Introduction
Several factors, such as congenital malformation of
the cleft lip and palate, drug treatment, local inflammation,
periodontitis, traumatic injury, malignant tumors and dental
surgery, can cause alveolar bone defects (1). Repairing alveolar bone injury or
periodontal soft tissue injury is a complex process (2). Due to its favorable osteogenesis and
osteoinduction characteristics, autogenous bone transplantation is
commonly performed for treating alveolar bone defects (2). However, autologous bone
transplantation depends on the donor site conditions,
transplantation failure rate and immune rejection of patients
(3). Furthermore, the sources of
autologous bone grafts are limited, the treatment is painful and
the donor site is damaged (3).
Other bone tissue substitutes, such as xenotransplantation, are
available. However, this approach is hindered by immune rejection
and high pathogen transmission (4). Due to the poor integration with
natural bone tissue, synthetic grafts often result in graft
failure. Therefore, the application of synthetic grafts is limited
(5). A suitable technique needs to
be developed to fully regenerate damaged bone tissues.
The main aim of repairing alveolar bone defects is
to stimulate or induce osteogenic potential and provide the
materials, space and environment needed for osteogenesis (6). Hertwig's epithelial root sheath
(HERS) is a double-layer epithelial structure formed by the fusion
of the inner and outer enamel epithelia in the tooth neck after the
formation of the tooth crown during tooth development (7). It differentiates into cementoblasts
through epithelial-mesenchymal transition (EMT) and promotes the
differentiation of medial dental papilla cells (DPCs) into
odontoblasts through epithelial-mesenchymal interaction, thus
secreting and producing dentin from the tooth root (7). Furthermore, HERS can differentiate
into cementum cells through EMT and secrete cementum to form
periodontal tissue (8). The
cementum is formed by differentiating cementoblasts, which have
similar phenotypes to osteoblasts, and expresses proteins such as
runt-related transcription factor 2 (RUNX2), bone sialoprotein
(BSP) and alkaline phosphatase (ALP) (9). The sources of the differentiation of
cementum may be dental follicle cells and epithelial cells
(7). Considering that the inner
cells of HERS initiate the differentiation of odontoblasts to form
root dentin, HERS is necessary for root development. For example,
if the continuity of HERS is damaged, it cannot induce DPCs to
differentiate into odontoblasts, resulting in dentin defects
(10). Additionally, if the
epithelial root sheath fails to break at a specific time and
adheres to the root dentin surface, the dental follicle cells
cannot differentiate into cementoblasts to form cementum (11).
Several studies have suggested that extracellular
vehicles, including microvesicles and exosomes produced by
mesenchymal stem cells (MSCs) and dental pulp stem cells (DPSCs),
promote their therapeutic potency by mediating cell-cell
communication and transporting paracrine factors to facilitate
angiogenesis, immune regulation and tissue regeneration (12,13).
Some studies have shown that DPSCs can secrete various molecules in
the medium that can be used in regenerative medicine (14,15).
Extracellular vesicles (EVs), which are small endocytic vesicles,
have attracted attention in bone regeneration research (16). Researchers have found that after
coculturing EVs derived from MSCs with chondrocytes, CD73-mediated
adenosine activation of AKT and extracellular regulated protein
kinase (ERK) signaling in EVs derived from MSCs promotes cell
migration, proliferation and matrix synthesis during cartilage
repair (17). The implantation of
EVs into an animal defect model can significantly increase the
number of chondrocytes and heal cartilage defects (18). Although the majority of preliminary
studies indicate that EVs stimulate osteogenesis and angiogenesis
(18,19), the exact mechanism remains
unelucidated. Therefore, reliable methods need to be developed to
identify and purify EVs. Exosomes are formed by the multivesicular
division of cells and have high therapeutic potential (19). However, they neither self-replicate
nor cause autoimmune reactions in the host (19). These paracrine factors can be
encapsulated in biomaterials to maintain their biological activity
and can undergo controlled release (20). Among the different types of
biomaterials studied for the infusion of EVs, hydrogels are the
most user-friendly, cost-effective and accessible material
(21). Therefore, in the present
study an alveolar bone defect model was used to study the
osteogenic effects of DPSC-EV-loaded hydrogels with the aim to
propose a new strategy for the osteogenic potential of DPSC-EVs for
safe cell-free therapy and regeneration of defective alveolar
bone.
Materials and methods
Animals
A total of 24 male Sprague-Dawley (SD) rats (age, 10
weeks old; weight, 200±10 g) purchased from Beijing Weitong Lihua
Biotechnology Co., Ltd. (cat. no. SCXK-2021-0006) were used in the
present study. The rats had free access to food and water. All
experimental manipulations were performed in accordance with the
National Institute of Health Guide for the Care and Use of
Laboratory Animals. All animals were housed in separate cages at
the Beijing MeDeKanNa (MDKN) Biotechnology Co., LTD. (experimental
animal use license no. SYXK-2020-0050), with an ambient temperature
of 20–23°C, a relative humidity of 60% and under a 12 h light-dark
cycle. Animals were fed in the animal room for 7 days before
conducting experimental modeling. The animal treatment groups were
as follows: i) Alveolar bone injury group (n=6); ii) alveolar bone
injury + EV intervention group (100 µg; n=6); iii) alveolar bone
injury + EV hydrogel intervention group (100 µg; n=6); and iv)
alveolar bone injury + EV hydrogel intervention group (100 µg) +
TGF-β1 inhibitor (gavage, 20 mg/kg; three times/week) group (n=6).
The present study was approved by The Animal Welfare Ethics
Committee of Beijing MDKN Biotechnology Co., LTD. (approval no.
MDKN-2022-052; Beijing, China).
Animal alveolar bone defect model
Numerous studies have selected the rat alveolar bone
defect model (22–25). In the present study, the rat
mandible was lifted to expose its maxillary dentition. Under
sterile conditions, a pointed scalpel was used to cut the
mesiopalatal gingiva of the first maxillary molar, the gingival
flap was gently opened, the mesioalveolar bone surface of the first
molar was exposed, and a dental cylindrical needle (diameter, 1.5
mm) was used to remove the mesioalveolar bone of the mesiomedial
root of the first maxillary molar in the rats. Intermittent
grinding was performed and water was sprayed on the alveolar bone.
Finally, a large ball drill (diameter, 4 mm) was used to repair the
hole pattern, and was made to meet the surgical standard of a
hemispherical defect with a diameter of 4 mm and volume of 16.76
mm3. Eventually, a buccal alveolar bone was formed at
the first to third mandibular molars, forming an ~12 mm3
bone defect rat model. After the surgery, treatment with EVs, and a
rehabilitation membrane (HealAll®; Yantai Zhenghai
Bio-Tech Co., Ltd.) was used to cover the alveolar bone defect site
to prevent the impact of other connective tissue. The rats were
euthanized with CO2 at a volume displacement of 30%
vol/min. After euthanasia, indicators such as breathing, heartbeat,
pupils and nerve reflexes were observed to confirm animal
death.
Isolation and culture of HERS
Cells
In the present study, the rats (n=15) were
euthanized, disinfected with 75% ethanol and their mandibles were
separated. Under a stereomicroscope, the first mandibular molar
embryos were separated via microscopic forceps. A total of ~1-mm
thick dental neck tissue was cut along the mineralized edge of the
crown, the separated dental neck tissue was rinsed twice with PBS,
cut into small pieces (1×1 cm), and digested at 37°C for 1 h in a
mixture of type I collagenase and dispase enzymes. The epithelial
culture medium (ScienCell Research Laboratories, Inc.; cat. no.
4101) was added to stop the digestion, the mixture was centrifuged
at 250 × g at 4°C for 5 min to remove the supernatant and the cells
were washed with PBS for 5 min twice. Subsequently, the supernatant
was removed, the cells were resuspended in epithelial culture
medium (ScienCell Research Laboratories, Inc.; cat. no. 4101), the
mixture was inoculated in a 25 ml culture flask and cultured at 5%
CO2 and 37°C, with the medium changed every two days. On
the third day after inoculation, the epithelial cells were purified
when they reached a growth rate of ~70%. The culture medium was
discarded, and the cells were washed once with PBS. After which,
0.5 ml of 0.25% trypsin EDTA was added, and the cells were placed
in a 37°C incubator for 3 min. When the spindle-shaped cells were
observed to shrink under the microscope, 2 ml serum-containing
culture medium (Gibco; Thermo Fisher Scientific, Inc.; cat. no.
16000-044) was added to terminate digestion. The medium was gently
aspirated to remove the shrunken cells, the medium was discarded
and 5 ml epithelial medium (Lifeline Cell Technology; cat. no.
LL-0023) was added for further cultivation at 37°C. After three
days, the aforementioned purification step was repeated once, and
purified P1-generation HERS epithelial cells were obtained.
DPSC isolation
The surface of the teeth was disinfected and
sterilized with 75% ethanol, and the pulp was removed under sterile
conditions. After rinsing the dental pulp in sterile phosphate
buffered saline (PBS), the pulp was cut into 1.0 mm3
fragments in α-MEM culture medium (Gibco; Thermo Fisher Scientific,
Inc.; cat. no. BC-M-042). A total of 0.5 ml each of 0.3% type I
collagenase and 0.4% neutral protease (dispase) was added, and the
centrifuge tube containing the dental pulp and digestive fluid was
gently shaken to ensure full contact between the dental pulp and
digestive fluid. Pieces of dental pulp were digested at 37°C for 30
min until the tissue mass became loose. After which, an equal
volume of α-MEM culture medium containing 20% fetal bovine serum
(FBS; Gemini Bio Products; cat. no. 900-108) was added to terminate
digestion. The cell precipitate and tissue mass were gently blown
and beaten, centrifuged at 100 × g at 4°C for 6 min and the
supernatant was discarded. An appropriate amount of α-MEM culture
medium containing 20% FBS was added to the cell precipitate, and
gently blown and beaten until a single-cell suspension was mixed
with the loose tissue mass. The samples was inoculated into a 25
cm2 culture bottle, where it was evenly dispersed and
spread flat at the bottom of the bottle and incubated in a 37°C
cell incubator. During the first week of adherent culture, to
prevent tissue blocks from floating, moving the culture bottle was
avoided. The fluid was changed once a week before cell growth and
once every 3 days after growth. When the growth of the dental pulp
cells reached 80% confluence, the passage was performed using α-MEM
culture medium containing 20% FBS, 1% penicillin streptomycin, and
1% glutamine. The samples was cultured in a moist incubator at 37°C
with 5% CO2, and the culture medium was changed every 3
days. Third-generation dental pulp cells were used for the
subsequent experiments.
Collection of extracellular
vesicles
DPSCs were inoculated in a 10 cm culture dish. When
cell growth reached 80% density, the samples were washed with PBS
and the medium was replaced with α-MEM without FBS (Gibco; Thermo
Fisher Scientific, Inc.; cat. no. BC-M-042). After 48 h, 180 ml
supernatant was collected and centrifuged at 300 × g at 4°C for 10
min, at 2,000 × g at 4°C for 10 min and at 10,000 × g at 4°C for 30
min to remove dead cells and cell fragments. Subsequently, the
samples were filtered through a 0.22 µm filter and centrifuged at
100,000 × g for 70 min at 4°C. After removing the supernatant,
vesicles were resuspended with PBS and centrifuged again at 100,000
× g at 4°C for 70 min (26,27).
Extracellular microspheres were resuspended in PBS and frozen at
−80°C for subsequent use.
Detection of extracellular
vesicles
The following steps were performed for detecting
EVs: i) Glutaraldehyde fixation of EVs; ii) cleaning: the samples
were washed three times with 1 ml PBS and left undisturbed for 15
min each time; iii) Acid fixation: 0.5 ml of 2% acid solution was
added, and the mixture was incubated at 37°C for 2 h for fixation;
iv) Cleaning: The samples were washed three times with PBS; v)
Dehydration: Gradient dehydration with 50, 70, 80 and 90% ethanol
was used at 37°C for 15 min, and then, 100% ethanol was used for
dehydration; vi) Replacement: The mixture was replaced with 1 ml
acetone twice; vii) Impregnation: The sample was soaked in a
suitable resin (Araldite; cat. no. GY250) for electron microscopy
observation after curing; viii) Embedding: The samples were placed
in an embedding plate containing Leica embedding agent (Leica
Biosystems; cat. no. 14020108926); ix) Polymerization: The embedded
plate was polymerized at 65°C for 48 h; x) Staining: The samples
were stained with uranium dioxide acetate at 37°C for 10 min and
cleaned; then, the samples were dyed with lead acetate at 37°C for
10 min and washed; and xi) Electron microscopy examination was
performed.
EV concentration detection
EVs were collected and their concentration was
measured using a BCA protein concentration assay kit, followed by
concentration treatment. The optical density was measured using a
microplate reader (SpectraMR; Dynex) at 562 nm. A standard curve
was drawn based on the absorbance and concentration of the standard
sample, and the protein concentration was calculated.
Nanoparticles tracking analysis (NTA)
of EVs
To observe the particle size distribution,
nanoparticles tracking analysis (NTA) of the isolated DPSCs-EVs was
implemented. A total of 20 µg DPSCs-EVs were uniformly dispersed in
1 ml PBS, followed by analysis by the NanoSight nanoparticle
tracking analyzer (Malvern Panalytical, Ltd.). The sample was
agitated on a vortex mixer for 5–10 sec, a 2.5 ml syringe was used
to aspirate the sample, it was inverted 2–3 times. The samples were
slowly injected and the interface particles were observed.
Immunofluorescence confocal
microscopy
Purified P1-generation HERS epithelial cells were
obtained. HERS cells were plated in 96-well flat bottom plates at a
density of 2×105 cells/well. After treatment with
DPSC-derived EVs with a concentration of 20 µg/ml at 37°C for 24 h,
the HERS cells were washed thrice with PBS, and fixed with 4%
paraformaldehyde in PBS at 37°C for 20 min. The cells were then
permeabilized with 0.3% Triton X-100 at 37°C for 5 min. The
sections were pre-blocked with 5% bovine serum albumin
(MilliporeSigma; cat. no. A7030-5G) in PBS at 37°C for 30 min and
stained with the following primary antibodies: Anti-E-cadherin
(Proteintech Group, Inc.; cat. no. 20874-1-AP; 1:200),
anti-vimentin (Novus Biologicals, Ltd.; Bio-Techne; cat. no.
NB300-223SS; 1:200) and anti-cytokeratin 14 (CK14) antibody
(Proteintech Group, Inc.; cat. no. 10143-1-AP; 1:200) overnight at
4°C. After being washed in PBS thrice, the HERS cells were stained
with 488-conjugated AffininPure Goat Anti-Mouse IgG (H+L; Jackson
ImmunoResearch Laboratories, Inc.; cat. no. 115-545-003) and
594-conjugated AffiniPure Donkey Anti-Rabbit IgG (H+L; Jackson
ImmunoResearch Laboratories, Inc.; cat. no. 711-585-152) overnight
at 4°C followed by washing with 3X PBS. After washing, the nuclei
were stained with Heochst (Sigma-Aldrich; Merck KGaA) at 37°C for 5
min. Finally, the cells were observed using a laser-scanning
confocal microscope (Leica Microsystems GmbH).
Cell counting kit-8 (CCK-8) assay
HERS cells were plated in 96-well flat bottom plates
at a density of 2×105 cells/well and incubated with PBS
and exosomes in a medium containing 10% fetal calf serum (FCS;
Gibco; Thermo Fisher Scientific, Inc.; cat. no. 10099-141) at 37°C
in 5% CO2 in humidified air for 48 h. After which, 10 µl
CCK-8 solution (Dojindo Laboratories, Inc.) was added to each well
for 2 h. The optical density was measured using a microplate reader
(SpectraMR; Dynex) at 540 nm.
Western blotting
Protein concentrations were measured using a BCA
protein assay kit (Thermo Fisher Scientific, Inc.). A total of 100
µg of the total protein was loaded per well. The proteins in the
HERS cells were separated via 10–12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (Beijing Solarbio
Science & Technology Co, Ltd.; cat. no. P1200) and transferred
electrophoretically onto an immobilon polyvinylidene difluoride
membrane. After blocking with 10% skim milk overnight at 4°C, the
membrane was incubated for 6 h at room temperature with anti-BSP
(CST Biological Reagents Co., Ltd.; cat. no. 5468), anti-ALP
(Abcam; cat. no. ab229126), anti-RUNX2 (CST Biological Reagents
Co., Ltd.; cat. no. 12556), anti-TGF-β1 (Abcam; cat. no. ab315254),
anti-TGFR1 (Abcam; cat. no. ab235578), anti-Smad2/3 (Abcam; cat.
no. ab202445), anti-phosphorylated (p-)Smad3 (Abcam; cat. no.
ab202445), anti-MAPK (Abcam; cat. no. ab308333), anti-ERK (Abcam;
cat. no. ab184699), anti-p-ERK (Abcam; cat. no. ab314200), anti-CD9
(Abcam; cat. no. ab307085), anti-CD63 (Abcam; cat. no. ab108950),
anti-CD81 (Abcam; cat. no. ab155760), anti-TSG101 (Abcam; cat. no.
ab125011), anti-Calnexin (Abcam; cat. no. ab22595) and anti-GAPDH
(Proteintech Group, Inc.; cat. no. 6004-1-1g) antibodies (all
1:1,000). The TBST buffer (TBS buffer containing 0.05% Tween-20)
was prepared and the membrane was washed three times with 1X TBST.
After which, the membrane was incubated with HRP-labeled Goat
Anti-Rabbit IgG H&L antibody (Abcam; cat. no. ab205718;
1:2,000) for 1 h at room temperature. The membrane was washed again
three times with TBST and developed via an enhanced
chemiluminescence (ECL) plus chemiluminescence kit (Tanon Science
& Technology, Co., Ltd.; cat. no. 180-501). Protein bands were
detected using an Odyssey System (LI-COR Biosciences).
ALP staining
The aforementioned grouped cells were cultured at
37°C in 5% CO2 in humidified air for 48 h, the
supernatant was discarded, washed twice with PBS, and fixed with
70% alcohol at 37°C for 2 min. Distilled water was used to remove
the fixative. Using alkaline phosphatase calcium cobalt staining
kit (Jiangsu Kaiji Biotechnology, Co., Ltd.; cat. no. KGA353),
incubation solution A and B were mixed to form the incubation
working solution before use. The sample was incubated with the
incubation solution at 37°C for 3 h. The sample was then rinsed
with water for 5–10 min. After which, the sample was incubated with
staining solution A at room temperature for 5 min and rinsed with
water for 2 min. Then, the sample was incubated with staining
solution B at room temperature for 1 min and rinsed again for 5
min. Images were captured using a biological inverted microscope
(Olympus Soft Imaging Solutions GmbH; cat. no. IX71).
Alizarin Red staining
determination
The medium was changed every 3 days, and after 14
and 21 days the culture medium was aspirated. The samples was
washed twice with PBS, fixed with 4% paraformaldehyde for 30 min at
37°C and washed with deionized water. After which, the sample was
stained with 2% Alizarin Red staining solution for 20–30 min at
37°C, and images were captured with a fluorescence microscope.
After which, decolorization treatment was performed, whereby 200 µl
5% perchloric acid solution was added to each well, the sample was
gently agitated on a shaker for 15 min and then transferred with
150 µl decolorization solution to a 96 well plate. The sample was
read with an enzyme-linked immunosorbent assay reader, the OD value
was recorded and detected at a wavelength of 490 nm.
Flow cytometry analysis
Cells were seeded in a 96-well culture plate, and
the number of cells in each well was adjusted to 1×105
using 10% FCS-RPMI 1640 medium (Thermo Fisher Scientific, Inc.;
cat. no. 22400089), with a final volume of 200 µl/well. A portion
from each group of cells was extracted for flow cytometry detection
of CD44, CD34, CD45 and CD90. The cells were washed with pre-cooled
PBS and resuspended in 100 µl PBS. Anti-CD44-PE (Abcam; cat. no.
ab23396), anti-CD34-PE (Abcam; cat. no. ab223930), anti-CD45-PE
(eBioscience; cat. no. 12-0451-83) and anti CD90-PE (Abcam; cat.
no. ab33694) was added to the samples and incubated at 4°C in the
dark for 30 min. The cells were washed twice with 2 ml 1X
transmembrane buffer, suspended in PBS and detected by flow
cytometry (BD Biosciences, C6 model), and analyzed by the
Cytek® NL-CLC full spectrum flow cytometer.
Transmission electron microscopy
(TEM)
The isolated exosomes were evaluated using
transmission electron microscopy (TEM). Firstly, 10 µl of each
sample was dropped onto a 200 mesh ultra-thin carbon coated copper
grid at 37°C for 2 min. After which, the grid was quickly dried on
filter paper. Negative staining was performed on the grid using 1%
uranyl acetate (filtered twice through a 0.22 µm filter). After
complete drying for 1 min, images were captured using Hitachi
HT-7700 TEM at a voltage of 100 kV.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Third-generation HERD cells were seeded in 6-well
plates at a density of 1×104 cells/well and co-cultured
at 37°C with osteogenic medium (ScienCell Research Laboratories,
Inc.; cat. no. YB4601) and scaffolds for 14 and 21 days to obtain
cell pellets. Total RNA was extracted with TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and cDNA was
synthesized using the PrimeScript RT Master Mix Perfect Real-Time
Kit at 37°C (Takara Biotechnology, Co., Ltd.). The relative gene
expression level was normalized with the internal parameter
(GAPDH). The reaction procedure was: 30 sec at 95°C, and 40 cycles
of 5 sec at 95°C, 30 sec at 60°C. SYBR Select MasterMix (Takara)
was used for q-PCR assay to detect mRNA levels of ALP, BSP, RUNX2,
TGF-β1, TGFR1, Smad2/3, MAPK and ERK1/2. All primers used in the
present study were purchased from GeneChem, Inc., (Table I).
 | Table I.Primer sequences for RT-qPCR. |
Table I.
Primer sequences for RT-qPCR.
| Primer | Sequence
(5′-3′) |
|---|
| Rattus GAPDH_F |
GCTGAGTATGTCGTGGAGTC |
| Rattus GAPDH_R |
GATGCATTGCTGACAATCTT |
| Rattus BSP_F |
CGGCCACGCTACTTTCTTTA |
| Rattus BSP_R |
CTCCAACTTTCCAGCGTCA |
| Rattus RUNX2_F |
CCTATGACCAGTCTTACCCC |
| Rattus RUNX2_R |
CAGAGGCAGAAGTCAGAGGT |
| Rattus ALP_F |
CGGCTATGAACAGTGTGATG |
| Rattus ALP_R |
ACCGCGTTCCAGACAGTAG |
| Rattus
TGF-β1_F |
CGCCTGCAGAGATTCAAGT |
| Rattus
TGF-β1_R |
CTCGACGTTTGGGACTGAT |
| Rattus
ERK1/2_F |
GAACATCATCGGCATCAATG |
| Rattus
ERK1/2_R |
TGAGGTCACGGTGCAGAA |
| Rattus p38
MAPK_F |
ACATCGTGTGGCAGTGAAGA |
| Rattus p38
MAPK_R |
ACGTGGTCATCGGTAAGCTT |
| Rattus Smad3_F |
GGCTACCTGAGTGAAGATGGA |
| Rattus Smad3_R |
GGCTGTAGGTCCAAGTTATTG |
| Rattus
TGFβR1_F |
TCACTAGATCGCCCTTTCAT |
| Rattus
TGFβR1_R |
TCGCCAAACTTCTCCAAAC |
Micro-CT analysis
SD rats in each group (n=6) were anesthetized with
3% pentobarbital sodium (50 mg/kg; intraperitoneally). After which,
a micro-CT system was used to calculate the bone mineral density
(BMD) and the bone volume/total volume (BV/TV) ratio of regenerated
bone in the calvarial defect. The CT Analyzer (version 1.20.3.0;
Bruker Corporation) was used to analyze the region of interest.
Unified parameters were set to calculate the TV, BV, BV/TV, bone
area (BS), number of trabeculae (Tb. N) and thickness of trabeculae
(Tb. th) of the tissue. The micro-CT equipment was set with a
tomographic rotation of 180° at 85 kV and 135 mA.
Statistical analysis
Statistical analyses were performed using the
GraphPad Prism 9.0 software (Dotmatics). The data were presented as
the mean ± standard deviation (SD). Data were consistent with a
normal distribution based on the Shapiro-Wilk test. One-way ANOVA
combined with Tukey's post-hoc test were employed for the
comparison of statistical difference. P<0.05. was considered to
indicate a statistically significant difference.
Results
Identification of DPSC-EVs and
validation of uptake experiments
First, the morphology of DPSCs was observed and
first generation DPSCs cells (P1) were shown to be short and
irregularly shuttle-shaped, while the third generation DPSCs cells
(P3) appeared to be long and regularly shuttle-shaped (Fig. 1A). DPSCs and DPSC-EVs were
isolated. The results of the flow cytometry assays with DPSC
markers showed positive expression of CD44 and CD90 and negative
expression of CD34 and CD45 (Fig.
1B). Alizarin red, ALP and Oil red O staining in DPSCs was also
performed and the results showed that Alizarin red staining could
clearly display calcified plaques (Fig. 1C) and ALP was expressed in DPSCs.
Next, the morphology of the EVs via TEM was observed (Fig. 1D), and the NTA particle size was
determined to be 115 nm (Fig. 1E).
The results of the western blotting analysis revealed that the
extracellular vesicle markers CD9, CD63, TSG101 and CD81 were
expressed, whereas, the DPSC marker calnexin was not expressed
(Fig. 1E).
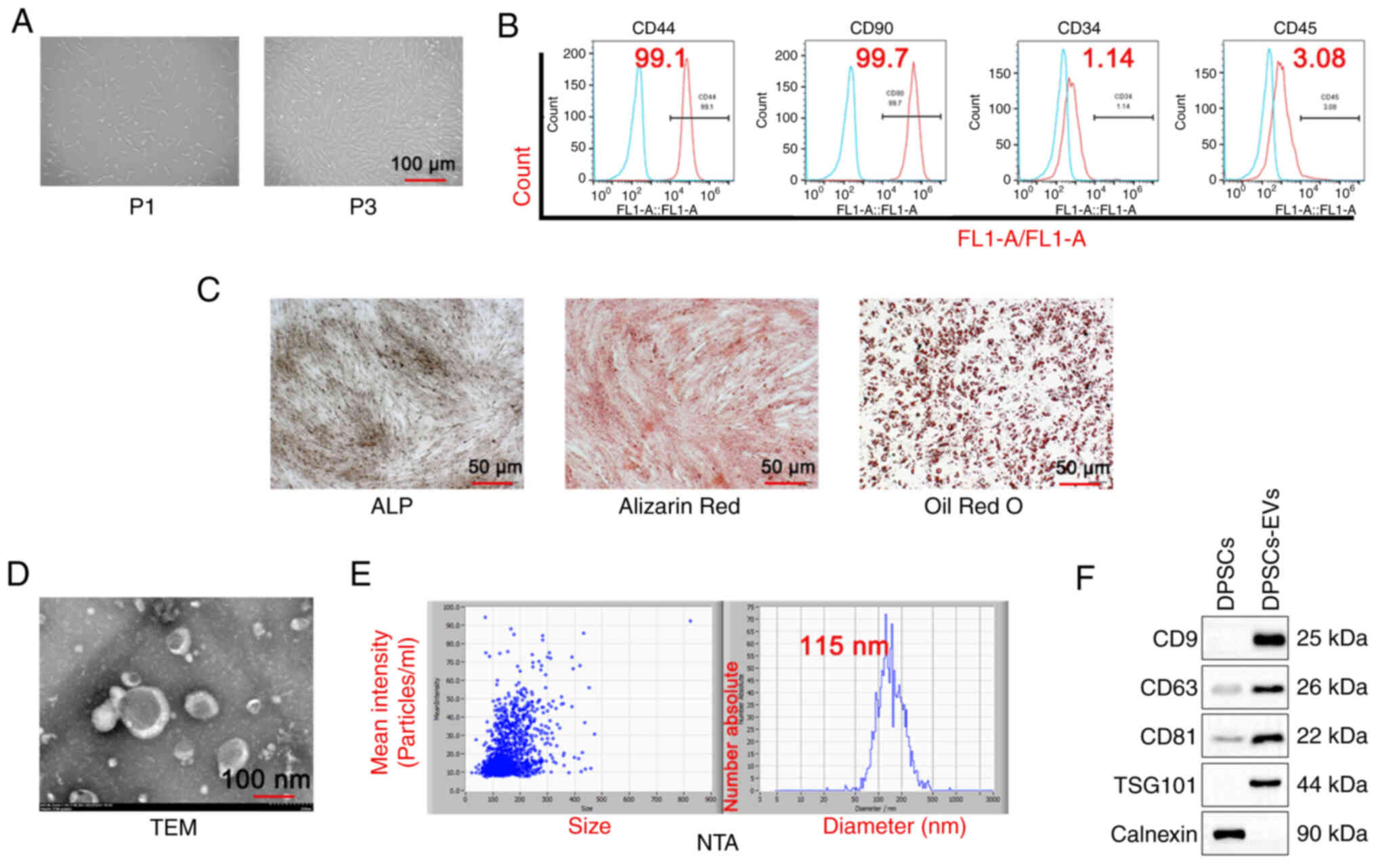 | Figure 1.Isolation and identification of DPSC
and DPSC-EVs. (A) DPSCs were isolated, cultured and observed under
a microscope. (B) Flow cytometry was performed to detect DPSC
markers (CD44, CD90, CD34 and CD45). (C) ALP staining, Alizarin red
staining and Oil Red O staining were performed to evaluate the
osteogenic capacity of DPSCs. (D) The morphological features of
DPSC-EVs were observed. The average size of EVs was 115 nm. (E) The
NTA particle size of DPSC-EVs was analyzed. (F) Western blotting
was performed to detect DPSC-EV markers (CD9, CD63, CD81, TSG101
and calnexin). Triplicate independent experiments were used. DPSC,
dental pulp stem cell; EV, extracellular vesicle; NTA,
nanoparticles tracking analysis; FL1, green fluorescent channel;
P1,first generation DPSCs cells; P3, third generation DPSCs cells;
TSG101,tumor susceptibility gene 101; TEM, transmission electron
microscopy. |
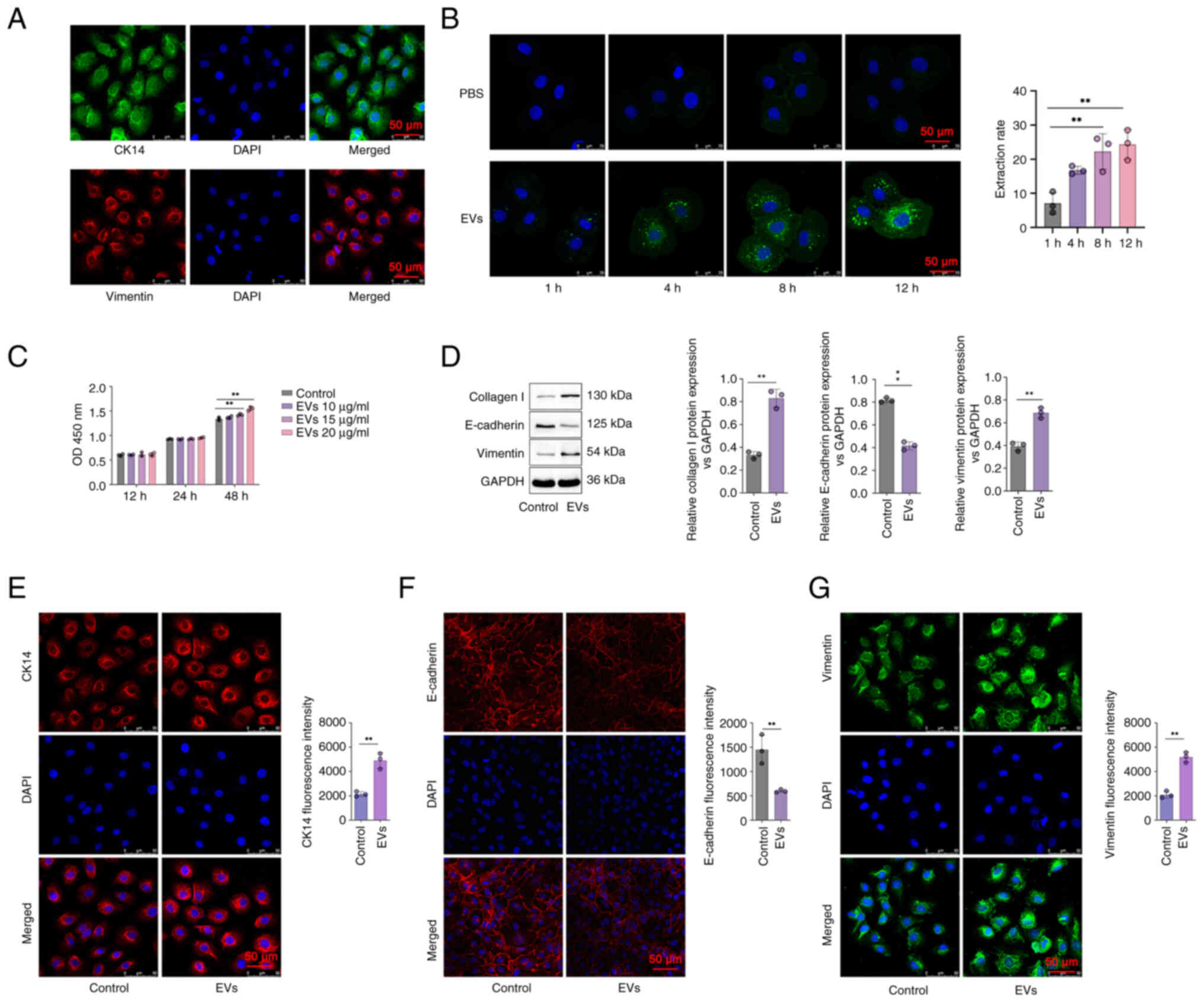
The HERS cells were isolated and co-cultured with
DPSC-EVs, and immunofluorescence staining was used to detect
epithelial cell markers, including CK14 and Vimentin.
Immunofluorescence staining showed CK14 and Vimentin were expressed
in the HERS cells (Fig. 2A). The
uptake of EVs was assessed via confocal microscopy. The EVs were
stained with green fluorescence. The range of green fluorescence
staining agents in the EVs in the DPSC-EV group increased, and the
color of green fluorescence became increasingly intense, indicating
that the number of EVs taken up increased with time. A comparison
between the PBS and EV groups revealed nearly no visible exocrine
uptake between these two groups at 1 h. Thus, the uptake of EVs
commenced after 1 h (P<0.05; Fig.
2B). The difference in the quantity of secretions ingested
between the two groups increased over time (Fig. 2B). Next, different concentrations
(10, 15 and 20 µg/ml) of DPSC-EVs were used to stimulate HERS cells
for 12, 24 and 48 h. The results showed that with a concentration
of 20 µg/ml at 48 h, compared with control, the cell proliferation
activity of HERS increased significantly after treatment with
DPSC-EVs (P<0.05; Fig. 2C).
DPSC-EVs promote EMT and osteogenic
differentiation of HERS cells
To further evaluate the ability of DPSC-EVs to
promote the osteogenic differentiation of HERS cells, changes in
epithelial and mesenchymal cell markers in HERS cells were
detected. After 24 h of coculture with DPSC-EVs and HERS cells, the
expression of the epithelial cell marker E-cadherin in HERS cells
decreased, whereas the expression of the mesenchymal cell marker
vimentin and type I collagen increased compared with control group
(P<0.05; Fig. 2D).
Immunofluorescence staining showed that CK14 expression was
increased in the DPSC-EVs group compared with control group
(P<0.05; Fig. 2E), the
expression of E-cadherin in HERS cells decreased in the DPSC-EVs
group compared with control group (P<0.05; Fig. 2F) and the expression of vimentin in
HERS cells increased in the DPSC-EVs group compared with control
group (P<0.05; Fig. 2G). These
findings indicated that HERS cells simultaneously express
epithelial and mesenchymal cell markers, with dual characteristics
of epithelial and mesenchymal cells, promoting
epithelial-to-mesenchymal cell transformation after being
stimulated by DPSC-EVs. Therefore, after treatment with DPSC-EVs,
HERS cells may differentiate into cementoblasts through EMT.
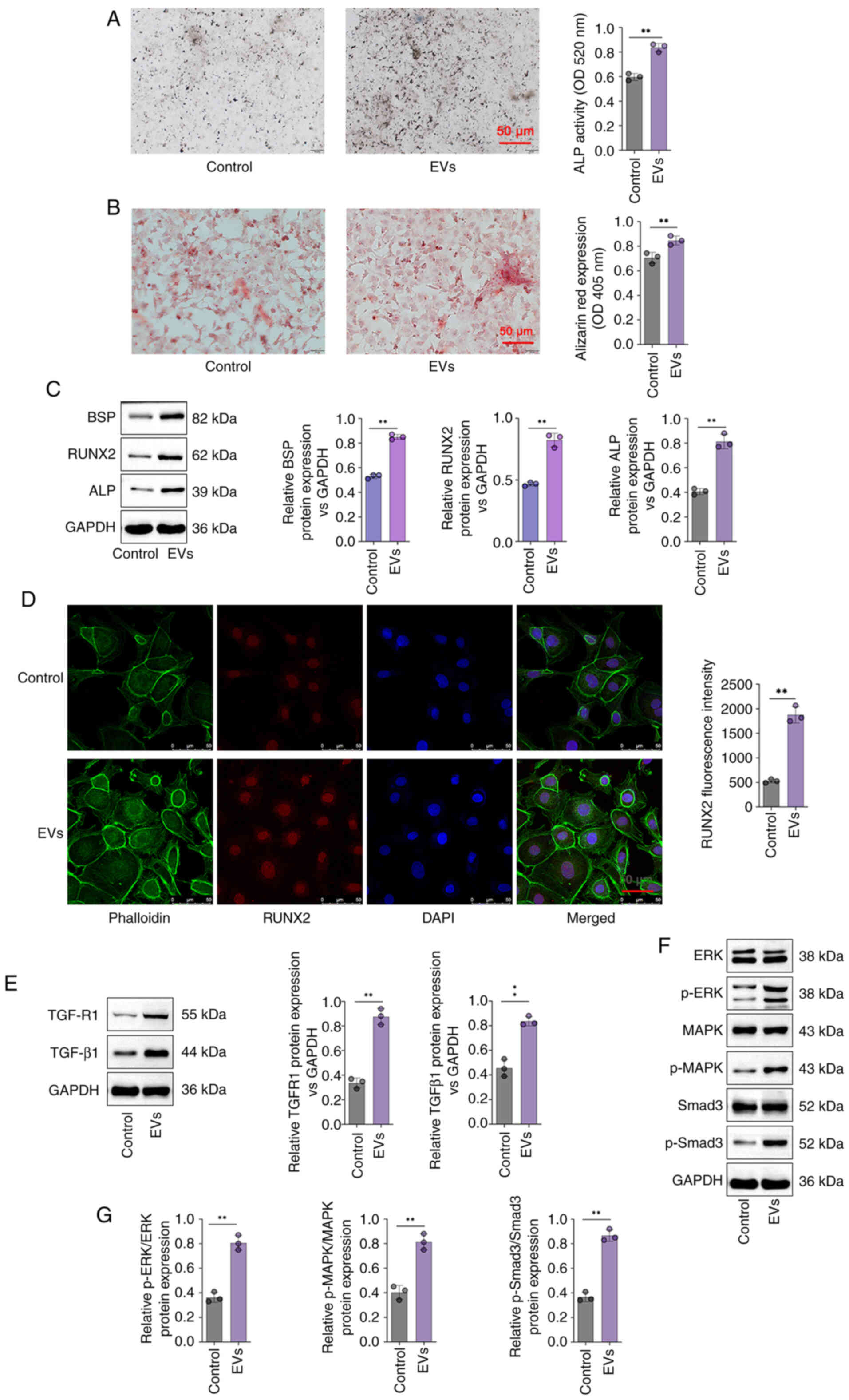
Furthermore, ALP activity was significantly greater
in the DPSC-EV group compared with control group (P<0.05;
Fig. 3A). Cell mineralization
determined via Alizarin red staining revealed that the Alizarin red
content in the DPSC-EV groups was significantly greater than that
in the control group (P<0.05; Fig.
3B). In addition, compared with control group, the expressions
of BSP, ALP and RUNX2 were increased significantly in the cells of
the DPSC-EV (HERS) group (P<0.05; Fig. 3C). mRNA levels of ALP (Fig. S1A), RUNX2 (Fig. S1B), BSP (Fig. S1C), TGFR1 (Fig. S1D), TGF-β1 (Fig. S1E), Smad3 (Fig. S1F), ERK (Fig. S1G) and MAPK (Fig. S1H) were increased significantly in
the cells of the DPSC-EV (HERS) group compared with control group.
Furthermore, RUNX2 expression in HERS cells was also studied via
RUNX2 staining. DPSC-EVs promoted the expression of RUNX2, thus
promoting osteogenic differentiation (P<0.05; Fig. 3D). Therefore, following
DPSC-EV-induction, HERS cells retained osteogenic potential, which
indicated that HERS cells can be used for cementum
regeneration.
DPSC-EV-induced osteogenic
differentiation is regulated by TGF-β1/ERK signaling
The involvement of TGF-β1 in DPSC-EV-mediated
osteogenic differentiation was further elucidated. In the present
study, the protein levels of TGF-β1, TGFR1, p-Smad3 and MAPK/ERK
were determined via Western blotting. The findings revealed that
the expression of TGFR1 and TGF-β1were significantly increased in
DPSC-EV-treated cells compared with control group (P<0.05;
Fig. 3E). Relative expression of
p-Smad3/Smad3, ERK/p-ERK and MAPK/p-MAPK were significantly
increased in DPSC-EV-treated cells compared with control group
(P<0.05; Fig. 3F and G). Using
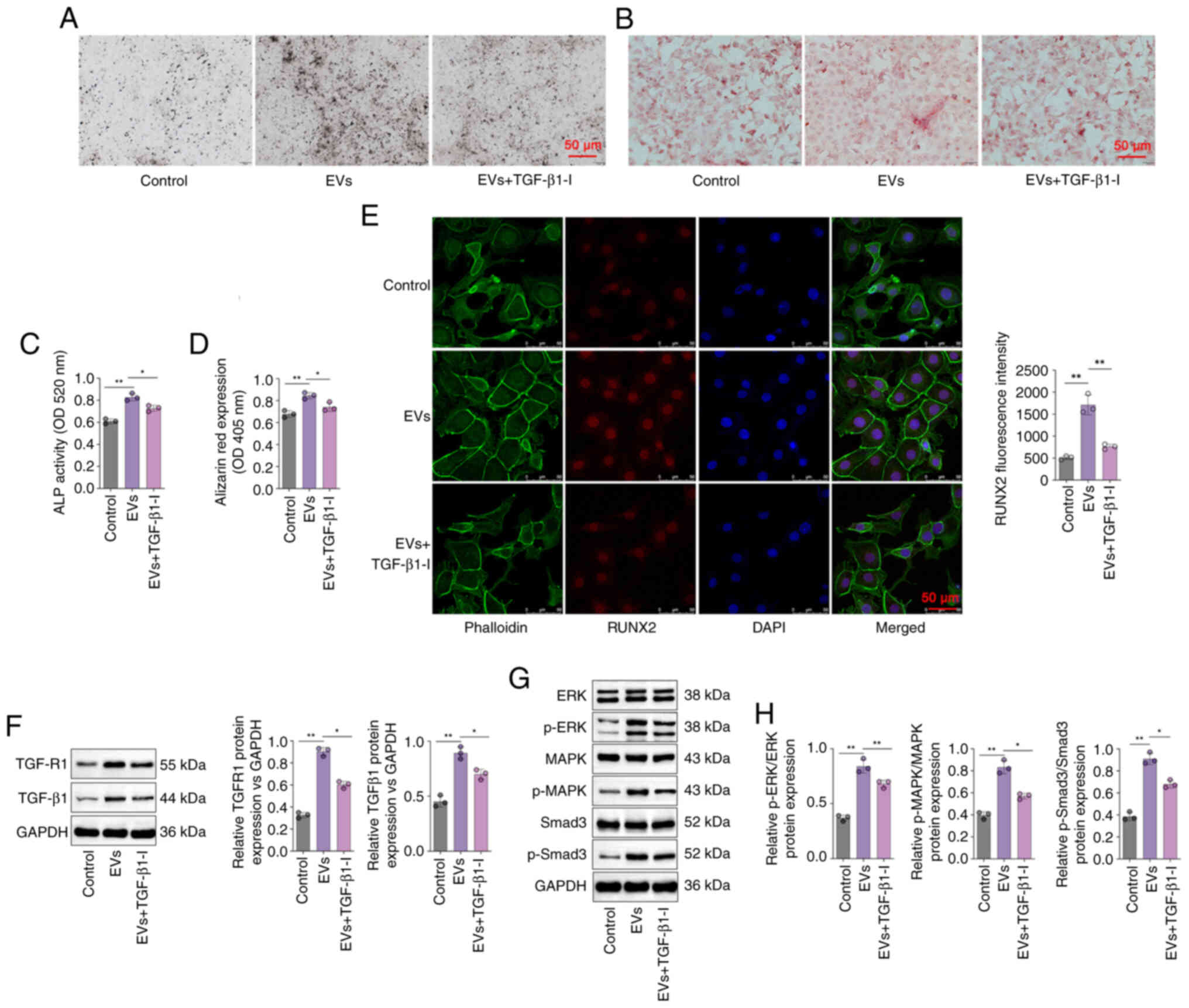
a specific inhibitor of TGF-β1, it was found that ALP expression in
the DPSC-EVs + TGF-β1inhibitor group was lower than that in the
DPSC-EV group (Fig. 4A), and ALP
activity was also significantly lower in the DPSC-EV + TGF-β1
inhibitor group compared with DPSC-EV group (P<0.05; Fig. 4B). Using a specific inhibitor of
TGF-β1, the Alizarin red content in the DPSC-EVs + TGF-β1 inhibitor
group was lower than that in the DPSC-EV group (Fig. 4C), and Alizarin red expression was
also significantly lower in the DPSC-EV + TGF-β1 inhibitor group
compared with the DPSC-EV group (P<0.05; Fig. 4D). The confocal microscopy results
revealed that TGF-β1 inhibition weakened the positive effect of
DPSC-EVs on the expression of RUNX2 (P<0.05; Fig. 4E). It was also found that treatment
with a TGF-β1 inhibitor reversed the increase in the expression of
TGFR1 and TGF-β1 induced by DPSC-EVs (P<0.05; Fig. 4F). The levels of MAPK, ERK, Smad3
and p-Smad3 were detected by Western blotting (Fig. 4G), and were increased in the
DPSC-EVs group compared with control group, and consistently lower
in the TGF-β1 inhibitor-treated group than in the DPSC-EVs group
(all P<0.05, Fig. 4H).
Furthermore, mRNA levels of TGF-β1 (Fig. S2A), TGFR1 (Fig. S2B), Smad3 (Fig. S2C), MAPK (Fig. S2D) and ERK (Fig. S2E) in the TGF-β1 inhibitor-treated
group were markedly decreased compared with the EVs only group.
DPSC-EV-loaded hydrogels accelerate
bone regeneration in rats with alveolar bone defects
After establishing the alveolar bone defect rat
model, micro-CT examinations were first performed to observe bone
regeneration in each rat group showing bone regeneration in rats
with alveolar bone defects after injecting DPSC-EVs and
DPSC-EV-loaded hydrogel (Fig. 5A).
New bone grew concentrically in the DPSC-EV and DPSC-EV-loaded
hydrogel groups, with greater bone mass in these groups than in the
blank control group, and the BV/TV increased significantly compared
with control group (Fig. 5B).
Furthermore, the effects of DPSC-EVs, DPSC-EV-loaded hydrogel or
DPSC-EV-loaded hydrogel and TGF-β1i on osteogenic differentiation
in alveolar bone defect model rats were also evaluated using
Western blotting (Fig. 5C). The
levels of TGF-β1 and p-Smad3/Smad3 were significantly greater in
the DPSC-EV-loaded hydrogel group than in the DPSC-EV group; thus,
treatment with the TGF-β1 inhibitor decreased the expression of
TGF-β1 and p-Smad3/Smad3 (P<0.05; Fig. 5D). Expressions of BSP, ALP and
RUNX2 were significantly greater in the DPSC-EV-loaded hydrogel
group than in the DPSC-EV group, and the expression of BSP, ALP and
RUNX2 were decreased upon treatment with the TGF-β1 inhibitor
(P<0.05; Fig. 5E). mRNA levels
of ALP (Fig. S3A), BSP (Fig. S3B), RUNX2 (Fig. S3C), TGF-β1 (Fig. S3D) and Smad3 (Fig. S3E) were significantly increased in
the DPSC-EV-loaded hydrogel group than in the DPSC-EV group, but
were decreased in the TGF-β1 inhibitor group.
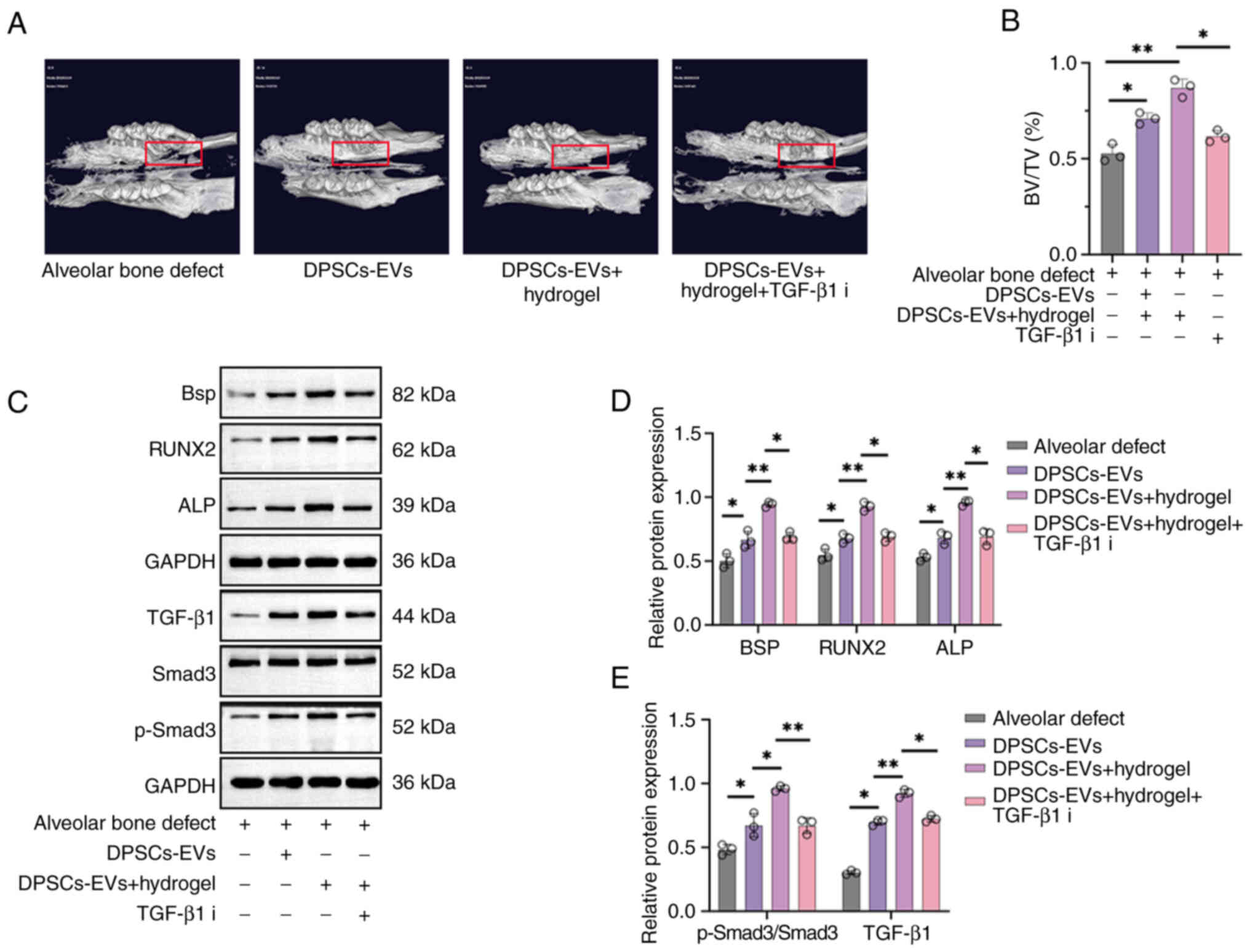 | Figure 5.DPSC-EV-loaded hydrogels accelerated
bone regeneration in rats with alveolar bone defects. (A) A rat
model of alveolar bone defects was established, and the rats were
treated with DPSC-EVs. The rats were euthanized after the
operation, and new bone regeneration was less intense in the
control group and hydrogel group. More new bones were found in the
hydrogel + DPSCs-EVs group. (B) Similarly, the BV/TV results showed
that the hydrogel + DPSCs-EV group formed more new bones than the
other groups. (C) The expression of osteogenesis-related proteins
(BSP, ALP and RUNX2) and TGF-β1/Smad signaling were determined via
Western blotting. (D) Expressions of TGF-β1 and p-Smad3/Smad3 were
significantly greater in the DPSC-EV-loaded hydrogel group than in
the DPSC-EV group. (E) Expressions of BSP, ALP and RUNX2 were
significantly greater in the DPSC-EV-loaded hydrogel group than in
the DPSC-EV group. The data from three independent experiments are
presented as the mean ± SD; *P<0.05 and **P<0.01. DPSC,
dental pulp stem cell; EV, extracellular vesicle; HERS, Hertwig's
epithelial root sheath; p, phosphorylated; RUNX2, runt-related
transcription factor 2; ALP, alkaline phosphatase; i, inhibitor;
BV/TV, bone volume/total volume; BSP, bone sialoprotein. |
Discussion
Bone tissue engineering is an interdisciplinary
field that combines the application of osteoblasts (stem
cells/progenitors), bioactive molecules and biocompatible
scaffolds, as well as, stem cells and signaling molecules (28). The key factors involved in bone
tissue engineering are cells, scaffolds and signaling molecules
(29). Similar to other bone
tissues, osteoclasts and osteoblasts are the primary factors
involved in bone formation and bone homeostasis in alveolar bone
(30,31). Additionally, a complex cellular
communication network plays a key role in maintaining bone coupling
and alveolar bone homeostasis (32). However, the invasion of pathogens
from the oral environment or blood transmission, orthodontic
treatment, drugs and mechanical stress caused by systemic
pathological factors induce complex inflammation, which in turn
activates osteoclasts and inhibits osteoblasts, thus disrupting the
balance between bone removal and regeneration, leading to alveolar
bone loss (33–36). In the present study, rat alveolar
bone defects model were used and the surgery was carried out as
described previously (25).
Animals were sacrificed from each group and tissue samples were
collected for micro-CT analysis. It was found that DPSC-EVs were
highly biocompatible and effectively promoted the proliferation and
migration of HERS cells when an appropriate number of DPSC-EVs was
used. The DPSC-EV-loaded hydrogel promoted osteogenic and
osteoinductive effects in alveolar bone defects in rats.
The present study investigated methods for promoting
the interaction between DPSC-EVs and HERS cells. It was found that
DPSC-EVs created a dimensional environment conducive to cell
growth. The high cell viability in the DPSC-EV culture indicated
that DPSC-EVs were biocompatible. DPSC-EVs also promoted the
differentiation of HERS cells via EMT. Bone formation is a complex
process, and the expression of genes closely related to bone
formation in osteoblasts is necessary for the differentiation of
osteoblasts into osteoblasts (37); such genes are called bone-related
genes (38). During in vivo
bone formation, bone-related gene expression has a strictly
regulated time sequence (39). The
orderly expression of bone-related genes is crucial for the bone
metabolism of osteoblasts, osteogenesis, bone growth and bone
reconstruction (40). ALP is
produced in the early stages of cell development and is commonly
found on cell surfaces and in matrix vesicles of bone and calcified
cartilage (41). Although some
osteocalcin genes are upregulated, ALP expression decreases
(42). The expression of RUNX2
marks the onset of osteoblast differentiation and promotes the
early maturation and differentiation of osteoblasts (43). Therefore, RUNX2 is the earliest and
most specific marker of the bone formation process and is generally
highly expressed in the early stage of bone cell differentiation
(44). BSP, a critical marker
protein, is highly expressed in the middle and late stages of
osteoblast differentiation (44).
The results of the present study suggested that ALP, BSP and RUNX2
expression increased considerably in HERS cultured with DPSC-EVs,
suggesting that DPSC-EVs promote osteogenic differentiation in HERS
cells at an early stage. The results in the present study are
consistent with previous studies, which indicated that HERS could
differentiate into cementum cells through EMT and secrete cementum
to form periodontal tissue (8,9).
However, in the present study the role and regulatory mechanism of
DPSC-EVs in the promotion of osteogenic differentiation of HERS
cells was also investigated.
The findings of the present study revealed that EVs
are crucial regulators of EMT and are essential for bone
development and regeneration. EVs play a dual role in the
aforementioned processes, activating endothelial cells to
accelerate their migration and proliferation and stimulating
osteogenesis by regulating osteogenic growth factors (45). EVs promote vascular invasion and
the recruitment of fragmented cartilage tissue to hypertrophic
cartilage (46). Thus, the results
of the present study indicated that EMT and osteogenesis are linked
and must be tightly coupled for the physiological function of
bones.
Although EV treatment has shown promising outcomes,
the exact underlying mechanism remains unclear. Several studies
have reported that TGF-β1 regulates various functions in normal
tissue homeostasis, such as cellular differentiation, apoptosis,
cell cycle arrest and cellular migration (47–49).
TGF-β1 binds to type I receptors and forms a heteromeric complex
with type II receptors (50). This
complex activates intracellular Smad transcription factors to
mediate downstream signaling events, which facilitates the movement
of the complex into the nucleus, where it regulates the expression
of its target genes (50). The
in vitro results of the present study showed that DPSC-EVs
mediate HERS cell functions through the cell surface presentation
of TGF-β1 to TGF-βR on target cells. Furthermore, with Smad2 and
Smad3 phosphorylation, the phosphorylated intermediate is
associated with a co-Smad in the cytoplasm, which migrates to the
nucleus, where transcription is regulated through direct DNA
binding by the Smad complex. Thereafter, the expression of Smad2/3
and p-Smad2/3 in HERS cells was determined, and p-Smad2/3 and
MAPK/ERK expression increased significantly following DPSC-EV
treatment, whereas inhibiting TGF-β1 expression in HERS cells
decreased p-Smad expression. These findings suggested that the
combination of TGF-β1 on the surface of HERS cells activates
downstream signal transducers of TGF-β1, including Smad2/3.
Furthermore, these findings suggested that DPSC-EVs modulate
osteogenesis via TGF-β1/ERK signaling. Micro-CT examinations were
performed to observe bone regeneration in each rat group after
injecting DPSC-EVs, DPSC-EV-loaded hydrogels or DPSC-EV-loaded
hydrogels with TGF-β1i. New bone grew concentrically in the DPSC-EV
or DPSC-EV-loaded hydrogel groups, with greater bone mass and
BV/TV. In vivo, the expression of osteogenesis-related
proteins was determined, and the expression of BSP, ALP and RUNX2
increased significantly following treatment with DPSC-EVs or
DPSC-EV-loaded hydrogels, indicating that DPSC-EVs or
DPSC-EV-loaded hydrogels might promote osteogenesis after alveolar
bone defects. The present study had several limitations, including
that the role of cytokines in EVs was not verified, and in
vivo new bone regeneration and BMD were not be detected via
histological examinations.
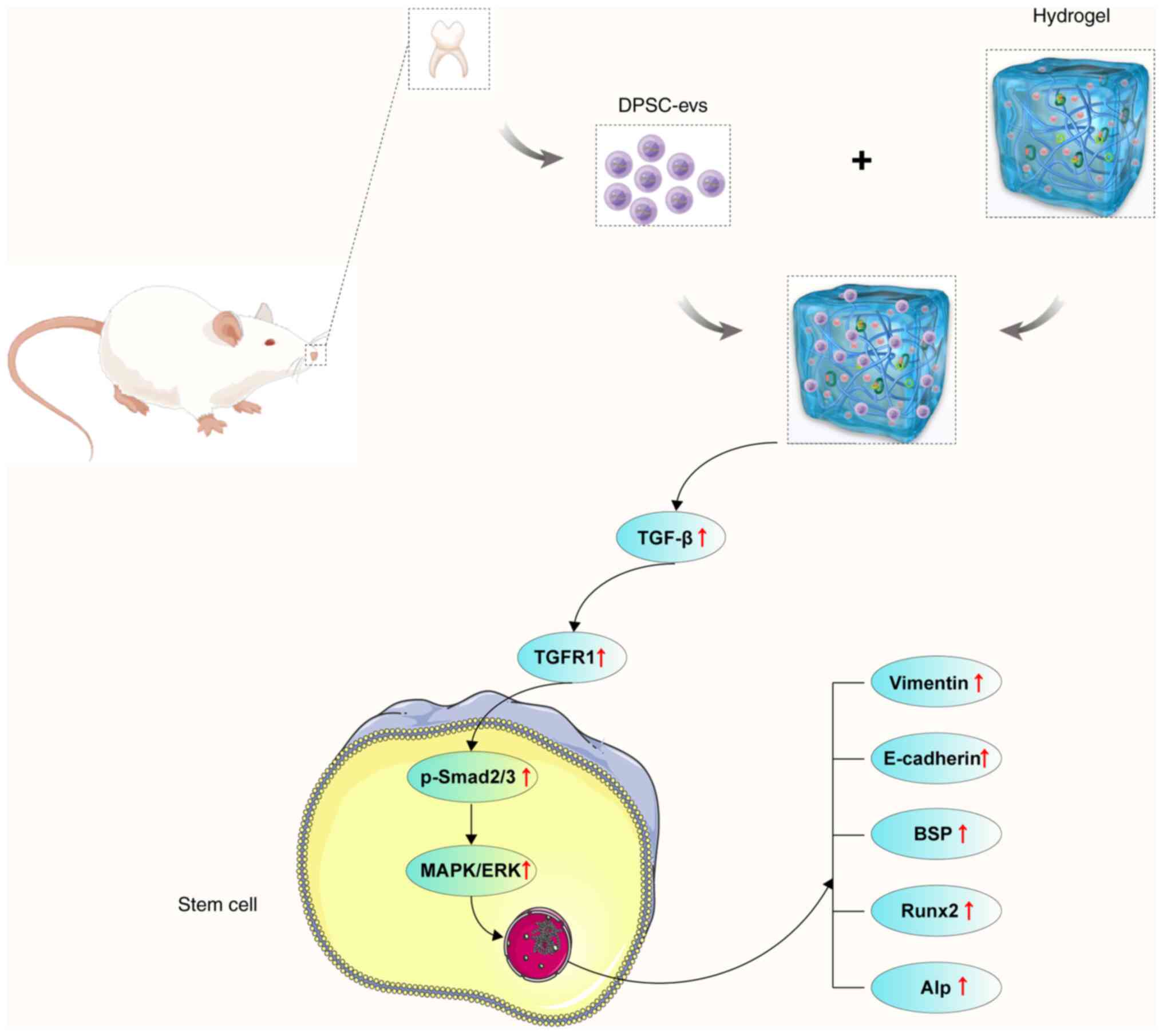
To summarize, the findings of the present study
revealed that DPSC-EVs promote EMT and osteogenesis. DPSC-EVs
promote the proliferation and EMT of HERS cells. The cell viability
and proliferation results indicated that DPSC-EVs provide suitable
conditions for HERS cells. Additionally, DPSC-EVs affect osteogenic
differentiation through the TGF-β1/ERK signaling pathway. The
DPSC-EVs and the DPSC-EV-loaded hydrogels greatly promoted
osteogenesis in alveolar bone defect model rats. These findings
suggested that DPSC-EVs or DPSC-EV-loaded hydrogels regulate
osteogenesis, which might be a key therapeutic target for alveolar
bone defects (Fig. 6).
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Beijing Nova Program
(grant no. 20220484231) and China Higher Education Innovation Fund
(grant no. 2021JH039).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
XH, XYC, KY and DLZ conceived and designed the
study. XH and XC performed the experiments, collected and analyzed
the data, produced the figures and wrote the manuscript. XC, YLX,
ZLL, YL and CYG performed bioinformatics and statistical analysis.
YYL and KY revised the manuscript. KY and DLZ confirm the
authenticity of all the raw data. All authors read and approved the
final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by The Animal Welfare
Ethics Committee of Beijing MDKN Biotechnology Co., LTD. (Beijing,
China; approval no. MDKN-2022-052).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hollý D, Klein M, Mazreku M, Zamborský R,
Polák Š, Danišovič Ľ and Csöbönyeiová M: Stem cells and their
derivatives-implications for alveolar bone regeneration: A
comprehensive review. Int J Mol Sci. 22:117462021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pervizaj-Oruqaj L, Selvakumar B, Ferrero
MR, Heiner M, Malainou C, Glaser RD, Wilhelm J, Bartkuhn M, Weiss
A, Alexopoulos I, et al: Alveolar macrophage-expressed Plet1 is a
driver of lung epithelial repair after viral pneumonia. Nat Commun.
15:872024. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu X, Yu S, Chen G, Zheng W, Peng J, Huang
X and Chen L: Insight into the roles of melatonin in bone tissue
and bone-related diseases (Review). Int J Mol Med. 47:822021.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baldwin P, Li DJ, Auston DA, Mir HS, Yoon
RS and Koval KJ: Autograft, allograft, and bone graft substitutes:
Clinical evidence and indications for use in the setting of
orthopaedic trauma surgery. J Orthop Trauma. 33:203–213. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang HD and Zhang YZ: Editorial
commentary: Reduce the failure risk: A challenge to reduce the risk
of using hybrid graft in anterior cruciate ligament reconstruction.
Arthroscopy. 34:2936–2938. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lei F, Li M, Lin T, Zhou H, Wang F and Su
X: Treatment of inflammatory bone loss in periodontitis by stem
cell-derived exosomes. Acta Biomater. 141:333–343. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li X, Zhang S, Zhang Z, Guo W, Chen G and
Tian W: Development of immortalized Hertwig's epithelial root
sheath cell lines for cementum and dentin regeneration. Stem Cell
Res Ther. 10:32019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ouchi T and Nakagawa T: Mesenchymal stem
cell-based tissue regeneration therapies for periodontitis. Regen
Ther. 14:72–78. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park KR, Kim S, Cho M and Yun HM: Limonoid
triterpene, obacunone increases runt-related transcription factor 2
to promote osteoblast differentiation and function. Int J Mol Sci.
22:24832021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bi F, Tang H, Zhang Z, Lyu Y, Huo F, Chen
G and Guo W: Hertwig's epithelial root sheath cells show potential
for periodontal complex regeneration. J Periodontol. 94:263–276.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang S, Yang Y, Jia S, Chen H, Duan Y, Li
X, Wang S, Wang T, Lyu Y, Chen G and Tian W: Exosome-like vesicles
derived from Hertwig's epithelial root sheath cells promote the
regeneration of dentin-pulp tissue. Theranostics. 10:5914–5931.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gupta D, Zickler AM and El Andaloussi S:
Dosing extracellular vesicles. Adv Drug Deliv Rev. 178:1139612021.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harrell CR, Jovicic N, Djonov V,
Arsenijevic N and Volarevic V: Mesenchymal stem cell-derived
exosomes and other extracellular vesicles as new remedies in the
therapy of inflammatory diseases. Cells. 8:16052019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mattei V and Delle Monache S: Dental pulp
stem cells (DPSCs) and tissue regeneration: Mechanisms mediated by
direct, paracrine, or autocrine effects. Biomedicines. 11:3862023.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mattei V, Martellucci S, Pulcini F,
Santilli F, Sorice M and Delle Monache S: Regenerative potential of
DPSCs and revascularization: direct, paracrine or autocrine effect?
Stem Cell Rev Rep. 17:1635–1646. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang X and Thomsen P: Mesenchymal stem
cell-derived small extracellular vesicles and bone regeneration.
Basic Clin Pharmacol Toxicol. 128:18–36. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang S, Chuah SJ, Lai RC, Hui JHP, Lim SK
and Toh WS: MSC exosomes mediate cartilage repair by enhancing
proliferation, attenuating apoptosis and modulating immune
reactivity. Biomaterials. 156:16–27. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hade MD, Suire CN and Suo Z: Mesenchymal
stem cell-derived exosomes: applications in regenerative medicin.
Cells. 10:19592021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Otsuru S, Desbourdes L, Guess AJ, Hofmann
TJ, Relation T, Kaito T, Dominici M, Iwamoto M and Horwitz EM:
Extracellular vesicles released from mesenchymal stromal cells
stimulate bone growth in osteogenesis imperfecta. Cytotherapy.
20:62–73. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Familtseva A, Jeremic N and Tyagi SC:
Exosomes: Cell-created drug delivery systems. Mol Cell Biochem.
459:1–6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu F, Lei N, Yang S, Zhou J, Chen M, Chen
C, Qiu L, Guo R, Li Y and Chang L: Treatment strategies for
intrauterine adhesion: Focus on the exosomes and hydrogels. Front
Bioeng Biotechnol. 11:12640062023. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chew JRJ, Chuah SJ, Teo KYW, Zhang S, Lai
RC, Fu JH, Lim LP, Lim SK and Toh WS: Mesenchymal stem cell
exosomes enhance periodontal ligament cell functions and promote
periodontal regeneration. Acta Biomater. 89:252–264. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kandalam U, Kawai T, Ravindran G, Brockman
R, Romero J, Munro M, Ortiz J, Heidari A, Thomas R, Kuriakose S, et
al: Predifferentiated gingival stem cell-induced bone regeneration
in rat alveolar bone defect model. Tissue Eng Part A. 27:424–436.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang CH, Sun CK, Lin YW, Hung MC, Lin HY,
Li CH, Lin IP, Chang HC, Sun JS and Chang JZ:
Metformin-incorporated gelatin/nano-hydroxyapatite scaffolds
promotes bone regeneration in critical size rat alveolar bone
defect model. Int J Mol Sci. 23:5582022. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo Q, Zheng J, Lin H, Han Z, Wang Z, Ren
J, Zhai J, Zhao H, Du R and Li C: Conditioned media of deer antler
stem cells accelerate regeneration of alveolar bone defects in
rats. Cell Prolif. 56:e134542023. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Witwer KW, Goberdhan DC, O'Driscoll L,
Théry C, Welsh JA, Blenkiron C, Buzás EI, Di Vizio D, Erdbrügger U,
Falcón-Pérez JM, et al: Updating MISEV: Evolving the minimal
requirements for studies of extracellular vesicles. J Extracell
Vesicles. 10:e121822021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gelibter S, Marostica G, Mandelli A,
Siciliani S, Podini P, Finardi A and Furlan R: The impact of
storage on extracellular vesicles: A systematic study. J Extracell
Vesicles. 11:e121622022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Matsuzaka Y and Yashiro R: Therapeutic
strategy of mesenchymal-stem-cell-derived extracellular vesicles as
regenerative medicine. Int J Mol Sci. 23:64802022. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Storti G, Scioli MG, Kim BS, Orlandi A and
Cervelli V: Adipose-derived stem cells in bone tissue engineering:
Useful tools with new applications. Stem Cells Int.
2019:36738572019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wan Z, Zhang P, Liu Y, Lv L and Zhou Y:
Four-dimensional bioprinting: Current developments and applications
in bone tissue engineering. Acta Biomater. 101:26–42. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen Y, Yang Q, Lv C, Chen Y, Zhao W, Li
W, Chen H, Wang H, Sun W and Yuan H: NLRP3 regulates alveolar bone
loss in ligature-induced periodontitis by promoting osteoclastic
differentiation. Cell Prolif. 54:e129732021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin W, Li Q, Zhang D, Zhang X, Qi X, Wang
Q, Chen Y, Liu C, Li H, Zhang S, et al: Mapping the immune
microenvironment for mandibular alveolar bone homeostasis at
single-cell resolution. Bone Res. 9:172021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kittaka M, Yoshimoto T, Schlosser C,
Rottapel R, Kajiya M, Kurihara H, Reichenberger EJ and Ueki Y:
Alveolar bone protection by targeting the SH3BP2-SYK axis in
osteoclasts. J Bone Miner Res. 35:382–395. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang X, Xie M, Xie Y, Mei F, Lu X, Li X
and Chen L: The roles of osteocytes in alveolar bone destruction in
periodontitis. J Transl Med. 18:4792020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan J, Dai A, Pan L, Zhang L, Wang Z, Ke
T, Sun W, Wu Y, Ding PH and Chen L: Inflamm-aging-related cytokines
of IL-17 and IFN-γ accelerate osteoclastogenesis and periodontal
destruction. J Immunol Res. 2021:99190242021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhu L, Zhou C, Chen S, Huang D, Jiang Y,
Lan Y, Zou S and Li Y: Osteoporosis and alveolar bone health in
periodontitis niche: A predisposing factors-centered review. Cells.
11:33802022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Setiawan M, Jäger A, Daratsianos N,
Reimann S, Chen J, Schmöle AC, Derichs-Schönthal D and Konermann A:
Impact of the endocannabinoid system on murine cranial and alveolar
bone phenotype. Ann Anat. 230:1515162020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martínez-Gil N, Mellibovsky L,
Manzano-López González D, Patiño JD, Cozar M, Rabionet R, Grinberg
D and Balcells S: On the association between Chiari malformation
type 1, bone mineral density and bone related genes. Bone Rep.
16:1011812022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vimalraj S: Alkaline phosphatase:
Structure, expression and its function in bone mineralization.
Gene. 754:1448552020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cirano FR, Pimentel SP, Ribeiro FV, Casati
MZ, Casarin RC, Gallafassi DF, Nishii D and Corrêa MG: Impact of
history of periodontitis on gene expression of bone-related factors
in young patients. Braz Oral Res. 34:e0142020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Balbaied T and Moore E: Overview of
optical and electrochemical alkaline phosphatase (ALP) biosensors:
Recent approaches in cells culture techniques. Biosensors (Basel).
9:1022019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Darjanki CM, Prahasanti C, Fitria AE,
Kusumawardani B, Wijaksana IKE and Aljunaid M: RUNX2 and ALP
expression in osteoblast cells exposed by PMMA-HAp combination: An
in vitro study. J Oral Biol Craniofac Res. 13:277–282. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim KM, Son HE, Min HY and Jang WG:
Vitexin enhances osteoblast differentiation through phosphorylation
of Smad and expression of Runx2 at in vitro and ex vivo. Mol Biol
Rep. 47:8809–8817. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gao RT, Zhan LP, Meng C, Zhang N, Chang
SM, Yao R and Li C: Homeobox B7 promotes the osteogenic
differentiation potential of mesenchymal stem cells by activating
RUNX2 and transcript of BSP. Int J Clin Exp Med. 8:10459–10470.
2015.PubMed/NCBI
|
|
45
|
Chen D, Kim DJ, Shen J, Zou Z and O'Keefe
RJ: Runx2 plays a central role in Osteoarthritis development. J
Orthop Translat. 23:132–139. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kazemi NY, Gendrot B, Berishvili E,
Markovic SN and Cohen M: The role and clinical interest of
extracellular vesicles in pregnancy and ovarian cancer.
Biomedicines. 9:12572021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gug C, Caba L, Mozos I, Stoian D, Atasie
D, Gug M and Gorduza EV: Rare splicing mutation in COL1A1 gene
identified by whole exomes sequencing in a patient with
osteogenesis imperfecta type I followed by prenatal diagnosis: A
case report and review of the literature. Gene. 741:1445652020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang L, Hu J, Meshkat BI, Liechty KW and
Xu J: LncRNA MALAT1 modulates TGF-β1-induced EMT in keratinocyte.
Int J Mol Sci. 22:118162021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Syed AM, Kundu S, Ram C, Kulhari U, Kumar
A, Mugale MN, Mohapatra P, Murty US and Sahu BD: Up-regulation of
Nrf2/HO-1 and inhibition of TGF-β1/Smad2/3 signaling axis by
daphnetin alleviates transverse aortic constriction-induced cardiac
remodeling in mice. Free Radic Biol Med. 186:17–30. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang J, Xiang H, Lu Y and Wu T: Role and
clinical significance of TGF-β1 and TGF-βR1 in malignant tumors
(Review). Int J Mol Med. 47:552021. View Article : Google Scholar : PubMed/NCBI
|