Introduction
Myocardial ischemia/reperfusion (MI/R) injury is an
acute adverse cardiac event, resulting from the deterioration or
loss of cardiac function, thereby reducing the efficacy of
reperfusion therapy (1). Despite
recent improvements in medical knowledge and management, MI/R and
related heart failure are still significant health problems
worldwide (2). The mechanisms of
MI/R pathogenesis are complex and multifactorial, including
endothelial cell injury, apoptosis, marked oxidative/nitrosative
stress, inflammation, mitochondrial dysfunction, dysregulated
vascular relaxation and intracellular calcium overload (3). Due to the lack of a comprehensive
understanding of the pathological processes occurring during I/R
injury, there are no definitive interventions to eliminate MI/R
injury (4,5). Therefore, it is important to explore
novel therapeutic strategies to alleviate MI/R injury and elucidate
the underlying mechanisms.
Troxerutin is a trihydroxyethylated derivative of
the natural bioflavonoid rutin and is found in tea, corn buds,
coffee, fruit and green vegetables (6,7).
Troxerutin has been found to have multiple biological properties,
including having anti-oxidative, anti-inflammatory,
anti-fibrinolytic, anti-neoplastic and neuroprotective activities
(7–9). Recently, emerging evidence has
revealed the cardioprotective potential of troxerutin (10,11).
Mokhtari et al (12)
reported that troxerutin attenuates myocardial reperfusion injury
in diabetic rats, which is dependent on its anti-apoptotic
function. Another previous study demonstrated that troxerutin
treatment, as well as ischemic postconditioning, inhibited the
activation of leukocyte-endothelial cell interactions and prevented
inflammatory-pathological changes under I/R insults in myocardial
cells (13). Although there are
studies that have reported the protective effect of troxerutin
preconditioning following MI/R injury, the specific effects of
troxerutin on MI/R injury, and the underlying mechanisms, have not
been fully elucidated under healthy and disease conditions.
Cardiomyocyte oxidative stress and inflammatory
responses have been recognized as hallmarks of MI/R injury
(14,15). Previous studies reported that
oxidative stress is increased or accelerated during I/R, and
partially contributes to the overall level of apoptosis and
cardiomyocyte death (16,17). Similarly, inflammation plays an
important role in the pathophysiology of MI/R injury (18). Reducing oxidative stress and the
inflammatory response minimizes cardiac injury induced by I/R
(19). In addition, the PI3K/AKT
signaling pathway has been identified as a potential therapeutic
target in the treatment of MI/R injury (20,21).
Hypoxia-inducible factor-1α (HIF-1α), a transcription factor that
is a central component of the oxygen sensing mechanism in mammalian
cells, has been shown to be an important regulator in the response
to hypoxia and ischemia (22,23).
Previous studies have revealed that enhancing the expression of
HIF-1α reduces infarct size, promotes angiogenesis and improves
cardiac function (24,25). PI3K/AKT and HIF-1α also play a
substantial role in mediating the inflammatory response and
oxidative stress (26,27). A previous study showed that HIF-1α
is a downstream target of the PI3K/AKT pathway and is involved in
lung I/R injury (28), cerebral
infarction (29) and oral squamous
cell carcinoma (30).
Nevertheless, the role of the PI3K/AKT/HIF-1α signaling pathway in
the pathogenesis of MI/R injury and the protective effect of
troxerutin on MI/R injury remains unclear.
The aim of the present study was to investigate the
effects of troxerutin on oxidative stress and inflammation
following MI/R injury in H9C2 cells, with an emphasis on the role
of the PI3K/AKT/HIF-1α signaling pathway.
Materials and methods
H9C2 cardiomyocyte culture
The H9C2 cardiomyocyte line, a subclone of the
original clonal cell line derived from embryonic rat heart tissue,
was obtained from the American Type Cell Culture Collection. The
cells were cultured in DMEM (Sigma-Aldrich; Merck KGaA)
supplemented with (v/v) 10% FBS (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 U/ml streptomycin in a
humidified incubator at 37°C with an atmosphere of 5%
CO2 and 95% air.
Establishment of the Oxygen-Glucose
Deprivation and Reoxygenation (OGD/R) injury model
To create an in vitro model of MI/R injury,
H9C2 cells were maintained in glucose-free Hanks' Balanced Salt
Solution (Invitrogen; Thermo Fisher Scientific, Inc.) within an
anaerobic chamber containing 95% N2 and 5%
CO2 at 37°C for 6 h, which induced OGD. Subsequently,
H9C2 cells from the OGD-treated groups were removed from the anoxic
incubator and cultured under normal condition as aforementioned for
a further 18 h to allow reoxygenation to occur. This process was
called MI/R injury.
Cell transfection
Small interfering (si)RNAs against HIF-1α
(HIF-1α-siRNA) and non-specific control siRNA (NS-siRNA) were
synthesized by Shanghai GenePharma Co., Ltd. The sense strand siRNA
sequences are as follows: HIF-1α-siRNA, 5′-GCCGCUCAAUUUAUGAAUATT-3′
and NS-siRNA, 5′-UUCUCCGAACGUGUCACGUTT-3′. H9C2 cells were
transfected with HIF-1α-siRNA (50 nM) or NS-siRNA (50 nM) using
Lipofectamine® RNAiMAX (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
Briefly, the cells were seeded in 6-well plates at a density of
5×105 cells/per well. HIF-1α-siRNA, NS-siRNA or
Lipofectamine® RNAiMAX transfection reagent was diluted
in Opti-MEM medium (Gibco; Thermo Fisher Scientific, Inc.). The
diluted siRNA was added to the diluted Lipofectamine®
RNAiMAX reagent in a 1:1 ratio. Following incubation at room
temperature for 5 min, the siRNA-lipid complexes were added to the
cells and incubated for a further 6 h, after which the media was
changed to complete media. The cells were harvested 48 h
post-transfection and the transfection efficiency was determined
using reverse transcription-quantitative (RT-q)PCR.
Experimental protocols
A previous study and preliminary experimental
results indicated that troxerutin (10 µM) addition to cultured
myocardial cells for 1 h before OGD exerts cardioprotective effects
(31). LY294002 (20 µM; Merck
KGaA), a specific inhibitor of the PI3K/AKT signaling pathway, was
used to pretreat H9C2 cardiomyocytes for 2 h before troxerutin
treatment (32).
In the present study, cells were divided into the
following 6 groups: i) Control group, H9C2 cells were cultured
under normal conditions in high-glucose DMEM; ii) OGD/R group, H9C2
cells were subjected to OGD for 6 h and reoxygenation for 18 h;
iii) troxerutin + OGD/R group, H9C2 cells were incubated with
troxerutin (10 µM) for 1 h followed by co-treatment with OGD/R; iv)
LY294002 + troxerutin + OGD/R group, cells were pre-conditioned
with LY2940021 (20 µM) for 2 h, troxerutin (10 µM) for 1 h and then
subjected to OGD/R; v) siRNA+ troxerutin + OGD/R group, H9C2 cells
were transfected with HIF-1α-siRNA (50 nM) or NS-siRNA (50 nM) for
24 h and then treated with troxerutin (10 µM) for 1 h followed by
OGD/R; and vi) LY294002 + siRNA, H9C2 cells were incubated with
LY2940021 (20 µM) for 24 h and treated with HIF-1α-siRNA (50 nM) or
NS-siRNA (50 nM) for 24 h.
Cell viability assay
The cell viability of H9C2 cells treated as
aforementioned was analyzed using an MTT assay. Briefly, the cells
were seeded in 96-well plates at a density of 1×104
cells/well. Following the aforementioned treatments, MTT solution
(0.5 mg/ml) was added to the each well and incubated with the cells
at 37°C for 4 h. Following this, the supernatants were removed and
150 µl DMSO was added to each well to dissolve the formazan
crystals. The absorbance at 570 nm was measured using a microplate
reader (Bio-Rad Laboratories, Inc.).
Cell cytotoxicity measurement
H9C2 cells were cultured in 96-well plates at a
density of 1×104 cells/well and subjected to the
aforementioned treatments. Extracellular lactate dehydrogenase
(LDH) activity was determined using a commercial LDH assay kit
(Nanjing KeyGen Biotech Co., Ltd.), according to the manufacturer's
instructions. The absorbance at 490 nm was measured using a
microplate reader.
Cell apoptosis assay
H9c2 cells were treated as aforementioned and the
level of apoptosis was determined using an Annexin V-FITC/PI kit
(Becton, Dickinson and Company), according to the manufacturer's
instructions. After treatment, the cells were harvested, washed
twice with PBS and stained with annexin V-FITC (5 µl) and propidium
iodide (10 µl) solution for 15 min at 37°C in the dark. The level
of apoptosis was determined using a flow cytometer (FACScan; BD
Biosciences) and quantified using CELL Quest 3.0 software (BD
Biosciences).
RT-qPCR
Total RNA was isolated from H9C2 cells using TRIzol
Reagent (Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. First-strand complementary DNA was
synthesized using 1 µg RNA using the PrimeScript RT Reagent kit
with gDNA Eraser (Takara Bio, Inc.). The temperature protocol used
for the RT reaction consisted of 37°C for 15 min and at 90°C for 5
sec. The mRNA levels of HIF-1α, interleukin (IL)-1β, IL-6 and tumor
necrosis factor (TNF)-α were analyzed using the SYBR Premix Ex Taq
kit (Takara Bio, Inc.) and an ABI PRISM 7500 PCR instrument
(Applied Biosystems; Thermo Fisher Scientific, Inc.). qPCR
thermocycling conditions were: 95°C for 90 sec for an initial
denaturation, 45 denaturation cycles at 94°C for 6 sec, followed by
60°C for 30 sec for annealing and elongation, and final 72°C for 8
min for extension. β-actin was used as an internal control and the
relative level of mRNA was analyzed using the 2−ΔΔCq
method (33). The primer sequences
used are as follows: HIF-1α forward, 5′-GATGACGGCGACATGGTTTAC-3′
and reverse, 5′-CTCACTGGGCCATTTCTGTGT-3′; IL-1β forward,
5′-GCCTCGTGCTGTCGGACCCATAT-3′ and reverse,
5′-TCCTTTGAGGCCCAAGGCCACA-3′; IL-6 forward,
5′-ACAACCACGGCCTTCCCTACTT-3′ and reverse,
5′-CACGATTTCCCAGAGAACATGTG-3′; TNF-α forward,
5′-CCCTCCTGGCCAACGGCATG-3′ and reverse,
5′-TCGGGGCAGCCTTGTCCCTT-3′); and β-actin forward,
5′-CGTGCGTGACATCAAAGAGAAG-3′ and reverse,
5′-CCAAGAAGGAAGGCTGGAAAA-3′.
Intracellular reactive oxygen species
(ROS) generation
H9C2 cells were seeded at a density of
1×105 cells/ml in 6-well plates and the production of
intracellular ROS was determined using an ROS assay kit (Beyotime
Institute of Biotechnology). Briefly, following the aforementioned
treatments, H9C2 cells were washed twice with PBS and incubated
with 2′,7′-dichlorodihydrofluorescein diacetate (10 µM) at 37°C for
20 min. The fluorescence intensity was observed under a
fluorescence microscope (Olympus Corporation, magnification, ×200).
For the quantitative assay, the fluorescence intensity was measured
using a flow cytometer at an excitation wavelength of 485 nm and an
emission wavelength of 525 nm.
Measurement of oxidative stress
biomarkers
The H9C2 cells from each group were homogenized with
Scientz-IID Ultrasound Cell lysis Instrument (Changchun, Hangzhou,
15 sec/3 times pyrolysis and 15 sec for interval) on ice to
determine the malondialdehyde (MDA) content, and the activities of
superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px). The
MDA content was determined using the thiobarbituric acid method,
the activity of was measured using the xanthine oxidase method and
the activity of GSH-Px was determined using the
dithio-dinitrotoluidine method (all Nanjing Jiancheng
Bioengineering Institute), according to the manufacturer's
instructions.
Western blot analysis
Following the aforementioned treatments, cells were
lysed in ice-cold RIPA Buffer (Beyotime Institute of Biotechnology)
containing 1% (v/v) Combined protease and phosphatase inhibitors
(Thermo Fisher Scientific, Inc.). The cell lysates were centrifuged
at 13,000 × g at 4°C for 10 min. The protein concentration in each
sample was determined using the bicinchoninic acid method. Equal
amounts of protein (30 µg) were separated on 10% SDS-PAGE and
subsequently transferred to PVDF membranes. After blocking with 5%
(w/v) non-fat dry milk in PBS with Tween (PBST) at room temperature
for 2 h. The following primary antibodies were diluted 1:2,000 and
incubated overnight at 4°C with the membranes (all Cell Signaling
Technology, Inc.): PI3K (cat. no. 4249), phosphorylated AKT (p-AKT,
cat. no. 9271), AKT (cat. no. 4691), HIF-1α (cat. no. 36169) and
GAPDH (cat. no. 5174). After washing with TBST three times, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies (Cell Signaling Technology, Inc., cat. no.
7074) at room temperature for 2 h. The membranes were washed and
the protein bands were visualized using enhanced chemiluminescence
(ECL; Pierce; Thermo Fisher Scientific, Inc.). Optical density
values for each group were normalized to GAPDH and quantified using
Quantity One 4.6.2 software (Bio-Rad, Laboratories, Inc.).
ELISA
Following the aforementioned treatments, the levels
of TNF-α (Thermo Fisher Scientific, Inc., cat. no. BMS622), IL-6
(Thermo Fisher Scientific, Inc., cat. no. BMS603-2) and IL-1β
(E-EL-R0012c, Elabscience) in the supernatant were evaluated using
ELISA, according to the manufacturer's instructions. The results of
the ELISAs were measured using a plate reader at 450 nm.
Statistical analysis
Statistical analysis was performed using a one-way
ANOVA followed by the least significant difference test using SPSS
software, version 17.0 (SPSS, Inc.). All data are presented as the
mean ± SD from three experiments. P<0.05 was considered to
indicate a statistically significant difference.
Results
Troxerutin alleviates OGD/R-induced
H9C2 cell injury
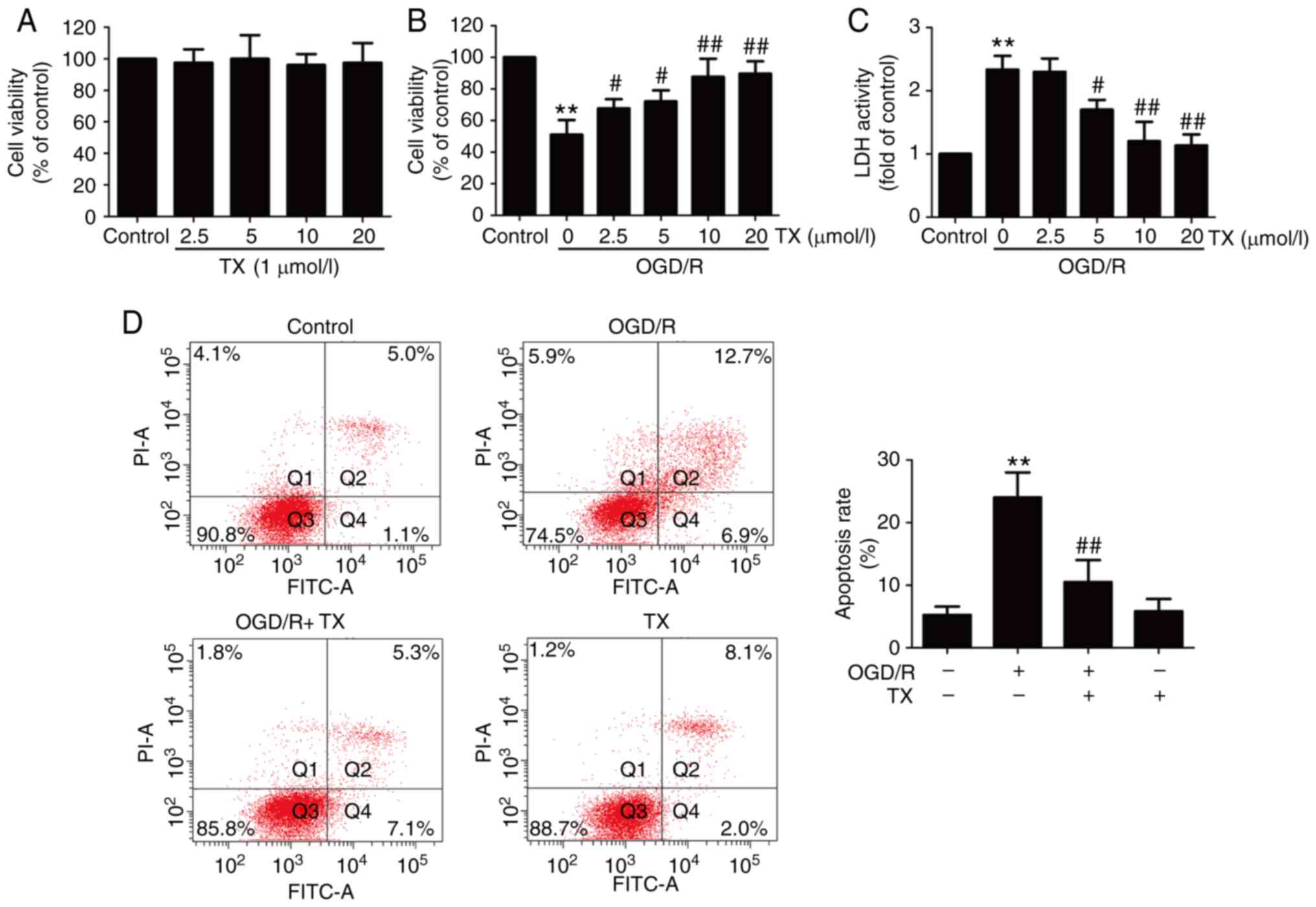
To examine the protective effect of troxerutin on
OGD/R injury, cell viability was assessed. The results revealed
that the different concentrations of troxerutin treatment had no
effect on cell viability (Fig.
1A), and Results from the MTT assay demonstrated that OGD/R
treatment resulted in reduced cell viability, which was reversed by
troxerutin (Fig. 1B). In addition,
ODG/R-induced an increase in LDH activity that was also blocked by
troxerutin (Fig. 1C).
Subsequently, the effect of troxerutin on apoptosis in
OGD/R-treated H9C2 cells was also measured. The results indicated
that OGD/R treatment induced apoptosis (Fig. 1D) in H9C2 cells. However, these
effects were attenuated by troxerutin treatment. These data
demonstrated that troxerutin exerted cardioprotective effects
against OGD/R injury in H9C2 cells.
Troxerutin activates the
PI3K/AKT/HIF-1α signaling pathway in OGD/R-treated H9C2 cells
To elucidate the potential underlying mechanism of
troxerutin, the impact of troxerutin on the PI3K/AKT/HIF-1α
signaling pathway was evaluated in OGD/R-injured H9C2 cells. OGD/R
treatment significantly reduced the expressions of PI3K (Fig. 2A and B) and p-AKT (Fig. 2A and C), indicating that OGD/R
inhibited the PI3K/AKT pathway. However, the expression of PI3K and
p-AKT was reversed by troxerutin. In addition, troxerutin also
significantly reversed the OGD/R-induced decrease in the expression
of HIF-1α (Fig. 2A and D). Taken
together, these data suggested that the PI3K/AKT/HIF-1α signaling
pathway may play an important role in the cardioprotective effect
of troxerutin against myocardial OGD/R damage.
PI3K/AKT/HIF-1α signaling pathway
mediates the troxerutin-induced suppression of H9C2 cell damage
under OGD/R conditions
Based on the aforementioned results, the effect of
the PI3K/AKT pathway inhibitor LY294002 on the cardioprotective
effects of troxerutin was investigated. LY294002 pretreatment
significantly mitigated the troxerutin-induced upregulation of PI3K
(Fig. 3A and B), p-AKT (Fig. 3A and C) and HIF-1α (Fig. 3A and D) in OGD/R-treated H9C2
cells, indicating that LY294002 results in the inhibition of the
PI3K/AKT/HIF-1α signaling pathway. Meanwhile, compared with
troxerutin + OGD/R group, pretreatment with LY294002 reversed
troxerutin-induced increased cell viability (Fig. 3E) and decreased LDH activity
(Fig. 3F) in OGD/R-treated H9C2
cells. These results suggested that troxerutin exerted its
cardioprotective effects against OGD/R injury by enhancing
PI3K/AKT-mediated HIF-1α signaling pathway.
 | Figure 3.Effects of LY294002 on the
PI3K/AKT/HIF-1α signaling pathway, cell viability and LDH activity
in the presence or absence of troxerutin in OGD/R-treated H9C2
cells. H9C2 cells were pretreated with PI3K pathway inhibitor
LY294002 (20 µM) for 2 h and then treated with troxerutin (10 µM)
followed by OGD (6 h)/R (18 h) treatment. (A) The expression levels
of PI3K, AKT, p-AKT and HIF-1α were determined using western blot
analysis. Quantitative analysis of (B) PI3K, (C) p-AKT/AKT and (D)
HIF-1α expression in H9C2 cells. (E) Cell viability in each group
was determined using the MTT assay. (F) LDH activity was determined
using a commercial LDH assay kit. Data are presented as the mean ±
SD (n=3). *P<0.05, **P<0.01 vs. -OGD/R-troxerutin-LY294002
group; #P<0.05, ##P<0.01 vs.
+OGD/R-troxerutin-LY294002 group; &P<0.05,
&&P<0.01 vs. +OGD/R +troxerutin-LY294002
group. OGD/R, oxygen-glucose deprivation and reoxygenation; LDH,
lactate dehydrogenase; HIF-1α, hypoxia-inducible factor-1α. |
HIF-1α knockdown blocks the
cardioprotective effects of troxerutin against OGD/R injury in H9C2
cells
Next, an siRNA against HIF-1α was used to further
investigate the underlying mechanism of troxerutin. H9C2 cells were
transfected with HIF-1α-siRNA or NS-siRNA and the transfection
efficiency was determined using RT-qPCR. The results showed that
compared with NS-siRNA transfection group, HIF-1α-siRNA
transfection significantly decreased the level of HIF-1α mRNA in
the control, OGD/R group and troxerutin + OGD/R groups (Fig. 4A). HIF-1α-siRNA further reduced
cell viability and reduced the troxerutin-induced increase in cell
viability in OGD/R-treated H9C2 cells (Fig. 4B). HIF-1α-siRNA also exacerbated
the OGD/R-mediated increase in LDH activity, and, notably,
HIF-1α-siRNA did not eliminate the inhibition of troxerutin on LDH
activity in troxerutin + OGD/R-treated H9C2 cells (Fig. 4C). Additionally, HIF-1α-siRNA
aggravated apoptosis in the OGD/R-treated group (compared with the
NS-siRNA + OGD/R group) and reduced the troxerutin-induced decrease
in apoptosis rate in the troxerutin + OGD/R-treated group (compared
with the troxerutin + OGD/R group) (Fig. 4D and E). Notably, the effects of
troxerutin on apoptosis were not abolished by HIF-1α-siRNA, meaning
other pathways may contribute to the cardioprotection of troxerutin
against apoptosis. These results indicated that troxerutin
protected against OGD/R-induced cytotoxicity partly depending on
the PI3K/AKT/HIF-1α signaling pathway, while the effects of
troxerutin attenuated apoptosis independent of the PI3K/AKT/HIF-1α
signaling pathway.
HIF-1α knockdown mitigates
troxerutin-induced inhibition of oxidative stress in OGD/R-treated
H9C2 cells
To further explore the effect of troxerutin on
oxidative stress and the role of the PI3K/AKT/HIF-1α signaling
pathway in this process, oxidative stress markers were assessed in
H9C2 cells. Compared with the control group, OGD/R treatment
increased ROS generation (Fig. 5A and
B) and the MDA content (Fig.
5C) in H9C2 cells, while compared with OGD/R group, troxerutin
pretreatment decreased ROS production (Fig. 5A and B) and the MDA content
(Fig. 5C) in OGD/R-treated H9C2
cells, highlighting the anti-oxidative properties of troxerutin
under OGD/R condition in H9C2 cells. However, compared with
NS-siRNA group, the inhibition of troxerutin on oxidative stress
was partly reversed by HIF-1α-siRNA in TX + OGD/R group. In
addition, troxerutin alleviated the OGD/R-induced decreases in
antioxidant stress indicators, including SOD (Fig. 5D) and GSH-Px activities (Fig. 5E), while these effects were reduced
by HIF-1α-siRNA. Compared with the NS-siRNA transfection group,
HIF-1α-siRNA induced oxidative stress in the OGD/R treatment group
as shown by the increased generation of ROS and MDA content, and
the reduced activities of SOD and GSH-Px. Notably, the effect of
troxerutin on oxidative stress was not totally abolished by the
knockdown of HIF-1α, the effect was only reduced, meaning that
other molecule or pathways may be involved in the protection of
troxerutin against oxidative stress. These results suggested that
troxerutin attenuated OGD/R-induced oxidative stress, partly by
enhancing PI3K/AKT/HIF-1α signaling pathway.
 | Figure 5.Effects of HIF-1α-siRNA on oxidative
and antioxidative products in the presence or absence of TX in
OGD/R-treated H9C2 cells. H9C2 cells were transfected with
HIF-1α-siRNA or NS-siRNA for 6 h, and then treated with TX (10 µM)
followed by OGD (6 h)/R (18 h) treatment. (A) Intracellular ROS
generation was measured using a DCFH-DA assay and the fluorescence
intensity was observed under a fluorescent microscope. (B) The
DCFH-DA assay was quantitatively analyzed using a flow cytometer.
The (C) MDA content, (D) SOD activity and (E) GSH-Px activity were
measured using commercial kits. Data are presented as the mean ± SD
(n=3). *P<0.05, **P<0.01 vs. NS-siRNA + control;
#P<0.05, ##P<0.01 vs. NS-siRNA + OGD/R;
&P<0.05 vs. NS-siRNA + TX + OGD/R group. OGD/R,
oxygen-glucose deprivation and reoxygenation; siRNA, small
interfering RNA; HIF-1α, hypoxia-inducible factor-1α; NS,
non-specific; TX, troxerutin; ROS, reactive oxygen species; MDA,
malonaldehyde; SOD, superoxide dismutase; GSH-Px, glutathione
peroxidase; DCFH-DA, 2′,7′-dichlorodihydrofluorescein
diacetate. |
HIF-1α knockdown eliminates the
protective effect of troxerutin on the inflammatory response in
OGD/R-treated H9C2 cells
The effect of troxerutin on inflammation in
OGD/R-treated H9C2 cells was investigated. Troxerutin treatment
mitigated OGD/R-induced increases in inflammatory markers including
IL-1β (Fig. 6A), IL-6 (Fig. 6B) and TNF-α (Fig. 6C) mRNA expression in H9C2 cells,
while these effects were reduced by HIF-1α-siRNA. In addition,
OGD/R-induced an increase in the levels of IL-1β (Fig. 6D), IL-6 (Fig. 6E), and TNF-α (Fig. 6F) in the culture supernatant, which
were also reversed by troxerutin treatment. However, these effects
of troxerutin on the inflammatory markers reduced by HIF-1α-siRNA.
These results indicated that troxerutin attenuated OGD/R-induced
inflammation by promoting the PI3K/AKT/HIF-1α signaling
pathway.
 | Figure 6.Effects of HIF-1α-siRNA on
inflammatory cytokines in the presence or absence of TX in
OGD/R-treated H9C2 cells. H9C2 cells were transfected with
HIF-1α-siRNA or NS-siRNA for 6 h and then treated with TX (10 µM)
followed by OGD (6 h)/R (18 h) treatment. The expression levels of
(A) IL-1β, (B) IL-6 and (C) TNF-α were determined using reverse
transcription-quantitative PCR. The levels of (D) IL-1β, (E) IL-6
and (F) TNF-α in the culture supernatant were analyzed using ELISA
kits. Data are presented as the mean ± SD (n=3). *P<0.05,
**P<0.01 vs. NS-siRNA + control; #P<0.05,
##P<0.01 vs. NS-siRNA + OGD/R;
&P<0.05, &&P<0.01 vs.
NS-siRNA + TX + OGD/R group. OGD/R, oxygen-glucose deprivation and
reoxygenation; siRNA, small interfering RNA; HIF-1α,
hypoxia-inducible factor-1α; NS, non-specific; TX, troxerutin; IL,
interleukin; TNF-α, tumor necrosis factor-α. |
Discussion
Mortality from MI/R remains high and there is an
unmet need for exploring innovative and effective therapeutic
strategies for MI/R injury (1,2).
Chinese herbal medicines have long been used as alternative
therapies against MI/R injury because of their anti-apoptotic,
anti-oxidative stress, and anti-inflammatory activities (33). Troxerutin, a trihydroxyethylated
derivative of the natural bioflavonoid rutin, elicits a therapeutic
action in cardiovascular disease (6,7).
However, the impacts of troxerutin on MI/R injury remain unclear.
The present study, for the first time to the best of our knowledge,
demonstrated that troxerutin attenuated MI/R injury by suppressing
oxidative stress and inflammation associated with enhancing the
activation of the PI3K/AKT/HIF-1α signaling pathway.
In recent years, emerging evidence has identified
the cardioprotective properties of troxerutin in MI/R (10,31).
Badalzadeh et al (13)
reported that troxerutin attenuates MI/R injury through
pharmacological preconditioning. In addition, preconditioning with
troxerutin significantly reduced myocardial infarct size, the
levels of inflammatory cytokines and inhibited cardiomyocyte
apoptosis, ameliorating MI/R injury (34). Consistent with these previous
studies, the present study revealed that troxerutin increased H9C2
cell viability, reduced LDH release and apoptosis under OGD/R
condition, attenuating OGD/R-induced H9C2 cell injury.
HIF-1α is an important transcription factor that
modulates the cellular response to hypoxia; an increase in the
expression of HIF-1α is one of the first adaptations of the
myocardium to ischemic injury (35,36).
Previous studies confirm that the PI3K/AKT pathway can activate
HIF-1α, thus playing an important role in cell growth and cell
survival (22,30). Research has shown that the
activation of the PI3K/AKT/HIF-1α pathway mediates protection
against lung I/R injury (28), and
the upregulation and activation of the PI3K/AKT/HIF-1α pathway is
involved in the protective effects of microRNAs on cardiomyocytes
(37). Troxerutin preconditioning
has been shown to protect against MI/R injury by inhibiting
apoptosis and activating the PI3K/AKT pathway (34). In present study, It was found that
OGD/R treatment significantly reduced the expression of PI3K, p-AKT
and HIF-1α, indicating that OGD/R led to the inhibition of the
PI3K/AKT/HIF-1α pathway. However, troxerutin pretreatment reversed
these effects. In addition, LY294002, an inhibitor of the PI3K
pathway, attenuated troxerutin-induced upregulation of the
PI3K/AKT/HIF-1α pathway and the protection against OGD/R-induced
H9C2 cell injury. HIF-1α-siRNA transfections partially reversed
troxerutin-induced protection against OGD/R injury. These results
indicated that the PI3K/AKT/HIF-1α signaling pathway mediated the
cardioprotective effect of troxerutin against OGD/R injury.
Oxidative damage plays a causal role in the
pathogenesis of MI/R (17),
oxidative stress induces injury through the accumulation of ROS in
I/R-injured hearts and plays important roles in cardiac apoptosis
(38). Although previous studies
have demonstrated that troxerutin has many different biological
activities, including antioxidant and anti-inflammatory activities,
in diabetic male rats (7),
cerebral ischemia (39) and
cardiac disease in metabolic syndrome patients (40), the effects of troxerutin on
oxidative stress in MI/R injury remain unclear. A number of
enzymes, such as SOD and GSH, provide cellular protection against
damage from oxygen-derived free radicals (41). MDA is the end-product of the
oxygen-derived free radicals and lipid oxidation, which reflects
the damage caused by ROS (42). In
present study, the results indicated that troxerutin pretreatment
eliminated OGD/R-induced oxidative stress as illustrated by the
enhancement of antioxidant systems and the inhibition of oxidative
stress. In addition, HIF-1α-siRNA partially reversed the inhibition
of troxerutin on oxidative stress under OGD/R condition. To the
best of our knowledge, the present study is the first to report
that troxerutin attenuates OGD/R-induced oxidative stress by
promoting the PI3K/AKT/HIF-1α signaling pathway.
Inflammation is an important contributor to MI/R
injury, which is triggered and aggravated during the process of
I/R, resulting in cardiomyocyte death (43). Cardiac inflammation is
characterized by the upregulation of pro-inflammatory cytokines,
including IL-1β, IL-6 and TNF-α, which are major contributors to
MI/R injury (43). Previous
studies revealed that troxerutin can regulate the inflammatory
response in a variety of diseases, such as cerebral I/R injuries
(44), kidney damage (45) and MI/R injury (13). Although, the PI3K/AKT signaling
pathway has been reported to be implicated in human malignancies,
it can suppress inflammation, however, the effects of HIF-1α
signaling on inflammation remain to be explored. The present study
demonstrated that troxerutin pretreatment mitigated OGD/R-induced
inflammatory response as shown by the decreases in the levels of
IL-1β, IL-6 and TNF-α. Furthermore, the present study was the
first, to the best of our knowledge, to find that HIF-1α knockdown
partially reversed the inhibition of troxerutin on inflammation in
OGD/R-treated H9C2 cells. Taken together, these results indicated
that the PI3K/AKT/HIF-1α signaling pathway contributes to the
protection of troxerutin against ODG/R injury by inhibiting the
inflammatory response.
The results of the present study suggested that
HIF-1a knockdown further increased apoptosis, oxidative stress and
inflammation compared with NS-siRNA, and in these cases, although
troxerutin still had a partial protective effect against
MI/R-induced apoptosis, oxidative stress and inflammation in H9C2
cell, the beneficial effects were reduced compared with the
NS-siRNA group. A previous study reported that troxerutin reduced
2,2,4,4-tetrabromodiphenyl ether-induced oxidative stress and
inflammatory injury, which was dependent on increasing nuclear
factor erythroid 2-reated factor 2 (Nrf2) activity and inhibiting
NACHT, LRR and PYD domains-containing protein 3 (NLRP3)
inflammasome signaling in kidney damage (46). ROS- NLRP3 inflammasome activation
and the Nrf2-antioxidant response element both play an important
role in MI/R injury (47,48). In addition, microRNAs was also
reported to be involved in the cardioprotective effects of
troxerutin during I/R injury (31). Hence, these pathways may also
contribute to the antioxidant, anti-inflammatory and
cardioprotective properties of troxerutin in MI/R injury, and
deserves further investigation in future experiments.
In conclusion, troxerutin exerted protective effects
against MI/R injury by inhibiting oxidative stress and
inflammation. The underlying mechanisms for these phenomena
involved the activation of the PI3K/AKT/HIF-1α signaling pathway.
The findings of the present study facilitate finding new molecular
therapeutic targets for MI/R injury and provide a novel insight
into therapeutic development as an adjuvant therapy for MI/R
injury.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZPY participated in designing the research and
writing of the paper. HQY and JL contributed equally to data
analysis. CL, XH and XSS were involved in designing the study,
performing the experiments and revising the paper. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interests.
References
|
1
|
Kalogeris T, Baines CP, Krenz M and
Korthuis RJ: Ischemia/reperfusion. Compr Physiol. 7:113–170. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang CF: Clinical manifestations and basic
mechanisms of myocardial ischemia/reperfusion injury. Ci Ji Yi Xue
Za Zhi. 30:209–215. 2018.PubMed/NCBI
|
|
3
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Donato M, Evelson P and Gelpi RJ:
Protecting the heart from ischemia/reperfusion injury: An update on
remote ischemic preconditioning and postconditioning. Curr Opin
Cardiol. 32:784–790. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hentia C, Rizzato A, Camporesi E, Yang Z,
Muntean DM, Săndesc D and Bosco G: An overview of protective
strategies against ischemia/reperfusion injury: The role of
hyperbaric oxygen preconditioning. Brain Behav. 8:e009592018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang Z, Zhang L, Wang Y, Gao H, Li X,
Huang X and Huang T: Effects of rutin and its derivatives on
citrinin production by Monascus aurantiacus Li AS3.4384 in liquid
fermentation using different types of media. Food Chem.
284:205–212. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zavvari Oskuye Z, Mirzaei Bavil F,
Hamidian GR, Mehri K, Qadiri A, Ahmadi M, Oghbaei H, Vatankhah AM
and Keyhanmanesh R: Troxerutin affects the male fertility in
prepubertal type 1 diabetic male rats. Iran J Basic Med Sci.
22:197–205. 2019.PubMed/NCBI
|
|
8
|
Raja B, Saranya D and Prabhu R: Role of
flavonoid troxerutin on blood pressure, oxidative stress and
regulation of lipid metabolism. Front Biosci (Elite Ed).
11:121–129. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xin X, Zhang M, Li X, Lai F and Zhao G:
Biocatalytic synthesis of acylated derivatives of troxerutin: Their
bioavailability and antioxidant properties in vitro. Microb Cell
Fact. 17:1302018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Najafi M, Noroozi E, Javadi A and
Badalzadeh R: Anti-arrhythmogenic and anti-inflammatory effects of
troxerutin in ischemia/reperfusion injury of diabetic myocardium.
Biomed Pharmacother. 102:385–391. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu Y and Zheng G: Troxerutin protects
against diabetic cardiomyopathy through NFkappaB/AKT/IRS1 in a rat
model of type 2 diabetes. Mol Med Rep. 15:3473–3478. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mokhtari B, Badalzadeh R, Alihemmati A and
Mohammadi M: Phosphorylation of GSK-3β and reduction of apoptosis
as targets of troxerutin effect on reperfusion injury of diabetic
myocardium. Eur J Pharmacol. 765:316–321. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Badalzadeh R, Baradaran B, Alihemmati A,
Yousefi B and Abbaszadeh A: Troxerutin preconditioning and ischemic
postconditioning modulate inflammatory response after myocardial
ischemia/reperfusion injury in rat model. Inflammation. 40:136–143.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gielis JF, Beckers PAJ, Briede JJ, Cos P
and Van Schil PE: Oxidative and nitrosative stress during pulmonary
ischemia-reperfusion injury: From the lab to the OR. Ann Transl
Med. 5:1312017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hernandez-Resendiz S, Chinda K, Ong SB,
Cabrera-Fuentes H, Zazueta C and Hausenloy DJ: The role of redox
dysregulation in the inflammatory response to Acute myocardial
ischaemia-reperfusion injury-adding fuel to the fire. Curr Med
Chem. 25:1275–1293. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cadenas S: ROS and redox signaling in
myocardial ischemia-reperfusion injury and cardioprotection. Free
Radic Biol Med. 117:76–89. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gonzalez-Montero J, Brito R, Gajardo AI
and Rodrigo R: Myocardial reperfusion injury and oxidative stress:
Therapeutic opportunities. World J Cardiol. 10:74–86. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Katsuki S, Matoba T, Koga JI, Nakano K and
Egashira K: Anti-inflammatory nanomedicine for cardiovascular
disease. Front Cardiovasc Med. 4:872017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sinning C, Westermann D and Clemmensen P:
Oxidative stress in ischemia and reperfusion: Current concepts,
novel ideas and future perspectives. Biomark Med. 11:11031–11040.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun XH, Wang X, Zhang Y and Hui J:
Exosomes of bone-marrow stromal cells inhibit cardiomyocyte
apoptosis under ischemic and hypoxic conditions via miR-486-5p
targeting the PTEN/PI3K/AKT signaling pathway. Thromb Res.
177:23–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yao H and Han X and Han X: The
cardioprotection of the insulin-mediated PI3K/Akt/mTOR signaling
pathway. Am J Cardiovasc Drugs. 14:433–442. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Agani F and Jiang BH: Oxygen-independent
regulation of HIF-1: Novel involvement of PI3K/AKT/mTOR pathway in
cancer. Curr Cancer Drug Targets. 13:245–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Brocato J, Chervona Y and Costa M:
Molecular responses to hypoxia-inducible factor 1α and beyond. Mol
Pharmacol. 85:651–657. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jianqiang P, Ping Z, Xinmin F, Zhenhua Y,
Ming Z and Ying G: Expression of hypoxia-inducible factor 1 alpha
ameliorate myocardial ischemia in rat. Biochem Biophys Res Commun.
465:691–695. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li M, Cui Y, He W, Deng X, Wang Y, Cai M,
Wang Y, Pei J, Mei X and Wu P: Effects of triple-mutated
hypoxia-inducible factor-1α on angiogenesis and cardiac function
improvement in rats with myocardial infarction. Cell Physiol
Biochem. 50:2329–2340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Man S, Chai H, Cui J, Yao J, Ma L and Gao
W: Antitumor and anti-metastatic mechanisms of Rhizoma paridis
saponins in Lewis mice. Environ Toxicol. 33:149–155. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Movafagh S, Crook S and Vo K: Regulation
of hypoxia-inducible factor-1a by reactive oxygen species: New
developments in an old debate. J Cell Biochem. 116:696–703. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liang S, Wang Y and Liu Y: Dexmedetomidine
alleviates lung ischemia-reperfusion injury in rats by activating
PI3K/Akt pathway. Eur Rev Med Pharmacol Sci. 23:370–377.
2019.PubMed/NCBI
|
|
29
|
Liang Z, Chi YJ, Lin GQ, Luo SH, Jiang QY
and Chen YK: MiRNA-26a promotes angiogenesis in a rat model of
cerebral infarction via PI3K/AKT and MAPK/ERK pathway. Eur Rev Med
Pharmacol Sci. 22:3485–3492. 2018.PubMed/NCBI
|
|
30
|
Wei J and Wu J, Xu W, Nie H, Zhou R, Wang
R, Liu Y, Tang G and Wu J: Salvianolic acid B inhibits glycolysis
in oral squamous cell carcinoma via targeting PI3K/AKT/HIF-1α
signaling pathway. Cell Death Dis. 9:5992018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shu L, Zhang W, Huang G, Huang C, Zhu X,
Su G and Xu J: Troxerutin attenuates myocardial cell apoptosis
following myocardial ischemia-reperfusion injury through inhibition
of miR-146a-5p expression. J Cell Physiol. 234:9274–9282. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu J, Sui H, Zhao J and Wang Y: Osmotin
protects H9c2 cells from simulated ischemia-reperfusion injury
through AdipoR1/PI3K/AKT signaling pathway. Front Physiol.
8:6112017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Q, Li J, Wang J, Li J, Janicki JS and
Fan D: Effects and mechanisms of chinese herbal medicine in
ameliorating myocardial ischemia-reperfusion injury. Evid Based
Complement Alternat Med. 2013:9256252013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shu L, Zhang W, Huang C, Huang G and Su G:
Troxerutin protects against myocardial ischemia/reperfusion injury
Via Pi3k/Akt pathway in rats. Cell Physiol Biochem. 44:1939–1948.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee SH, Wolf PL, Escudero R, Deutsch R,
Jamieson SW and Thistlethwaite PA: Early expression of angiogenesis
factors in acute myocardial ischemia and infarction. N Engl J Med.
342:626–633. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Semenza GL: HIF-1: Mediator of
physiological and pathophysiological responses to hypoxia. J Appl
Physiol (1985). 88:1474–1480. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun N, Meng F, Xue N, Pang G, Wang Q and
Ma H: Inducible miR-145 expression by HIF-1a protects
cardiomyocytes against apoptosis via regulating SGK1 in simulated
myocardial infarction hypoxic microenvironment. Cardiol J.
25:268–278. 2018.PubMed/NCBI
|
|
39
|
Yu L, Zhang W, Huang C, Liang Q, Bao H,
Gong Z, Xu M, Wang Z, Wen M and Cheng X: FoxO4 promotes myocardial
ischemia-reperfusion injury: The role of oxidative stress-induced
apoptosis. Am J Transl Res. 10:2890–2900. 2018.PubMed/NCBI
|
|
40
|
Ma W, Wang S, Liu X, Tang F, Zhao P, Cheng
K, Zheng Q, Zhuo Y, Zhao X, Li X and Feng W: Protective effect of
troxerutin and cerebroprotein hydrolysate injection on cerebral
ischemia through inhibition of oxidative stress and promotion of
angiogenesis in rats. Mol Med Rep. 19:3148–3158. 2019.PubMed/NCBI
|
|
41
|
Geetha R, Sathiya Priya C and Anuradha CV:
Troxerutin abrogates mitochondrial oxidative stress and myocardial
apoptosis in mice fed calorie-rich diet. Chem Biol Interact.
278:74–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sies H: Oxidative stress: A concept in
redox biology and medicine. Redox Biol. 4:180–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Czerska M, Mikołajewska K, Zieliński M,
Gromadzińska J and Wąsowicz W: Today's oxidative stress markers.
Med Pr. 66:393–405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ong SB, Hernández-Reséndiz S,
Crespo-Avilan GE, Mukhametshina RT, Kwek XY, Cabrera-Fuentes HA and
Hausenloy DJ: Inflammation following acute myocardial infarction:
Multiple players, dynamic roles, and novel therapeutic
opportunities. Pharmacol Ther. 186:73–87. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhào H, Liu Y, Zeng J, Li D, Zhang W and
Huang Y: Troxerutin and cerebroprotein hydrolysate injection
protects neurovascular units from oxygen-glucose deprivation and
reoxygenation-induced injury in vitro. Evid Based Complement
Alternat Med. 2018:98596722018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Shan Q, Zheng GH, Han XR, Wen X, Wang S,
Li MQ, Zhuang J, Zhang ZF, Hu B, Zhang Y and Zheng YL: Troxerutin
protects kidney tissue against BDE-47-induced inflammatory damage
through CXCR4-TXNIP/NLRP3 signaling. Oxid Med Cell Longev.
2018:98654952018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shan Q, Zhuang J, Zheng G, Zhang Z, Zhang
Y, Lu J and Zheng Y: Troxerutin reduces kidney damage against
BDE-47-induced apoptosis via inhibiting NOX2 activity and
increasing Nrf2 activity. Oxid Med Cell Longev. 2017.6034692.
View Article : Google Scholar
|
|
48
|
Qiu Z, Lei S, Zhao B, Wu Y, Su W, Liu M,
Meng Q, Zhou B, Leng Y and Xia ZY: NLRP3 inflammasome
activation-mediated pyroptosis aggravates myocardial
ischemia/reperfusion injury in diabetic rats. Oxid Med Cell Longev.
2017:97432802017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shen Y, Liu X, Shi J and Wu X: Involvement
of Nrf2 in myocardial ischemia and reperfusion injury. Int J Biol
Macromol. 125:496–502. 2019. View Article : Google Scholar : PubMed/NCBI
|