Introduction
Cardiovascular disease is a significant hazard to
human life and health. Research has established that the death of
endothelial cells (ECs), inflammation, endothelial-to-mesenchymal
transition, proliferation and migration, resulting from
hypertension, hyperlipidemia, diabetes and smoking, serve crucial
roles in the development of cardiovascular diseases (1). In addition, numerous studies
(2,3) have shown that changes in the
autophagic activity of ECs have important roles in the pathogenesis
of cardiovascular diseases.
Autophagy is the process of encapsulating proteins
and organelles to be degraded, and transporting them to lysosomes
for degradation. Subsequently, the resulting amino acids, lipids,
nucleic acids and carbohydrates are released into cells to support
metabolic requirements (4). Under
physiological conditions, when the nutrient supply is sufficient,
the level of autophagy is very low, and a low level of basal
autophagy is essential for cell survival; however, if the autophagy
activity is inhibited to zero, the cells will die quickly (5). ECs are an important part of the
vascular wall of the cardiovascular system. According to the time
and degree of EC autophagy activation, the EC autophagy process
serves a dual role in cardiovascular disease (3). Zhou et al (6) reported that resveratrol can protect
human umbilical vein ECs (HUVECs) from palmitic acid-induced
oxidative damage by inducing autophagy, thereby reducing
endothelial oxidative damage in a transcription factor EB-dependent
manner. Peng et al (7)
demonstrated that the traditional Chinese medicine compound
13-methyl berberine (13-MB) may possess anti-atherosclerosis
effects; 13-MB was shown to exert a protective role in a model of
H2O2-induced cell injury by activating
autophagy in HUVECs and inhibiting activation of the NLRP3
inflammasome. Zhang et al (8) reported that knockout of the RAGE gene
reduced myocardial fibrosis by inhibiting excessive
autophagy-mediated transformation of ECs into mesenchymal cells.
Niu et al (9) revealed that
metformin downregulated autophagy through the Hedgehog pathway,
thereby reducing hyperglycemia-induced endothelial injury.
Therefore, in cardiovascular diseases, autophagy may protect cells
and clear damaged cell components; when cells are stimulated by
stressors, such as nutrient deficiency, hypoxia and infection,
autophagy can act to promote cell survival. However, conversely,
excessive activation of autophagy can lead to excessive degradation
of intracellular components, which in turn leads to cell death
(10).
Notably, improving understanding of the effects of
autophagy on the body and its potential for precise regulation is
highly significant in the context of preventing or treating
cardiovascular diseases. However, research has suggested that the
mechanism of autophagy is complex and is regulated by a number of
signaling pathways; among these, the regulation of autophagy by
microRNAs (miRNAs/miRs) has become the focus of extensive research.
miRNAs are small non-coding RNAs that negatively regulate gene
expression by binding to the 3′ untranslated region (3′-UTR) of
different target mRNAs, thereby inducing mRNA degradation or
inhibiting translation (11).
Previous studies (12,13) have identified genetic factors
leading to the onset of cardiovascular disease and have discussed
the role of miRNAs in its pathogenesis. Among these, miR-92a is a
member of the miR-17-92 family that has been reported to be related
to the formation of vascular ECs (14–16).
As a mechanosensitive miRNA closely related to cardiovascular
diseases (17,18), miR-92a serves a significant role in
regulating the dynamic equilibrium of vascular ECs. Several studies
(19,20) have shown that miR-92a is associated
with cardiovascular disease. In addition, clinical trials have
demonstrated that miR-92a levels may be significantly increased in
the serum of patients with cardiovascular diseases (21–25).
Further receiver operating characteristic (ROC) analysis confirmed
that miR-92a alone showed 85% sensitivity and 82.50% specificity
for predicting cardiovascular diseases (26). The upregulation of miR-92a has a
significant role in EC damage by inducing apoptosis, inflammation,
endothelial-mesenchymal transition, oxidative stress, proliferation
and migration; these processes contribute to the onset and
progression of cardiovascular illnesses. Wu et al (17) revealed that the expression of
miR-92a was elevated in the ECs of the aortic arch in mice and pigs
that were subjected to disrupted blood flow; this increase in
miR-92a led to heightened inflammation and permeability in the ECs
by suppressing the expression of phosphatidic acid phosphatase type
2 B. Furthermore, Wiese et al (27) transfected miR-92a into human aortic
ECs (HCAECs) and demonstration that overexpression of miR-92a
directly regulated FAM220A to increase STAT3 phosphorylation,
thereby promoting inflammatory responses to cholesterol and lipid
accumulation in HCAECs. Shang et al (28) found that high expression of miR-92a
promoted EC apoptosis and subsequent vascular inflammation by
inhibiting the expression of the endothelial protective molecules
Krüppel-like factor (KLF)2 and KLF4. However, whether miR-92a
regulates the autophagic activity of EA.hy926 RECs by targeting the
expression of FOXO3 has not been reported.
The present study aimed to examine the expression of
miR-92a in a cell model of autophagy comprising EA.hy926 cells
treated with the autophagy inducer rapamycin (rapa-EA.hy926 cells).
Additionally, the study sought to elucidate the molecular mechanism
via which miR-92a regulates autophagy in EA.hy926 cells.
Materials and methods
Cell culture and establishment of the
autophagy model
EA.hy926 cells (Shanghai Zhongqiao Xinzhou
Biotechnology Co., Ltd.) were cultured in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.)
containing 10% fetal bovine serum (Biological Industries; Sartorius
AG) and 1% streptomycin/penicillin (Beyotime Institute of
Biotechnology) at 37°C in a saturated humidity incubator containing
5% CO2. When cell confluence reached 70–80%, the cells
were starved in serum-free medium for 12 h to synchronize the cell
cycle and were then incubated with rapamycin (cat. no. B20714;
Shanghai Yuanye Biotechnology Co., Ltd.) at a concentration of
1,000 nM in a 5% CO2 cell incubator at 37°C for 6 h to
establish the autophagy model. The cells in the control group did
not receive any treatment.
Western blot analysis
LC3I/LC3II, p62, Beclin1 and FOXO3 expression levels
were determined using western blotting. Total protein was extracted
from EA.hy926 cells using radioimmunoprecipitation assay buffer
(cat. no. P0013B; Beyotime Institute of Biotechnology) and were
quantified using a BCA kit (cat. no. P0009; Beyotime Institute of
Biotechnology). A Bio-Rad iMark microplate reader (Bio-Rad
Laboratories, Inc.) was used to measure the absorbance value of
each sample at a wavelength of 540 nm. The protein concentration
was calculated according to the standard curve, after which, equal
samples of protein (20 µg) were separated by SDS-PAGE on 10 or 20%
gels, and were transferred to a polyvinylidene fluoride membrane.
After being blocked with 5% skim milk for 1 h at room temperature,
the membrane was incubated with rabbit primary antibodies against
LC3I/LC3II (cat. no. WL01506; 16/14 kDa), p62 (cat. no. WL02385; 55
kDa), Beclin1 (cat. no. WL02508; 60 kDa) and FOXO3 (cat. no.
WL02891; 70 kDa) (all from Shenyang Wanlei Biotechnology Co., Ltd.
and diluted to 1:1,000) at 4°C overnight. Subsequently, the
membrane was incubated with a horseradish peroxidase-conjugated
goat anti-rabbit IgG secondary antibody (cat. no. WLA023; 1:5,000,
Shenyang Wanlei Biotechnology Co., Ltd.) at 37°C for 1 h. Target
bands were developed using a chemiluminescence substrate kit (cat.
no. P0018S; Beyotime Institute of Biotechnology) and were analyzed
using Image-Pro Plus software (version 6.0; Media Cybernetics,
Inc.). β-actin (cat. no. WL0002d; 43 kDa; Shenyang Wanlei
Biotechnology Co., Ltd.; diluted to 1:1,000) was used as an
internal reference.
Immunofluorescence assay
Immunofluorescence analysis was performed to assess
the expression of LC3B. The cells were fixed with 4%
paraformaldehyde (cat. no. P0099; Beyotime Institute of
Biotechnology) for 15 min at room temperature and permeabilized
with 0.1% Triton X-100 (cat. no. P0096; Beyotime Institute of
Biotechnology) for 15 min at room temperature. Goat serum (cat. no.
C0265; Beyotime Institute of Biotechnology) was used to block the
sections for 30 min at room temperature. Subsequently, the cells
were incubated with an anti-LC3B rabbit antibody (cat. no. WL01506;
1:200; Shenyang Wanlei Biotechnology Co., Ltd.) at 4°C overnight.
The cells were then incubated with an AlexaFluor®
594-conjugated goat anti-rabbit IgG secondary antibody (cat. no.
RS3611; 1:1,000; ImmunoWay Biotechnology Company) at room
temperature for 1 h. The nuclei were stained at room temperature
with DAPI (1 µg/ml; cat. no. C1006; Beyotime Institute of
Biotechnology) for 10 min. A Leica TCS SP laser scanning confocal
microscope (Leica Microsystem GmbH) was used to obtain micrographs.
The immunofluorescence intensity was measured using Image-ProPlus
6.0 software (Media Cybernetics, Inc.).
Transmission electron microscopy
(TEM)
The treated cells were moved to a 1.5-ml centrifuge
tube using a cell scraper and then centrifuged at 1,006.2 × g for 5
min at room temperature. Subsequently, the supernatant was
discarded, and 1 ml 2.5% glutaraldehyde (Beyotime Institute of
Biotechnology) was added to the sediment for fixation at 4°C for 4
h. The fixed samples were then washed with phosphate-buffered
saline, dehydrated at 4°C with successively increasing percentages
of acetone (50, 70, 90 and 100%; 15 min each) and soaked overnight
at room temperature with Spurr's resin. Subsequently, the samples
were incubated overnight in a 37°C oven and baked for 48 h in a
60°C oven. The samples were then cut into ultrathin slices (60–80
nm) using an ultrathin microtome (Leica Microsystems GmbH), stained
with uranyl acetate for 15 min at room temperature, rinsed with
distilled water, stained with lead citrate for 15 min at room
temperature, rinsed again with distilled water and observed by TEM
(Hitachi, Ltd.) after air-drying overnight.
Reverse transcription-quantitative PCR
(RT-qPCR)
The expression levels of miR-92a and FOXO3 were
measured using RT-qPCR. Briefly, total RNA was isolated from
EA.hy926 cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. RNA was then reverse transcribed into cDNA using the
Evo M-MLV Reverse Transcription Kit (cat. no. AG11705; Accurate
Biotechnology Co., Ltd.) or the miRNA 1st strand cDNA synthesis kit
(cat. no. AG11717; Accurate Biotechnology Co., Ltd.), according to
the manufacturer's protocols, and qPCR reaction was carried out
according to the following amplification conditions:
Pre-denaturation at 94°C for 2 min for one cycle; followed by 35
cycles of denaturation at 94°C for 30 sec, annealing at 55°C for 30
sec and extension at 72°C for 2 min; and a final extension step at
72°C for 6 min. The relative changes in mRNA expression were
calculated using the 2−ΔΔCq formula (29). Among them, the reverse miR-92a
primer was obtained from its RT reagent kit (cat. no. AG11717;
Accurate Biotechnology Co., Ltd.). GAPDH and U6 were used as
internal controls for mRNA and miRNA, respectively, and the primers
for qPCR were synthesized by General Biology (Anhui) Co., Ltd. with
the following sequences: FOXO3, forward 5′-CTCTCTCGCCCATGCTCTAC-3′,
reverse 5′-CCGAGCCCTTGGTGGTATA-3′; miR-92a, forward
5′-TATTGCACTTGTCCCGGCCTG-3′; GAPDH, forward
5′-GGACCTGACCTGCCGTCTAG-3′, reverse 5′-GAGGAGTGGGTGTCGCTGTT-3′; and
U6, forward 5′-CTCGCTTCGGCAGCACA-3′ and reverse
5′-CTCGCTTCACGAATTTGCGT-3′.
Cell transfection
Transfection of miR-92a mimics and inhibitor.
miR-92a mimics, miR-92a inhibitor and their corresponding negative
controls (NCs) were purchased from General Biosystems (Anhui)
Corporation Ltd. Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) was used for transfection according to the
manufacturer's instructions. Briefly, the cells were uniformly
inoculated into 6-well plates (2×105/well) and cultured
at 37°C and 5% CO2 until the cell confluence reached
30–50% after 24 h. A miRNA storage solution was prepared at a final
concentration of 20 µM by dissolving 2.5 µmol freeze-dried miRNA
mimics, inhibitor, mimics NC and inhibitor NC in 125 µl deionized
water. Subsequently, 10 µl miRNA storage solution was mixed with
240 µl serum-free DMEM; the final concentration of miRNA mimics and
inhibitors was 100 nM. The miR-92a mimics, miR-92a inhibitor and
their corresponding NCs were then transfected into cells using
Lipofectamine 2000 at room temperature for 4–6 h. The subsequent
experiments were conducted 48 h post-transfection.
The transfected cells were then cultured in
conventional medium for 48–72 h. The sequences were as follows:
miR-92a mimics, 5′-UAUUGCACUUGUCCCGGCCUGU-3′; miR-92a inhibitor,
5′-ACAGGCCGGGACAAGUGCAAUA-3′; miR-92a mimics NC,
5′-UUCUCCGAACGUGUCACGUTT-3′; miR-92a inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAA-3′.
Small interfering RNA (siRNA)
transfection
FOXO3 siRNA (siFOXO3) and NC siRNA (siFOXO3 NC) were
acquired from General Biosystems (Anhui) Corporation Ltd. The cells
were evenly seeded into 6-well plates (2×105/well) and
incubated at 37°C and 5% CO2 until cell confluence
reached 30–50% after 24 h. The cells were cultured with a mixture
of siRNA and Lipofectamine 2000 in 100 µl serum-free DMEM,
according to the manufacturer's instructions. The medium was
replaced after 4–6 h, and the cells were cultured in a 5%
CO2 incubator at 37°C for 48 h. The sequences were as
follows: si FOXO3, sense 5′-AAAUAGCUACUUACCUUUGCAGU-3′, antisense
5′-ACUGCAAAGGUAAGUAGCUAUUU-3′; and siFOXO3 NC, sense
5′-UGAAUUGUAAUACGACUCACUAU-3′, antisense
5′-AUAGUGAGUCGUAUUACAAUUCA-3′.
Co-transfection of cells with miR-92a
inhibitor + siFOXO3 and miR-92a inhibitor + siFOXO3 NC
A miRNA storage solution was prepared at a final
concentration of 20 µM by dissolving 2.5 µmol freeze-dried miR-92a
inhibitor in 125 µl deionized water. A total of 24 h before
transfection, the cells were plated in 24-well cell culture plates
in complete DMEM (5×104 cells/well). Cells were then
transfected with 100 nM miR-92a inhibitor and 100 nM siFOXO3 or
siFOXO3 NC for 4–6 h at room temperature using Lipofectamine 2000.
Subsequent experiments were performed 72 h post-transfection.
Dual-luciferase reporter assay
The pmirGLO vector was synthesized by General
Biology (Anhui) Co., Ltd. The potential target genes of miR-92a
were predicted using StarBase (http://starbase.sysu.edu.cn/) database, which showed
that the FOXO3 3′-UTR has a miR-92a-binding site. Logarithmic-phase
cells were inoculated into 24-well plates at a density of
2×105 cells/well. Subsequently, the cells were
co-transfected with luciferase reporter vectors (FOXO3-WT,
5′-GTAAATTGTTGTGCAATTGTGG-3′; FOXO3-MUT,
5′-GTAAATTGTTGTATGATTGTGGTTA-3′) and miR-92a mimics or NCs using
Lipofectamine 2000. Luciferase activity was measured 24 h after
transfection using a dual-luciferase reporter kit (Dual Luciferase
Reporter Gene Assay Kit; Beyotime Institute of Biotechnology) and
luciferase activity was normalized to Renilla luciferase
activity.
Statistical analysis
Each experiment was conducted at least three times
and data are presented as the mean ± standard deviation. GraphPad
Prism (version 9.0; Dotmatics) was used for statistical analysis.
Comparisons between two groups were performed using an unpaired
Student's t-test (independent-samples t-test). Comparisons among
multiple groups were performed with one-way ANOVA and Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Rapamycin promotes EA.hy926 cell
autophagy and inhibits the expression of miR-92a
Rapamycin, a strong and effective autophagy inducer,
is known to stimulate ECs to undergo autophagy when incubated at a
concentration of 1,000 nM for 6 h (30). In order to examine the association
between miR-92a and autophagy in the EA.hy926 cell line, the cells
were exposed to rapamycin at a dose of 1,000 nM for a 6 h to create
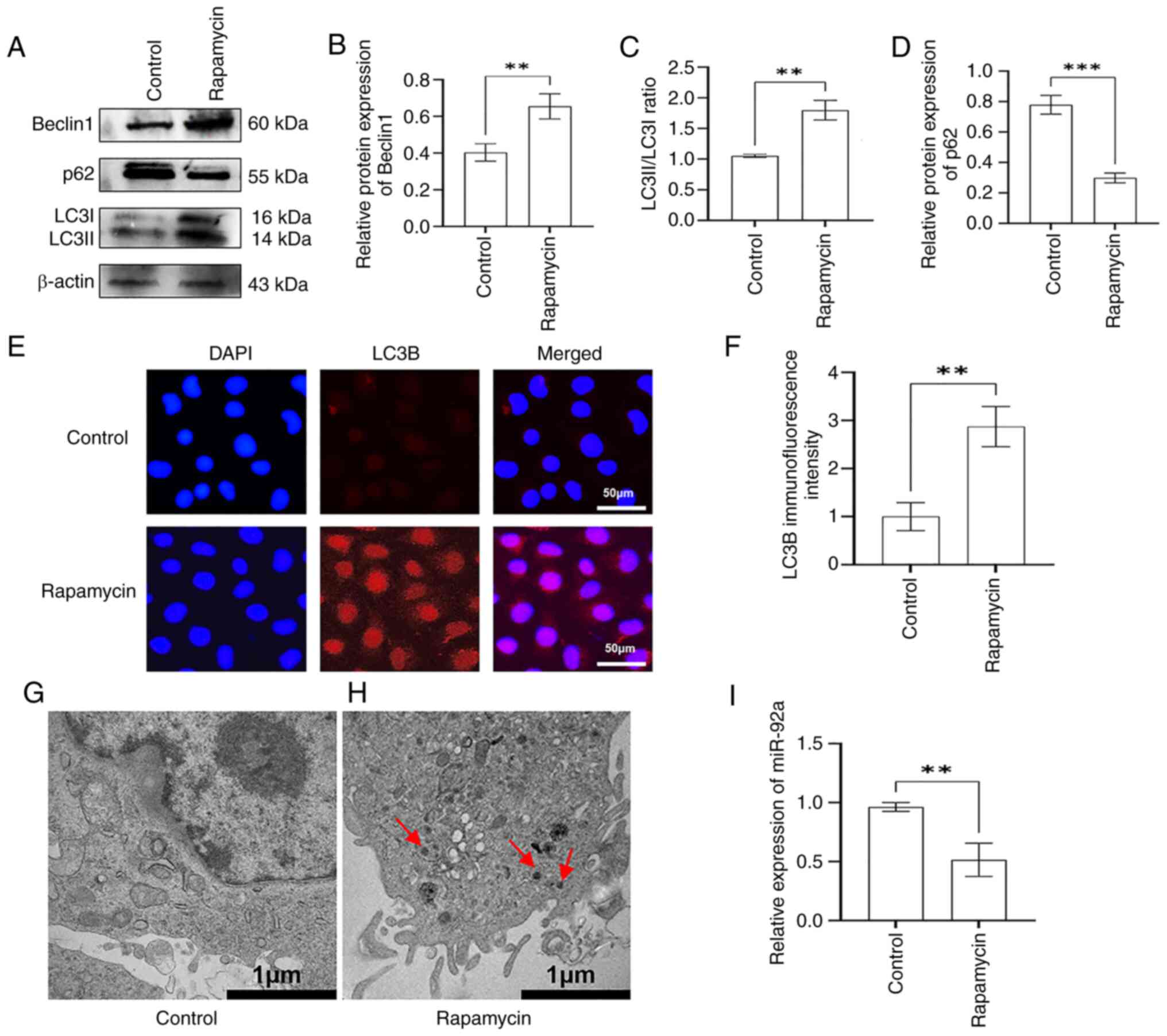
an autophagy model in EA.hy926 cells. The results of western
blotting showed that, compared with in the control group, the
expression levels of autophagy-related Beclin1 and the LC3II/I
ratio were significantly increased in the rapamycin group (Fig. 1A-C), whereas the expression levels
of p62 were decreased (Fig. 1A and
D). The results of immunofluorescence assay showed
significantly increased fluorescence intensity of LC3B in the
rapamycin group compared with that in the control group (Fig. 1E and F). TEM was used to count the
number of autophagosomes in EA.hy926 cells in the control and
rapamycin-treated groups. The results found that no obvious
autophagosomes were present in the cytoplasm of the control group
(Fig. 1G), whereas there was an
increased number of autophagosomes in the cytoplasm of the
rapamycin-treated group (Fig. 1H).
These results indicated that rapamycin activated autophagy in
EA.hy926 cells, and the rapa-EC.hy926 autophagy model was
established successfully.
To assess the association between miR-92a and
autophagy in ECs, RT-qPCR was performed to detect the expression
levels of miR-92a in the rapa-EA.hy926 cell autophagy model. The
findings demonstrated a significant decrease in the expression of
miR-92a in the rapamycin group compared with that in the control
group (Fig. 1I). These findings
indicated that miR-92a was abnormally expressed in the
rapa-EA.hy926 autophagy model, suggesting that miR-92a may be
involved in autophagy in this model system.
miR-92a regulates the autophagy
function of rapa-EA.hy926 cells
To assess the impact of miR-92a on autophagy in
EA.hy926 cells, miR-92a mimics or a miR-92a inhibitor were
introduced into rapa-EA.hy926 cells using Lipofectamine. This
allowed for the increase or decrease in the levels of miR-92a,
hence creating experimental variations in miR-92a expression. The
results of RT-qPCR indicated that the miR-92a mimics group
exhibited a significant increase in the expression levels of
miR-92a, indicating overexpression, compared with those in the
mimics NC control group (Fig. 2A).
Conversely, the miR-92a inhibitor group showed a significant
decrease in the expression level of miR-92a, indicating inhibition.
Western blot analysis demonstrated that, compared with those in the
control group, the expression levels of Beclin1 and the LC3II/LC3I
ratio in the miR-92a mimics group were decreased, and the protein
expression levels of p62 were increased (Fig. 2B-E). By contrast, the expression
levels of Beclin1 and the LC3II/LC3I ratio were higher in the
miR-92a inhibitor group than those in the control group, whereas
the protein expression levels of p62 were decreased. In addition,
immunofluorescence staining results showed that the fluorescence
intensity of LC3B in the miR-92a mimics group was significantly
decreased compared with that in the control group, whereas the
fluorescence intensity of LC3B in the miR-92a inhibitor group was
increased compared with that in the control group (Fig. 2F and G). These findings indicated
that the autophagy marker protein LC3B accumulated in the cytoplasm
of the miR-92a inhibitor group. Furthermore, as number of
autophagosomes were observed by TEM in the cytoplasm of the miR-92a
inhibitor group, whereas no autophagosomes were apparent in the
miR-92a mimics group, and only a small number of autophagosomes
were observed in the miR-92a inhibitor and miR-92a mimic NC groups
(Fig. 2H-K). These results
suggested that overexpression of miR-92a may inhibit autophagy,
whereas inhibition of miR-92a could enhance autophagy in
rapa-EA.hy926 cells.
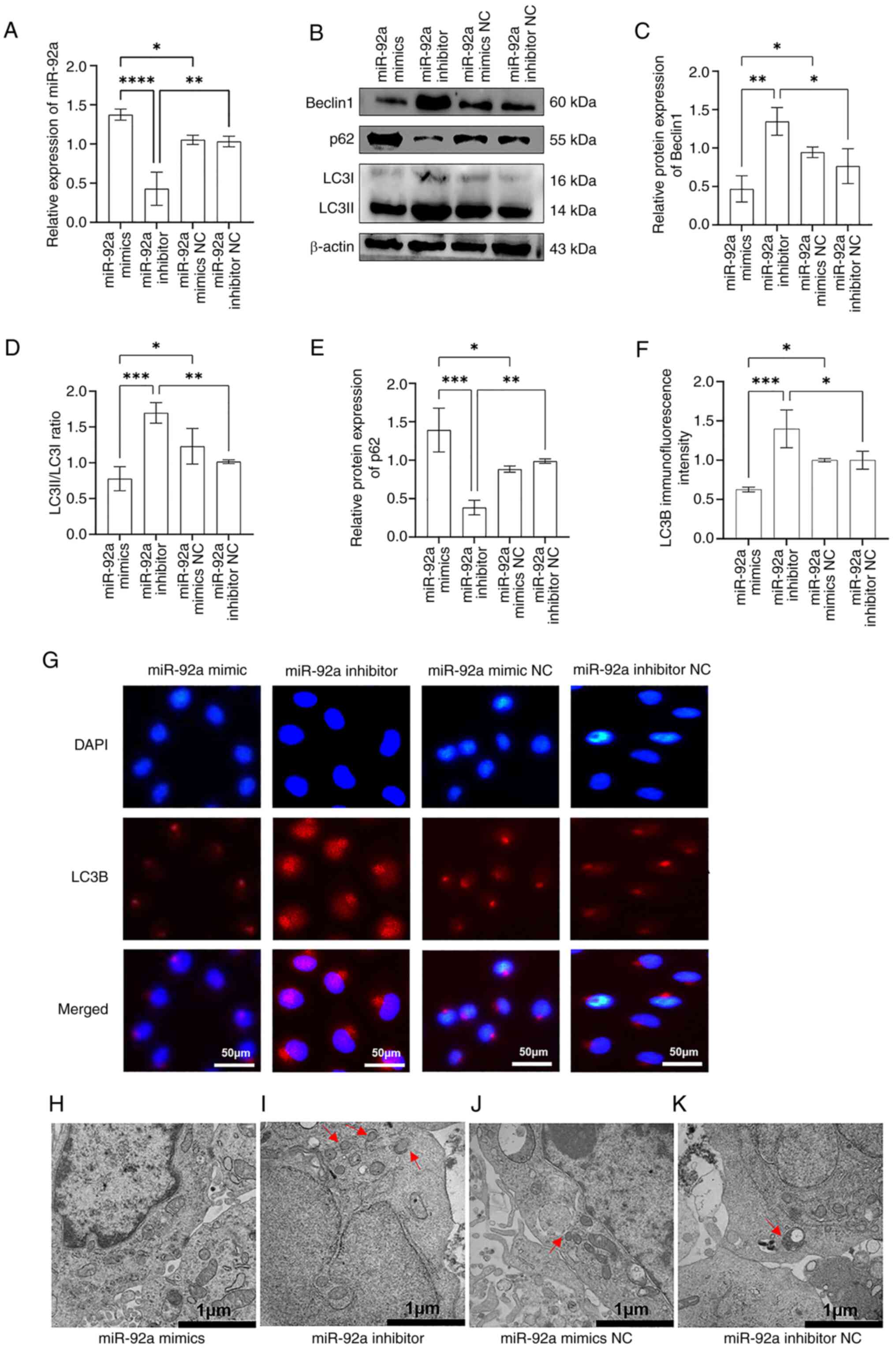 | Figure 2.miR-92a inhibits the autophagic
activity of rapamycin-treated EA.hy926 cells. (A) Reverse
transcription-quantitative PCR results showed that after 48 h of
transfection of rapamycin-treated EA.hy926 cells with miR-92a
mimics, miR-92a inhibitor or corresponding NCs, the expression of
miR-92a was upregulated in the miR-92a mimics group, whereas it was
downregulated in the miR-92a inhibitor group, indicating that
miR-92a mimics or inhibitors were successfully transfected into the
cells. (B) Western blotting was used to detect the expression
levels of autophagy-related proteins Beclin1, p62 and LC3II/I in
the miR-92a mimics, miR-92a inhibitor or corresponding NC groups.
Semi-quantitative analysis of western blotting showed that the (C)
expression of Beclin1 and the (D) LC3II/I ratio were decreased in
the miR-92a mimics group and were increased in the miR-92a
inhibitor group, whereas (E) p62 was increased in the miR-92a
mimics group and decreased in the miR-92a inhibitor group. (F)
Results of LC3B immunofluorescence semi-quantitative analysis
showed that the expression of LC3B in the miR-92a mimics group was
decreased, whereas it was increased in the miR-92a inhibitor group.
(G) Immunofluorescence assay was used to detect the
immunofluorescence intensity of LC3B in the miR-92a mimics, miR-92a
inhibitor or corresponding NC groups. Red, LC3B protein; blue, DAPI
nuclear staining. Results of transmission electron microscopy in
the (H) miR-92a mimics group, (I) miR-92a inhibitor group, (J)
miR-92a mimics NC group and (K) miR-92a inhibitor NC group (red
arrows indicate autophagosomes). *P<0.05, **P<0.01,
***P<0.001, ****P<0.0001. miR, microRNA; NC, negative
control. |
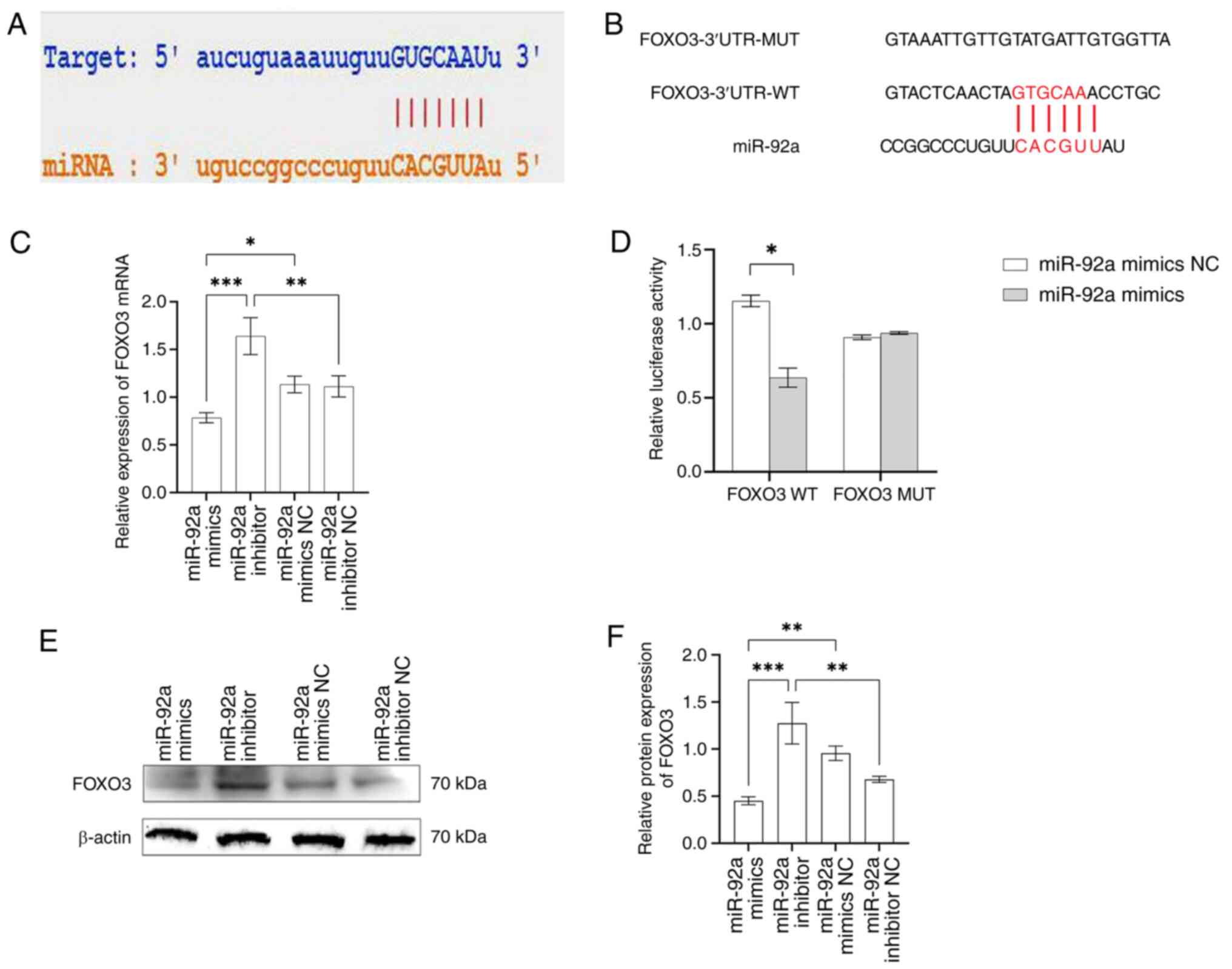
miR-92a directly binds to the FOXO3
3′-UTR region and negatively regulates FOXO3 expression
According to reports (31,32),
miRNAs control several biological processes of cells by suppressing
the transcription or translation of specific target genes at the
post-transcriptional stage. Therefore, whether miR-92a is involved
in the regulation of autophagy in EA.hy926 cells by acting on
downstream target genes was explored. The bioinformatics software
StarBase was used to analyze the downstream target genes of
miR-92a. The results showed a binding site for miR-92a in the
3′-UTR region of FOXO3 (Fig. 3A);
therefore, it was hypothesized that FOXO3 is a target gene for
miR-92a. To assess this, FOXO3 wild-type and mutant luciferase
reporter plasmids were constructed (Fig. 3B). Notably, luciferase activity in
the miR-92a mimics + wild-type FOXO3 plasmid group was
significantly lower than that in the miR-92a mimic NC + wild-type
FOXO3 plasmid group (Fig. 3D).
There was no significant difference in luciferase activity between
the miR-92a mimic + mutant FOXO3 plasmid group and the miR-92a
mimic NC + mutant FOXO3 plasmid group, which further confirmed that
miR-92a directly binds to FOXO3 3′-UTR.
Subsequently, the impact of miR-92a on the control
of FOXO3 gene expression was investigated. Western blotting and
RT-qPCR showed that FOXO3 mRNA and protein expression levels were
inhibited in the miR-92a mimics group compared with those in the
control group, whereas FOXO3 mRNA and protein expression levels
were enhanced in the miR-92a inhibitor group (Fig. 3C, E and F). In addition, FOXO3 mRNA
and protein expression levels were significantly downregulated in
the miR-92a mimics group compared with those in the miR-92a
inhibitor group. These results indicated that the overexpression or
inhibition of miR-92a can affect the expression of FOXO3 mRNA and
protein. In summary, these findings suggested that FOXO3 may be a
direct target of miR-92a in EA.hy926 cells and that miR-92a
directly binds to a specific site on the FOXO3 3′-UTR to regulate
its expression.
FOXO3 gene knockdown inhibits the
autophagic activity of rapa-EA.hy926 cells
The present study demonstrated that miR-92a serves a
role in controlling autophagy in EA.hy926 cells and that FOXO3 is a
gene targeted by miR-92a. In addition, previous research (33) has indicated that FOXO3 has a
significant role as an autophagic protein. Therefore, it was
hypothesized that miR-92a might regulate the autophagy of EA.hy926
cells through FOXO3. To assess this hypothesis, the present study
first investigated whether FOXO3 affected autophagy in EA.hy926
cells. siFOXO3 was transfected into cells using Lipofectamine to
silence FOXO3. RT-qPCR and western blot analysis showed that FOXO3
mRNA and protein expression levels were significantly inhibited in
the in siFOXO3 group of rapa-EA.hy926 cells compared with in the
siFOXO3 NC group (Fig. 4A-C),
indicating that FOXO3 siRNA was successfully transfected into
cells. Western blotting was used to detect the expression levels of
autophagy-related proteins in siFOXO3 cells. The results showed
that, compared with in the siFOXO3 NC group, siFOXO3 transfection
significantly decreased the expression levels of Beclin1 and the
LC3II/I ratio, whereas p62 protein expression levels were increased
(Fig. 4A and D-F). In addition,
the immunofluorescence intensity of LC3B in siFOXO3 cells was
weaker than that in the siFOXO3 NC cells (Fig. 4G and H). Furthermore, TEM revealed
that autophagosomes were readily apparent in the siFOXO3 NC group,
whereas no autophagosome formation was observed in the siFOXO3
group (Fig. 4I and J). Therefore,
it was suggested that FOXO3 gene silencing inhibited autophagy in
rapa-EA.hy926 cells.
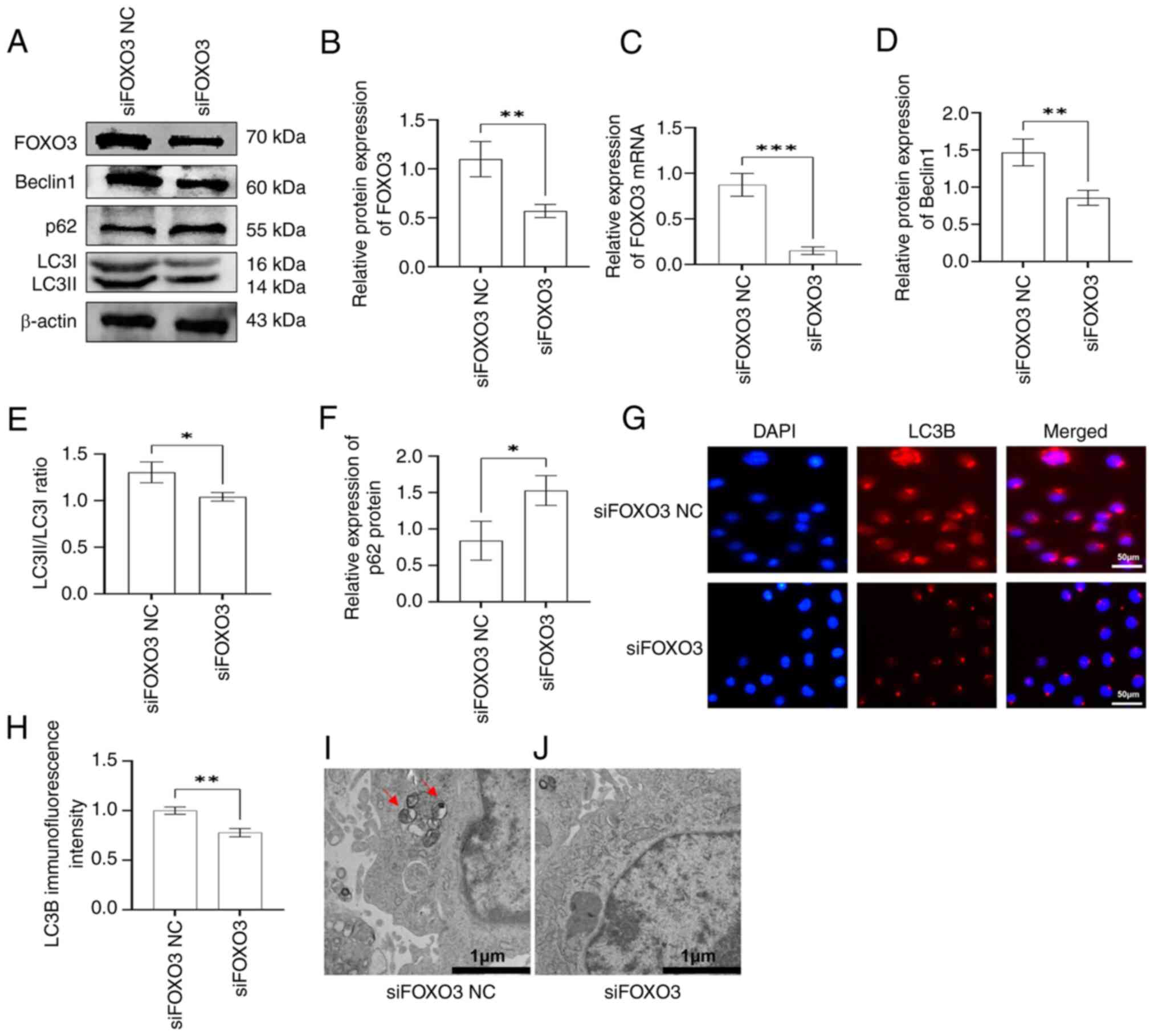 | Figure 4.siFOXO3 inhibits the autophagic
activity of rapa-EA.hy926 cells. (A) Western blotting was used to
detect the expression levels of autophagy-related proteins Beclin1,
p62, LC3II/I and FOXO3 in EA.hy926 cells in the siFOXO3 and siFOXO3
NC groups. (B) Semi-quantitative analysis showed that the
expression of FOXO3 was downregulated in EA.hy926 cells in the
siFOXO3 group, indicating that FOXO3 was successfully inhibited.
(C) Reverse transcription-quantitative PCR results showed that the
mRNA expression levels of FOXO3 were lower in the siFOXO3 group,
indicating that FOXO3 was successfully inhibited. Semi-quantitative
analysis of western blotting showed that (D) Beclin1 expression and
the (E) LC3II/I ratio were lower in the siFOXO3 group, whereas (F)
p62 expression was increased in the siFOXO3 group. (G)
Immunofluorescence assay was used to determine the
immunofluorescence intensity of LC3B in siFOXO3 and siFOXO3 NC
groups. Red, LC3B protein expression; blue, DAPI nuclear staining.
(H) Immunofluorescence semi-quantitative analysis of LC3B showed
that the expression of LC3B was lower in the siFOXO3 group. Results
of transmission electron microscopy showed that there were (I)
obvious autophagosomes in the siFOXO3 NC group and (J) no obvious
autophagosome formation in the siFOXO3 group (red arrows indicate
autophagosomes). *P<0.05, **P<0.01, ***P<0.001. NC,
negative control; si, small interfering. |
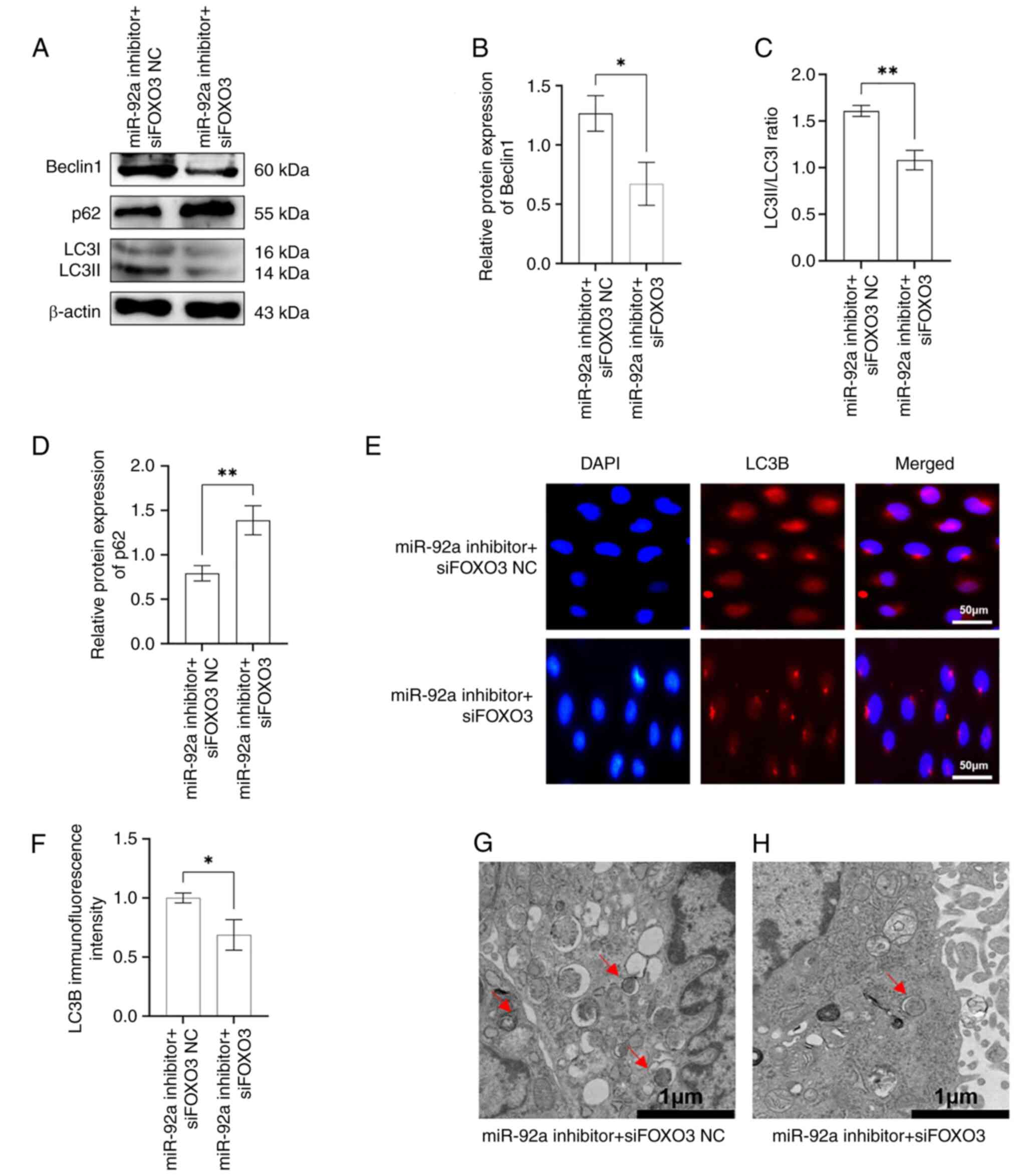
siFOXO3 partially reversed the promoting effect of
miR-92a inhibitors on rapa-EA.hy926 cell autophagy. To evaluate the
role of miR-92a in regulating autophagy in EA.hy926 cells via the
suppression of FOXO3 expression, the rapa-EA.hy926 cells were
transfected with miR-92a inhibitor + siFOXO3 or miR-92a inhibitor +
siFOXO3 NC. Western blotting results showed that the expression
levels of Beclin1 and the LC3II/I ratio in the miR-92a inhibitor +
siFOXO3 group were decreased compared with those in the miR-92a
inhibitor + siFOXO3 NC group, whereas the expression levels of p62
showed an opposite trend (Fig.
5A-D). Immunofluorescence results also showed that the
fluorescence intensity of LC3B in the miR-92a inhibitor + siFOXO3
group was significantly weaker than that in the miR-92a inhibitor +
siFOXO3 NC group (Fig. 5E and F).
Similarly, only a few autophagosomes were detected in the cells of
the miR-92a inhibitor + siFOXO3 group by TEM (Fig. 5 G and H). In conclusion, the
results of siFOXO3 and miR-92a inhibitor co-transfection suggested
that siFOXO3 can partially reverse the promoting effect of the
miR-92a inhibitor on the autophagic activity of rapa-EA.hy926
cells. This further verified that miR-92a regulates autophagy in
EA.hy926 cells by targeting FOXO3.
Discussion
Studies have confirmed that rapamycin is an
autophagy inducer, and it is commonly used in the construction of
various cell models of autophagy (34–36).
To assess the association between miR-92a and autophagy in ECs, the
stable human endothelial cell line EA.hy926 was subjected to
treatment with rapamycin (1,000 nM) for 6 h to develop the
rapa-EA.hy926 cell autophagy model. Rapa-EA.hy926 cells had
increased Beclin1 protein expression, an increased LC3II/I ratio,
decreased p62 expression, significantly increased LC3B
immunofluorescence intensity, and increased formation of
autophagosomes, as detected by TEM, compared with the control
cells, suggesting that the rapa-EA.hy926 cell autophagy model was
successfully constructed. This result is consistent with a previous
report that ECs incubated with 1,000 nM rapamycin for 6 h exhibited
a readily apparent increase in autophagy (30). The results of RT-qPCR showed
significant downregulation of miR-92a in the EA.hy926 cell
autophagy model, suggesting a role for miR-92a in the regulation of
EA.hy926 cell autophagy. To further determine the effects of
miR-92a overexpression and knockdown on the autophagic activity of
EA.hy926 cells, rapa-EA.hy926 cells were transfected with miR-92a
mimics or a miR-92a inhibitor. The results indicated that miR-92a
overexpression could inhibit the autophagic activity of
rapa-EA.hy926 cells, whereas miR-92a knockdown enhanced the
autophagic activity of rapa-EA.hy926 cells. These experimental
results suggested that inhibition of EC autophagy by miR-92a may be
another miR-92a-related cause of endothelial dysfunction. However,
the mechanism by which miR-92a inhibits the autophagic activity of
EC has not yet been reported.
Research (33) has
demonstrated that miRNAs attach themselves to the 3′-UTR region of
downstream target genes to control and influence cellular
functions. In addition, has been suggested that miRNAs have a novel
function in controlling EC autophagy (37). For example, miR-103 has been shown
to protect coronary ECs from H2O2-induced
oxidative stress damage through BNIP3-mediated end-stage autophagy
(38), whereas miR-130a targets
the expression of Runx3 to maintain the normal autophagy levels of
endothelial progenitor cells and promote their survival (39). Lu et al (40) proposed that propofol treatment
could induce the expression of miR-30b, thereby alleviating hypoxia
and reoxygenation-induced HUVEC damage, and upregulated miR-30b was
shown to regulate HUVEC autophagic activity by targeting the
expression of Beclin1.
To further analyze the mechanism by which miR-92a
inhibits EC autophagy, the present study used bioinformatics
software to screen and predict the target genes of miR-92a, and
revealed that FOXO3 was a potential downstream target gene of
miR-92a, which was confirmed by subsequent dual-luciferase
experiments. Western blotting and RT-qPCR analysis showed that
miR-92a mimics suppressed FOXO3 mRNA and protein expression,
whereas the miR-92a inhibitor promoted FOXO3 mRNA and protein
expression. Therefore, these results confirmed FOXO3 to be a target
gene of miR-92a, and miR-92a may inhibit FOXO3 protein and mRNA
expression by binding to its 3′-UTR region.
FOXO3 is a transcription factor that has a role in
regulating several cellular activities. Previous studies have has
demonstrated that FOXO3 functions as a tumor suppressor by
controlling the expression of genes involved in apoptosis, cell
cycle arrest and resistance to oxidative stress (41,42).
In addition, numerous studies have shown that FOXO3 is associated
with longevity and autophagy by directly activating the expression
of autophagy-related proteins, thereby complementing core
components of autophagy (43–46).
In addition, it has been reported that FOXO3 is negatively
regulated by miRNAs and participates in the process of autophagy.
Li et al (47) reported
that the expression of miR-221 was increased and the levels of
FOXO3 were decreased in a rat model of myocardial infarction after
myocardial fibrosis. By contrast, inhibition of miR-221 expression
could inhibit the autophagic activity of cardiomyocytes, and the
results of a luciferase reporter gene assay confirmed that FOXO3
was its downstream target gene. In addition, Song et al
(48) proposed that miR-34a can
inhibit autophagy of alveolar type II epithelial cells in acute
lung injury by inhibiting FOXO3 expression, and FOXO3 was
identified as a downstream target gene of miR-34a. Long et
al (49) showed that
miR-223-3p can regulate autophagy by acting on its target gene
FOXO3, thus participating in the osteogenic differentiation of bone
marrow mesenchymal stem cells. In summary, FOXO3 is known to serve
a significant role in the process of autophagy as a target gene of
miRNAs.
The present study validated that the excessive
production of miR-92a can impede the process of autophagy in
EA.hy926 cells and FOXO3 was identified as a target gene of
miR-92a. Therefore, it was hypothesized that miR-92a may regulate
the autophagic function of EA.hy926 cells by regulating the
expression of FOXO3. To further verify this hypothesis, the effect
of FOXO3 on the autophagy of EA.hy926 cells was studied. siFOXO3
was transfected into rapa-EA.hy926 cells to silence the FOXO3 gene,
and the results showed that siFOXO3 significantly inhibited the
autophagic activity of rapa-EA.hy926 cells. In addition, the
miR-92a inhibitor and siFOXO3 were co-transfected into
rapa-EA.hy926 cells to observe whether FOXO3 silencing could
reverse the effects of the miR-92a inhibitor on autophagy in
rapa-EA.hy926 cells. The results showed that knockdown of FOXO3
could reverse the promoting effect of the miR-92a inhibitor on the
autophagic activity of rapa-EA.hy926 cells, further confirming that
miR-92a may regulate the autophagic activity of EA.hy926 cells
through targeted inhibition of FOXO3 expression.
The present study provided initial evidence to
suggest that miR-92a may suppress the autophagic activity of
EA.hy926 cells by inhibiting the expression of FOXO3. Therefore, it
may be hypothesized that miR-92a not only induces EC apoptosis and
inflammation (48,50) but also inhibits the autophagic
activity of ECs by inhibiting FOXO3 expression, resulting in EC
injury. Because of the significant role of impaired EC autophagic
activity in the pathological processes of cardiovascular disease,
miR-92a inhibitors may be used as EC protective factors that could
provide a novel therapeutic approach for cardiovascular disease;
however, one miRNA can regulate multiple target genes, and the
signaling pathways of miRNAs regulating autophagy are extremely
complex. Therefore, whether miR-92a also regulates EC autophagy by
regulating the expression of other target genes or signaling
pathways needs further study. In addition, the rapamycin-treated
EA.hy926 cell line was an effective research model for in
vitro experiments; however, it is necessary to perform
experiments on animals to verify whether miR-92a influences
autophagic activity via an identical mechanism in living organisms.
With the in-depth study of miR-92a in cardiovascular diseases, the
relationship between cardiovascular diseases and miR-92a will
become clearer, a development that may provide innovative ideas for
the clinical diagnosis and treatment of various cardiovascular
diseases.
Acknowledgements
Not applicable.
Funding
This study was supported by the Natural Science Foundation of
Heilongjiang Province (grant no. LH2022H028).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
LH contributed to the experimental design, revised
the paper, and guided and supervised the entire experimental
process. WC performed the experiments and authored the paper. BZ,
LG, XS and ZZ participated in the data analysis, and the literature
review and collation. LH and BZ confirm the authenticity of all the
raw data. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Little PJ, Askew CD, Xu S and Kamato D:
Endothelial dysfunction and cardiovascular disease: History and
analysis of the clinical utility of the relationship. Biomedicines.
96:6992021. View Article : Google Scholar
|
|
2
|
Lin X, Ouyang S, Zhi C, Li P, Tan X, Ma W,
Yu J, Peng T, Chen X, Li L and Xie W: Focus on ferroptosis,
pyroptosis, apoptosis and autophagy of vascular endothelial cells
to the strategic targets for the treatment of atherosclerosis. Arch
Biochem Biophys. 715:1090982022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao F, Satyanarayana G, Zhang Z, Zhao J,
Ma XL and Wang Y: Endothelial autophagy in coronary microvascular
dysfunction and cardiovascular disease. Cells. 11:20812022.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mameli E, Martello A and Caporali A:
Autophagy at the interface of endothelial cell homeostasis and
vascular disease. FEBS J. 28911:2976–2991. 2021.
|
|
5
|
Li A, Gao M, Liu B, Qin Y, Chen L, Liu H,
Wu H and Gong G: Mitochondrial autophagy: Molecular mechanisms and
implications for cardiovascular disease. Cell Death Dis.
135:4442022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou X, Yang J, Zhou M, Zhang Y, Liu Y,
Hou P, Zeng X, Yi L and Mi M: Resveratrol attenuates endothelial
oxidative injury by inducing autophagy via the activation of
transcription factor EB. Nutr Metab (Lond). 16:422019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Peng Z, Zhan H, Shao Y, Xiong Y, Zeng L,
Zhang C, Liu Z, Huang Z, Su H and Yang Z: 13-methylberberine
improves endothelial dysfunction by inhibiting NLRP3 inflammasome
activation via autophagy induction in human umbilical vein
endothelial cells. Chin Med. 15:82020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang L, He J, Wang J, Liu J, Chen Z, Deng
B, Wei L, Wu H, Liang B, Li H, et al: Knockout RAGE alleviates
cardiac fibrosis through repressing endothelial-to-mesenchymal
transition (EndMT) mediated by autophagy. Cell Death Dis.
12:4702021. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Niu C, Chen Z, Kim KT, Sun J, Xue M, Chen
G, Li S, Shen Y, Zhu Z, Wang X, et al: Metformin alleviates
hyperglycemia-induced endothelial impairment by downregulating
autophagy via the Hedgehog pathway. Autophagy. 15:843–870. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ren H, Dai R, Nik Nabi WN, Xi Z, Wang F
and Xu H: Unveiling the dual role of autophagy in vascular
remodelling and its related diseases. Biomed Pharmacother.
168:1156432023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao S, Wang H, Xu H, Tan Y, Zhang C, Zeng
Q, Liu L and Qu S: Targeting the microRNAs in exosome: A potential
therapeutic strategy for alleviation of diabetes-related
cardiovascular complication. Pharmacol Res. 173:1058682021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wronska A: The role of microRNA in the
development, diagnosis, and treatment of cardiovascular disease:
Recent developments. J Pharmacol Exp Ther. 3841:123–132. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wojciechowska A, Osiak A and
Kozar-Kamińska K: MicroRNA in cardiovascular biology and disease.
Adv Clin Exp Med. 26:868–874. 2017. View Article : Google Scholar
|
|
14
|
Doebele C, Bonauer A, Fischer A, Scholz A,
Reiss Y, Urbich C, Hofmann WK, Zeiher AM and Dimmeler S: Members of
the microRNA-17-92 cluster exhibit a cell-intrinsic antiangiogenic
function in endothelial cells. Blood. 115:4944–4950. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bonauer A, Carmona G, Iwasaki M, Mione M,
Koyanagi M, Fischer A, Burchfield J, Fox H, Doebele C, Ohtani K, et
al: MicroRNA-92a controls angiogenesis and functional recovery of
ischemic tissues in mice. Science. 324:1710–1713. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li M, Guan X, Sun Y, Mi J, Shu X, Liu F
and Li C: miR-92a family and their target genes in tumorigenesis
and metastasis. Exp Cell Res. 323:1–6. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu C, Huang RT, Kuo CH, Kumar S, Kim CW,
Lin YC, Chen YJ, Birukova A, Birukov KG, Dulin NO, et al:
Mechanosensitive PPAP2B regulates endothelial responses to
atherorelevant hemodynamic forces. Circ Res. 174:e41–e53.
2015.PubMed/NCBI
|
|
18
|
Kumar S, Kim CW, Simmons RD and Jo H: Role
of flow-sensitive microRNAs in endothelial dysfunction and
atherosclerosis: Mechanosensitive athero-miRs. Arterioscler Thromb
Vasc Biol. 34:2206–2216. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Y, Cheng J, Chen F, Wu C, Zhang J,
Ren X, Pan Y, Nie B, Li Q and Li Y: Circulating endothelial
microparticles and miR-92a in acute myocardial infarction. Biosci
Rep. 37:BSR201700472017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Carena MC, Badi I, Polkinghorne M,
Akoumianakis I, Psarros C, Wahome E, Kotanidis CP, Akawi N,
ntonopoulos AS, Chauhan J, et al: Role of human epicardial adipose
tissue-derived miR-92a-3p in myocardial redox state. J Am Coll
Cardiol. 82:317–332. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Y, Li Q, Hosen MR, Zietzer A, Flender
A, Levermann P, Schmitz T, Fruhwald D, Goody P, Nickenig G, et al:
Atherosclerotic conditions promote the packaging of functional
MicroRNA-92a-3p into endothelial microvesicles. Circ Res.
124:575–587. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Parahuleva MS, Lipps C, Parviz B,
Holschermann H, Schieffer B, Schulz R and Euler G: MicroRNA
expression profile of human advanced coronary atherosclerotic
plaques. Sci Rep. 8:78232018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang H, Xie Y, Salvador AM, Zhang Z, Chen
K, Li G and Xiao J: Exosomes: Multifaceted messengers in
atherosclerosis. Curr Atheroscler Rep. 22:572020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park CS, Kim I, Oh GC, Han JK, Yang HM,
Park KW, Cho HJ, Kang HJ, Koo BK, Chung WY, et al: Diagnostic
utility and pathogenic role of circulating MicroRNAs in vasospastic
angina. J Clin Med. 9:13132020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alexandru N, Constantin A, Nemecz M,
Comarita IK, Vilcu A, Procopciuc A, Tanko G and Georgescu A:
Hypertension associated with hyperlipidemia induced different
MicroRNA expression profiles in plasma, platelets, and
Platelet-derived microvesicles; Effects of endothelial progenitor
cell therapy. Front Med (Lausanne). 6:2802019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang W, Li Z, Zheng Y, Yan M, Cui Y and
Jiang J: Circulating microRNA-92a level predicts acute coronary
syndrome in diabetic patients with coronary heart disease. Lipids
Health Dis. 18:222019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wiese CB, Zhong J, Xu ZQ, Zhang Y, Ramirez
Solano MA, Zhu W, Linton MF, Sheng Q, Kon V and Vickers KC: Dual
inhibition of endothelial miR-92a-3p and miR-489-3p reduces renal
injury-associated atherosclerosis. Atherosclerosis. 282:121–131.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shang F, Wang SC, Hsu CY, Miao Y, Martin
M, Yin Y, Wu CC, Wang YT, Wu G, Chien S, et al: MicroRNA-92a
Mediates endothelial dysfunction in CKD. J Am Soc Nephrol.
2811:3251–3261. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 254:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang P, Luo XL, Mo GJ, Tao XJ, Shen F and
Ou HS: The effects of miR-24 on proliferation, metastasis, and
autophagy of human umbilical vein endothelial cells. Shandong Med
J. 57:24–27. 2017.
|
|
31
|
Feng Y, Yang H, Yue Y and Tian F:
MicroRNAs and target genes in epileptogenesis. Epilepsia.
61:2086–2096. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Seok H, Ham J, Jang ES and Chi SW:
MicroRNA target recognition: Insights from Transcriptome-Wide
Non-canonical interactions. Mol Cells. 39:375–381. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Long J, Wang J, Dong Y, Yang J, Xie G and
Tong Y: Prolyl isomerase Pin1 promotes autophagy and cancer cell
viability through activating FoxO3 signalling. Cell Signal.
113:1109402024. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liang L, Hui K, Hu C, Wen Y, Yang S, Zhu
P, Wang L, Xia Y, Qiao Y, Sun W, et al: Autophagy inhibition
potentiates the anti-angiogenic property of multikinase inhibitor
anlotinib through JAK2/STAT3/VEGFA signaling in non-small cell lung
cancer cells. J Exp Clin Cancer Res. 381:712019. View Article : Google Scholar
|
|
35
|
Yang Z, Huang C, Wen X, Liu W, Huang X, Li
Y, Zang J, Weng Z, Lu D, Tsang CK, et al: Circular RNA circ-FoxO3
attenuates blood-brain barrier damage by inducing autophagy during
ischemia/reperfusion. Mol Ther. 303:1275–1287. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang S, Tian W, Duan X, Zhang Q, Cao L,
Liu C, Li G, Wang Z, Zhang J, Li J, et al: Melatonin attenuates
diabetic cardiomyopathy by increasing autophagy of cardiomyocytes
via regulation of VEGF-B/GRP78/PERK signaling pathway. Cardiovasc
Diabetol. 231:192024. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou C, Shen L, Mao L, Wang B, Li Y and Yu
H: miR-92a is upregulated in cervical cancer and promotes cell
proliferation and invasion by targeting FBXW7. Biochem Biophys Res
Commun. 4581:63–69. 2015. View Article : Google Scholar
|
|
38
|
Wang Y, Song X, Li Z, Liu N, Yan Y, Li T,
Sun W, Guan Y, Li M, Yang Y, et al: MicroRNA-103 protects coronary
artery endothelial cells against H2O2-Induced
oxidative stress via BNIP3-mediated End-stage autophagy and
antipyroptosis pathways. Oxid Med Cell Longev.
2020:83513422020.PubMed/NCBI
|
|
39
|
Xu Q, Meng S, Liu B, Li MQ, Li Y, Fang L
and Li YG: MicroRNA-130a regulates autophagy of endothelial
progenitor cells through Runx3. Clin Exp Pharmacol Physiol.
41:351–357. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lu Z, Shen J, Chen X, Ruan Z, Cai W, Cai
S, Li M, Yang Y, Mo J, Mo G, et al: Propofol upregulates
MicroRNA-30b to inhibit excessive autophagy and apoptosis and
attenuates Ischemia/Reperfusion injury in vitro and in patients.
Oxid Med Cell Longev. 2022:21098912022. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu D, Liang M, Dang H, Fang F, Xu F and
Liu C: Hydrogen protects against hyperoxia-induced apoptosis in
type II alveolar epithelial cells via activation of PI3K/Akt/Foxo3a
signaling pathway. Biochem Biophys Res Commun. 4952:1620–1627.
2018. View Article : Google Scholar
|
|
42
|
Shi XY, Ding W, Li TQ, Zhang YX and Zhao
SC: Histone deacetylase (HDAC) inhibitor, suberoylanilide
hydroxamic Acid (SAHA), induces apoptosis in prostate cancer cell
lines via the Akt/FOXO3a signaling pathway. Med Sci Monit.
23:5793–5802. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hao W, Dian M, Zhou Y, Zhong Q, Pang W, Li
Z, Zhao Y, Ma J, Lin X, Luo R, et al: Autophagy induction promoted
by m6A reader YTHDF3 through translation upregulation of FOXO3
mRNA. Nat Commun. 13:58452022. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao C, Li X, Sun G, Liu P, Kong K, Chen
X, Yang F and Wang X: CircFOXO3 protects against osteoarthritis by
targeting its parental gene FOXO3 and activating PI3K/AKT-mediated
autophagy. Cell Death Dis. 13:9322022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nho RS and Hergert P: FoxO3a and disease
progression. World J Biol Chem. 5:346–354. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sun L, Zhao M, Wang Y, Liu A, Lv M, Li Y,
Yang X and Wu Z: Neuroprotective effects of miR-27a against
traumatic brain injury via suppressing FoxO3a-mediated neuronal
autophagy. Biochem Biophys Res Commun. 482:1141–1147. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li F, Long TY, Bi SS, Sheikh SA and Zhang
CL: circPAN3 exerts a profibrotic role via sponging miR-221 through
FoxO3/ATG7-activated autophagy in a rat model of myocardial
infarction. Life Sci. 257:1180152020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Song L, Zhou F, Cheng L, Hu M, He Y, Zhang
B, Liao D and Xu Z: MicroRNA-34a suppresses autophagy in alveolar
type II epithelial cells in acute lung injury by inhibiting FoxO3
expression. Inflammation. 40:927–936. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Long C, Cen S, Zhong Z, Zhou C and Zhong
G: FOXO3 is targeted by miR-223-3p and promotes osteogenic
differentiation of bone marrow mesenchymal stem cells by enhancing
autophagy. Hum Cell. 34:14–27. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhou Y, Wei W, Shen J, Lu L, Lu T, Wang H
and Xue X: Alisol A 24-acetate protects oxygen-glucose
deprivation-induced brain microvascular endothelial cells against
apoptosis through miR-92a-3p inhibition by targeting the B-cell
lymphoma-2 gene. Pharm Biol. 59:513–524. 2021. View Article : Google Scholar : PubMed/NCBI
|