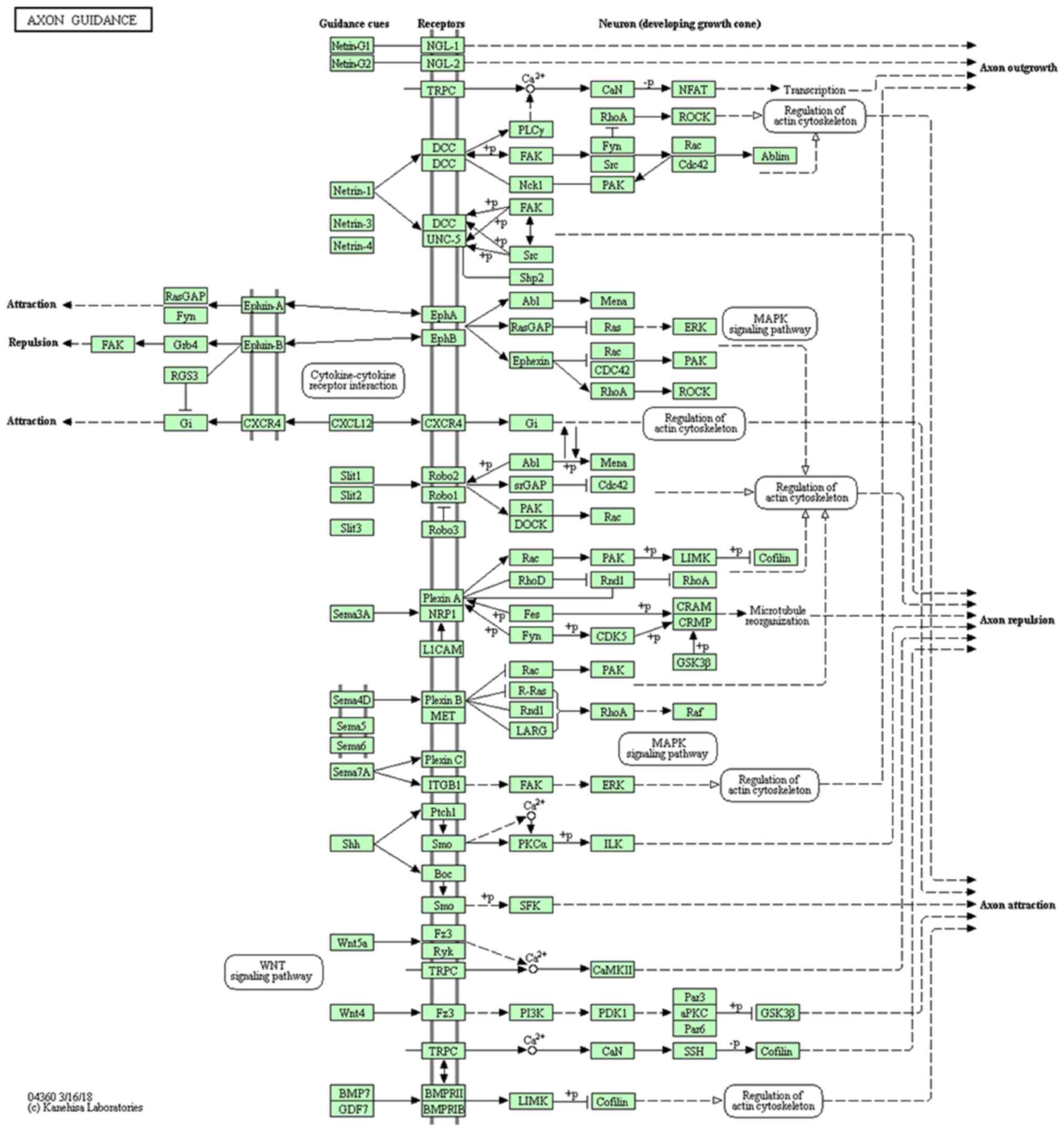
The results of the pathway analysis indicated that PKA7
participated in axon guidance pathway (Fig. 3).
| Table II.Top 6 pathways enriched by up- and
downregulated mRNAs. |
Table II.
Top 6 pathways enriched by up- and
downregulated mRNAs.
| A, Upregulated
mRNAs |
|
| Pathway ID | P-value
(adjusted) | Enriched genes |
|---|
|
KEGG_NEUROACTIVE_LIGAND_RECEPTOR_INTERACTION |
3.32×10−2 | GABRA3, CHRNA4,
HRH3, CHRM1, GABRB3… |
|
KEGG_CALCIUM_SIGNALING_PATHWAY |
3.32×10−2 | CACNA1E, CHRM1,
HTR6, PLCB1, GNAL… |
|
KEGG_AXON_GUIDANCE |
3.32×10−2 | PAK1, PAK3, PAK6,
PAK7, L1CAM… |
|
| B, Downregulated
mRNAs |
|
| Pathway
ID | P-value | Enriched
genes |
|
|
KEGG_NEUROACTIVE_LIGAND_RECEPTOR_INTERACTION |
3.32×10−2 | GABRA3, CHRNA4,
HRH3, CHRM1, GABRB3… |
|
KEGG_CALCIUM_SIGNALING_PATHWAY |
3.32×10−2 | CACNA1E, CHRM1,
HTR6, PLCB1, GNAL… |
|
KEGG_AXON_GUIDANCE |
3.32×10−2 | PAK1, PAK3, PAK6,
PAK7, L1CAM… |
Co-expression network analysis
A total of 312 mRNAs and 22 lncRNAs showed positive
correlation with PAK7. Following miRNA prediction, a total of
25,440 miRNA-lncRNA-mRNA regulatory interactions were reported from
the screening of lncRNA-mRNA pairs regulated by the same miRNA. The
selection of disease-related miRNAs revealed a total of 185
miRNA-lncRNA-mRNA relationships. Combined positive co-expression
lncRNA-mRNA relationships revealed a total of 159 miRNA-lncRNA-mRNA
regulatory associations. The ceRNA network was constructed based on
network connectivity (Fig. 4). The
results revealed 103 nodes (including 6 miRNAs, 92 mRNAs and 5
lncRNAs) and 281 interactions (including 115 miRNA-mRNA
interactions, 7 miRNA-lncRNA interactions and 159 lncRNA-mRNA
interactions), including miR-185-5p-LINC00599-PAK7 in the current
ceRNA network.
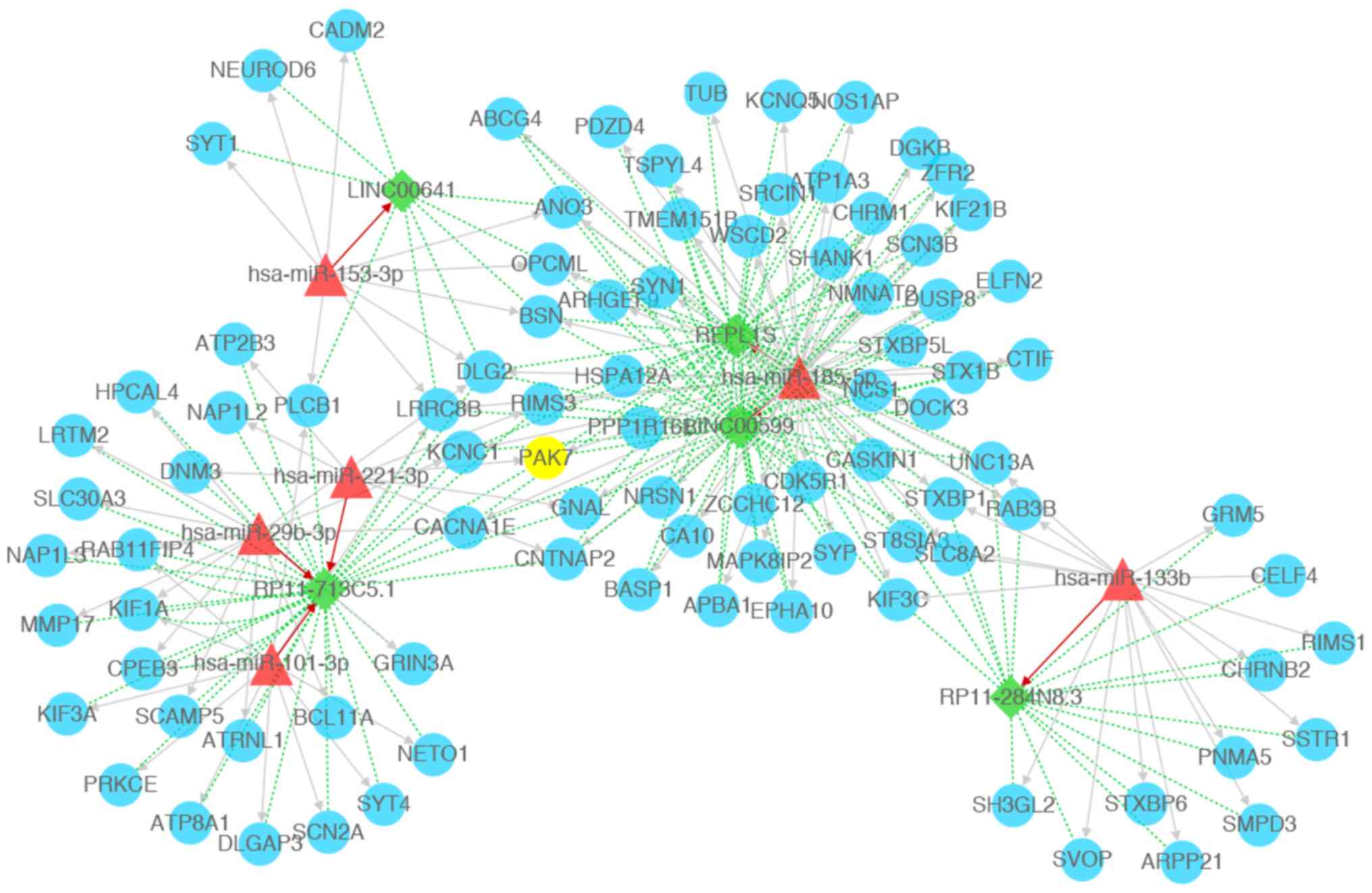 | Figure 4.Competing endogenous RNA network
constructed with miRNAs, lncRNAs, and mRNAs. Green diamonds
represent lncRNAs, blue circles represents mRNAs (PAK7 is
highlighted in yellow), red triangles represent miRNAs, red arrow
represents miRNA-lncRNA regulatory relations, gray arrows represent
miRNA-mRNA regulatory relations, green dotted lines represent
mRNA-lncRNA co-expression relations. PAK7, p21 (RAC1)-activated
kinase 7; miRNA/miR, microRNA; lncRNA, long non-coding RNA. |
The mRNAs and lncRNAs regulated by the same miRNAs
as PAK7 were investigated, followed by the construction of a core
ceRNA network. As shown in Fig. 5,
a total of 63 nodes (2 miRNAs, 58 mRNAs and 3 lncRNAs) and 171
interactions (62 miRNA-mRNA relations, 3 miRNA-lncRNA relations and
106 lncRNA-mRNA relations) were observed in the current core ceRNA
network. These mRNAs and lncRNAs were regulated by hsa-mir-185-5p
or hsa-mir-221-3p together with PAK7.
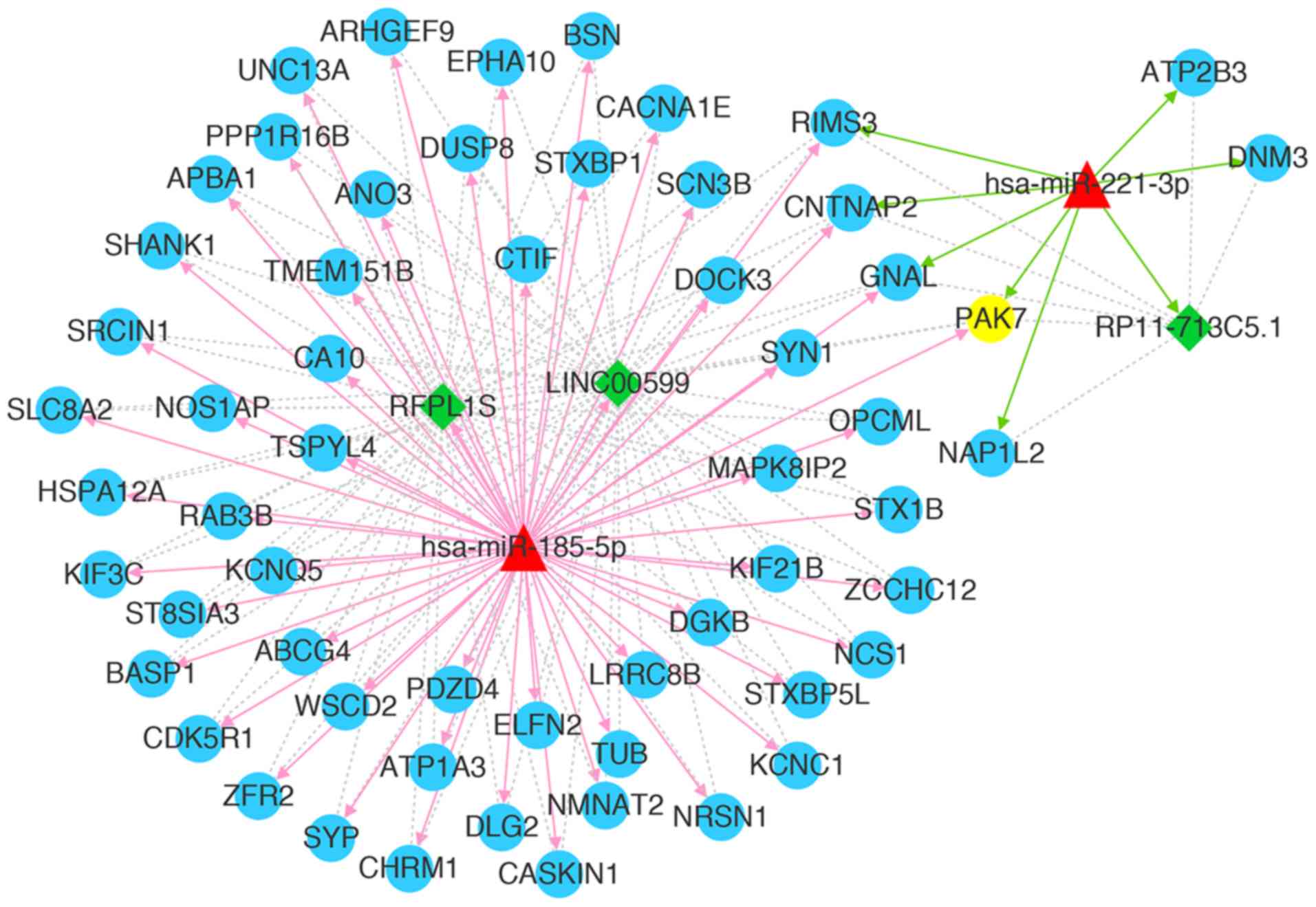 | Figure 5.Core competing endogenous RNA network
associated with PAK7. Green diamonds represent lncRNAs, blue
circles represent mRNAs (PAK7 is highlighted in yellow), red
triangles represent miRNAs, red arrows represent miRNA-lncRNA
regulatory relations, gray lines represent miRNA-mRNA regulatory
relations, green dotted lines represent mRNA-lncRNA co-expression
relations, red arrows represent relations with hsa-miR-185-5p,
green arrows represent relations with hsa-miR-221-3p. PAK7, p21
(RAC1)-activated kinase 7; miRNA/miR, microRNA; lncRNA, long
non-coding RNA. |
Survival analysis for core ceRNAs
A total of 143 samples with OS prognosis
information, and mRNA and lncRNA data were explored. PNMA5 was a
unique mRNA associated with OS prognosis upon grouping of all
mRNAs/lncRNAs based on the median values of expression (Fig. 6A). Hippocalcin-like 4 and SSTR1
were two outstanding mRNAs associated with OS prognosis following
grouping of all mRNAs/lncRNAs based on the mean value of expression
(Fig. 6B and C). KM analysis
showed that PNMA5, Hippocalcin-like 4 and SSTR1 may be the factors
associated with prognosis in patients with GBM.
RT-qPCR validation
The results of RT-qPCR showed that the mRNA levels
of PAK7 and SSTR1 were significantly downregulated in GBM samples
compared with control samples. Furthermore, LINC00599 expression
was downregulated and miR-185-5p expression was upregulated in GBM
samples (Fig. 7). These results
were consistent with the bioinformatics analysis results. However,
contrary to the bioinformatics analysis results, the mRNA
expression of PNMA5 was significantly downregulated in GBM samples
compared with control samples.
Discussion
GBM is the most common malignant brain tumor in
adults (35). Targeted gene
therapy such as PAK7 serves as a therapeutic strategy for glioma
(11). However, the detailed
mechanisms underlying the effects of PAK7 expression on GBM
progression remain incompletely understood. In the present study,
gene expression data of GBM were downloaded and reanalyzed using
bioinformatic tools. The results revealed a total of 954
upregulated and 1,234 downregulated DEGs between GBM and control
samples. These DEGs, including PAK7, were mainly enriched in
pathways such as axon guidance. ceRNA network analysis revealed
several outstanding ceRNA relationships, including
miR-185-5p-LINC00599-PAK7. Moreover, PNMA5 and SSTR1 were two
outstanding prognostic genes associated with OS in survival
analysis.
Bioinformatic approaches have been widely used for
the identification of genes that are associated with GBM. Wei et
al (36) identified DEGs
regulated by transcription factors in GBM by analyzing the GSE4290
dataset downloaded from the Gene Expression Omnibus (GEO) database.
Zhang et al (37) selected
GSE50161 from GEO database to identify potential oncogenes
associated with GBM progression. Moreover, Liu et al
(38) analyzed the glioma gene
expression profile dataset GSE4290 for the identification of DEGs.
These studies were performed based on datasets downloaded from GEO
database. In particular, the expression profile data in GEO usually
lack clinical data and may be unsuitable for survival analysis. In
the present study, a dataset obtained from TCGA database that
included clinical data was used, permitting survival analysis. In
addition to DEM selection, the potential lncRNAs were also
selected, and a regulatory network was constructed to explore the
potential molecular mechanism of GBM.
Axon guidance (also called axon pathfinding) is a
subfield of neural development related to the process by which
neurons send out axons to reach the correct targets (39). Agrawal et al (40) reported the development of acute
axonal polyneuropathy after the resection of GBM, indicative of the
close relationship between axon guidance and GBM. Axon guidance is
accomplished with the involvement of relatively few guidance
molecules (41). Certain guidance
molecules, such as PLEXIN-B2, may promote the tumorigenesis of GBM
through axon guidance pathways (42). The dysregulation in gene expression
implicated in the canonical axon guidance pathway influences the
glioma cell cycle (43).
Furthermore, PAK family members, including PAK1, PAK4 and PAK6,
commonly participate in tumorigenesis as guide molecules (44–46).
Hing et al (47)
demonstrated that PAK was not only a critical regulator of axon
guidance, but also served as a downstream effector of Dock in
vivo. Axon guidance was shown to be involved in the PAK-small
GTPase signaling in patients with schizophrenia (48). In the present study, PAK7 was one
of the most outstanding DEGs enriched in axon guidance pathway.
Thus, we speculate that PAK7 may participate in the tumorigenesis
of GBM via the axon guidance pathway.
Studies have shown that miRNAs may suppress the
process of tumorigenesis through the induction of cell
differentiation and cycle arrest (49). It was shown that the negative
expression of miR-185 in GBM may inhibit the proliferation of
glioma cell lines (50). Thus,
miR-185 is commonly used as a predictive biomarker for the
prognosis of malignant glioma (51,52).
miRNAs play important roles in tumor progression by targeting
lncRNAs (53). Tran et al
(54) reported that lncRNAs such
as LINC00176 may regulate cell cycle and survival by titrating the
tumor suppressor miR-185. Despite miRNA-lncRNA interactions, the
lncRNA-mRNA regulatory relations are also vital for tumor
progression (55). A comprehensive
analysis on the functional miRNA-mRNA regulatory network revealed
the association of miRNA signatures with glioma (56). As an mRNA, PAK7 is related with
tumor progression by targeting of miRNAs, including miR-492
(57). Pan et al (58) showed that miR-106a-5p inhibited the
migration and invasion of renal cell carcinoma cells by targeting
PAK7. Neuronal activity regulates spine formation, at least in
part, by increasing miRNA transcription, which in turn activates
PAK actin remodeling pathway (59). However, the association between
miR-185-PAK7 interactions and GBM progression is unclear. In the
present study, ceRNA network analysis showed that the
miR-185-5p-LINC00599-PAK7 axis was one of the regulatory
relationships most strongly associated with GBM. Thus, it is
hypothesized that the miR-185-5p may play an important role in GBM
progression by sponging the PAK7-LINC00599 interaction.
The members of the PNMA family have been identified
as onconeuronal antigens that are aberrantly expressed on cancer
cells (60). Distinct functional
domains of PNMA5 mediate protein-protein interactions, nuclear
localization and apoptosis signaling in human cancer cells
(60). As a novel tumor-associated
gene, PNMA5 plays an important role in maintaining the malignant
phenotype of hepatocellular carcinoma (61). However, studies on PNMA5 expression
in glioma or GBM are very rare. SSTR1 acts at several sites to
inhibit the release of hormones or secretory proteins (62). Kiviniemi et al (63) indicated that SSTR2 may serve as a
prognostic marker in high-grade glioma. The activation of SSTR1 was
shown to induce glioma growth arrest in vitro and in
vivo (64). However, whether
SSTR1 can be used as a prognostic marker in GBM is unclear. In the
present study, the survival analysis results showed that PNMA5 and
SSTR1 were the two most notable DEGs associated with overall
survival in disease. This result indicated that PNMA5 and SSTR1 may
be used as novel prognostic markers for GBM. However, certain
limitations of this study include the small sample size and lack of
verification. Thus, a larger sample size with more extensive
verification analysis is warranted in future investigations.
In conclusion, the present study suggested that PAK7
expression was downregulated in GBM, and that this gene may
participate in the tumorigenesis of GBM through the ceRNA pathway
of miR-185-5p-LINC00599-PAK7. Although the expression levels of
miR-185-5p, LINC00599 and PAK7 in GBM tissue were detected, the
regulatory relation of this ceRNA needs to be validated in future
studies.
Acknowledgements
Not applicable.
Funding
This work was supported by Torch Research Fund From
The Fourth Affiliated Hospital Of Harbin Medical University (grant
no. HYDSYHJ201902).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW was involved in the conception and design of the
research, participated in the acquisition of data and drafted the
manuscript. SL performed analysis and interpretation of data. ZS
performed analysis. PZ conceived the study, participated in its
design and co-ordination, and aided in drafting the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by Ethics Committee of The
Fourth Affiliated Hospital of Harbin Medical University and
Heilongjiang Provincial Hospital. Informed consent was provided by
all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol. 15
(Suppl 2):ii1–ii56. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Young RM, Jamshidi A, Davis G and Sherman
JH: Current trends in the surgical management and treatment of
adult glioblastoma. Ann Transl Med. 3:1212015.PubMed/NCBI
|
|
3
|
Nam JY and de Groot JF: Treatment of
glioblastoma. J Oncol Pract. 13:629–638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gallego O: Nonsurgical treatment of
recurrent glioblastoma. Curr Oncol. 22:e273–e281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bo LJ, Wei B, Li ZH, Wang ZF, Gao Z and
Miao Z: Bioinformatics analysis of miRNA expression profile between
primary and recurrent glioblastoma. Eur Rev Med Pharmacol Sci.
19:3579–3586. 2015.PubMed/NCBI
|
|
6
|
Guo Q, Zhang M, Hu G, Dai Y, Liu D and Yu
S: Bioinformatics analysis of differentially expressed genes in
glioblastoma. Acta Med Univ Sci Technol Huazhong. ((Issue 1)):
38–43. 2018.(In Chinese).
|
|
7
|
Gu X, Wang C, Wang X, Ma G, Li Y, Cui L,
Chen Y, Zhao B and Li K: Efficient inhibition of human glioma
development by RNA interference-mediated silencing of PAK5. Int J
Biol Sci. 11:230–237. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rane CK and Minden A: P21 activated
kinases: Structure, regulation, and functions. Small GTPases.
5(pii): e280032014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dan C, Nath N, Liberto M and Minden A:
PAK5, a new brain-specific kinase, promotes neurite outgrowth in
N1E-115 cells. Mol Cell Biol. 22:567–577. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pandey A, Dan I, Kristiansen TZ, Watanabe
NM, Voldby J, Kajikawa E, Khosravi-Far R, Blagoev B and Mann M:
Cloning and characterization of PAK5, a novel member of
mammalianp21-activated kinase-II subfamily that is predominantly
expressed in brain. Oncogene. 21:3939–3948. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han ZX, Wang XX, Zhang SN, Wu JX, Qian HY,
Wen YY, Tian H, Pei DS and Zheng JN: Downregulation of PAK5
inhibits glioma cell migration and invasion potentially through the
PAK5-Egr1-MMP2 signaling pathway. Brain Tumor Pathol. 31:234–241.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhai J, Qu S, Li X, Zhong J, Chen X, Qu Z
and Wu D: miR-129 suppresses tumor cell growth and invasion by
targeting PAK5 in hepatocellular carcinoma. Biochem Biophys Res
Commun. 464:161–167. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harrow J, Frankish A, Gonzalez JM,
Tapanari E, Diekhans M, Kokocinski F, Aken BL, Barrell D, Zadissa
A, Searle S, et al: GENCODE: The reference human genome annotation
for The ENCODE project. Genome Res. 22:1760–1774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Quadrianto N and Buntine WL: Linear
regression. Encyclopedia of Machine Learning. Springer. (Boston,
MA). 2016.
|
|
15
|
Koop G: Bayesian methods for empirical
macroeconomics with big data. Rev Econ Anal. 9:33–56. 2017.
|
|
16
|
Smyth GK: Limma: Linear models for
microarray data. Bioinformatics and Computational Biology Solutions
Using R and Bioconductor. Statistics for Biology and Health, .
Gentleman R, Carey VJ, Huber W, Irizarry RA and Dudoit S: Springer;
New York, NY: pp. 397–420. 2005, View Article : Google Scholar
|
|
17
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J Roy Stat Soc B. 57:289–300. 1995.
|
|
18
|
Yekti YND and Yassierli: Kansei
engineering using non-metric multidimensional scaling (NMDS) and
cluster analysis methods. Seanes International Conference on Human
Factors and Ergonomics in South-East Asia. 2016.
|
|
19
|
Oksanen J, Blanchet F, Kindt R, Legendre
P, Minchin R, O'Hara R, Oksanen J, Blanchet FG, Kindt R, Legendre
P, et al: vegan: Community ecology package version 2.0–10. J Stat
Softw. 48:103–132. 2013.
|
|
20
|
Clark KR: Non-parametric multivariate
analysis of changes in community structure. Aust J Ecol.
18:117–143. 1993. View Article : Google Scholar
|
|
21
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47(D1): D590–D595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Breuer K, Foroushani AK, Laird MR, Chen C,
Sribnaia A, Lo R, Winsor GL, Hancock RE, Brinkman FS and Lynn DJ:
InnateDB: Systems biology of innate immunity and beyond-recent
updates and continuing curation. Nucleic Acids Res. 41((Database
Issue)): D1228–D1233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dweep H and Gretz N: miRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Moreno R, Miranda DR, Fidler V and Van SR:
Evaluation of two outcome prediction models on an independent
database. Crit Care Med. 26:50–61. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X: miRDB: A microRNA target
prediction and functional annotation database with a wiki
interface. RNA. 14:1012–1017. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vejnar CE and Zdobnov EM: MiRmap:
Comprehensive prediction of microRNA target repression strength.
Nucleic Acids Res. 40:11673–11683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miranda K, Huynh T, Tay Y, Ang YS, Tam WL,
Thomson AM, Lim B and Rigoutsos I: A pattern-based method for the
identification of MicroRNA binding sites and their corresponding
heteroduplexes. Cell. 126:1203–1217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Grimson A, Farh KK, Johnston WK,
Garrett-Engele P, Lim LP and Bartel DP: MicroRNA targeting
specificity in mammals: Determinants beyond seed pairing. Mol Cell.
27:91–105. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database Issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Das S, Ghosal S, Sen R and Chakrabarti J:
lnCeDB: Database of human long noncoding RNA acting as competing
endogenous RNA. PLoS One. 9:e989652014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alberti C, Timsit JF and Chevret S:
Survival analysis-the log rank test. Rev Mal Respir. 22:829–832.
2005.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bland JM and Altman DG: Survival
probabilities (the Kaplan-Meier method). BMJ. 317:15721998.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ni Y, Zhang F, An M, Yin W and Gao Y:
Early candidate biomarkers found from urine of glioblastoma
multiforme rat before changes in MRI. Sci China Life Sci.
61:982–987. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei B, Wang L, Du C, Hu G, Wang L, Jin Y
and Kong D: Identification of differentially expressed genes
regulated by transcription factors in glioblastomas by
bioinformatics analysis. Mol Med Rep. 11:2548–2554. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Xia Q and Lin J: Identification
of the potential oncogenes in glioblastoma based on bioinformatic
analysis and elucidation of the underlying mechanisms. Oncol Rep.
40:715–725. 2018.PubMed/NCBI
|
|
38
|
Liu M, Xu Z, Du Z, Wu B, Jin T, Xu K, Xu
L, Li E and Xu H: The identification of key genes and pathways in
glioma by bioinformatics analysis. J Immunol Res. 2017:12780812017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Suter TACS, DeLoughery ZJ and Jaworski A:
Meninges-derived cues control axon guidance. Dev Biol. 430:1–10.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Agrawal R, Chugh C, Mukherji JD and Singh
P: Acute axonal polyneuropathy following resection of a
glioblastoma multiforme. Neurol India. 65:1422–1423. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pace KR, Dutt R and Galileo DS: Exosomal
L1CAM stimulates glioblastoma cell motility, proliferation, and
invasiveness. Int J Mol Sci. 20(pii): E39822019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang Y, Tejero-Villalba R, Kesari S, Zou
H and Friedel R: ANGI-19. The axon guidance receptor Plexin-B2
promotes tumorigenicity of glioblastoma. Neuro Oncol. 19 (Suppl
6):vi252017. View Article : Google Scholar
|
|
43
|
Kunapuli P, Lo K, Hawthorn L and Cowell
JK: Reexpression of LGI1 in glioma cells results in dysregulation
of genes implicated in the canonical axon guidance pathway.
Genomics. 95:93–100. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang G, Song Y, Liu T, Wang C, Zhang Q,
Liu F, Cai X, Miao Z, Xu H, Xu H, et al: PAK1-mediated MORC2
phosphorylation promotes gastric tumorigenesis. Oncotarget.
6:9877–9886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
He LF, Xu HW, Chen M, Xian ZR, Wen XF,
Chen MN, Du CW, Huang WH, Wu JD and Zhang GJ: Activated-PAK4
predicts worse prognosis in breast cancer and promotes
tumorigenesis through activation of PI3K/AKT signaling. Oncotarget.
8:17573–17585. 2016. View Article : Google Scholar
|
|
46
|
Roberts JA: An investigation of the role
of PAK6 in tumorigenesis. Diss Theses Gradworks. 2012.
|
|
47
|
Hing H, Xiao J, Harden N, Lim L and
Zipursky SL: Pak functions downstream of Dock to regulate
photoreceptor axon guidance in Drosophila. Cell. 97:853–863. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen SY, Huang PH and Cheng HJ:
Disrupted-in-Schizophrenia 1-mediated axon guidance involves
TRIO-RAC-PAK small GTPase pathway signaling. Proc Natl Acad Sci
USA. 108:5861–5866. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhao Z, Ma X, Sung D, Li M, Kosti A, Lin
G, Chen Y, Pertsemlidis A, Hsiao TH and Du L: microRNA-449a
functions as a tumor suppressor in neuroblastoma through inducing
cell differentiation and cell cycle arrest. RNA Biol. 12:538–554.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Morrow C, Smirnov I, Adai A, Yeh RF,
Mistra A and Feuerstein B: MIR-185 is lost in glioblastoma
multiforme (GBM) and inhibits proliferation in glioma cell lines.
Meet Soc Neuro Oncol. 7672008.
|
|
51
|
Tang H, Liu Q, Liu X, Ye F, Xie X, Xie X
and Wu M: Plasma miR-185 as a predictive biomarker for prognosis of
malignant glioma. J Cancer Res Ther. 11:630–634. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tang H, Wang Z, Liu X, Liu Q, Xu G, Li G
and Wu M: LRRC4 inhibits glioma cell growth and invasion through a
miR-185-dependent pathway. Curr Cancer Drug Targets. 12:1032–1042.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Paraskevopoulou MD and Hatzigeorgiou AG:
Analyzing MiRNA-LncRNA interactions. Methods Mol Biol.
1402:271–286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tran DDH, Kessler C, Niehus SE, Mahnkopf
M, Koch A and Tamura T: Myc target gene, long intergenic noncoding
RNA, Linc00176 in hepatocellular carcinoma regulates cell cycle and
cell survival by titrating tumor suppressor microRNAs. Oncogene.
37:75–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Amirkhah R, Schmitz U, Linnebacher M,
Wolkenhauer O and Farazmand A: MicroRNA-mRNA interactions in
colorectal cancer and their role in tumor progression. Genes
Chromosomes Cancer. 54:129–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li Y, Xu J, Chen H, Bai J, Li S, Zhao Z,
Shao T, Jiang T, Ren H, Kang C and Li X: Comprehensive analysis of
the functional microRNA-mRNA regulatory network identifies miRNA
signatures associated with glioma malignant progression. Nucleic
Acids Res. 41:e2032013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Song X, Xie Y, Liu Y, Shao M and Yang W:
MicroRNA-492 overexpression exerts suppressive effects on the
progression of osteosarcoma by targeting PAK7. Int J Mol Med.
40:891–897. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pan YJ, Wei LL, Wu XJ, Huo FC, Mou J and
Pei DS: MiR-106a-5p inhibits the cell migration and invasion of
renal cell carcinoma through targeting PAK5. Cell Death Dis.
8:e31552017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Impey S, Davare M, Lasiek A, Fortin D,
Ando H, Varlamova O, Obrietan K, Soderling TR, Goodman RH and
Wayman GA: An activity-induced microRNA controls dendritic spine
formation by regulating Rac1-PAK signaling. Mol Cell Neurosci.
43:146–156. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lee YH, Pang SW, Poh CL and Tan KO:
Distinct functional domains of PNMA5 mediate protein-protein
interaction, nuclear localization, and apoptosis signaling in human
cancer cells. J Cancer Res Clin Oncol. 142:1967–1977. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Teng XM, Deng Q, Han ZG and Huang J:
Expression of PNMA5 in hepatocellular carcinoma tissues and its
function. Tumor. 28:911–915. 2008.
|
|
62
|
Bruns C, Weckbecker G, Raulf F, Kaupmann
K, Schoeffter P, Hoyer D and Lübbert H: Molecular pharmacology of
somatostatin receptor subtypes. Ann N Y Acad Sci. 733:138–146.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kiviniemi A, Gardberg M, Frantzén J,
Pesola M, Vuorinen V, Parkkola R, Tolvanen T, Suilamo S, Johansson
J, Luoto P, et al: Somatostatin receptor subtype 2 in high-grade
gliomas: PET/CT with (68)Ga-DOTA-peptides, correlation to
prognostic markers, and implications for targeted radiotherapy.
EJNMMI Res. 5:252015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Gatti M, Pattarozzi A, Würth R, Barbieri F
and Florio T: Somatostatin and somatostatin receptors 1, 2 and 5
selective agonists inhibit C6 glioma cell growth in vitro and in
vivo: Analysis of activated intracellular pathways. Regulat
Peptides. 164:382010. View Article : Google Scholar
|