Introduction
Angiogenesis, the development of capillaries from
pre-existing blood vessels, is essential for the growth of
malignant tumours (1). The vascular
endothelial growth factor (VEGF) system plays a significant role in
regulating this process (2), and
the clinical benefit from targeting this system in patients with
metastatic colorectal cancer (mCRC) is well documented (3).
Pre-clinical studies demonstrated a role of
microRNAs (miRNAs) in the regulation of angiogenesis and the VEGF
system (4,5). miRNA-126 has been reported to play an
essential role in angiogenesis by modulating VEGFR-2-related signal
transduction through the RAS/ERK and PI3K/AKT pathways by
inhibiting regulatory units (6–9).
miRNAs are a group of small single-stranded
non-coding RNAs with a length of approximately 22 nucleotides. They
negatively regulate gene transcripts at a post-transcriptional
level by targeting mRNAs leading to mRNA degradation or
translational repression, depending on the complementarities
between miRNA and its targets (10). It has been demonstrated that one
miRNA may target several different mRNAs, and that one mRNA may be
targeted by several miRNAs; several other principles have been
suggested to explain miRNA-mediated gene regulation.
Approximately 1,000 unique human miRNAs have been
identified (11). Most of the
miRNAs are located in introns of mRNA transcripts, whereas others
have been identified in exons, in the 3′ untranslated region
(3′UTR) of a gene transcript or in non-coding transcripts,
including miRNA clusters. A large fraction of the identified miRNAs
are located in areas of the genome known to be involved in the
development of cancer and are consequently thought to play a
prominent role in malignant transformation (10). The expression levels of miRNAs are
often dysregulated in malignant tissue compared to the
corresponding normal tissue, which enables miRNAs to act as either
tumour suppressors or oncogenes depending on the function of their
target networks (10). An
increasing number of reports on the clinical significance of miRNAs
in various types of cancer, including CRC, have been published.
Differences in expression levels between CRC and normal colorectal
tissue have been described (12–14),
and certain studies have also provided evidence of prognostic
(15,16) and predictive (17) value related to miRNAs.
At present, little is known about the possible
relationship between miRNAs and the VEGF system in patients with
CRC. The aim of this study was to analyse the relationship between
miRNA-126, VEGFR-2 and neo-angiogenesis in patients with CRC.
Materials and methods
Study population
This study included 81 consecutive patients with CRC
during the period between February 2004 and July 2005, all
undergoing surgical resection for histologically verified
adenocarcinomas of the colon or rectum at the Department of
Surgery, Vejle Hospital, Denmark. Patients who had received
preoperative chemoradiation for rectal cancer were excluded.
Pre-treatment examinations included a chest X-ray and ultrasound or
CT scan of the abdomen. Postoperatively, the tumours were
histologically classified and staged according to the pTNM system.
Information regarding patient characteristics was based on patient
records and registries. The study was approved by the Regional
Scientific Ethical Committee for Southern Denmark according to
Danish law, J.nr. S-VF-20040047. Informed consent was obtained from
all patients enrolled in the study.
Tissue sampling
Immediately after surgery the removed bowel segment
was brought to the Department of Clinical Pathology and tissue from
the tumour was sampled by a pathologist. Samples for mRNA and miRNA
analyses were placed in RNAlater™ (Qiagen, CA, USA) and
stored at −20˚C. Samples for quantitative protein analysis were
frozen and stored at −80˚C. The samples were frozen within 30 min
of surgical removal. Samples intended for later
immunohistochemistry (IHC) and in situ hybridisation (ISH)
followed routine fixation and paraffin embedding. Samples for
quantitative protein analysis and IHC were sampled in close
proximity to each other. Based on a microscopic examination of
H&E-stained FFPE tumour sections, it was semi-quantitatively
estimated that the tissue used for quantitative protein analyses
was dominated by carcinoma cells (>50%).
VEGFR-2 mRNA and protein analyses
The VEGFR-2 gene expression and protein analyses
have previously been described (18). Briefly, tissue samples were
homogenised and total RNA was isolated according to the
RNeasy® mini handbook of June 2001 (Qiagen, MD, USA).
RNA was quantified using spectrophotometry (Eppendorf, Hamburg,
Germany) followed by cDNA synthesis using a M-MLV RT kit
(Invitrogen Co., Carlsbad, CA, USA). The geometrical mean of
β-2-microglobulin and β-actin was used for normalisation. Gene
expression analyses were performed using fluorescence-based
real-time reverse transcription-polymerase chain reaction (RT-PCR).
The analyses were performed on the ABI PRISM 7900 HT fast real-time
PCR system, TaqMan (Applied Biosystems, Foster City, CA, USA).
After protein extraction, VEGFR-2 was analysed using
Quantikine ELISA kits (DVR200; R&D Systems, Minneapolis, MN,
USA). The controls were also purchased from R&D Systems. The
assay employs the quantitative sandwich enzyme immunoassay
technique. Results are presented in pg/mg of total protein. Tissue
samples, standards and controls were assayed in duplicate and the
mean was recorded. The total coefficients of variation on three
levels (low, medium and high concentrations of VEGFR-2) were 11.3,
10.1 and 8.3%, respectively.
CD105 and caldesmon immunostaining and
MVD counting
The staining and counting procedures have previously
been described (19). Briefly, MVD
was measured using tissue sections stained by antibodies against
CD105 and caldesmon in order to visualise immature microvessels. A
kit from Dako (Glostrup, Denmark; code K5361, EnVision™ G|2
doublestain system, rabbit/mouse, DAB+/permanent red) was used for
the detection of primary antibodies. The anti-CD105 antibody was
obtained from Novocastra (Bristol, UK; endoglin NCL-CD105, clone
4G11) and used at a dilution of 1:25. The anti-caldesmon antibody
was obtained from Dako (code M3557, clone h-CD) and used at a
dilution of 1:50.
Microvessels were counted at the invasive tumour
front by two observers unaware of the clinical parameters. The mean
MVD from three hotspots was used for statistical analysis. Any
stained endothelial cell or endothelial cell cluster clearly
separated from adjacent microvessels by tumour cells and/or stroma
elements was considered a single countable microvessel. Vessel
lumen was not necessary for a structure to be counted as a
microvessel. Consensus counts resolved any discrepancy between the
observers. Regarding the CD105 (brown) and caldesmon (red)
staining, only microvessels without associated red staining were
counted in order to preferentially estimate the MVD of the immature
vessels.
miRNA-126 gene expression analyses
miRNA quantification was carried out using
predesigned TaqMan miRNA assays (Applied Biosystems). cDNA
synthesis, pre-amplification and qPCR were performed according to
the manufacturer's instructions. In brief, RNA was reversely
transcribed using the TaqMan miRNA reverse transcription kit in
combination with the stem-loop Megaplex™ primer pool A v2.0,
allowing simultaneous reverse transcription of 377 unique miRNAs.
Subsequently, the Megaplex RT product was pre-amplified using
TaqMan PreAmp master mix and Megaplex PreAmp primer pool A v2.0.
RT-PCR amplification of miRNA-126 (part no. 4395339) was performed
in TaqMan Universal PCR master mix. Cycling conditions were as
follows: 95˚C for 10 min followed by 40 cycles of 95˚C for 15 sec
and 60˚C for 1 min. PCR reactions were performed on the ABI PRISM
7900 HT Fast RT-PCR system (Applied Biosystems). Raw Cq values were
calculated using the SDS software v2.1 using automatic baseline and
threshold settings. Expression levels of Hsa-miR-340 (part no.
4427975) were used for normalisation, since this miRNA was found by
the NormFinder algorithm (20), a
rigorous statistical approach for evaluating expression variation
of normalisation gene candidates, to be the most stably expressed
transcript throughout the tissue samples.
miRNA-126 in situ hybridisation
ISH was performed as described elsewhere (21). In brief, ISH was carried out on 6-μm
tissue sections containing CRC and normal colorectal tissue using
double DIG-labelled LNA probes for human miRNA-126 (Exiqon A/S,
Vedbaek, Denmark). The DIG-labelled probe was detected with
alkaline phosphatase-conjugated sheep anti-DIG Fab fragments
followed by NBT-BCIP chromogenic staining and nuclear fast red
staining.
Statistical analysis
The Fisher's exact test was used for two-group
comparisons. Median values were compared using the Wilcoxon rank
sum test. Disease-free survival (DFS) was defined as the time from
surgery until the first documented tumour recurrence or death.
Progression-free survival (PFS) was defined as the time from
surgery until the first documented tumour recurrence, progression
or death. Overall survival (OS) was defined as the time from
surgery until death. Survival curves were determined according to
the Kaplan-Meier method, and the log-rank test was used to test for
differences between the groups. Survival data from patients
diagnosed with a new malignancy following their surgical resection
for CRC (5 patients) were censored from the date of their new
cancer diagnosis. These data were excluded to prevent any potential
bias related to the presence of a new cancer or the
chemotherapeutic treatments used. Statistical calculations were
carried out using NCSS statistical software (NCSS Statistical
Software, version 2007; Kaysville, UT, USA). P<0.05 was
considered to be statistically significant and all tests were
two-sided.
Results
Patient characteristics
The patient characteristics, along with miRNA-126
expression levels in CRC obtained by PCR analysis, are shown in
Table I. Female patients presented
with a significantly lower median miRNA-126 expression in their
tumour tissue compared to their male counterparts. Furthermore,
non-significant associations were observed with regard to age at
diagnosis and tumour localisation. Younger patients and patients
with left-sided colon cancers presented with a higher miRNA-126
expression. No differences were observed between the patient
characteristics, as listed in Table
I, and gene or protein levels of VEGFR-2 or MVD. The 5-year
survival rate for the entire cohort was 60%.
 | Table IPatient characteristics. |
Table I
Patient characteristics.
| No. (%)
(n=81) | miRNA-126 |
|---|
|
|---|
| Median (95% CI) | p-value |
|---|
| Gender | | | 0.04 |
| Male | 37 (46) | 1.00 (0.87–1.28) | |
| Female | 44 (54) | 0.83 (0.59–1.08) | |
| Age (years) | | | 0.09 |
| Mean (SD) | 71.7 (11.1) | | |
| Range | 47–91 | | |
| >mean | 44 (54) | 0.85 (0.89–1.48) | |
| <mean | 37 (46) | 1.02 (0.89–1.48) | |
| T category | | | 0.72 |
| 1–3 | 63 (78) | 0.97 (0.82–1.12) | |
| 4 | 18 (22) | 0.79 (0.59–1.48) | |
| N category | | | 0.81 |
| 0 | 46 (57) | 0.99 (0.72–1.13) | |
| 1–2 | 35 (43) | 0.96 (0.77–1.28) | |
| M category | | | 0.57 |
| 0 | 65 (80) | 0.96 (0.81–1.12) | |
| 1 | 16 (20) | 0.90 (0.70–1.69) | |
| Stage | | | 0.73 |
| I–II | 43 (53) | 1.01 (0.68–1.13) | |
| III–IV | 38 (47) | 0.95 (0.77–1.28) | |
| Localisation | | | |
| Rectum | 20 (25) | 0.88 (0.70–1.48) | 0.88 |
| Colon | 61 (75) | 0.98 (0.77–1.23) | 0.07 |
| Left colon | 28 (46) | 1.05
(0.94–1.47) | |
| Right colon | 33 (54) | 0.77
(0.56–1.13) | |
| MSI status | | | 0.33 |
| MSI | 15 (19) | 0.72
(0.38–1.24) | |
| MSS | 66 (81) | 0.96
(0.82–1.08) | |
| Tumour grade | | | 0.74 |
| 1 and 2 | 61 (75) | 0.95
(0.79–1.08) | |
| 3 | 20 (25) | 0.96
(0.59–1.58) | |
| Vascular
invasion | | | 0.42 |
| Yes | 10 (12) | 1.08
(0.70–1.73) | |
| No | 71 (88) | 0.96
(0.79–1.02) | |
| Neuronal
invasion | | | 0.18 |
| Yes | 10 (12) | 1.32
(0.70–1.87) | |
| No | 71 (88) | 0.94
(0.77–1.02) | |
| Peritoneal
perforation | | | 0.33 |
| Yes | 17 (21) | 0.77
(0.59–1.36) | |
| No | 64 (79) | 0.97
(0.83–1.12) | |
miRNA-126 expression
Paired samples of CRC and normal colorectal tissue
were obtained from 28 patients. The median gene expression of
miRNA-126 in CRC, 0.85 (95% CI, 0.71–0.97), did not differ
significantly from the median gene expression in normal colorectal
tissue, 1.01 (95% CI, 0.80–1.11), p=0.56. The distribution of
miRNA-126 expression levels in the CRC samples for the entire
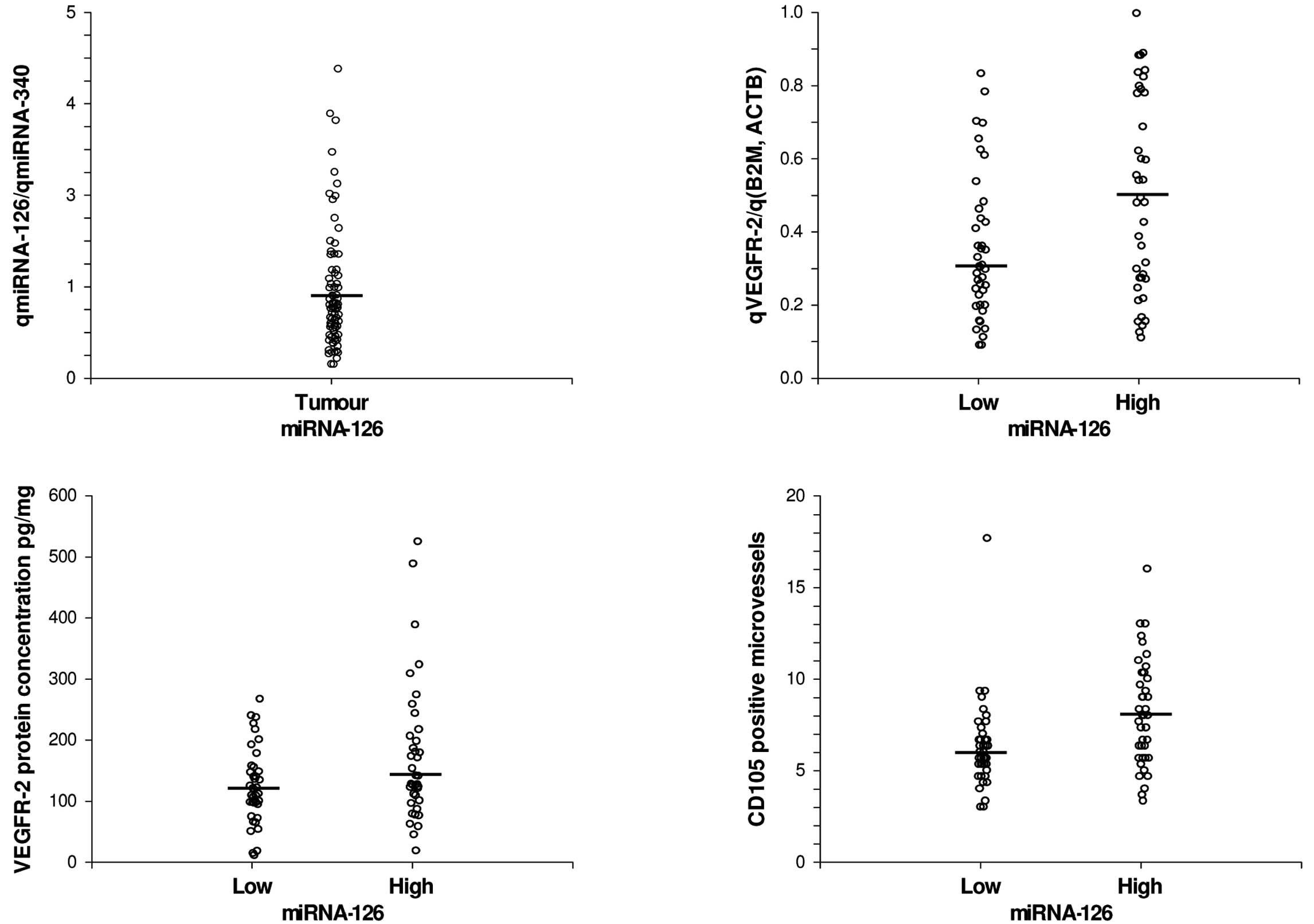
patient cohort is shown in Fig. 1A.
Tumours were classified as low or high miRNA-126-expressing tumours
using the median as a cut-off.
The distribution of VEGFR-2 gene expression in CRC
based on miRNA-126 levels is shown in Fig. 1B. The median gene expression level
of VEGFR-2 was significantly lower in the tumours expressing low
levels of miRNA-126, 0.30 (95% CI, 0.24–0.36), compared to those
expressing high levels, 0.48 (95% CI, 0.28–0.60), p=0.02.
The distribution of VEGFR-2 protein concentrations
in CRC based on miRNA-126 levels is shown in Fig. 1C. The median protein concentration
of VEGFR-2 was slightly lower, although not significant, in the
tumours expressing low levels of miRNA-126, 115 pg/mg (95% CI,
98–140), compared to those expressing high levels, 134 pg/mg (95%
CI, 121–180), p=0.06.
The median MVD was significantly lower in the
tumours expressing low levels of miRNA-126, 5.8 (95% CI,
5.33–6.67), compared to those expressing high levels, 8.0 (95% CI,
6.33–9.00), p<0.01 (Fig.
1D).
miRNA-126 in situ hybridisation
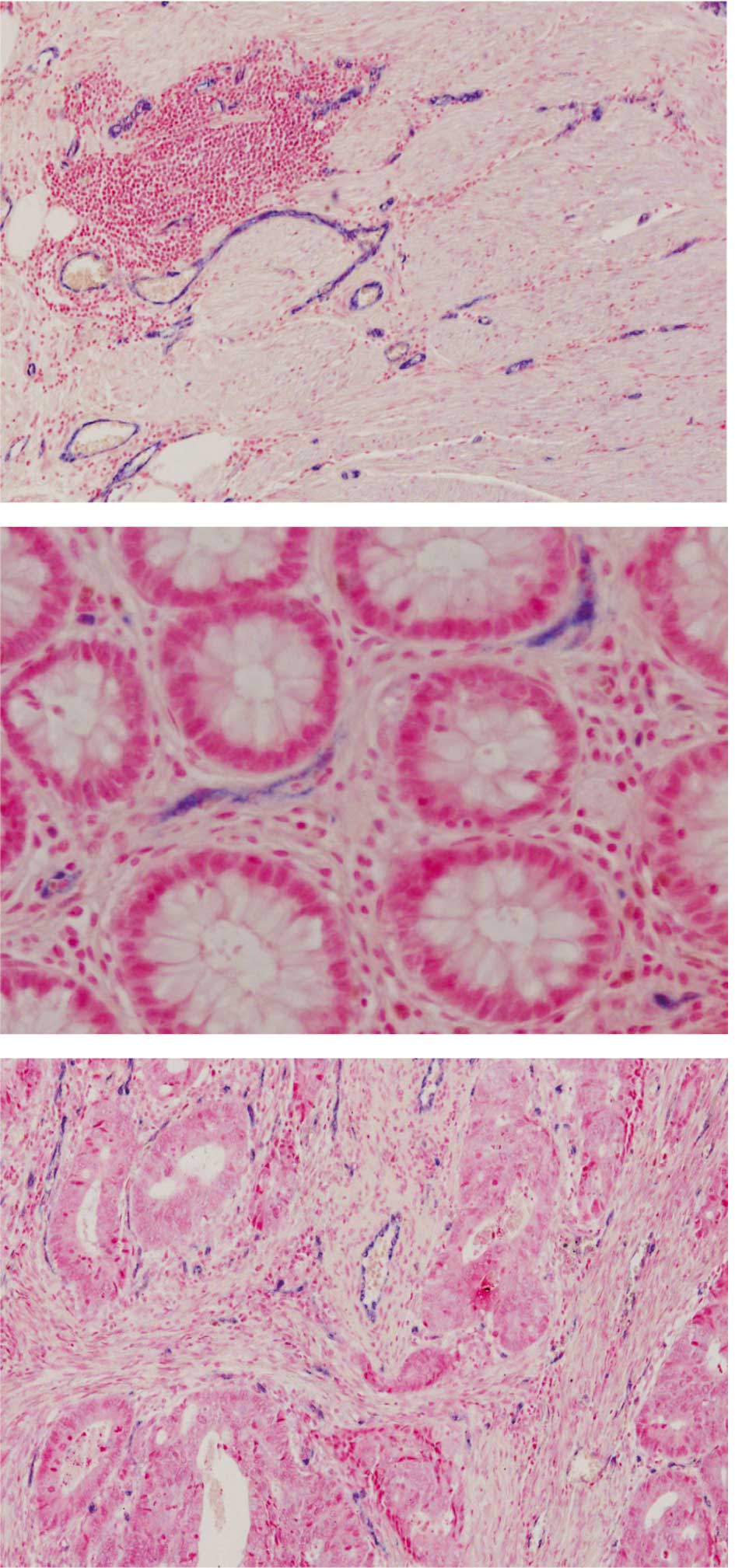
ISH analyses using high-affinity LNA probes revealed
intense ISH signals in endothelial cells. miRNA-126-positive
vessels were observed in the tumour stroma, as well as in the
lamina propria of normal mucosa and in the deep unaffected bowel
wall (Fig. 2).
miRNA-126 and prognosis
The possible prognostic value of miRNA-126
expression in CRC was assessed by comparing patients with a low
expression of miRNA-126 to those with a high expression, using the
median as the cut-off. Neither PFS [hazard ratio 0.85 (95% CI,
0.44–1.64)] nor OS [hazard ratio 0.99 (95% CI, 0.49–1.98)] differed
significantly between the two groups (p=0.64 and p=0.98,
respectively).
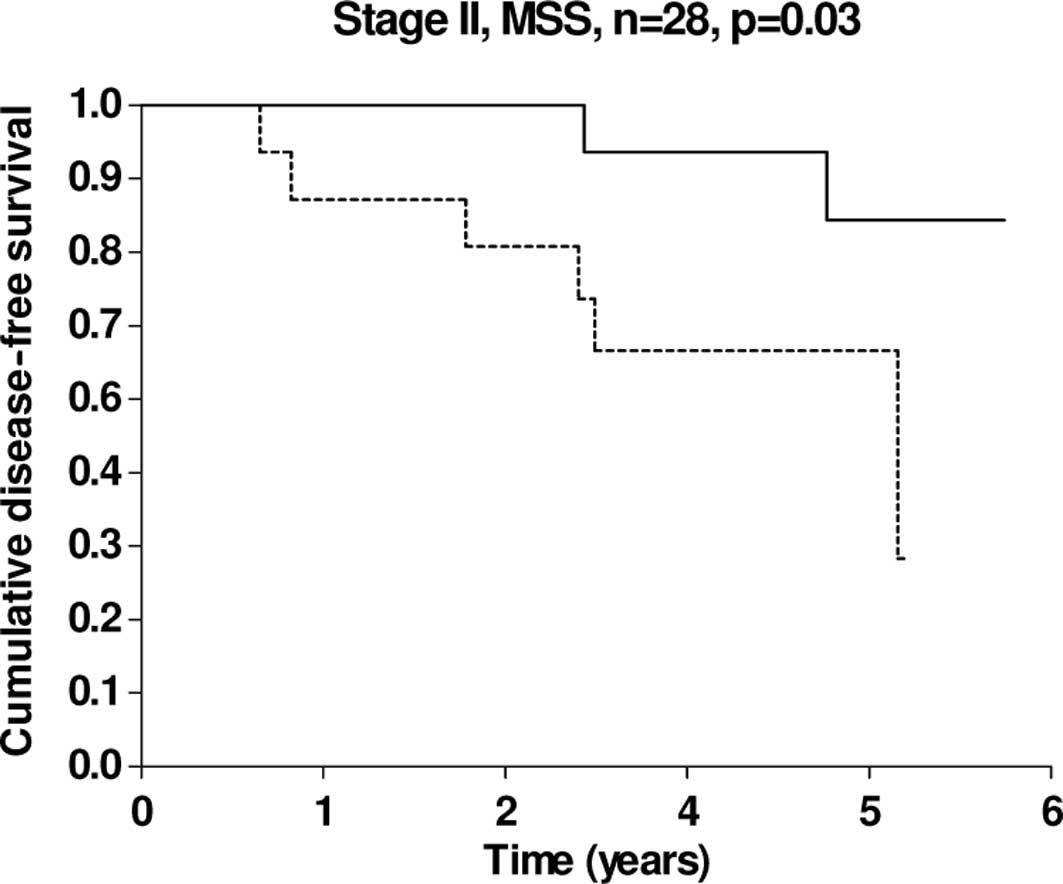
Our patient cohort included a total of 28 patients
with microsatellite stable (MSS) stage II CRC and, within this
subgroup, short DFS was significantly linked to high miRNA-126
expression levels (Fig. 3).
Discussion
In the present pilot study, a direct correlation was
found between miRNA-126, VEGFR-2 and angiogenesis in samples from
patients with CRC.
Paired samples of CRC and normal colorectal tissue
were obtained from 28 patients, and the median miRNA-126 expression
level in the two tissue types did not differ significantly. Guo
et al (7) demonstrated that
the miRNA-126 level was significantly lower in colon cancer
compared to normal colon tissue in a panel of 6 paired samples. In
both cases, the results are based on a rather limited number of
paired samples and should therefore be interpreted with caution.
Furthermore, the use of different normalisation methods (Guo et
al used 5S rRNA) may also be significant in this context.
The miRNA-126 expression in CRC was significantly
lower in females compared to males, and a possible, albeit
non-significant, difference was also observed regarding age and
tumour localisation. The miRNA-126 expression tended to be lower in
the older group of patients and in patients with right-sided colon
cancers.
Based on these findings, a correlation with
microsatellite instable (MSI) status was expected. However, no such
correlation was found in the present study. Díaz et al
(15) analysed several miRNAs in
110 patients who underwent surgery for CRC. miRNA-126 expression
was found to be significantly lower in the youngest group of
patients and no significant association with gender was documented.
These discrepancies are difficult to explain since the two studies
involved a similar number of patients, included Caucasians only,
used almost the same methodology, although Díaz et al used
5S for normalisation, and also included patients with stage I–IV
disease. The study by Díaz et al used fresh-frozen tumour
tissue compared to the present study in which miRNA was extracted
from tissue preserved in RNAlater. A possible effect on
miRNA-126 expression levels by directly comparing preservation in
RNAlater and freezing of the tissue specimens remains to be
clarified.
A high miRNA-126 expression was correlated to a high
gene expression of VEGFR-2 and likely also to high protein
concentrations of VEGFR-2 in CRC, although the latter correlation
did not reach statistical significance (p=0.06). Furthermore, a
high miRNA-126 expression was significantly correlated to a higher
density of newly formed micro vessels. These results, obtained in a
clinical setting, suggest a relationship between the VEGF system,
neo-angiogenesis and miRNA-126, and correlate well with results
reported in the pre-clinical literature. Two recent studies
(6,9) have demonstrated the significant role
of miRNA-126 in media-ting developmental angiogenesis and
identified two specific targets for miRNA-126, i.e., the
sprouty-related, EVH1 domain-containing protein 1 (SPRED-1) and the
phospho-inositol-3 kinase regulatory subunit 2 (PIK3R2), both
acting as negative regulators of the RAS/ERK and PI3K/AKT pathways,
respectively. miRNA-126 is capable of potentiating the angiogenic
signalling downstream of VEGFR-2 by modulating the signals through
these pathways. Other studies have demonstrated a similar
regulatory role of miRNA-126 with regard to these pathways
(7,22). In 2010, Nicoli et al
(8) published results suggesting
that blood flow stimulates angiogenesis in the zebrafish by a
genetic pathway in which a transcription factor (klf2a) induces the
expression of miRNA-126 and thereby enhances VEGF-A signalling.
These pre-clinical studies, confirm that high levels of miRNA-126
increase signalling downstream of VEGFR-2, ultimately resulting in
increased angiogenesis. To a certain extent, this increase is
supported by the present results obtained in a clinical
setting.
Our results showed a relationship between miRNA-126,
VEGFR-2 and neo-angiogenesis estimated by CD105 MVD, suggesting
that miRNA-126 expression was correlated to tissue vascularisation.
The miRNA-126 ISH analyses revealed an expression in endothelial
cells in malignant and normal colorectal tissue similar to results
from various in vitro studies (5,6,8,9).
These observations indicate high and specific miRNA-126 expression
levels in endothelial cells. The clinical value of the miRNA-126
ISH analyses, which may be an alternative to the MVD technique
based on IHC, deserves further analysis using larger patient
cohorts.
The possible prognostic value of miRNA-126
expression in CRC was analysed. However, no significant correlation
with survival was found in the unselected patient cohort, which
corresponds to the results obtained by Díaz et al (15). A subgroup analysis consisting of
patients with stage II disease only and MSS tumours is relevant due
to the need to identify new prognostic markers in stage II CRC, and
the well-known prognostic difference between MSI and MSS tumours
(23) justifies this selection.
Although this subgroup only consisted of 28 patients in the present
study, it is significant that our observations are consistent with
the findings reported by Schepeler et al (16). These authors demonstrated that
miRNA-126, among other miRNAs, was up-regulated in the primary
tumour tissue from patients with stage II MSS colon cancers who
experienced relapse significantly later compared to those that did
not. A meaningful multivariate survival analysis was not conducted
due to the small sample size. Therefore, although miRNA-126
expression in CRC does not appear to be of prognostic value in the
entire patient cohort, it may harbour a prognostic impact for a
selected group of CRC patients. The meta-analysis by Des et
al demonstrated a correlation between high MVD and poor
prognosis in CRC (24). The present
relationship between high MVD and high miRNA-126 expression in the
present study provides a plausible explanation for these findings,
although this hypothesis requires validation in a larger patient
cohort.
In conclusion, significant associations were found
between miRNA-126 levels in CRC, VEGFR-2 gene expression and
neo-angiogenesis, although a great overlap was observed between the
compared groups. Conclusions should therefore be drawn with
caution, and additional testing in larger and more homogeneous
study populations is necessary to validate the results. The present
results correspond well with the pre-clinical literature,
indicating a pivotal role of miRNA-126 in the modulation of
angiogenesis (6–9,25).
Thus, miRNA-126 may harbour predictive information with regard to
anti-angiogenetic therapy. A better understanding of how this miRNA
is regulated is of great significance. Further studies that analyse
the clinical value of miRNA-126 in CRC, are therefore required.
Acknowledgements
The authors are grateful for the technical
assistance provided by Lone Frischknecht, Lone Hartmann Hansen,
Sara Egsgaard, Birgit Roed Sørensen, Pamela Celis and Karin Larsen.
This study was supported by The Cancer Foundation, which had no
influence on any part of the study.
References
|
1
|
Folkman J: What is the evidence that
tumors are angiogenesis dependent? J Natl Cancer Inst. 82:4–6.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wagner AD, Arnold D, Grothey AA, Haerting
J and Unverzagt S: Anti-angiogenic therapies for metastatic
colorectal cancer. Cochrane Database Syst Rev.
8:CD0053922009.PubMed/NCBI
|
|
4
|
Hua Z, Lv Q, Ye W, et al: MiRNA-directed
regulation of VEGF and other angiogenic factors under hypoxia. Plos
One. 1:E1162006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuehbacher A, Urbich C, Zeiher AM and
Dimmeler S: Role of Dicer and Drosha for endothelial microRNA
expression and angiogenesis. Circ Res. 101:59–68. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fish JE, Santoro MM, Morton SU, Yu S, Yeh
RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY and Srivastava D:
miR-126 regulates angiogenic signaling and vascular integrity. Dev
Cell. 15:272–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo C, Sah JF, Beard L, Willson JK,
Markowitz SD and Guda K: The noncoding RNA, miR-126, suppresses the
growth of neoplastic cells by targeting phosphatidylinositol
3-kinase signaling and is frequently lost in colon cancers. Genes
Chromosomes Cancer. 47:939–946. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nicoli S, Standley C, Walker P, Hurlstone
A, Fogarty KE and Lawson ND: MicroRNA-mediated integration of
haemodynamics and Vegf signalling during angiogenesis. Nature.
464:1196–1200. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang S, Aurora AB, Johnson BA, Qi X,
McAnally J, Hill JA, Richardson JA, Bassel-Duby R and Olson EN: The
endothelial-specific microRNA miR-126 governs vascular integrity
and angiogenesis. Dev Cell. 15:261–271. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Esquela-Kerscher A and Slack FJ: Oncomirs
– microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269.
2006.
|
|
11
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Faber C, Kirchner T and Hlubek F: The
impact of microRNAs on colorectal cancer. Virchows Arch.
454:359–367. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Michael MZ, O'Connor SM, van Holst
Pellekaan NG, Young GP and James RJ: Reduced accumulation of
specific microRNAs in colorectal neoplasia. Mol Cancer Res.
1:882–891. 2003.PubMed/NCBI
|
|
14
|
Yamamichi N, Shimomura R, Inada K, et al:
Locked nucleic acid in situ hybridization analysis of miR-21
expression during colorectal cancer development. Clin Cancer Res.
15:4009–4016. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Díaz R, Silva J, Garcia JM, Lorenzo Y,
Garcia V, Pena C, Rodriguez R, Munoz C, Garcia F, Bonilla F and
Dominguez G: Deregulated expression of miR-106a predicts survival
in human colon cancer patients. Genes Chromosomes Cancer.
47:794–802. 2008.PubMed/NCBI
|
|
16
|
Schepeler T, Reinert JT, Ostenfeld MS, et
al: Diagnostic and prognostic microRNAs in stage II colon cancer.
Cancer Res. 68:6416–6424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schetter AJ, Leung SY, Sohn JJ, Zanetti
KA, Bowman ED, Yanaihara N, Yuen ST, Chan TL, Kwong DL, Au GK, Liu
CG, Calin GA, Croce CM and Harris CC: MicroRNA expression profiles
associated with prognosis and therapeutic outcome in colon
adenocarcinoma. Jama. 299:425–436. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hansen TF, Spindler KL, Lorentzen KA,
Olsen DA, Andersen RF, Lindebjerg J, Brandslund I and Jakobsen A:
Quantitative analysis of vascular endothelial growth factor
receptors 1 and 2 in colorectal cancer. Mol Med Reports. 2:787–792.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hansen TF, Sorensen FB, Spindler KL, Olsen
DA, Andersen RF, Lindebjerg J, Brandslund I and Jakobsen A:
Microvessel density and the association with single nucleotide
polymorphisms of the vascular endothelial growth factor receptor 2
in patients with colorectal cancer. Virchows Arch. 456:251–260.
2010. View Article : Google Scholar
|
|
20
|
Andersen CL, Jensen JL and Orntoft TF:
Normalization of real-time quantitative reverse transcription-PCR
data: a model-based variance estimation approach to identify genes
suited for normalization, applied to bladder and colon cancer data
sets. Cancer Res. 64:5245–5250. 2004. View Article : Google Scholar
|
|
21
|
Jorgensen S, Baker A, Moller S and Nielsen
BS: Robust one-day in situ hybridization protocol for
detection of microRNAs in paraffin samples using LNA probes.
Methods. 52:375–381. 2010.
|
|
22
|
Kuhnert F, Mancuso MR, Hampton J,
Stankunas K, Asano T, Chen CZ and Kuo CJ: Attribution of vascular
phenotypes of the murine Egfl7 locus to the microRNA miR-126.
Development. 135:3989–3993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ionov Y, Peinado MA, Malkhosyan S, Shibata
D and Perucho M: Ubiquitous somatic mutations in simple repeated
sequences reveal a new mechanism for colonic carcinogenesis.
Nature. 363:558–561. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Des GG, Uzzan B, Nicolas P, Cucherat M,
Morere JF, Benamouzig R, Breau JL and Perret GY: Microvessel
density and VEGF expression are prognostic factors in colorectal
cancer. Meta-analysis of the literature. Br J Cancer. 94:1823–1832.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kuehbacher A, Urbich C and Dimmeler S:
Targeting microRNA expression to regulate angiogenesis. Trends
Pharmacol Sci. 29:12–15. 2008. View Article : Google Scholar
|