Introduction
Acute myocardial infarction (AMI), also known as a
heart attack, is a major global cause of mortality, which is
frequently linked to conditions such as high cholesterol levels,
high blood pressure, smoking, diabetes, obesity, lack of physical
activity, metabolic syndrome and depression (1). In the case of AMI, it is crucial to
restore blood flow promptly to prevent loss of heart function,
decreasing the size of the damaged area and improving overall
clinical prognosis. However, reperfusion, which occurs when the
blood flow is restored, can actually worsen the condition and cause
additional damage to the heart; this phenomenon is known as
myocardial ischemia-reperfusion (MI/R) injury (2). Common techniques for reconstructing
coronary arteries in AMI cases include percutaneous coronary
intervention and surgical coronary artery bypass grafting. However,
these methods do not offer significant benefits in terms of
reducing reperfusion injury. Currently, standard medications, such
as β-blockers, angiotensin-converting enzyme inhibitors and
statins, are administered post-reperfusion (3). Nevertheless, these drugs do not
effectively mitigate myocardial injury by addressing the persistent
risk factors that induce myocardial ischemia and the activation of
inflammatory signals after ischemia. Researchers have recently
focused on investigating drugs that can alleviate the inflammatory
cascade in MI/R injury (4–7). However, these studies often use young
animals as subjects, which does not accurately reflect the
situation of MI/R injury in older individuals with comorbidities
such as diabetes, hypertension and metabolic syndrome (8). Therefore, the identification of more
effective strategies to control MI/R injury remains a challenging
and urgent task.
Platelets serve a crucial role in the entire process
of ischemic heart disease, starting from the occurrence of ischemia
to reperfusion (9). Physiological
platelet activators (thrombin and thromboxin-A2) and external
reactive oxygen species (ROS) promote the activation of platelets
through extracellular signal-regulated kinase 5 after cardiac
ischemia (10). Additionally,
platelets release chemokines that stimulate the migration of
leukocytes to the injury site within blood vessels, thereby
exacerbating inflammatory damage to the myocardium. Presently,
although the use of antiplatelet medications has become a vital
approach to prevent and manage coronary artery disease (CAD), the
mortality rate remains high (11,12),
which may be due to the fact that antiplatelet drugs cannot
directly eliminate the induction factors affecting platelet
hyperreactivity. Therefore, it is necessary to further investigate
the regulatory mechanisms that affect the enhancement of platelet
reactivity, which may result in the development of more efficient
antiplatelet drugs capable of achieving a balance between the
antithrombotic effect and other complications, including excessive
bleeding.
Lipocalin 2 (LCN2), originally considered to be a
25-kDa glycoprotein isolated from human neutrophilic granulocytes,
is expressed in various tissues, including the liver, lung and
kidney (13,14). LCN2 has also been detected in human
atherosclerotic tissues, and it has the ability to upregulate the
production of interleukin (IL)-6, IL-8 and MCP-1 in macrophages,
endothelial cells and smooth muscle cells (15). In addition, LCN2 may have a
significant impact on the development of metabolic syndrome
(16). Notably, obese patients
have been shown to exhibit increased mRNA and protein expression
levels of LCN2 in their visceral adipose tissue compared with in
lean subjects (17). Moreover,
serum levels of LCN2 have been reported to be positively associated
with obesity, hyperglycemia, hypertriglyceridemia and insulin
resistance index (18–20), all of which are crucial risk
factors for ischemic cardiomyopathy. Numerous studies have
confirmed the crucial involvement of LCN2 in cardiovascular
diseases. For example, the knockout of LCN2 in mice with myocardial
ischemia has been shown to lead to a reduction in cardiac
dysfunction (20). Nonetheless, it
remains uncertain whether LCN2 directly affects platelet activation
to regulate the occurrence of MI/R injury.
The present study aimed to investigate the potential
exacerbating effect of high-fat diet (HFD)-induced LCN2 on MI/R
injury through its impact on platelet function. To verify the
hypothesis, a series of experiments were carried out. Firstly, the
expression patterns of LCN2 in the liver and serum of mice
subjected to a HFD were assessed by comparing them to mice not
exposed to a HFD. Secondly, the effects of LCN2 on various aspects
of platelet activity, including activation, aggregation, adhesion
and plaque contraction, were examined. Finally, a MI/R injury model
were established in both wild-type (WT) and LCN2 knockout
(LCN2−/−) mice after HFD feeding, in order to assess the
extent of MI and evaluate the recruitment of platelets,
neutrophils, T cells and B cells in the heart using
immunohistochemistry to detect CD42b, Ly6G, CD3 and B220
expression.
Materials and methods
Establishing a mouse model of
obesity
The animal experimental center at Guizhou Medical
University (Guiyang, China) was responsible for raising mice under
controlled conditions, namely a constant temperature of 22–24°C, a
relative humidity of 55–60% and a 12-h light/dark cycle. The
experimental procedures were approved by the animal experiment
ethics committee of Guizhou Medical University (approval no.
2100328). To determine the effect of a HFD on LCN2 expression, male
C57BL/6J WT mice (Cyagen Biosciences Inc.; weight, 18–20 g; age, 6
weeks) were fed either a HFD (60% kcal from fat; D12492 formula;
Research Diets, Inc.) or normal chow diet for 24 weeks, and then
blood and liver samples were collected to detect LCN2 expression.
These mice (n=5/group) were given free access to food/water. To
investigate the effect of LCN2 gene knockout on MI/R injury in
HFD-fed mice, male WT and LCN2−/− mice (Cyagen
Biosciences Inc.; weight, 18–20 g; age, 6 weeks) were fed a HFD for
24 weeks, and then underwent MI/R injury or sham surgery. The mice
were divided into the following four groups: WT + HFD + sham, WT +
HFD + I/R, LCN2−/− + HFD + sham and LCN2−/− +
HFD + I/R. The hearts (n=5/group) collected from the four groups
underwent hematoxylin and eosin (H&E) staining. The degree of
MI in the hearts of the four groups was assessed through TTC
staining (n=5/group). Immunohistochemistry was performed to assess
the expression of CD42b in the hearts collected from the four
groups (n=5/group). In addition, immunohistochemistry was employed
to detect the recruitment of Ly6G+, CD3+ and
B220+ cells in the myocardial tissues collected from the
mice in the WT + HFD + I/R and LCN2−/− +HFD + I/R groups
(n=5/group).
Identification of mouse genes
Cyagen Biosciences, Inc. provided C57BL/6J mice with
a background that lacked the LCN2 gene (LCN2−/−). Mouse
tail genomic DNA was extracted using lysis buffer (cat. no.
GD01-02; Bioland Scientific LLC) from mouse tail tissue, which was
mixed with DNA stabilization buffer. PCR was then conducted using
the 2X Es Taq Master Mix (cat. no. CW0690M; Jiangsu CoWin Biotech
Co., Ltd.), as follows: 35 cycles of 94°C for 30 sec, 60°C for 30
sec and 72°C for 30 sec. The following PCR primers were utilized:
LCN2, forward (F) 5′-CAACTCAGAACTTGATCCCTGCC-3′ and reverse (R)
5′-TTTCCCTAAGTCCCGTTCAATCC-3′. Agarose gel electrophoresis (2%) was
used to determine the size of the PCR products and the gels were
stained with nucleic acid dye SuperRed/GelRed (cat. no. BS354A;
Biosharp Life Sciences). For WT mice, a PCR product of 1,287 bp was
generated. By contrast, a PCR product of 586 bp was generated for
LCN2−/− mice.
Establishment of a MI/R injury model
in mice
Male mice were anesthetized by intraperitoneal
injection of Avertin (1.25%, 250 mg/kg) (cat. no. BR4108423;
Bioleaper). After anesthesia, the animals naturally laid down;
their head, limbs, tail and whiskers did not respond to touch,
their heartbeat and breathing were uniform in a supine position,
and their muscles were relaxed. These signs indicated full
anesthesia and that the subsequent experimental operations could be
carried out. The tongue was pulled out with tweezers and
endotracheal intubation was performed through the glottis. A 1.5-cm
incision was made at the intersection of the skin between the left
midclavicular line and the fourth intercostal space in mice, and
hemostatic forceps were used to stretch the fourth intercostal
space to expose the heart. The left anterior descending (LAD)
coronary artery was identified, and a 1.5-mm suture was made at the
lower margin of the left auricle through the LAD coronary artery
and tied into a knot. In addition, the sham operation group
underwent the same procedure with the suture passing under the LAD
branch, but without binding.
H&E and TTC staining
Mice were intraperitoneally injected with 1%
pentobarbital sodium (150 mg/kg) and were sacrificed by cervical
dislocation. Death was verified by the absence of a heartbeat and
dilated pupils. Liver and heart samples were then obtained and
fixed with 4% paraformaldehyde (cat. no. P0099; 100 ml; Beyotime
Institute of Biotechnology) for 24 h at room temperature. The liver
and heart tissues were then dehydrated, embedded in paraffin and
sliced into 5-µm sections. These tissue sections were placed onto
slides and subjected to dewaxing before staining with the H&E
staining kit (cat. no. C0105M; Beyotime Institute of
Biotechnology). The staining procedure was carried out according to
the manufacturer's instructions at room temperature, with
hematoxylin staining performed for 6 min and eosin staining for 1
min. The degree of MI was assessed through TTC staining (cat. no.
T8170; Beijing Solarbio Science & Technology Co., Ltd.)
according to the manufacturer's instructions. Images of the tissue
sections were captured using an optical microscope (80i; Nikon
Corporation).
Immunohistochemistry
The heart tissues of mice collected from different
treatment groups were fixed with 4% paraformaldehyde solution for
24 h and dehydrated in an ascending series of alcohol at room
temperature. After permeabilization with xylene for 30 min, heart
tissues were then embedded in paraffin and sliced into 5-µm
sections. These tissue sections were attached to slides and dewaxed
with xylene. For antigen retrieval, they were placed in dyeing
tanks containing citrate buffer, and were boiled in a pressure
cooker for 15 min. To inhibit the activity of endogenous
peroxidase, a 3% solution of hydrogen peroxide was applied for 10
min at room temperature. The heart tissue sections were then
blocked with goat serum (cat. no. SAP-9100; Beijing Zhongshan
Jinqiao Biotechnology Co., Ltd.) at 24°C for 30 min, and were
subsequently incubated with primary antibodies [anti-CD42b antibody
(1:200; cat. no. ab183345; Abcam), anti-CD3 antibody (1:200; cat.
no. ab16669; Abcam), anti-B220 antibody (1:200; cat. no. ab10558;
Abcam) and anti-Ly6G antibody (1:200; cat. no. ab238132; Abcam)] at
4°C for 12 h, followed by incubation with a horseradish peroxidase
(HRP)-IgG secondary antibody (1:50; cat. no. A0208; Beyotime
Institute of Biotechnology) at 24°C for 60 min. The expression of
specific proteins in the heart tissues was visualized using DAB
staining. Images of the tissue sections were captured using an
optical microscope (80i; Nikon Corporation). Three visual field
images of the infarct area were obtained from each histochemical
section. The DAB-positive cell number in each image was manually
counted by ImageJ (version 1.8.0; National Institutes of Health),
and the mean value was defined as the positive cell number in the
section.
Platelet separation
All blood samples used in the present study were
obtained from healthy volunteers (age, 18–26 years) who provided
written informed consent. Blood samples were collected from 18
volunteers, including 10 men and 8 women, with an mean age of 24
years, between March 2021 and March 2023. The Human Ethics
Committee of Guizhou Medical University approved the experimental
procedures [(approval no. 2021 (35)]. The collected blood was stored in a
VACUETTE® vessel and mixed with 0.109 M sodium citrate
at a ratio of 9:1. Subsequently, centrifugation at 120 × g for 20
min at 25°C was conducted to obtain the washed platelets for
further experiments.
ELISA
After mice were intraperitoneally injected with 1%
pentobarbital sodium (150 mg/kg), the blood serum was obtained by
centrifuging blood samples (150 µl) collected from the tail veins
of mice at 3,000 × g for 15 min at room temperature. The serum LCN2
levels were determined using a mouse ELISA kit (cat. no. ab199083;
Abcam) according to the manufacturer's instructions.
Detection of platelet aggregation,
activation and adhesion
Platelets were exposed to LCN2 (2 µg/ml; cat. no.
1757-LC-050; R&D Systems, Inc.) for 5 min at 37°C, and were
then stimulated with thrombin (0.04 U/ml; cat. no. T6884;
Sigma-Aldrich; Merck KGaA), collagen (1 µg/ml; cat. no. P/N 385;
Chrono-Log Corporation) or adenosine diphosphate (ADP; 2.5 µM; cat.
no. A2754; Sigma-Aldrich; Merck KGaA) for 5 min at 37°C. An
equivalent volume of normal saline was used as a vehicle.
Aggregation of platelets was then assessed using an aggregator
(Model 700; Chrono-Log Corporation), while constant stirring was
maintained at a rate of 1,200 rpm for 5 min. The expression levels
of P-selectin were analyzed by performing flow cytometric analysis.
Platelets were stimulated with or without LCN2 (2 µg/ml; cat. no.
1757-LC-050; R&D Systems, Inc.) for 5 min at 37°C, and were
then exposed to thrombin for 5 min at 37°C; untreated platelets
were used as a control. After incubating with FITC-conjugated
P-selectin antibodies (0.2 µg/ml; cat. no. ab33279; Abcam) for 20
min at 37°C, the expression of P-selectin on the platelet surface
was detected by flow cytometry (CytoFLEX S; Beckman Coulter, Inc.).
Flow cytometry data were analyzed using CytExpert software (version
2.3.1.22; Beckman Coulter, Inc.).
To investigate the adhesion of platelets, cellular
coverslips were categorized into three distinct groups: The first
group was coated with 1% bovine serum albumin (cat. no. A8020;
Beijing Solarbio Science & Technology Co., Ltd.) for 1 h at
20°C and subsequently incubated with non-treated platelets
(2×107). The remaining two groups were coated with 5
µg/ml collagen for 14 h at 4°C, and incubated with 2 µg/ml
LCN2-stimulated platelets (2×107) and unstimulated
platelets (2×107) at 37°C. After a 45-min incubation,
the platelets were stained with iFluor™ 680-phalloidin
(1 µg/ml; cat. no. 40788ES75; Shanghai Yeasen Biotechnology Co.,
Ltd.) for 25 min at 37°C, and the adhesion of platelets was
observed using a fluorescence microscope.
Assessment of clot retraction
Platelets were incubated with or without LCN2 (2
µg/ml) for 5 min at 37°C, and subsequently stimulated with thrombin
(0.4 U/ml) at 37°C. The process of clot retraction was monitored
and recorded at intervals of 0, 20, 40 and 60 min after thrombin
stimulation by capturing images of platelet clots.
RT-qPCR
Liver tissues were collected for extraction of total
RNA using a DNA/RNA co-extraction kit (cat. no. DP422; Tiangen
Biotech Co., Ltd.). Subsequently, cDNA synthesis was carried out
using the FastKing gDNA Dispelling RT SuperMix (cat. no. KR118;
Tiangen Biotech Co., Ltd.) according to the manufacturer's
instruction. qPCR was performed according to the protocol specified
by the manufacturer of FastFire qPCR PreMix (SYBR Green) (cat. no.
FP207; Tiangen Biotech Co., Ltd.). The qPCR program consisted of an
initial phase of pre-denaturation at 95°C for 1 min, followed by 40
cycles at 95°C for 5 sec, 55°C for 10 sec and 72°C for 15 sec. For
the expression analysis of the target genes, GAPDH was employed as
the internal reference for normalization. The primer sequences were
as follows, LCN2, F 5′-AAGGCAGCTTTACGATGT-3′, R
5′-TGGTTGTAGTCCGTGGTG-3′; GAPDH, F 5′-GCACAGTCAAGGCCGAGAAT-3′ and R
5′-GCCTTCTCCATGGTGGTGAA-3′. Data were normalized to GAPDH levels as
a reference. Subsequently, the gene expression levels were
calculated using the 2−ΔΔCq method (21).
Statistical analysis
The data were analyzed using GraphPad Prism v5.01
(Dotmatics), and the results are presented as the mean ± SEM
derived from ≥3 independent experiments. Differences between two
groups were assessed using unpaired Student's t-test, while
differences among multiple groups were determined by one-way ANOVA
and Tukey's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
LCN2 expression is increased in the
liver and serum of mice fed a HFD
Notably, LCN2 has been shown to be highly expressed
in adipose tissue, and to serve a role in both obesity and
obesity-related diseases (22,23).
Considering that obesity is an important risk factor for CAD and
MI/R injury, a mouse model of obesity was established in the
present study through the use of a HFD. The results revealed a
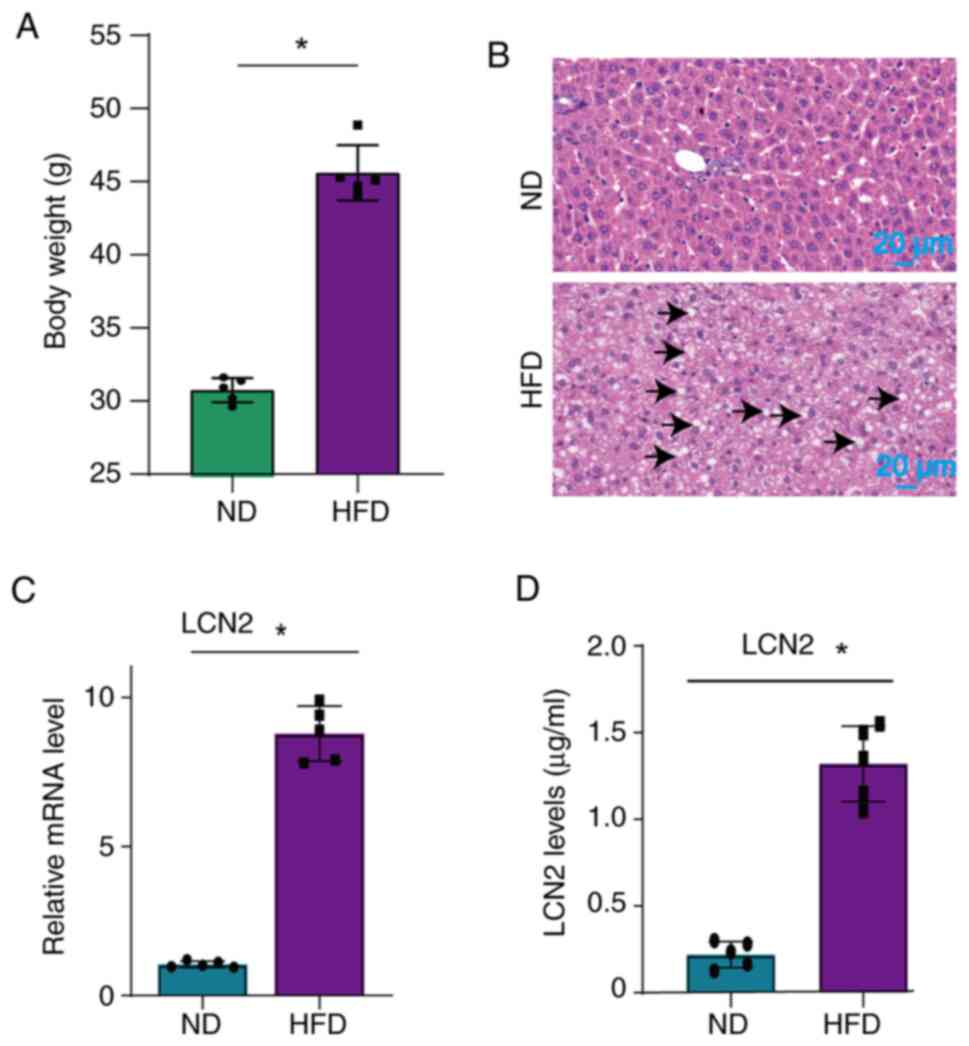
significant increase in the weight of the mice (Fig. 1A), as well as the accumulation of
large lipid droplets in their liver tissues (Fig. 1B) following administration of a
HFD. Additionally, a significant upregulation of LCN2 levels was
observed in both the liver (Fig.
1C) and serum (Fig. 1D) of
mice fed a HFD.
LCN2 potentiates platelet
function
In cardiovascular disorders, the formation of
atherosclerotic plaques and microthrombi are the consequences of
platelet aggregation, adhesion and activation (24). These processes also contribute to
the worsening of MI/R injury. Previous research has indicated that
endogenous proteins possessing pro-inflammatory properties can
enhance platelet hyperresponsiveness, and exacerbate
atherosclerosis and MI/R injury by promoting platelet aggregation
(25,26). The present study investigated the
effects of LCN2 on platelet function. When platelet agonists
(thrombin, collagen and ADP) were applied, incubation with LCN2
promoted platelet aggregation (Fig.
2A). Additionally, compared with the untreated platelets, a
higher number of platelets adhered to collagen-coated coverslips
following treatment with LCN2 (Fig.
2B). Furthermore, it was observed that LCN2-treated platelets
had more pronounced clot retraction (Fig. 2C) and higher expression of
P-selectin (Fig. 2D) upon
stimulation with thrombin.
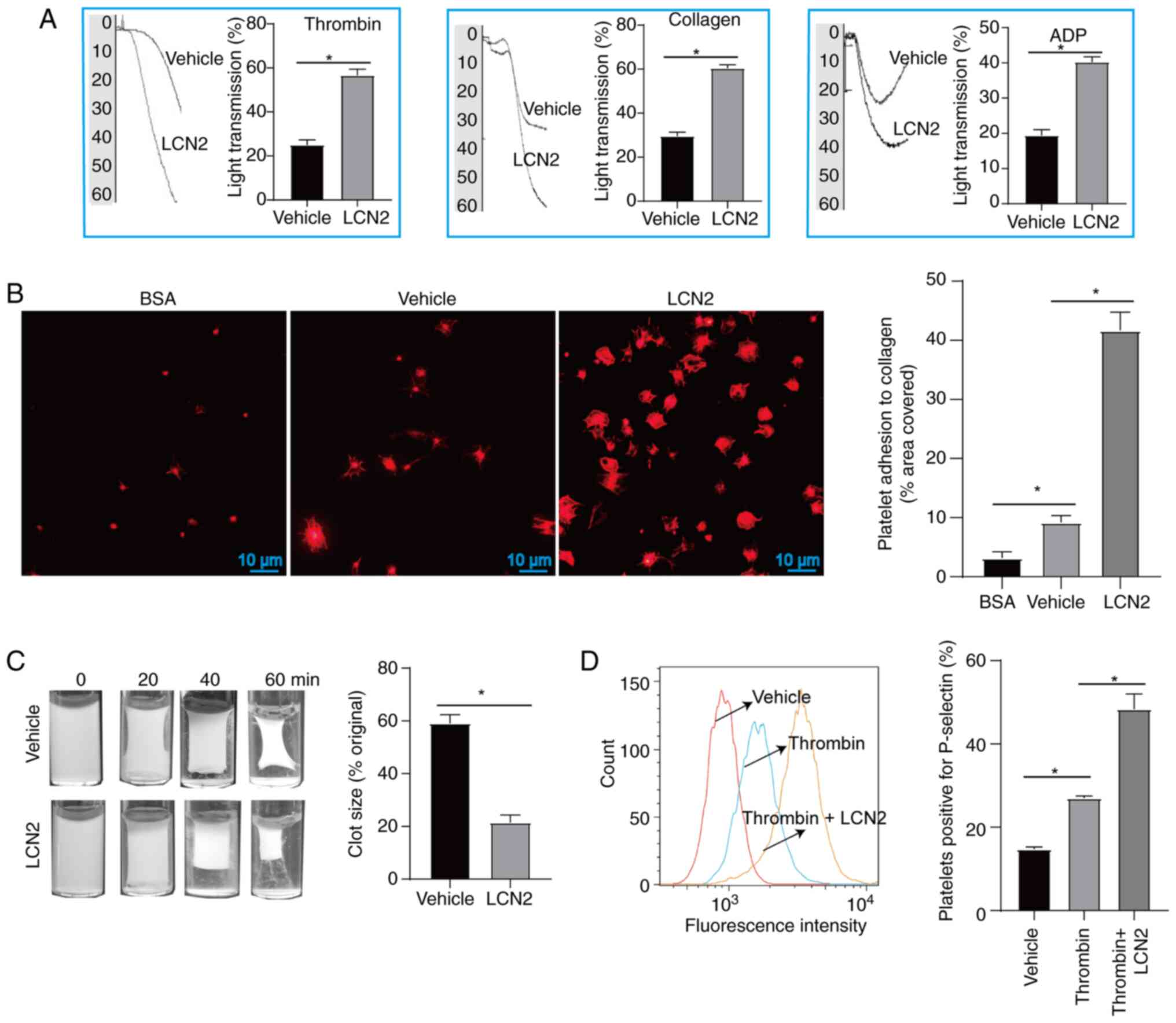 | Figure 2.Effect of LCN2 on platelet function.
(A) After platelets were incubated with or without LCN2, they were
further stimulated with ADP, thrombin or collagen to detect
platelet aggregation (5 platelet samples were collected from 5
volunteers). (B) After platelets from different treatment groups
were incubated on cellular coverslips for 45 min, they were stained
with iFluor™ 680-phalloidin, and the adhesion of
platelets was observed under a fluorescence microscope (5 platelet
samples were collected from 5 volunteers). (C) After incubation
with or without LCN2, the platelets were stimulated with thrombin.
Clot retraction was observed after stimulation with thrombin for 0,
20, 40 and 60 min (5 platelet samples were collected from 5
volunteers). (D) Upon stimulation with or without LCN2, the
platelets were incubated with thrombin, and untreated platelets
were used as a control. The expression of P-selectin on the
platelet surface was detected by flow cytometry (3 platelet samples
were collected from 3 volunteers). Data are presented as the mean ±
SEM. *P<0.05. ADP, adenosine diphosphate; BSA, bovine serum
albumin; LCN2, lipocalin 2. |
Effect of LCN2 knockout on MI/R injury
in HFD-treated mice
Obesity serves an important role in the development
of cardiovascular diseases. One of the main contributors to MI/R
injury in obese individuals is the increase in endogenous bioactive
molecules, which stimulate platelet activation (25). The gene phenotypes of WT mice and
LCN2−/− mice were determined by PCR (Fig. 3A). For WT mice, a PCR product of
1,287 bp was generated, whereas a PCR product of 586 bp was
generated for LCN2−/− mice. The PCR product of 586 bp in
LCN2−/− mice indicated that LCN2 was knocked out,
whereas the PCR product of 1,287 bp in WT mice indicated that LCN2
was not knocked out. These mice were then subjected to MI/R injury
or sham operation after being fed a HFD. Subsequently, cardiac
pathology was observed by H&E staining (Fig. 3B). After sham operation, the
myocardia of WT mice and LCN2−/− mice were closely
arranged, with normal structure and no obvious inflammatory cell
infiltration. WT mice and LCN2−/− mice with MI/R injury
exhibited notable histological features of myocardial tissue injury
and inflammatory cell infiltration. In the case of MI/R injury,
there was less structural damage to the myocardium and relatively
less inflammatory cell infiltration in LCN2−/− mice
compared with that in the WT mice (Fig. 3B). TTC staining showed no obvious
ischemic features in the myocardia after sham operation in WT mice
and LCN2−/− mice. After MI/R injury, the myocardial
infarct size of LCN2−/− mice was smaller than that of WT
mice (Fig. 3C).
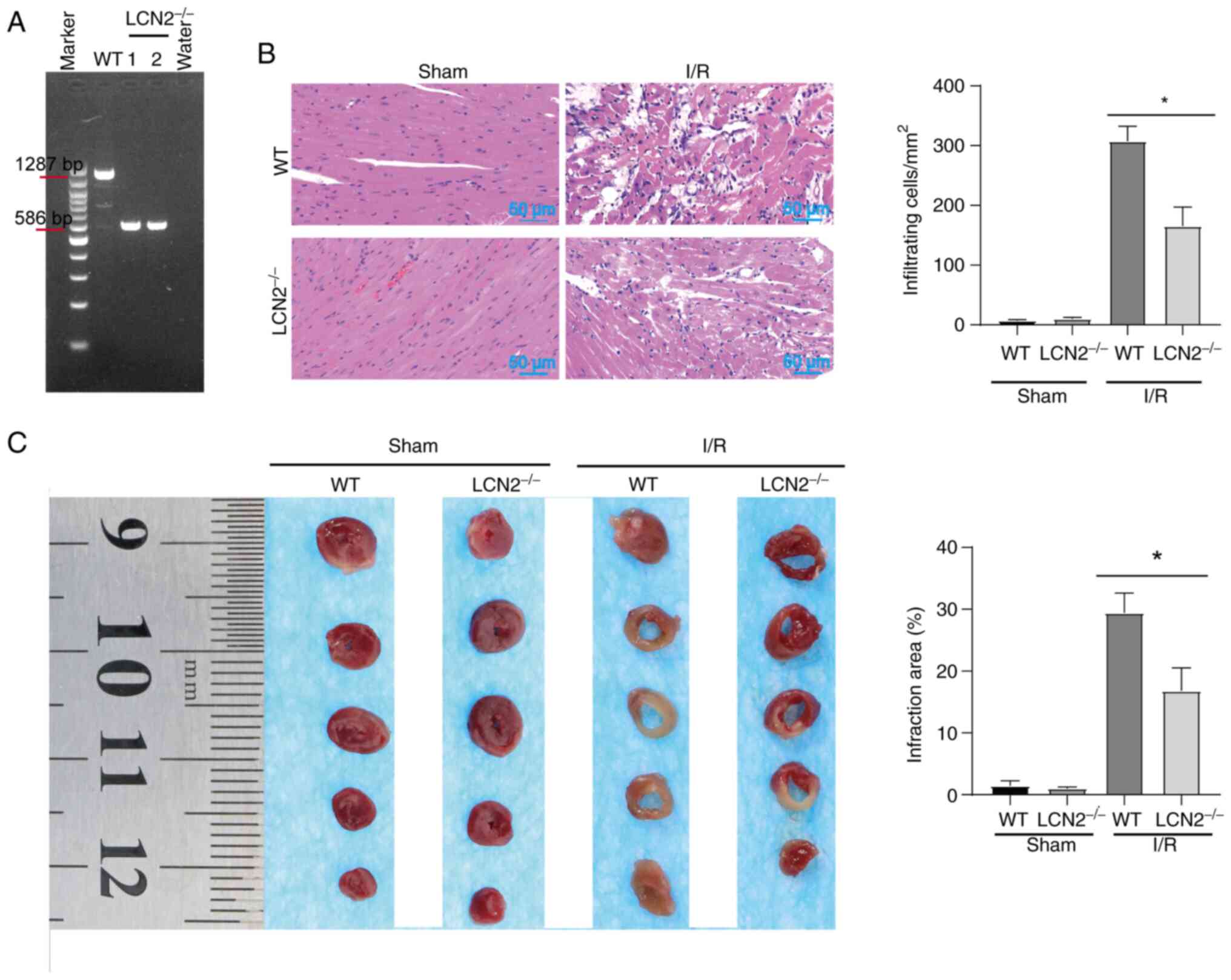 | Figure 3.Effect of LCN2 knockout on MI/R
injury in HFD-fed mice. (A) Mouse tails were utilized for DNA
extraction, followed by PCR amplification and electrophoresis. For
WT mice, a PCR product of 1,287 bp was obtained; conversely,
LCN2−/− mice had a product of 586 bp. Subsequently, both
WT and LCN2−/− mice underwent MI/R injury and sham
surgery after being fed a HFD. (B) Mouse hearts were procured for
hematoxylin and eosin staining. (C) Degree of infarction was
assessed through TTC staining. Data are presented as the mean ± SEM
(n=5). *P<0.05. HFD, high-fat diet; LCN2, lipocalin 2; MI/R,
myocardial ischemia-reperfusion; TTC, 2,3,5-triphenyltetrazolium
chloride; WT, wild-type. |
Effect of LCN2 knockout on the
recruitment of platelets and inflammatory cells in the myocardial
tissue of HFD-fed mice subjected to MI/R injury
During MI/R injury, platelets tend to form
microthrombi within the microvessels of the heart and worsen
myocardial damage by releasing certain bioactive molecules
(24). Thus, the current study
aimed to investigate whether the absence of LCN2 affects the
accumulation of platelets induced by I/R injury in myocardial
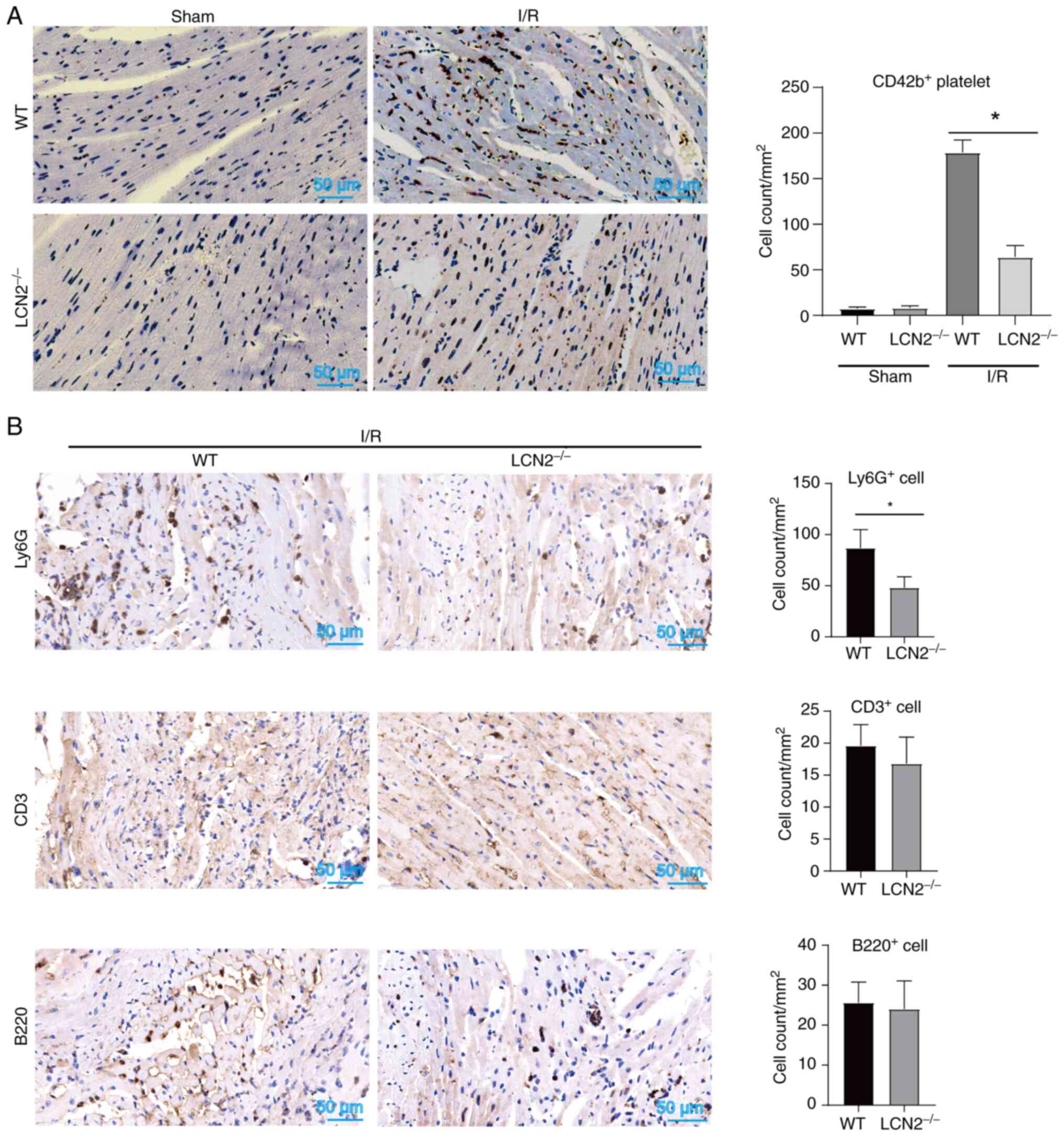
tissue. As shown in Fig. 4A, the
number of CD42b+ platelets in myocardial tissues was
lower in LCN2−/− mice compared with that in WT mice
following reperfusion post-ischemia. These findings suggested that
LCN2 induced by HFD may enhance platelet accumulation in myocardial
tissue during MI/R injury. Notably, activated platelets form
complexes with leukocytes and adhere to microvessels within the
heart, thereby exacerbating MI/R injury. Suppression of platelet
activation can reduce the recruitment of inflammatory cells in
myocardial tissue, ultimately mitigating MI/R injury (24,25).
In the present study, it was observed that LCN2 gene knockout
decreased the accumulation of Ly6G+ cells within the
region of myocardial injury induced by I/R, while having no
significant impact on the recruitment of CD3+ or
B220+ cells (Fig. 4B).
These results indicated that LCN2 knockout might alleviate MI/R
injury by inhibiting platelet and Ly6G+ cell
recruitment.
Discussion
In diseases related to obesity, an increase in
platelet volume is observed. Additionally, there is an increase in
the expression of thrombolane B2, prostaglandin F2, soluble
P-selectin and soluble CD40L. These changes occur due to continuous
stimulation from free fatty acids, oxidized low-density
lipoprotein, inflammatory cytokines, enhanced oxidative stress,
elevated stress proteins and other factors. Additionally, platelets
release a higher number of particles, indicating a hyperreactive
state (27–29). This hyperreactivity of platelets
leads to their adherence to damaged endothelial cells, which serves
a role in the development of atherosclerosis. Furthermore,
activated platelets secrete certain specific bioactive molecules,
such as myeloperoxidase and ROS, which participate in MI/R injury
(30–34). Therefore, understanding how obesity
impacts platelet function is crucial in identifying its role in
exacerbating MI/R injury.
Currently, the focus of research on the impact of
platelets on MI/R injury lies primarily in the analysis of the
active molecules that are secreted during platelet activation, the
surface molecules of platelets and the increased expression of
endogenous bioactive molecules affecting platelet function. A
previous study confirmed a significant positive correlation between
plasma galectin-3 and platelet aggregation in patients with CAD.
Moreover, galectin-3 was found to enhance platelet activation and
thrombosis in animal models (25).
Thus, the present findings supplement the understanding of the
involvement of LCN2 in cardiovascular disease and contribute
valuable insights to clinical science. Currently, known
antiplatelet drugs do not aim to reduce the expression of
endogenous molecules that activate platelets (24). This particular phenomenon may help
to explain the uncertainty surrounding the efficacy of antiplatelet
drugs in MI/R treatment.
The current research strategy was similar to that of
Chen et al. In this study, the endogenous molecule
galectin-3 was found to enhance platelet function to aggravate MI/R
injury (25). Since a HFD, which
is a risk factor of cardiovascular disease, could induce an
increase in LCN2 expression, the present study further confirmed
that LCN2 could promote platelet aggregation, adhesion and plaque
retraction through in vitro experiments. These findings
suggested that, similar to the function of galectin-3, LCN2 may
have a role in enhancing platelet function. Subsequently, the
current study further explored whether LCN2 could enhance the local
accumulation of platelets in injured myocardium during MI/R in
HFD-treated mice, as platelet accumulation has been considered to
be an important factor in exacerbating I/R-induced myocardial
injury. Under a HFD, the pathological damage of myocardial tissues
was significantly lower in LCN2−/− mice than in WT mice
when suffering from MI/R injury. Furthermore, fewer platelets
accumulated in the injured myocardial tissues of LCN2−/−
mice, which indicated that LCN2 was an important inducer of MI/R
injury in HFD mice. Combined with the in vitro evidence that
LCN2 promoted platelet function, it was hypothesized that
HFD-induced LCN2 may aggravate MI/R injury, which could be fully or
partially dependent on the enhancement of platelet activation and
aggregation. In addition to the aforementioned results, it was
observed that fewer Ly6G+ cells, rather than
CD3+ and B220+ cells, accumulated in the
injured myocardium of LCN2−/− mice when MI/R injury
occurred. A previous study reported that LCN2 can aggravate
psoriasis by inducing neutrophil chemotaxis and activation
(35), and the present results
also suggested that the effect of LCN2 on neutrophil function may
serve a role in promoting MI/R injury. However, additional research
is needed to fully confirm these possible mechanisms.
LCN2 has previously been reported to be involved in
heart disease, as demonstrated by Jang et al (36). In this previous study, a mouse
model of cardiomyopathy was established using adriamycin. Notably,
it was observed that LCN2 knockout mice experienced milder
myocardial injury after adriamycin induction, and this effect of
LCN2 was related to its function of inhibiting autophagic flow.
While the present study supports the potential pro-atherosclerotic
effects of LCN2, specifically its regulatory function on platelets,
it also raises important scientific questions for further
exploration. Firstly, it is known that MI/R injury is a consequence
of functional changes in multiple cell types. Beyond cardiomyocytes
and cardiac microvascular endothelial cells, T helper (Th)17 cell
differentiation has been implicated in the MI/R process (37). Notably, a previous study
demonstrated that LCN2 exacerbated psoriasis-like skin inflammation
by enhancing the Th17 response (38), which suggests that LCN2 may
potentially worsen MI/R injury by enhancing the Th17 reaction.
Furthermore, LCN2 is capable of activating various signaling
pathways, including JAK/STAT and NF-κB, which have important roles
in MI/R injury (39,40). Therefore, it is worth investigating
whether LCN2 aggravates MI/R injury by influencing the activation
of these signaling pathways. Moreover, potential receptors for
LCN2, such as the glycoproteins GP330 and SLC22A17, have been
identified (40). However, it
remains unclear whether these receptors are expressed in platelets
and whether LCN2 exerts its function through these receptors.
Finally, exploring the development of drugs that effectively
inhibit LCN2 function could lead to clinical applications for
improving MI/R injury. These scientific questions will be the focus
of further exploration in subsequent research.
In conclusion, the present study demonstrated that
HFD-induced LCN2 exacerbated MI/R injury by promoting platelet
activation, aggregation and adhesion.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Guizhou Provincial Health
Commission (grant nos. gzwkj2022-031, gzwkj2022-254 and
gzwkj2022-174), the Guizhou Administration of Traditional Chinese
Medicine (grant no. QZYY-2021-145), and the Guiyang Science and
Technology Bureau [grant no. (2019)9-12-3].
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
XL and QW conceived and designed the experiments. PL
performed the experiments. PL, JC and MW analyzed the data and
prepared the figures. XL and PL confirm the authenticity of all the
raw data. All authors contributed to the article, and read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The experimental procedures carried out in the
present study were approved by the animal experiment ethics
committee of Guizhou Medical University (approval no. 2100328) and
the Human Ethics Committee of Guizhou Medical University [approval
no. 2021 (35)]. All blood samples
used in the current study were obtained from healthy volunteers who
provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Harrington DH, Stueben F and Lenahan CM:
ST-elevation myocardial infarction and non-ST-elevation myocardial
infarction: Medical and surgical interventions. Crit Care Nurs Clin
North Am. 31:49–64. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu MY, Yiang GT, Liao WT, Tsai AP, Cheng
YL, Cheng PW, Li CY and Li CJ: Current mechanistic concepts in
ischemia and reperfusion injury. Cell Physiol Biochem.
46:1650–1667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Visseren FLJ, Mach F, Smulders YM,
Carballo D, Koskinas KC, Bäck M, Benetos A, Biffi A, Boavida JM,
Capodanno D, et al: 2021 ESC guidelines on cardiovascular disease
prevention in clinical practice. Eur Heart J. 42:3227–3337. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Algoet M, Janssens S, Himmelreich U, Gsell
W, Pusovnik M, Van den Eynde J and Oosterlinck W: Myocardial
ischemia-reperfusion injury and the influence of inflammation.
Trends Cardiovasc Med. 33:357–366. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei Y, Xing J, Su X, Li X, Yan X, Zhao J
and Tao H: IL-38 attenuates myocardial ischemia-reperfusion injury
by inhibiting macrophage inflammation. Immun Inflamm Dis.
11:e8982023. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang J, Hu S, Gao Y, Wei X, Qu Y, Gao R,
Lv Y, Wang J, Wang Y, Yang J, et al: Galangin alleviated myocardial
ischemia-reperfusion injury by enhancing autophagic flux and
inhibiting inflammation. Eur J Pharmacol. 945:1756212023.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen LQ, Wang WS, Li SQ and Liu JH:
Minocycline relieves myocardial ischemia-reperfusion injury in rats
by inhibiting inflammation, oxidative stress and apoptosis. Eur Rev
Med Pharmacol Sci. 26:3001–3009. 2022.PubMed/NCBI
|
|
8
|
Seeger JP, Benda NM, Riksen NP, van Dijk
AP, Bellersen L, Hopman MT, Cable NT and Thijssen DH: Heart failure
is associated with exaggerated endothelial ischaemia-reperfusion
injury and attenuated effect of ischaemic preconditioning. Eur J
Prev Cardiol. 23:33–40. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wereski R, Kimenai DM, Bularga A, Taggart
C, Lowe DJ, Mills NL and Chapman AR: Risk factors for type 1 and
type 2 myocardial infarction. Eur Heart J. 43:127–135. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cameron SJ, Ture SK, Mickelsen D,
Chakrabarti E, Modjeski KL, McNitt S, Seaberry M, Field DJ, Le NT,
Abe J and Morrell CN: Platelet extracellular regulated protein
kinase 5 is a redox switch and triggers maladaptive platelet
responses and myocardial infarct expansion. Circulation. 132:47–58.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schanze N, Bode C and Duerschmied D:
Platelet contributions to myocardial ischemia/reperfusion injury.
Front Immunol. 10:12602019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van der Meijden PEJ and Heemskerk JWM:
Platelet biology and functions: New concepts and clinical
perspectives. Nat Rev Cardiol y. 16:166–179. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jaberi SA, Cohen A, D'Souza C, Abdulrazzaq
YM, Ojha S, Bastaki S and Adeghate EA: Lipocalin-2: Structure,
function, distribution and role in metabolic disorders. Biomed
Pharmacother. 142:1120022021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ghosh S, Stepicheva N, Yazdankhah M, Shang
P, Watson AM, Hose S, Liu H, Weiss J, Zigler JS Jr, Valapala M, et
al: The role of lipocalin-2 in age-related macular degeneration
(AMD). Cell Mol Life Sci. 77:835–851. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eilenberg W, Stojkovic S,
Piechota-Polanczyk A, Kaun C, Rauscher S, Gröger M, Klinger M,
Wojta J, Neumayer C, Huk I and Demyanets S: Neutrophil
gelatinase-associated lipocalin (NGAL) is associated with
symptomatic carotid atherosclerosis and drives pro-inflammatory
state in vitro. Eur J Vasc Endovasc Surg. 51:623–631. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mosialou I, Shikhel S, Luo N, Petropoulou
PI, Panitsas K, Bisikirska B, Rothman NJ, Tenta R, Cariou B, Wargny
M, et al: Lipocalin-2 counteracts metabolic dysregulation in
obesity and diabetes. J Exp Med. 217:e201912612020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Catalán V, Gómez-Ambrosi J, Rodríguez A,
Ramírez B, Silva C, Rotellar F, Gil MJ, Cienfuegos JA, Salvador J
and Frühbeck G: Increased adipose tissue expression of lipocalin-2
in obesity is related to inflammation and matrix
metalloproteinase-2 and metalloproteinase-9 activities in humans. J
Mol Med (Berl). 87:803–813. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Flo TH, Smith KD, Sato S, Rodriguez DJ,
Holmes MA, Strong RK, Akira S and Aderem A: Lipocalin 2 mediates an
innate immune response to bacterial infection by sequestrating
iron. Nature. 432:917–921. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Lam KS, Kraegen EW, Sweeney G,
Zhang J, Tso AW, Chow WS, Wat NM, Xu JY, Hoo RL and Xu A:
Lipocalin-2 is an inflammatory marker closely associated with
obesity, insulin resistance, and hyperglycemia in humans. Clin
Chem. 53:34–41. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jang Y, Lee JH, Wang Y and Sweeney G:
Emerging clinical and experimental evidence for the role of
lipocalin-2 in metabolic syndrome. Clin Exp Pharmacol Physiol.
39:194–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu G, Li H, Zhou M, Fang Q, Bao Y, Xu A
and Jia W: Mechanism and clinical evidence of lipocalin-2 and
adipocyte fatty acid-binding protein linking obesity and
atherosclerosis. Diabetes Metab Res Rev. 30:447–456. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Abella V, Scotece M, Conde J, Gómez R,
Lois A, Pino J, Gómez-Reino JJ, Lago F, Mobasheri A and Gualillo O:
The potential of lipocalin-2/NGAL as biomarker for inflammatory and
metabolic diseases. Biomarkers. 20:565–571. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schanze N, Hamad MA, Nührenberg TG, Bode C
and Duerschmied D: Platelets in myocardial ischemia/reperfusion
injury. Hamostaseologie. 43:110–121. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen Y, Fu W, Zheng Y, Yang J, Liu Y, Qi
Z, Wu M, Fan Z, Yin K, Chen Y, et al: Galectin 3 enhances platelet
aggregation and thrombosis via Dectin-1 activation: A translational
study. Eur Heart J. 43:3556–3574. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen J, Liu G, Hong Y, Han J, Yang Z, Yang
Y, Li H, Wang S, Jue L and Wang Q: Regulation of atherosclerosis by
toll-like receptor 4 induced by serum amyloid 1: A systematic in
vitro study. Biomed Res Int. 2022:48875932022. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Anfossi G, Russo I and Trovati M: Platelet
dysfunction in central obesity. Nutr Metab Cardiovasc Dis.
19:440–449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Santilli F, Vazzana N, Liani R, Guagnano
MT and Davì G: Platelet activation in obesity and metabolic
syndrome. Obes Rev. 13:27–42. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barale C and Russo I: Influence of
cardiometabolic risk factors on platelet function. Int J Mol Sci.
21:6232020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mauler M, Herr N, Schoenichen C, Witsch T,
Marchini T, Härdtner C, Koentges C, Kienle K, Ollivier V, Schell M,
et al: Platelet serotonin aggravates myocardial
ischemia/reperfusion injury via neutrophil degranulation.
Circulation. 139:918–931. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Davidson SM, Andreadou I, Barile L,
Birnbaum Y, Cabrera-Fuentes HA, Cohen MV, Downey JM, Girao H,
Pagliaro P, Penna C, et al: Circulating blood cells and
extracellular vesicles in acute cardioprotection. Cardiovasc Res.
115:1156–1166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gumiężna K, Baruś P, Sygitowicz G,
Wiśniewska A, Ochijewicz D, Pasierb K, Klimczak-Tomaniak D,
Kuca-Warnawin E, Kochman J, Grabowski M, et al: Immature platelet
fraction in cardiovascular diagnostics and antiplatelet therapy
monitoring. Cardiol J. 30:817–824. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Margraf A and Zarbock A: Platelets in
inflammation and resolution. J Immunol. 203:2357–2367. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huilcaman R, Venturini W, Fuenzalida L,
Cayo A, Segovia R, Valenzuela C, Brown N and Moore-Carrasco R:
Platelets, a key cell in inflammation and atherosclerosis
progression. Cells. 11:10142022. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shao S, Cao T, Jin L, Li B, Fang H, Zhang
J, Zhang Y, Hu J and Wang G: Increased lipocalin-2 contributes to
the pathogenesis of psoriasis by modulating neutrophil chemotaxis
and cytokine secretion. J Invest Dermatol. 136:1418–1428. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jang HM, Lee JY, An HS, Ahn YJ, Jeong EA,
Shin HJ, Kim KE, Lee J, Koh JS and Roh GS: LCN2 deficiency
ameliorates doxorubicin-induced cardiomyopathy in mice. Biochem
Biophys Res Commun. 588:8–14. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li D, Yang Z, Gao S, Zhang H and Fan G:
Tanshinone IIA ameliorates myocardial ischemia/reperfusion injury
in rats by regulation of NLRP3 inflammasome activation and Th17
cells differentiation. Acta Cir Bras. 37:e3707012022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hau CS, Kanda N, Tada Y, Shibata S, Uozaki
H, Fukusato T, Sato S and Watanabe S: Lipocalin-2 exacerbates
psoriasiform skin inflammation by augmenting T-helper 17 response.
J Dermatol. 43:785–794. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen PC, Ho CH, Fan CK, Liu SP and Cheng
PC: Antimicrobial peptide LCN2 inhibited uropathogenic escherichia
coli infection in bladder cells in a high-glucose environment
through JAK/STAT signaling pathway. Int J Mol Sci. 23:157632022.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim SL, Shin MW, Seo SY and Kim SW:
Lipocalin 2 potentially contributes to tumorigenesis from colitis
via IL-6/STAT3/NF-κB signaling pathway. Biosci Rep.
42:BSR202124182022. View Article : Google Scholar : PubMed/NCBI
|