α-1 Antitrypsin (AAT) is an acute phase glycoprotein
with a molecular weight of 52 kDa, belonging to the serine protease
inhibitor (SERPIN) superfamily (1,2). It
is encoded by the SERPIN family A member 1 (SERPINA1)
gene located on the long arm of chromosome 14 (14q31-32.3), which
spans 12.2 kb and exhibits structural plasticity (3,4). The
structure of AAT comprises three β-folds (A-C) and nine α-helices
(A-I), along with a reaction center loop (RCL) that protrudes from
the molecule. Plasma AAT is primarily synthesized by hepatocytes,
but it is also produced by monocytes, macrophages and epithelial
cells (5). During the acute phase
response, circulating levels of AAT increase markedly. AAT has
anti-inflammatory, immunomodulatory, anti-infective and tissue
repair properties (6).
Changes in AAT expression levels are associated with
a variety of inflammatory and immune-mediated inflammatory
diseases, such as chronic obstructive pulmonary disease (COPD) and
rheumatoid arthritis (7–9). In addition, its expression levels are
also correlated with environmental exposure factors (10–14),
highlighting the role of this protein in diseases associated with
environmental exposure. Consequently, elucidating the association
between AAT and environmental factors is important to advance
AAT-related therapeutic developments. The present study reviews the
impact of AAT on inflammation, immune-mediated inflammatory
diseases and other conditions associated with genetic mutations and
environmental exposures. Mutations in the AAT gene are implicated
in lung inflammation, hepatitis, cirrhosis and liver cancer
(7,8,15–17).
Furthermore, AAT is discussed as a novel immunomodulator in
autoimmune diseases, as it is involved in complex signaling
pathways and interactions with multiple cytokines. It is proposed
that AAT may serve as a potential therapeutic target for
inflammatory diseases.
AAT, as other secreted proteins, requires processing
by the endoplasmic reticulum (ER) and Golgi apparatus. The stable
conformation of AAT is established through protein folding
(18). AAT synthesis is regulated
by both ER cargo receptors and biosynthetic quality control
systems. The process of AAT monomer extension to polymer secretion
is not accomplished in cells with disruption of the ER cargo
receptors lectin mannose binding 1 and surfeit protein locus 4. ER
cargo receptors modulate the synthesis of AAT within the ER and can
influence the accumulation of polymeric AAT by controlling the
concentration of precursor monomers and facilitating the secretion
of polymers (19). The
biosynthetic quality control system first enhances the structural
maturation of AAT and subsequently selectively eliminates immature
molecules, thereby promoting AAT secretion (20). The transport pathway of AAT is
intricate; it is transported to the pulmonary epithelium and
interstitium through apical endothelial cells via endocytosis and
transcytosis, with secretion occurring at the basolateral surface
(21). Additionally, AAT undergoes
bidirectional uptake and secretion between lung endothelial cells
and alveolar epithelial cells, as well as the air chambers
(22). The conformational
polymorphisms of the AAT protein contribute to the complexity of
its biological functions.
AAT is a multifunctional protein with several key
roles, including anti-inflammatory, antibacterial and antiapoptotic
functions, as well as the inhibition of serine proteases (23). As a unique endogenous
anti-inflammatory agent, AAT inhibits the synthesis and release of
inflammatory mediators while suppressing the production of
inflammatory cytokines. Its anti-inflammatory activities include
NF-κB-dependent mechanisms, such as the induction of the IL-1
receptor antagonist (24). The
antibacterial properties of AAT are primarily demonstrated through
its inhibition of Streptococcus pneumoniae in the lungs of
mice. A previous study reveals that lung clearance in untreated
mice infected with S. pneumoniae is compromised due to the
degradation of surfactant proteins A and D (which are important for
phagocytic activity) by neutrophil elastase (25). Conversely, AAT was shown to inhibit
neutrophil elastase-mediated degradation, thereby alleviating the
bacterial infection in the lungs (25). Additionally, AAT exerts
antiapoptotic effects on structural lung endothelial cells
(26). Previous studies have
investigated the role of AAT in disease regulation, particularly in
autoimmune diseases, diabetes and cell transplantation (9,27).
Furthermore, there is an adaptive immune response to AAT in
AAT-deficient lungs (28). In
addition, the anti-inflammatory and immunomodulatory properties of
AAT remain intact despite its anti-elastase effect (29). AAT therapy can prevent or reverse
type 1 diabetes and acute graft-vs.-host disease (GvHD) in
preclinical models of autoimmunity and transplantation, in which
alterations in cytokine and transcriptional profiles, as well as T
cell subset tolerance, are observed (30,31).
AAT may have a role in cellular senescence.
Oxidative stress is central to the cellular aging process (32) and the supplementation of exogenous
AAT can increase antioxidant levels such as SOD and reduces
oxidative stress (33). The
balance between oxidants and antioxidants is indirectly restored
through the antiapoptotic and anti-inflammatory effects of AAT,
although AAT does not directly facilitate the clearance of
oxidants. One of the key physiological functions of AAT is to
protect lung tissue from serine proteases (34), and AAT specifically inhibits
neutrophil elastase, proteinase 3 and proteinase G (35). The identification of AAT as a
potent inhibitor of neutrophil elastase led to the proposal of the
protease-antiprotease imbalance concept, which links the pulmonary
destruction associated with AAT deficiency (AATD) to the unchecked
activity of proteases (36). AATD
results in the loss of inhibition of neutrophil serine proteases,
leading to local tissue damage, as highlighted in the
protease-antiprotease hypothesis (37). Furthermore, AATD is associated with
the overexpression of inflammatory cytokine, which triggers
inflammation in lung cells, resulting in both lung and liver
disease.
AATD is an autosomal codominant disorder caused
mainly by point mutations in the SERPINA1 gene that can
cause lung related diseases such as emphysema (39). Decreased serum and tissue levels of
AAT increase the risk of developing COPD and emphysema (7,8).
Compared with healthy individuals, patients with COPD, emphysema or
bronchiectasis have an increased susceptibility to AATD (15,16).
AAT influences exacerbation patterns in patients with COPD,
particularly in those with frequent exacerbations of AATD (40). The levels of AAT protein differ
across various lung diseases, including cystic fibrosis,
interstitial pneumonia and bronchiectasis. In cystic fibrosis, AAT
levels remain normal, but neutrophil elastase levels increase to
levels that exceed the protective effect (41). Lower serum AAT levels are prevalent
in patients with non-idiopathic interstitial pneumonia compared
with patients with idiopathic interstitial pneumonia (42). Reduced AAT levels can also
instigate bronchiectasis (43). In
contrast to COPD, AATD is associated with an increased abundance
and activity of primary granule proteins, including neutrophil
elastase, on the cell surface (44). Coronavirus disease 2019 represents
a novel challenge with an unprecedented impact on human health and
development (45). AAT inhibits
severe acute respiratory syndrome coronavirus 2 infection by
blocking transmembrane serine protease 2 (46).
In addition to the lung diseases mentioned above,
AAT is also associated with a poor prognosis for cancer. In
non-small lung cancer cell lines, the presence of exogenous AAT
inhibits staurosporine (STS)-induced apoptosis. At the same time,
CLU (a pro-tumorigenic gene coding clusterin protein) expression
was higher (38). Furthermore, the
expression of AAT is associated with the metastasis of lung
adenocarcinoma cells, potentially promoting their spread by
upregulating fibronectin (47).
The upregulation of the expression of AAT enhances the adhesion
between lung adenocarcinoma cells and human umbilical vein
endothelial cells (47). This
adhesion represents a key step in the processes of tumor invasion
and metastasis.
In summary, AAT is associated with a variety of lung
inflammatory diseases. The expression level of AAT is different in
different pulmonary inflammatory diseases. The upregulated
expression of AAT can inhibit the inflammatory factors produced by
lung inflammation, while the lack of AAT can promote the expression
of inflammatory cytokines in the lung (Table I).
AATD is associated with neonatal hepatitis,
cirrhosis hepatocellular carcinoma and other liver-related diseases
(17). Misfolded AAT accumulates
in the ER of hepatocytes, leading to mitochondrial dysfunction
(48). Defective AAT results in
swelling and damage to hepatocytes, which can progress to cirrhosis
and pancreatitis (49,50). Hepatic steatosis can exacerbate
acute pancreatitis (51) and
panniculitis may be the initial presentation of both AATD and
pancreatic disease (52). Hepatic
steatosis acute pancreatitis is associated with reduced levels of
AAT and these levels associate with increased disease severity
(51). A previous study indicates
that AAT may be effective in treating acute liver failure and
pancreatic disease (53).
Furthermore, AATD may predispose patients to panniculitis (50). In addition to being associated with
a variety of lung inflammatory diseases, AAT is also associated
with liver inflammation. The main manifestation of AATD is that it
can cause diseases such as hepatitis, panniculitis, cirrhosis and
pancreatitis.
AAT exhibits anti-inflammatory and immunomodulatory
effects in various lung diseases; however, there is increasing
evidence that it also has a role in diseases such as rheumatoid
arthritis, systemic lupus erythematosus (9,31,54,55).
Autoimmune diseases are characterized by an overactive immune
system that attacks the tissues and organs of the host. Numerous
mechanisms and factors can trigger these diseases, including the
inflammatory response. Consequently, anti-inflammatory therapies
hold promise for the treatment of autoimmune diseases. AAT, an
anti-inflammatory protein, can prevent and reverse type 1 diabetes
and improve conditions such as rheumatoid arthritis and systemic
lupus erythematosus (SLE) (9,54).
Wegener's granulomatosis (WG), another autoimmune disease, is
classified as a necrotizing granulomatous vasculitis. Proteinase 3
is predominant in WG, and AAT acts as the main inhibitor of
proteinase 3; thus, AATD may contribute to the pathogenesis of WG
(55,56). Furthermore, AAT is hypothesized to
be a novel immunomodulator in transplantation (27). Acute graft-vs.-host disease (GvHD)
arises from the interaction of donor T cells, host
antigen-presenting cells and various proinflammatory cytokines
(such as TNF-α and IL-1β). However, exogenous AAT can mitigate
clinical manifestations of GvHD (31). Autoimmune diseases are caused by an
active immune system that produces a number of antibodies that
attack its own tissues, leading to inflammation and tissue damage.
In this process, AAT has an important anti-inflammatory role, and
AAT has become a potential therapeutic target for immune-mediated
inflammation.
AAT is associated with heart disease and
neurodegenerative diseases. Plasma-derived AAT reduces cardiac
infarct size in mice with acute myocardial infarction (57). Furthermore, exogenous AAT decreases
caspase-1 activity in the ischemic myocardium, thereby offering
myocardial protection (58). A
previous study indicated an association between AAT and the
regulation of vascular function by lipoproteins (59). Neuroinflammation contributes to the
degeneration of nerve cells, which is a hallmark of
neurodegenerative diseases. Blocking neuroinflammation by
downregulating inflammasome expression levels by altering the
expression of AAT may be beneficial in delaying the onset of
neurodegenerative diseases, as demonstrated in rd1 mice, a mouse
model for retinal degeneration (60,61).
AAT also exhibits therapeutic potential in diabetes,
with its activity being altered in both type 1 and type 2 diabetes
mellitus (62). Additionally, AAT
protects pancreatic β-cells from cytokine-induced apoptosis
(63), with these effects
potentially being mediated through the cAMP pathway (64). Upregulation of AAT expression
levels reduces the extent of intervertebral disc degeneration
(65). In addition, there is a
positive correlation between AAT levels and different types of
cancer, including pancreatic, ovarian, breast and colorectal
cancers (66–69). AAT enhances the resistance of
non-small cell lung cancer cells to anticancer drug-induced
apoptosis and autophagy (70).
Additionally, higher concentrations of AAT are observed in the sera
of patients with colorectal cancer compared with healthy controls
(66). Patients with gastric
cancer also have increased AAT levels in the gastric fluid compared
with that of healthy individuals and patients with benign
gastrointestinal diseases (71);
however, the precise mechanism underlying this observation remains
unclear and warrants further investigation. Nonetheless, AAT has a
potential use as a biomarker for gastric-related diseases (72) (Table
II).
AAT is expressed at different levels in, and is
associated with the mechanisms underlying the pathogenesis of, a
range of diseases (38,47,70).
The functions of AAT are associated with multiple factors such as
genetic polymorphisms of AAT, complex cellular signaling pathways
and multiple cytokines (73).
Genetic polymorphisms are associated with disease
development and prognosis. The SERPINA1 gene, which encodes
AAT, is polymorphic, with >100 known variants (74). Following gene mutation, plasma AAT
can undergo three different fates: Intracellular storage,
intracellular degradation or lack of synthesis (75). These alterations result in a
compromised defense mechanism in the lungs against serine proteases
(73). Wild-type AAT proteins
exhibit variable folding patterns. A more stable conformation is
achieved when the active central loop is incorporated as the fourth
strand in β-sheet A. The formation of these more stable
conformations can render AAT susceptible to mutations (76). Severely misfolded mutants trigger
the unfolded protein response (UPR), which enhances protein
folding. Conversely, if these mutants fail to activate the UPR,
they can promote NF-κB-mediated ER overload responses (77). The protease inhibitor (Pi) M
homozygotes represents the normal genotype, characterized by normal
AAT plasma levels (78).
Heterozygotes composed of M-type and other phenotypic (S/Z) alleles
exhibit AAT deficiency (78). By
contrast, the PiZ, PiS and Null alleles are defective variants,
with the PiZ allele most frequently associated with severe defects
and disease (79,80). Missense mutations identified in the
PiZ variant of AAT (Z-AAT) may accelerate misfolding and/or lead to
the formation of aggregates (81),
resulting in increased N-glycosylation of Z-AAT.
Serum AAT regulates ligand-receptor interactions,
which in turn modulate cytokine and neutrophil intracellular
signaling (85). In vitro,
demonstrate that AAT reduces the expression of Superoxide anion
(O2−) in neutrophils and inhibits the
stimulation of cyclic adenosine monophosphate receptors as well as
the phosphorylation of extracellular signal-regulated kinase (ERK)
1/2 (86). Neutrophil elastase, a
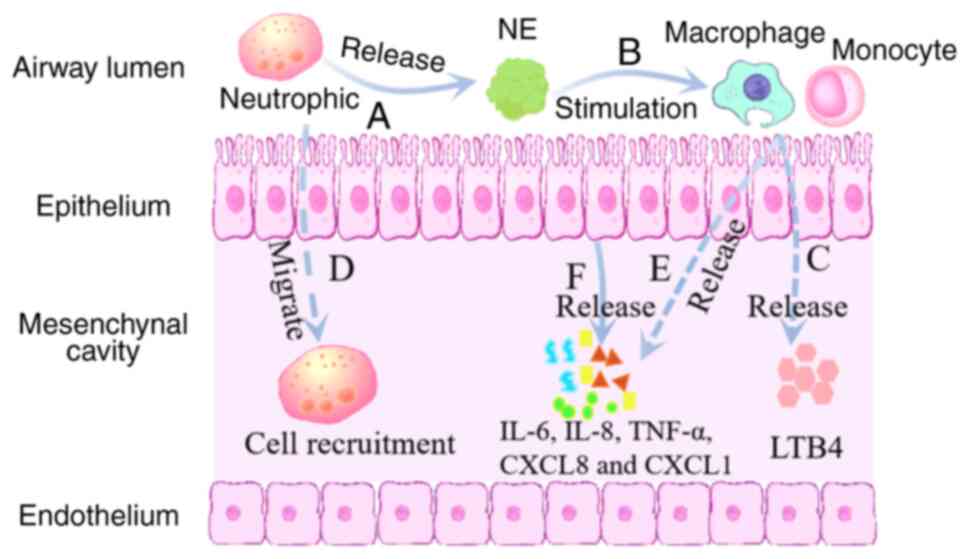
serine protease, is a key enzyme produced by neutrophils (87). In patients with AATD, AAT levels
are decreased, inactive polymers of AAT are present in the plasma
and neutrophil elastase levels are increased, resulting in an
imbalance in the pulmonary protease-antitrypsin system (88–91),
as shown in Fig. 1. AAT acts as a
serine protease inhibitor through RCL. At normal levels, AAT binds
to the serine protease and causes aberration and inactivation of
the serine protease. If the level of AAT is reduced or the level of
serine protease is increased, the inhibitory effect cannot be
played, resulting in the occurrence of disease (28,91).
In addition, another serine protease, protease 3, is also thought
to have the same or even greater impact on the disease process
(92).
In addition to directly interacting with proteases,
AAT also interacts indirectly with a variety of cytokines. The
mechanisms of the innate immunity can be modulated by the
anti-inflammatory activity of AAT, which is mediated through
interactions at the cell surface (93). AAT inhibits the secretion of
proinflammatory cytokines (94).
Both native and oxidized forms of AAT inhibit the ATP-induced
release of IL-1β from human monocytes (95,96),
independent of the antielastase activity of AAT. In addition, AAT
regulates ATP-induced IL-1β release through a novel triple
transmembrane signaling pathway. This triple transmembrane
signaling pathway includes lipid scavenger receptor CD36,
calcium-independent phospholipase A2β and the release of a small
soluble mediator (96). This
mediator activates nicotinic acetylcholine receptors, thereby
inhibiting the ATP-induced release of IL-1β from human monocytes
(96). In addition, glycosylated
AAT binds to IL-8, the ligand for C-X-C motif chemokine receptor 1
(Cxcr1), and obstructs the interaction of IL-8 with Cxcr1, thereby
inhibiting the release of pro-inflammatory cytokines (97). In the presence of Z-AAT and AATD,
neutrophils remain active and accumulate in the interstitial space,
exacerbating connective tissue destruction. It causes macrophages,
monocytes, alveolar epithelial cells and endothelial cells to
release inflammatory cytokines such as IL-6, IL-8, tumor necrosis
factor-α (TNF-α) and C-X-C motif chemokine ligands 8 and 1
(8,95,96,98,99),
as shown in Fig. 1. Circulating
serine protease inhibitors, including human AAT (hAAT), inhibit the
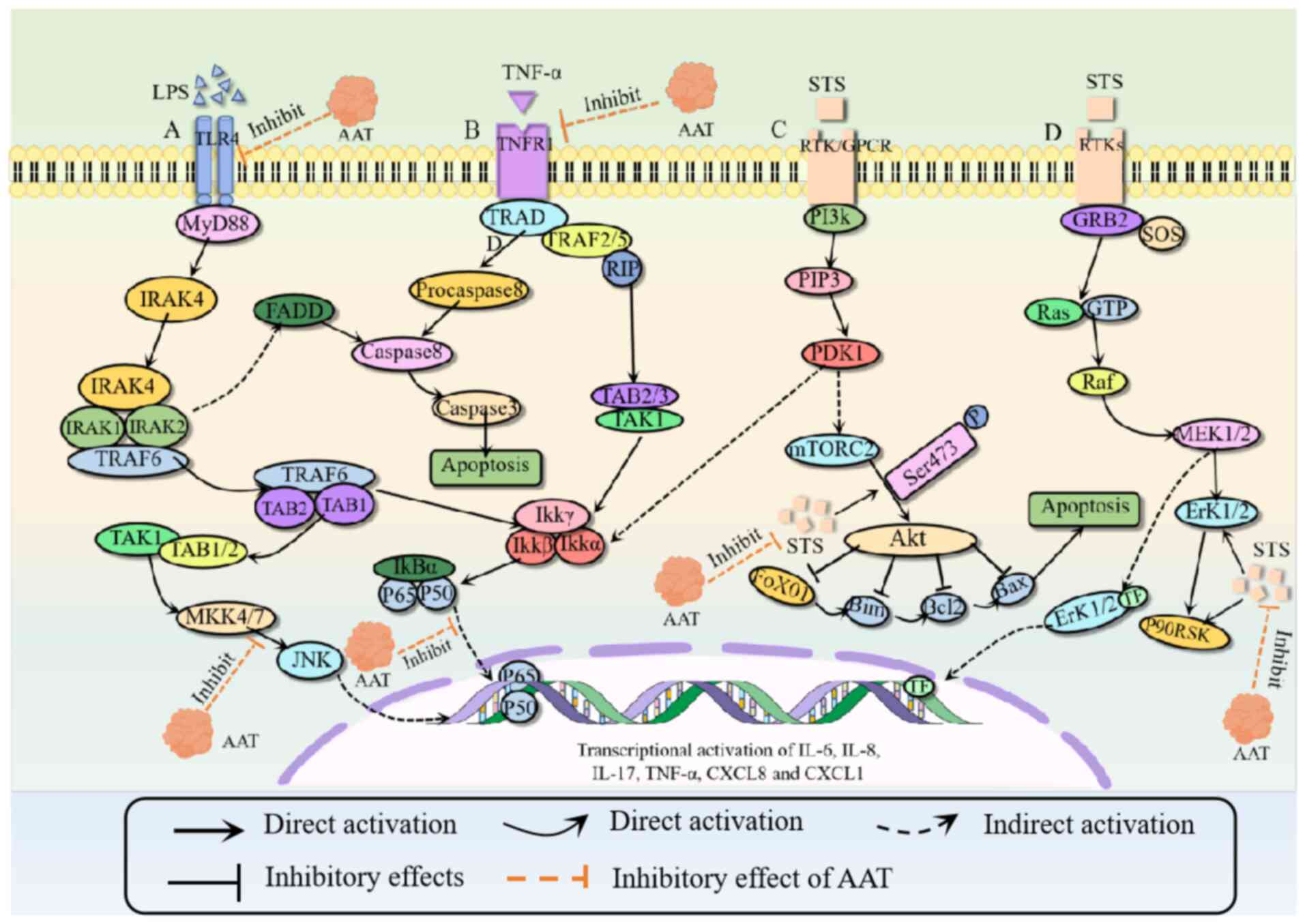
secretion of proinflammatory cytokines IL-17 and IL-6 (100). hAAT has a notable role in
inducing the production of anti-inflammatory cytokines. It enhances
the expression of IL-1 receptor (IL-1R) in macrophages and human
monocytes and promotes distinct phosphorylation and nuclear
translocation patterns of p65, a key transcription factor necessary
for the expression of IL-1R (101) as shown in Fig. 2. Additionally, hAAT increases the
number of T-regulatory cells as well as the expression of C-C
chemokine receptor type 6 in animal models (100,102).
AAT is involved in a variety of complex signaling
pathways. It attenuates coagulation and inhibits the
cytokine-induced activation of JNK and NF-κB in the instant
blood-mediated inflammatory response (103). Its anti-inflammatory activity
also involves NF-κB-dependent mechanisms (24). The accumulation of the Z-AAT
variant activates the NF-κB signaling pathway, leading to the
hypothesis that the downstream targets of NF-κB are components of
the proteostasis response network in this specific type of
proteinopathy (104).
Furthermore, the reduction of Z-AAT monomers may stimulate the
expression of the PiZ by decreasing the activation of hepatic NF-κB
and IL-6 levels (105).
Additionally, respiratory epithelial cells induce oxidative stress
and activate the NF-κB signaling pathway under senescent conditions
(106).
Oxidative stress is implicated in both the
physiological and pathological processes of AATD, suggesting that
AAT may have a role in cellular senescence (107,108). Additionally, TNF-α is key to the
pathogenesis of both hereditary AATD and non-hereditary COPD. TNF-α
can induce signal transduction in immune cells and lung endothelial
cells, and AAT is a key regulator of the TNF-α signaling pathway
(109). AAT inhibits the activity
of TNF-α-converting enzyme, suppresses the upregulation of TNF-α
receptor 1 and reduces the expression of TNF-α (Fig. 2). Calpain is activated by TNF-α and
AAT inhibits calpain activity, leading to a decrease in the level
of AAT itself (99). In addition,
AAT can inhibit the phosphorylation of IκBα, thereby reducing the
activation of NF-κB and inducing target gene transcription
(110). In alveolar epithelial
cells, lipopolysaccharide (LPS) induces toll-like receptor (TLR)
signaling pathways (54,96,111). AAT exerts anti-inflammatory
effects by inhibiting the expression of TLR4 and the
phosphorylation of IκBα (Fig. 2).
This signaling pathway also activates the JNK signaling pathway to
produce proinflammatory cytokines (103). However, AAT can inhibit the
phosphorylation of JNK and thus inhibits the proinflammatory
pathways (103,111). AAT also inhibits the TNF-α
induced activation of the WNT/β-catenin signaling pathway in human
bone marrow cells (61).
Lifestyle factors such as smoking can cause lung
disease. AAT protects the lung by blocking the constant influence
of damage associated molecular patterns and/or pathogen associated
molecular patterns caused by cigarette smoke, pollutants or
infections (114). In patients
with AATD, smoking exacerbates lung disease (10,115,116).
Numerous studies suggest that chronic
non-communicable diseases such as COPD develop as a result of a
combination of exposure to various environmental factors (such as
smoke, organic dusts, irradiation, toxic agents and metal
substances) and genetic predispositions (11,13,115,117–119). Levels of AAT vary under different
exposure conditions, and continued exposure of patients with AATD
to certain environmental factors accelerates disease progression.
There is a complex interrelationship between smoke exposure,
circulating AAT levels, systemic inflammation and lung function
(120). Furthermore, AAT levels
differ between smokers and non-smokers (117). Cigarette smoke has been shown to
inhibit AAT uptake in dermal cells and the lungs of mice (10); this inhibition is mediated by
neutrophil-derived serine proteases, primarily neutrophil elastase,
which can induce connective tissue rupture, leading to alveolar
space enlargement and emphysema in animal models (118). Exogenous AAT is protective and
can inhibit thrombin and plasma proteins that leak into the lungs
following cigarette smoke exposure, thereby preventing
protease-activated receptor type 1 activation and the release of
MMP-12 and TNF-α, which inhibits matrix degradation (121,122). Furthermore, cigarette smoke acts
as a proinflammatory agent (123). In individuals with genetic
defects in AAT, exposure to cigarette smoke accelerates the
development of COPD, eliciting an inflammatory response from AATD
macrophages to cigarette smoke-induced extracellular vesicles
(124). This is evidenced by the
additive role of smoking and intermediate AAT levels in PiMZ
heterozygotes in the development of emphysema (125), suggesting that gene-environment
interactions are key in the pathogenesis of COPD (119). AAT may mitigate smoking-induced
inflammation and stromal breakdown through an anti-inflammatory
mechanism that is associated with the inhibition of TNF-α,
providing partial protection against emphysema (126).
Various sources of exposure, including toxic agents,
metals and irradiation, notably impact the expression of AAT.
Chronic tramadol exposure leads to the dysregulation of
α-1-antitrypsin (encoded by SERPINA1b) (11), while exposure to sulfur mustard
notably increases AAT levels in saliva (12). In addition, arsenic metalloid
exposure in tap water reduces AAT in sputum (13). Irradiation also alters AAT levels;
in a mouse model subjected to total body irradiation with 11 Gy of
cobalt-60 γ radiation, AAT expression levels were increased
compared with that of non-irradiated controls (127,128). Furthermore, upregulation of the
AAT precursor expression level was noted in the plasma of CBA/CaJ
mice exposed to either 0 or 3 Gy of 137Cs gamma radiation (129). In a low-dose irradiated rat
model, established using intratracheal drip injection of uranium
tailings suspension, AAT expression levels were similarly
upregulated (14). The protective
mechanisms of AAT in environmental exposure injuries remain unclear
and warrant further investigation.
AAT has several key physiological and pathological
functions, and alterations in its activity can result in disease. A
deficiency or the abnormal expression levels of AAT can contribute
to lung and liver-related disorders. AAT may have the potential to
treat or prevent a range of diseases. In the context of autoimmune
and immune-mediated inflammatory diseases, AAT may serve as a novel
immunomodulator. Furthermore, AAT may influence cellular aging and
has been demonstrated to enhance antioxidant activity and mitigate
oxidative stress; however, the underlying mechanisms remain unclear
and warrant further investigation. Environmental factors such as
exposure to cigarette smoke, toxic substances and radiation can
impair lung function and alter AAT expression levels, particularly
in individuals with AATD. AAT is garnering increasing attention as
a key regulator of inflammatory and immune-mediated diseases, and
it may be a potential therapeutic target for these diseases.
Continued research may yield new therapeutic strategies for
specific diseases, offering more precise and effective treatment
options for patients. The application of AAT as a small molecule
immunomodulator also presents potential.
Not applicable.
This work was supported by Hunan Natural Science Foundation
(grant no. 2022JJ30478), Scientific Research Innovation Project for
Graduate Students in Hunan Province (grant no. CX20230991) and
Innovative Entrepreneurship Training Program for college students
in Hunan Province (grant nos. 2023-2600, 2023-2609 and
2023-4827).
Not applicable.
LY and TW contributed to the conceptualization of
the present study. TW, PS, SH and YL carried out the literature
search and were involved in the study design and conceptualization.
QW, CG and WW conducted the data/information search and critically
revised article content. PS, SH and YL performed the analysis. The
original draft was written by TW. The manuscript was subsequently
reviewed and edited by TW and LY. Data authentication is not
applicable. All authors read and approved the final version of the
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Sabina J and Tobias W: Augmentation
therapy with alpha1-antitrypsin: Novel perspectives. Cardiovasc
Hematol Disord Drug Targets. 13:90–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lechowicz U, Rudzinski S, Jezela-Stanek A,
Janciauskiene S and Chorostowska-Wynimko J: Post-translational
modifications of circulating alpha-1-antitrypsin protein. Int J Mol
Sci. 21:91872020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Santangelo S, Scarlata S, Poeta ML, Bialas
AJ, Paone G and Incalzi RA: Alpha-1 antitrypsin deficiency: Current
perspective from genetics to diagnosis and therapeutic approaches.
Curr Med Chem. 24:65–90. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haq I, Irving JA, Saleh AD, Dron L,
Regan-Mochrie GL, Motamedi-Shad N, Hurst JR, Gooptu B and Lomas DA:
Deficiency mutations of alpha-1 antitrypsin. Effects on folding,
function, and polymerization. Am J Respir Cell Mol Biol. 54:71–80.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van't Wout EF, van Schadewijk A, Savage
ND, Stolk J and Hiemstra PS: α1-Antitrypsin production by
proinflammatory and antiinflammatory macrophages and dendritic
cells. Am J Respir Cell Mol Biol. 46:607–613. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
de Serres F and Blanco I: Role of alpha-1
antitrypsin in human health and disease. J Intern Med. 276:311–335.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rahaghi FF and Miravitlles M: Long-term
clinical outcomes following treatment with alpha 1-proteinase
inhibitor for COPD associated with alpha-1 antitrypsin deficiency:
A look at the evidence. Respir Res. 18:1052017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stockley RA: Alpha1-antitrypsin review.
Clin Chest Med. 35:39–50. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song S: Alpha-1 antitrypsin therapy for
autoimmune disorders. Chronic Obstr Pulm Dis. 5:289–301.
2018.PubMed/NCBI
|
|
10
|
Serban KA, Petrusca DN, Mikosz A, Poirier
C, Lockett AD, Saint L, Justice MJ, Twigg HL III, Campos MA and
Petrache I: Alpha-1 antitrypsin supplementation improves alveolar
macrophages efferocytosis and phagocytosis following cigarette
smoke exposure. PLoS One. 12:e01760732017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang S, Liu G, Yuan H, Xu E, Xia W, Zhang
X, Liu J and Gao L: Changes on proteomic and metabolomic profile in
serum of mice induced by chronic exposure to tramadol. Sci Rep.
11:14542021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yarmohammadi ME, Hassan ZM, Mostafaie A,
Ebtekar M, Yaraee R, Pourfarzam S, Jalali-Nadoushan M, Faghihzadeh
S, Vaez-Mahdavi MR, Soroush MR, et al: Salivary levels of secretary
IgA, C5a and alpha 1-antitrypsin in sulfur mustard exposed patients
20 years after the exposure, sardasht-Iran cohort study (SICS). Int
Immunopharmacol. 17:952–957. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Burgess JL, Kurzius-Spencer M, Poplin GS,
Littau SR, Kopplin MJ, Stürup S, Boitano S and Clark Lantz R:
Environmental arsenic exposure, selenium and sputum alpha-1
antitrypsin. J Expo Sci Environ Epidemiol. 24:150–155. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yi L, Cui J, Hu N, Li L, Chen Y, Mu H, Yin
J, Wei S, Gong Y, Wei Y, et al: iTRAQ-based proteomic profiling of
potential biomarkers in rat serum for uranium tailing suspension
intratracheal instillation. J Proteome Res. 20:995–1004. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Veith M, Tüffers J, Peychev E, Klemmer A,
Kotke V, Janciauskiene S, Wilhelm S, Bals R, Koczulla AR,
Vogelmeier CF and Greulich T: The distribution of alpha-1
antitrypsin genotypes between patients with COPD/emphysema, asthma
and bronchiectasis. Int J Chron Obstruct Pulmon Dis. 15:2827–2836.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Alam S, Li Z, Atkinson C, Jonigk D,
Janciauskiene S and Mahadeva R: Z α1-antitrypsin confers a
proinflammatory phenotype that contributes to chronic obstructive
pulmonary disease. Am J Respir Crit Care Med. 189:909–931. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ordóñez A, Snapp EL, Tan L, Miranda E,
Marciniak SJ and Lomas DA: Endoplasmic reticulum polymers impair
luminal protein mobility and sensitize to cellular stress in
alpha1-antitrypsin deficiency. Hepatology. 57:2049–2060. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Giri Rao VVH and Gosavi S: On the folding
of a structurally complex protein to its metastable active state.
Proc Natl Acad Sci USA. 115:1998–2003. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ordóñez A, Harding HP, Marciniak SJ and
Ron D: Cargo receptor-assisted endoplasmic reticulum export of
pathogenic α1-antitrypsin polymers. Cell Rep. 35:1091442021.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ronzoni R, Berardelli R, Medicina D, Sitia
R, Gooptu B and Fra AM: Aberrant disulphide bonding contributes to
the ER retention of alpha1-antitrypsin deficiency variants. Hum Mol
Genet. 25:642–650. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lockett AD: Alpha-1 antitrypsin
transcytosis and secretion. Methods Mol Biol. 1639:173–184. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lockett AD, Brown MB, Santos-Falcon N,
Rush NI, Oueini H, Oberle AJ, Bolanis E, Fragoso MA, Petrusca DN,
Serban KA, et al: Active trafficking of alpha 1 antitrypsin across
the lung endothelium. PLoS One. 9:e939792014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim M, Cai Q and Oh Y: Therapeutic
potential of alpha-1 antitrypsin in human disease. Ann Pediatr
Endocrinol Metab. 23:131–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schuster R, Motola-Kalay N, Baranovski BM,
Bar L, Tov N, Stein M, Lewis EC, Ayalon M and Sagiv Y: Distinct
anti-inflammatory properties of alpha1-antitrypsin and
corticosteroids reveal unique underlying mechanisms of action. Cell
Immunol. 356:1041772020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ostermann L, Maus R, Stolper J, Schütte L,
Katsarou K, Tumpara S, Pich A, Mueller C, Janciauskiene S, Welte T
and Maus UA: Alpha-1 antitrypsin deficiency impairs lung
antibacterial immunity in mice. JCI Insight. 6:e1408162021.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Serban KA and Petrache I: alpha-1
antitrypsin and lung cell apoptosis. Ann Am Thorac Soc. 13 (Suppl
2):S146–S149. 2016.PubMed/NCBI
|
|
27
|
Han L, Wu X, Wang O, Luan X, Velander WH,
Aynardi M, Halstead ES, Bonavia AS, Jin R, Li G, et al: Mesenchymal
stromal cells and alpha-1 antitrypsin have a strong synergy in
modulating inflammation and its resolution. Theranostics.
13:2843–2862. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baraldo S, Turato G, Lunardi F, Bazzan E,
Schiavon M, Ferrarotti I, Molena B, Cazzuffi R, Damin M, Balestro
E, et al: Immune activation in α1-antitrypsin-deficiency emphysema.
Beyond the protease-antiprotease paradigm. Am J Respir Crit Care
Med. 191:402–409. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jonigk D, Al-Omari M, Maegel L, Müller M,
Izykowski N, Hong J, Hong K, Kim SH, Dorsch M, Mahadeva R, et al:
Anti-inflammatory and immunomodulatory properties of α1-antitrypsin
without inhibition of elastase. Proc Natl Acad Sci USA.
110:15007–15012. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ehlers MR: Immune-modulating effects of
alpha-1 antitrypsin. Biol Chem. 395:1187–1193. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tawara I, Sun Y, Lewis EC, Toubai T, Evers
R, Nieves E, Azam T, Dinarello CA and Reddy P: Alpha-1-antitrypsin
monotherapy reduces graft-versus-host disease after experimental
allogeneic bone marrow transplantation. Proc Natl Acad Sci USA.
109:564–569. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Blas-García A and Apostolova N: Novel
therapeutic approaches to liver fibrosis based on targeting
oxidative stress. Antioxidants (Basel). 12:15672023. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Feng Y, Xu J, Zhou Q, Wang R, Liu N, Wu Y,
Yuan H and Che H: Alpha-1 antitrypsin prevents the development of
preeclampsia through suppression of oxidative stress. Front
Physiol. 7:1762016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chapman KR, Chorostowska-Wynimko J,
Koczulla AR, Ferrarotti I and McElvaney NG: Alpha 1 antitrypsin to
treat lung disease in alpha 1 antitrypsin deficiency: Recent
developments and clinical implications. Int J Chron Obstruct Pulmon
Dis. 13:419–432. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Janciauskiene S and Welte T: Well-known
and less well-known functions of alpha-1 antitrypsin. its role in
chronic obstructive pulmonary disease and other disease
developments. Ann Am Thorac Soc. 13 (Suppl 4):S280–S288. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cosio MG, Bazzan E, Rigobello C, Tinè M,
Turato G, Baraldo S and Saetta M: Alpha-1 antitrypsin deficiency:
beyond the protease/antiprotease paradigm. Ann Am Thorac Soc. 13
(Suppl 4):S305–S310. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Stockley RA: The multiple facets of
alpha-1-antitrypsin. Ann Transl Med. 3:1302015.PubMed/NCBI
|
|
38
|
Schwarz N, Tumpara S, Wrenger S, Ercetin
E, Hamacher J, Welte T and Janciauskiene S: Alpha1-antitrypsin
protects lung cancer cells from staurosporine-induced apoptosis:
The role of bacterial lipopolysaccharide. Sci Rep. 10:95632020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Meghadri SH, Martinez-Delgado B, Ostermann
L, Gomez-Mariano G, Perez-Luz S, Tumpara S, Wrenger S, DeLuca DS,
Maus UA, Welte T and Janciauskiene S: Loss of Serpina1 in mice
leads to altered gene expression in inflammatory and metabolic
pathways. Int J Mol Sci. 23:104252022. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Stolk J, Tov N, Chapman KR, Fernandez P,
MacNee W, Hopkinson NS, Piitulainen E, Seersholm N, Vogelmeier CF,
Bals R, et al: Efficacy and safety of inhaled α1-antitrypsin in
patients with severe α1-antitrypsin deficiency and frequent
exacerbations of COPD. Eur Respir J. 54:19006732019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
McElvaney NG: Alpha-1 antitrypsin therapy
in cystic fibrosis and the lung disease associated with alpha-1
antitrypsin deficiency. Ann Am Thorac Soc. 13 (Suppl 2):S191–S196.
2016.PubMed/NCBI
|
|
42
|
Demir N, Erçen Diken Ö, Karabulut HG,
Karnak D and Kayacan O: Alpha-1 antitrypsin levels and
polymorphisms in interstitial lung diseases. Turk J Med Sci.
47:476–482. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Oriano M, Amati F, Gramegna A, De Soyza A,
Mantero M, Sibila O, Chotirmall SH, Voza A, Marchisio P, Blasi F
and Aliberti S: Protease-antiprotease imbalance in bronchiectasis.
Int J Mol Sci. 22:59962021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Murphy MP, McEnery T, McQuillan K,
McElvaney OF, McElvaney OJ, Landers S, Coleman O, Bussayajirapong
A, Hawkins P, Henry M, et al: α1 Antitrypsin therapy
modulates the neutrophil membrane proteome and secretome. Eur
Respir J. 55:19016782020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ritzmann F, Chitirala P, Krüger N,
Hoffmann M, Zuo W, Lammert F, Smola S, Tov N, Alagem N, Lepper PM,
et al: Therapeutic application of alpha-1 antitrypsin in COVID-19.
Am J Respir Crit Care Med. 204:224–227. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang C, Keshavjee S and Liu M: Alpha-1
antitrypsin for COVID-19 treatment: Dual role in antiviral
infection and anti-inflammation. Front Pharmacol. 11:6153982020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li Y, Miao L, Yu M, Shi M, Wang Y, Yang J,
Xiao Y and Cai H: α1-Antitrypsin promotes lung adenocarcinoma
metastasis through upregulating fibronectin expression. Int J
Oncol. 50:1955–1964. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Khodayari N, Wang RL, Oshins R, Lu Y,
Millett M, Aranyos AM, Mostofizadeh S, Scindia Y, Flagg TO and
Brantly M: The mechanism of mitochondrial injury in alpha-1
antitrypsin deficiency mediated liver disease. Int J Mol Sci.
22:132552021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tanash HA and Piitulainen E: Liver disease
in adults with severe alpha-1-antitrypsin deficiency. J
Gastroenterol. 54:541–548. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Franciosi AN, Ralph J, O'Farrell NJ,
Buckley C, Gulmann C, O'Kane M, Carroll TP and McElvaney NG:
Alpha-1 antitrypsin deficiency-associated panniculitis. J Am Acad
Dermatol. 87:825–832. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang Q, Du J, Yu P, Bai B, Zhao Z, Wang S,
Zhu J, Feng Q, Gao Y, Zhao Q and Liu C: Hepatic steatosis depresses
alpha-1-antitrypsin levels in human and rat acute pancreatitis. Sci
Rep. 5:178332015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yu Y, Rubin AG, Gee S, Banker S and Kim
CN: Ulcerative panniculitis with fevers and pleural effusions: A
unique case of α1-antitrypsin deficiency. JAAD Case Rep. 1:1–2.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jedicke N, Struever N, Aggrawal N, Welte
T, Manns MP, Malek NP, Zender L, Janciauskiene S and Wuestefeld T:
α-1-antitrypsin inhibits acute liver failure in mice. Hepatology.
59:2299–2308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Elshikha AS, Lu Y, Chen MJ, Akbar M,
Zeumer L, Ritter A, Elghamry H, Mahdi MA, Morel L and Song S: Alpha
1 antitrypsin inhibits dendritic cell activation and attenuates
nephritis in a mouse model of lupus. PLoS One. 11:e01565832016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Pervakova MY, Emanuel VL, Titova ON, Lapin
SV, Mazurov VI, Belyaeva IB, Chudinov AL, Blinova TV and Surkova
EA: The diagnostic value of alpha-1-antitrypsin phenotype in
patients with granulomatosis with polyangiitis. Int J Rheumatol.
2016:78314102016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mota A, Sahebghadam Lotfi A, Jamshidi AR
and Najavand S: Alpha 1-antitrypsin activity is markedly decreased
in Wegener's granulomatosis. Rheumatol Int. 34:553–558. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Mauro AG, Mezzaroma E, Marchetti C,
Narayan P, Del Buono MG, Capuano M, Prestamburgo A, Catapano S,
Salloum FN, Abbate A and Toldo S: A preclinical translational study
of the cardioprotective effects of plasma-derived alpha-1
anti-trypsin in acute myocardial infarction. J Cardiovasc
Pharmacol. 69:273–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Toldo S, Mauro AG, Marchetti C, Rose SW,
Mezzaroma E, Van Tassell BW, Kim S, Dinarello CA and Abbate A:
Recombinant human alpha-1 antitrypsin-Fc fusion protein reduces
mouse myocardial inflammatory injury after ischemia-reperfusion
independent of elastase inhibition. J Cardiovasc Pharmacol.
68:27–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lockett AD, Petrusca DN, Justice MJ,
Poirier C, Serban KA, Rush NI, Kamocka M, Predescu D, Predescu S
and Petrache I: Scavenger receptor class B, type I-mediated uptake
of A1AT by pulmonary endothelial cells. Am J Physiol Lung Cell Mol
Physiol. 309:L425–L434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhou T, Huang Z, Zhu X, Sun X, Liu Y,
Cheng B, Li M, Liu Y, He C and Liu X: Alpha-1 antitrypsin
attenuates M1 microglia-mediated neuroinflammation in retinal
degeneration. Front Immunol. 9:12022018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ebrahimi T, Rust M, Kaiser SN, Slowik A,
Beyer C, Koczulla AR, Schulz JB, Habib P and Bach JP:
α1-Antitrypsin mitigates NLRP3-inflammasome activation in amyloid
β1-42-stimulated murine astrocytes. J Neuroinflammation.
15:2822018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Park SS, Rodriguez Ortega R, Agudelo CW,
Perez Perez J, Perez Gandara B, Garcia-Arcos I, McCarthy C and
Geraghty P: Therapeutic potential of alpha-1 antitrypsin in type 1
and type 2 diabetes mellitus. Medicina (Kaunas). 57:3972021.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fleixo-Lima G, Ventura H, Medini M, Bar L,
Strauss P and Lewis EC: Mechanistic evidence in support of
alpha1-antitrypsin as a therapeutic approach for type 1 diabetes. J
Diabetes Sci Technol. 8:1193–1203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kalis M, Kumar R, Janciauskiene S, Salehi
A and Cilio CM: α 1-antitrypsin enhances insulin secretion and
prevents cytokine-mediated apoptosis in pancreatic β-cells. Islets.
2:185–189. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liu W and Wang Y: Protective role of the
alpha-1-antitrypsin in intervertebral disc degeneration. J Orthop
Surg Res. 16:5162021. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Pérez-Holanda S, Blanco I, Menéndez M and
Rodrigo L: Serum concentration of alpha-1 antitrypsin is
significantly higher in colorectal cancer patients than in healthy
controls. BMC Cancer. 14:3552014. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tountas Y, Sparos L, Theodoropoulos C and
Trichopoulos D: Alpha 1-antitrypsin and cancer of the pancreas.
Digestion. 31:37–40. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Vasishta A, Baker PR, Preece PE, Wood RA
and Cuschieri A: Serum proteinase-like peptidase activities and
proteinase inhibitors in women with breast disease. Eur J Cancer
Clin Oncol. 20:197–202. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Warwas M, Gerber J and Pietkiewicz A:
Haptoglobin and proteinase inhibitors in the blood serum of women
with inflammatory, benign and neoplastic lesions of the ovary.
Neoplasma. 33:79–84. 1986.PubMed/NCBI
|
|
70
|
Janciauskiene S, Wrenger S, Günzel S,
Gründing AR, Golpon H and Welte T: Potential roles of acute phase
proteins in cancer: Why do cancer cells produce or take up
exogenous acute phase protein alpha1-antitrypsin? Front Oncol.
11:6220762021. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hsu PI, Chen CH, Hsiao M, Wu DC, Lin CY,
Lai KH and Lu PJ: Diagnosis of gastric malignancy using gastric
juice alpha1-antitrypsin. Cancer Epidemiol Biomarkers Prev.
19:405–411. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wu W, Juan WC, Liang CR, Yeoh KG, So J and
Chung MC: S100A9, GIF and AAT as potential combinatorial biomarkers
in gastric cancer diagnosis and prognosis. Proteomics Clin Appl.
6:152–162. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Geramizadeh B, Jowkar Z, Karami L,
Masoumpour M, Mehrabi S and Ghayoumi MA: Alpha-1 antitrypsin
deficiency in Iranian patients with chronic obstructive pulmonary
disease. Iran Red Crescent Med J. 15:e75082013. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Matamala N, Lara B, Gomez-Mariano G,
Martínez S, Retana D, Fernandez T, Silvestre RA, Belmonte I,
Rodriguez-Frias F, Vilar M, et al: Characterization of novel
missense variants of SERPINA1 gene causing alpha-1 antitrypsin
deficiency. Am J Respir Cell Mol Biol. 58:706–716. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Foil KE: Variants of SERPINA1 and the
increasing complexity of testing for alpha-1 antitrypsin
deficiency. Ther Adv Chronic Dis (12 Suppl). 204062232110159542021.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tsutsui Y, Dela Cruz R and Wintrode PL:
Folding mechanism of the metastable serpin α1-antitrypsin. Proc
Natl Acad Sci USA. 109:4467–4472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Marciniak SJ, Ordóñez A, Dickens JA,
Chambers JE, Patel V, Dominicus CS and Malzer E: New concepts in
alpha-1 antitrypsin deficiency disease mechanisms. Ann Am Thorac
Soc. 13 (Suppl 4):S289–S296. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Börner FR, Lechowicz U, Wrenger S,
Martinez-Delgado B, Olejnicka B, Welte T, Chorostowska-Wynimko J,
Kiehntopf M and Janciauskiene S: Plasma levels of
α1-antitrypsin-derived C-terminal peptides in PiMM and
PiZZ COPD patients. ERJ Open Res. 9:00329–2023. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Fra AM, Gooptu B, Ferrarotti I, Miranda E,
Scabini R, Ronzoni R, Benini F, Corda L, Medicina D, Luisetti M and
Schiaffonati L: Three new alpha1-antitrypsin deficiency variants
help to define a C-terminal region regulating conformational change
and polymerization. PLoS One. 7:e384052012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Salahuddin P: Genetic variants of
alpha1-antitrypsin. Curr Protein Pept Sci. 11:101–117. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Karatas E and Bouchecareilh M: Alpha
1-antitrypsin deficiency: A disorder of proteostasis-mediated
protein folding and trafficking pathways. Int J Mol Sci.
21:14932020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Miranda E, Ferrarotti I, Berardelli R,
Laffranchi M, Cerea M, Gangemi F, Haq I, Ottaviani S, Lomas DA,
Irving JA and Fra A: The pathological Trento variant of
alpha-1-antitrypsin (E75V) shows nonclassical behaviour during
polymerization. FEBS J. 284:2110–2126. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Laffranchi M, Berardelli R, Ronzoni R,
Lomas DA and Fra A: Heteropolymerization of α-1-antitrypsin mutants
in cell models mimicking heterozygosity. Hum Mol Genet.
27:1785–1793. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Laffranchi M, Elliston ELK, Gangemi F,
Berardelli R, Lomas DA, Irving JA and Fra A: Characterisation of a
type II functionally-deficient variant of alpha-1-antitrypsin
discovered in the general population. PLoS One. 14:e02069552019.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bergin DA, Reeves EP, Hurley K, Wolfe R,
Jameel R, Fitzgerald S and McElvaney NG: The circulating proteinase
inhibitor α-1 antitrypsin regulates neutrophil degranulation and
autoimmunity. Sci Transl Med. 6:217ra12014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Hawkins P, McEnery T, Gabillard-Lefort C,
Bergin DA, Alfawaz B, Shutchaidat V, Meleady P, Henry M, Coleman O,
Murphy M, et al: In vitro and in vivo modulation of NADPH oxidase
activity and reactive oxygen species production in human
neutrophils by α1-antitrypsin. ERJ Open Res.
7:00234–2021. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Voynow JA and Shinbashi M: Neutrophil
elastase and chronic lung disease. Biomolecules. 11:10652021.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Miravitlles M: Alpha-1-antitrypsin and
other proteinase inhibitors. Curr Opin Pharmacol. 12:309–314. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hurley K, Lacey N, O'Dwyer CA, Bergin DA,
McElvaney OJ, O'Brien ME, McElvaney OF, Reeves EP and McElvaney NG:
Alpha-1 antitrypsin augmentation therapy corrects accelerated
neutrophil apoptosis in deficient individuals. J Immunol.
193:3978–3991. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
O'Dwyer CA, O'Brien ME, Wormald MR, White
MM, Banville N, Hurley K, McCarthy C, McElvaney NG and Reeves EP:
The BLT1 inhibitory function of α-1 antitrypsin augmentation
therapy disrupts leukotriene B4 neutrophil signaling. J Immunol.
195:3628–3641. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
McCarthy C, Reeves EP and McElvaney NG:
The role of neutrophils in alpha-1 antitrypsin deficiency. Ann Am
Thorac Soc. 13 (Suppl 4):S297–S304. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Fazleen A and Wilkinson T: The emerging
role of proteases in α1-antitrypsin deficiency and
beyond. ERJ Open Res. 7:00494–2021. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
O'Brien ME, Murray G, Gogoi D, Yusuf A,
McCarthy C, Wormald MR, Casey M, Gabillard-Lefort C, McElvaney NG
and Reeves EP: A review of alpha-1 antitrypsin binding partners for
immune regulation and potential therapeutic application. Int J Mol
Sci. 23:24412022. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Joosten LA, Crişan TO, Azam T, Cleophas
MC, Koenders MI, van de Veerdonk FL, Netea MG, Kim S and Dinarello
CA: Alpha-1-anti-trypsin-Fc fusion protein ameliorates gouty
arthritis by reducing release and extracellular processing of IL-1β
and by the induction of endogenous IL-1Ra. Ann Rheum Dis.
75:1219–1227. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Agné A, Richter K, Padberg W,
Janciauskiene S and Grau V: Commercial α1-antitrypsin preparations
markedly differ in their potential to inhibit the ATP-induced
release of monocytic interleukin-1β. Pulm Pharmacol Ther.
68:1020202021. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Siebers K, Fink B, Zakrzewicz A, Agné A,
Richter K, Konzok S, Hecker A, Zukunft S, Küllmar M, Klein J, et
al: Alpha-1 antitrypsin inhibits ATP-mediated release of
interleukin-1β via CD36 and nicotinic acetylcholine receptors.
Front Immunol. 9:8772018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Bergin DA, Reeves EP, Meleady P, Henry M,
McElvaney OJ, Carroll TP, Condron C, Chotirmall SH, Clynes M,
O'Neill SJ and McElvaney NG: α-1 Antitrypsin regulates human
neutrophil chemotaxis induced by soluble immune complexes and IL-8.
J Clin Invest. 120:4236–4250. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lee J, Lu Y, Oshins R, West J, Moneypenny
CG, Han K and Brantly ML: Alpha 1 antitrypsin-deficient macrophages
have impaired efferocytosis of apoptotic neutrophils. Front
Immunol. 11:5744102020. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Lockett AD, Kimani S, Ddungu G, Wrenger S,
Tuder RM, Janciauskiene SM and Petrache I:
α1-Antitrypsin modulates lung endothelial cell
inflammatory responses to TNF-α. Am J Respir Cell Mol Biol.
49:143–150. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhukovsky N, Silvano M, Filloux T,
Gonzalez S and Krause KH: Alpha-1 antitrypsin reduces disease
progression in a mouse model of charcot-marie-tooth type 1A: A role
for decreased inflammation and ADAM-17 inhibition. Int J Mol Sci.
23:74052022. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Abecassis A, Schuster R, Shahaf G, Ozeri
E, Green R, Ochayon DE, Rider P and Lewis EC: α1-Antitrypsin
increases interleukin-1 receptor antagonist production during
pancreatic islet graft transplantation. Cell Mol Immunol.
11:377–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ozeri E, Mizrahi M, Shahaf G and Lewis EC:
α-1 antitrypsin promotes semimature, IL-10-producing and readily
migrating tolerogenic dendritic cells. J Immunol. 189:146–153.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Wang J, Sun Z, Gou W, Adams DB, Cui W,
Morgan KA, Strange C and Wang H: α-1 antitrypsin enhances islet
engraftment by suppression of instant blood-mediated inflammatory
reaction. Diabetes. 66:970–980. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Mukherjee A, Hidvegi T, Araya P, Ewing M,
Stolz DB and Perlmutter DH: NFκB mitigates the pathological effects
of misfolded α1-antitrypsin by activating autophagy and an
integrated program of proteostasis mechanisms. Cell Death Differ.
26:455–469. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Pastore N, Blomenkamp K, Annunziata F,
Piccolo P, Mithbaokar P, Maria Sepe R, Vetrini F, Palmer D, Ng P,
Polishchuk E, et al: Gene transfer of master autophagy regulator
TFEB results in clearance of toxic protein and correction of
hepatic disease in alpha-1-anti-trypsin deficiency. EMBO Mol Med.
5:397–412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Rivas M, Gupta G, Costanzo L, Ahmed H,
Wyman AE and Geraghty P: Senescence: Pathogenic driver in chronic
obstructive pulmonary disease. Medicina (Kaunas). 58:8172022.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Saferali A, Lee J, Sin DD, Rouhani FN,
Brantly ML and Sandford AJ: Longer telomere length in COPD patients
with α1-antitrypsin deficiency independent of lung function. PLoS
One. 9:e956002014. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Escribano A, Pastor S, Reula A, Castillo
S, Vicente S, Sanz F, Casas F, Torres M, Fernández-Fabrellas E,
Codoñer-Franch P and Dasí F: Accelerated telomere attrition in
children and teenagers with α1-antitrypsin deficiency. Eur Respir
J. 48:350–358. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hurley K, Reeves EP, Carroll TP and
McElvaney NG: Tumor necrosis factor-α driven inflammation in
alpha-1 antitrypsin deficiency: A new model of pathogenesis and
treatment. Expert Rev Respir Med. 10:207–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Li Z, Alam S, Wang J, Sandstrom CS,
Janciauskiene S and Mahadeva R: Oxidized {alpha}1-antitrypsin
stimulates the release of monocyte chemotactic protein-1 from lung
epithelial cells: Potential role in emphysema. Am J Physiol Lung
Cell Mol Physiol. 297:L388–L400. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Subramaniyam D, Glader P, von Wachenfeldt
K, Burneckiene J, Stevens T and Janciauskiene S: C-36 peptide, a
degradation product of alpha1-antitrypsin, modulates human monocyte
activation through LPS signaling pathways. Int J Biochem Cell Biol.
38:563–575. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Antonsson A and Persson JL: Induction of
apoptosis by staurosporine involves the inhibition of expression of
the major cell cycle proteins at the G(2)/m checkpoint accompanied
by alterations in Erk and Akt kinase activities. Anticancer Res.
29:2893–2898. 2009.PubMed/NCBI
|
|
113
|
Campos MA, Geraghty P, Holt G, Mendes E,
Newby PR, Ma S, Luna-Diaz LV, Turino GM and Stockley RA: The
biological effects of double-dose alpha-1 antitrypsin augmentation
therapy. A pilot clinical trial. Am J Respir Crit Care Med.
200:318–326. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Hunt JM and Tuder R: Alpha 1 anti-trypsin:
One protein, many functions. Curr Mol Med. 12:827–835. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Berman R, Jiang D, Wu Q and Chu HW:
α1-Antitrypsin reduces rhinovirus infection in primary human airway
epithelial cells exposed to cigarette smoke. Int J Chron Obstruct
Pulmon Dis. 11:1279–1286. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Mehta AJ, Thun GA, Imboden M, Ferrarotti
I, Keidel D, Künzli N, Kromhout H, Miedinger D, Phuleria H, Rochat
T, et al: Interactions between SERPINA1 PiMZ genotype, occupational
exposure and lung function decline. Occup Environ Med. 71:234–240.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Wrześniak M, Kepinska M, Królik M and
Milnerowicz H: The influence of tobacco smoke on protein and metal
levels in the serum of women during pregnancy. PLoS One.
11:e01613422016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Pemberton PA, Kobayashi D, Wilk BJ,
Henstrand JM, Shapiro SD and Barr PJ: Inhaled recombinant alpha
1-antitrypsin ameliorates cigarette smoke-induced emphysema in the
mouse. COPD. 3:101–108. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Molloy K, Hersh CP, Morris VB, Carroll TP,
O'Connor CA, Lasky-Su JA, Greene CM, O'Neill SJ, Silverman EK and
McElvaney NG: Clarification of the risk of chronic obstructive
pulmonary disease in α1-antitrypsin deficiency PiMZ heterozygotes.
Am J Respir Crit Care Med. 189:419–427. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Stearns K, Goldklang M, Xiao R, Zelonina
T, Blomenkamp K, Teckman J and D'Armiento JM: Knockdown of alpha-1
antitrypsin with antisense oligonucleotide does not exacerbate
smoke induced lung injury. PLoS One. 16:e02460402021. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Rangaraju M and Turner AM: Why is disease
penetration so variable in alpha-1 antitrypsin deficiency? The
contribution of environmental factors. Chronic Obstr Pulm Dis.
7:280–289. 2020.PubMed/NCBI
|
|
122
|
Churg A, Wang X, Wang RD, Meixner SC,
Pryzdial EL and Wright JL: Alpha1-antitrypsin suppresses TNF-alpha
and MMP-12 production by cigarette smoke-stimulated macrophages. Am
J Respir Cell Mol Biol. 37:144–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Thun GA, Ferrarotti I, Imboden M, Rochat
T, Gerbase M, Kronenberg F, Bridevaux PO, Zemp E, Zorzetto M,
Ottaviani S, et al: SERPINA1 PiZ and PiS heterozygotes and lung
function decline in the SAPALDIA cohort. PLoS One. 7:e427282012.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Khodayari N, Oshins R, Mehrad B, Lascano
JE, Qiang X, West JR, Holliday LS, Lee J, Wiesemann G, Eydgahi S
and Brantly M: Cigarette smoke exposed airway epithelial
cell-derived EVs promote pro-inflammatory macrophage activation in
alpha-1 antitrypsin deficiency. Respir Res. 23:2322022. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Al Ashry HS and Strange C: COPD in
individuals with the PiMZ alpha-1 antitrypsin genotype. Eur Respir
Rev. 26:1700682017. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Geraghty P, Eden E, Pillai M, Campos M,
McElvaney NG and Foronjy RF: α1-Antitrypsin activates protein
phosphatase 2A to counter lung inflammatory responses. Am J Respir
Crit Care Med. 190:1229–1242. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Rosen E, Fatanmi OO, Wise SY, Rao VA and
Singh VK: Gamma-tocotrienol, a radiation countermeasure, reverses
proteomic changes in serum following total-body gamma irradiation
in mice. Sci Rep. 12:33872022. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Rosen E, Fatanmi OO, Wise SY, Rao VA and
Singh VK: Tocol prophylaxis for total-body irradiation: A proteomic
analysis in murine model. Health Phys. 119:12–20. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Rithidech KN, Honikel L, Rieger R, Xie W,
Fischer T and Simon SR: Protein-expression profiles in mouse
blood-plasma following acute whole-body exposure to (137)Cs gamma
rays. Int J Radiat Biol. 85:432–447. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zutler M, Quinlan PJ and Blanc PD:
Alpha-1-antitrypsin deficient man presenting with lung function
decline associated with dust exposure: A case report. J Med Case
Rep. 5:1542011. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Bolund AC, Miller MR, Sigsgaard T and
Schlünssen V: The effect of organic dust exposure on long-term
change in lung function: A systematic review and meta-analysis.
Occup Environ Med. 74:531–542. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Liao SY, Lin X and Christiani DC:
Occupational exposures and longitudinal lung function decline. Am J
Ind Med. 58:14–20. 2015. View Article : Google Scholar : PubMed/NCBI
|