Introduction
In the last decade, progress has been made in the
treatment of patients with gastric cancer, which is one of the most
prevalent malignancies worldwide. However, a significant percentage
of gastric cancer patients fail to achieve complete remission,
leading to a relapse or poor prognosis. Metastasis and multidrug
resistance (MDR) have been regarded as major mechanisms underlying
tumor relapse following successful surgical resections (1–3).
Metastasis is the advanced stage of solid tumor progression and is
a multi-step process that involves invasion, intravasation,
extravasation to distant organs and finally growth of the secondary
tumor. Chemotherapy is an important option in tumor treatment and
contributes to the prevention of cancer metastasis and recurrence
for surgical resection patients, but MDR is a significant clinical
problem associated with chemotherapy (4,5).
Chromatin architecture plays a significant role in
the regulation of nuclear function and is crucial for the global
changes in gene expression during tumor progression (6). Special AT-rich binding protein 1
(SATB1) is a well-characterized matrix attachment region
(MAR)-binding protein. SATB1 organizes the chromatin into distinct
loops via periodic anchoring of MARs to the nuclear matrix and
functionally interacts with chromatin modifiers to suppress gene
expression through histone deacetylation and nucleosome remodeling
(7–10). SATB1 has been shown to directly
regulate genes involved in cell proliferation, including c-myc
(11,12). Furthermore, previous studies have
suggested that SATB1 is a master regulator of breast cancer growth
and metastasis (1,13). Notably, Li et al recently
reported that SATB1 was able to upregulate certain genes associated
with MDR and suppress drug-induced apoptosis in breast cancer,
indicating the role of SATB1 in MDR (14).
Immunohistochemical assays revealed that the
expression of SATB1 was higher in gastric cancer tissues than in
normal gastric mucosa (15).
Nevertheless, few studies have addressed the potential role of
SATB1 in gastric cancer. Therefore, we investigated the role of
SATB1 in gastric cancer with a particular focus on the biological
characteristics of gastric cancer cells that are associated with
MDR and metastasis.
Materials and methods
Cell culture
The human gastric adenocarcinoma cell line SGC-7901
(maintained in our laboratory) and its corresponding VCR-resistant
variant SGC7901/VCR (obtained from the Xiangya Center Laboratory,
Changsha, Hunan, China) were cultured in DMEM medium supplemented
with 10% fetal bovine serum (FBS), 100 U/ml penicillin and 100
mg/ml streptomycin in a 37°C incubator with 100% humidity and 5%
CO2. In addition, to maintain the MDR phenotype, the
drug-resistant cells were stimulated with 1 μg/ml VCR.
Plasmid construction and
transfection
The coding region of SATB1 (GeneBank Accession
number NM_002971) was amplified using the primers: forward:
5′-CCGACAACCACTACCTGA-3′ and reverse: 5′-GTGGCACCTTCCAGGGTC-3′, and
subcloned into the mammalian expression vector pEX-M29
(GeneCopoeia, Rockville, MD, USA) to generate the plasmid
pEX-SATB1. The recombinant construction was verified by DNA
sequencing. pEX-SATB1 was prepared using the Fastfilter Endo-Free
Plasmid Midi kit (Omega Bio-Tek, Norcross, GA, USA) according the
manufacturer's instructions and transfected into SGC7901 or
SGC7901/VCR cells using lipofectamine 2000 (Invitrogen, Carlsbad,
CA, USA) according to the manufacturer's instructions. The cells
were harvested after 36 or 48 h for the following experiments.
RNA interference
Three SATB1 siRNA oligonucleotides and one negative
control sequence were designed based on the SATB1 sequence and
synthesized by Guangzhou RiboBio Co., Ltd. (Guangzhou, China).
SGC7901 cells were seeded at 2×105 cells/well in 6-well
plates and cultured in serum-free medium. The following day, when
the cells reached 70% confluence, 50 nM siRNA was transfected into
the cells using lipofectamine 2000 (Invitrogen) according to the
manufacturer's instructions. After 6 h, the cells were cultured in
DMEM medium supplemented with 10% FBS. The cells were harvested at
the indicated time points for the subsequent experiments.
RT-PCR assay
Total RNA was isolated from the cultured cells using
TRIzol reagent (Takara Bio, Inc., Shiga, Japan) according to the
manufacturer's instructions. RT-PCR was performed using the
StepOnePlus™ Real-Time PCR system (Applied Biosystems, Carlsbad,
CA, USA). The primers used were: SATB1, forward:
5′-GTGGGTACGCGATCAACTGA-3′ and reverse: 5′-TGTTAAAGCCAGTGCAA-3′;
GAPDH (internal control), forward: 5′-TTGGTATCGTGGAAGGACTCA-3′ and
reverse: 5′-TGTCATCATATTTGGCAGGTT-3′. The PCR cycling conditions
were denaturation at 48 cycles of 95°C for 30 sec, annealing at
52°C for 40 sec and extension at 72°C for 40 sec. Finally, the PCR
products were visualized on a 1.0% agarose gel containing 5 μg/ml
ethidium bromide.
Western blotting
The cells were lysed with RIPA lysis buffer (Santa
Cruz Biotechnology, Santa Cruz, CA, USA) and the protein
concentration of the cell lysates was quantitated using the BCA
method. Equal amounts of protein were loaded and separated via 10%
SDS-PAGE, then transferred onto nitrocellulose membranes
(Millipore, Billerica, MA, USA). The membranes were blocked with 5%
non-fat milk for 2 h and then incubated with the SATB1 antibody
(1:1,000 dilution; Epitomics, Burlingame, CA, USA) and β-actin
antibody (1:2,000 dilution, Sigma, St. Louis, MO, USA) at 4°C
overnight. Following three washes with TBST, the membranes were
incubated with HRP-conjugated secondary antibodies (Boster, Wuhan,
China) for 4 h at room temperature. The membranes were developed
using an ECL kit (Santa Cruz Biotechnology) and exposed to X-ray
film. β-actin was used as a loading control.
In vitro invasion assay
The invasion behaviors of SGC7901 and its variant
cells were detected using a Matrigel transmembrane invasion assay
as described in a previous study (16). Briefly, 15 μl of ice-cold Matrigel
(Sigma) was spread over the transwell chambers (8-μm pore size,
Millipore) and incubated for 1 h at room temperature. Cells
(1×105 cells/well) were washed with PBS twice and then
transferred to the upper chamber with 200 μl serum-free medium. The
lower chamber was filled with serum containing medium with the
addition of fibronectin as the chemoattractant. The cells were
allowed to invade across the Matrigel-coated membrane for 48 h at
37°C in 5% CO2. The filter was removed gently and the
cells on the upper surface were eliminated by wiping with a cotton
swab. The cells that had invaded across the Matrigel and attached
to the bottom were fixed and stained with hematoxylin and eosin
(H&E). The stained cells were counted under a microscope in 10
randomly selected high-power fields. The assay was performed for a
minimum of three times independently.
In vitro drug sensitivity assay
The sensitivity of cells to VCR was evaluated using
3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)
assay as described previously (17). Briefly, cells in the logarithmic
phase were seeded in 96-well plates at a density of
1×104 cells/well. The following day, the cells were
cultured in serum containing medium supplemented with a different
concentration of VCR. Following 72 h, 50 μl MTT (Sigma) was added
into each well. After incubation for 4 h, the supernatant was
replaced with 150 μl dimethyl sulfoxide (Sigma). The optical
density was read at a wavelength of 490 nm on a spectrophotometer
(MPR-2100, Syntron, Carlsbad, CA, USA). The relative inhibitory
rate on cell growth of different concentrations of VCR was
calculated according to the following formula:
R=(V1-V2)/V2,
where R equaled the relative inhibitory rate, V1 was the
absorbance value of the control cells without VCR treatment and
V2 was the absorbance value in the presence of VCR.
Flow cytometric analysis of
apoptosis
The apoptotic rate was determined in SGC7901 and its
variant cells using flow cytometry of propidium iodide (PI)-stained
cells on a FACS Calibur flow cytometer (Becton Dickinson, San Jose,
CA, USA). The cells were plated into 24-well plates and then
treated with 1 μg/ml VCR the following day. After two days, the
pEZ-SATB1 plasmid or siRNA was tansfected into the cells. The cells
were harvested, washed with PBS, fixed in 70% ethanol overnight at
4°C and stained with 50 mg/ml PI in the dark at room temperature
for 30 min. The sub-G1 peak was measured and analyzed
using Cell Quest software (Becton Dickinson).
Intracellular VCR concentration
analysis
Flow cytometry was used to detect the fluorescence
intensity of intracellular VCR as described previously (18). Cells in the log phase were seeded in
six-well plates at a density of 1×106 cells/well and
cultured overnight at 37°C. The cells were then incubated with 5
μg/ml VCR for 1 h. The cells were trypsinized and harvested for the
detection of VCR accumulation, or alternatively cultured in
RPMI-1640 for a further 30 min to detect the retention. Following
washing twice with ice-cold PBS, the cells were analyzed by flow
cytometry with an excitation wavelength of 488 nm and an emission
of 575 nm to detect the intracellular VCR concentration.
Statistical analysis
Statistical analyses were performed using SPSS
software (Chicago, IL, USA). Quantitative values are shown as the
mean ± SD. ANOVA and the Student's t-test were used to compare the
differences. P<0.05 was considered to indicate a statistically
significant result.
Results
Expression of SATB1 is upregulated in MDR
gastric cancer cells
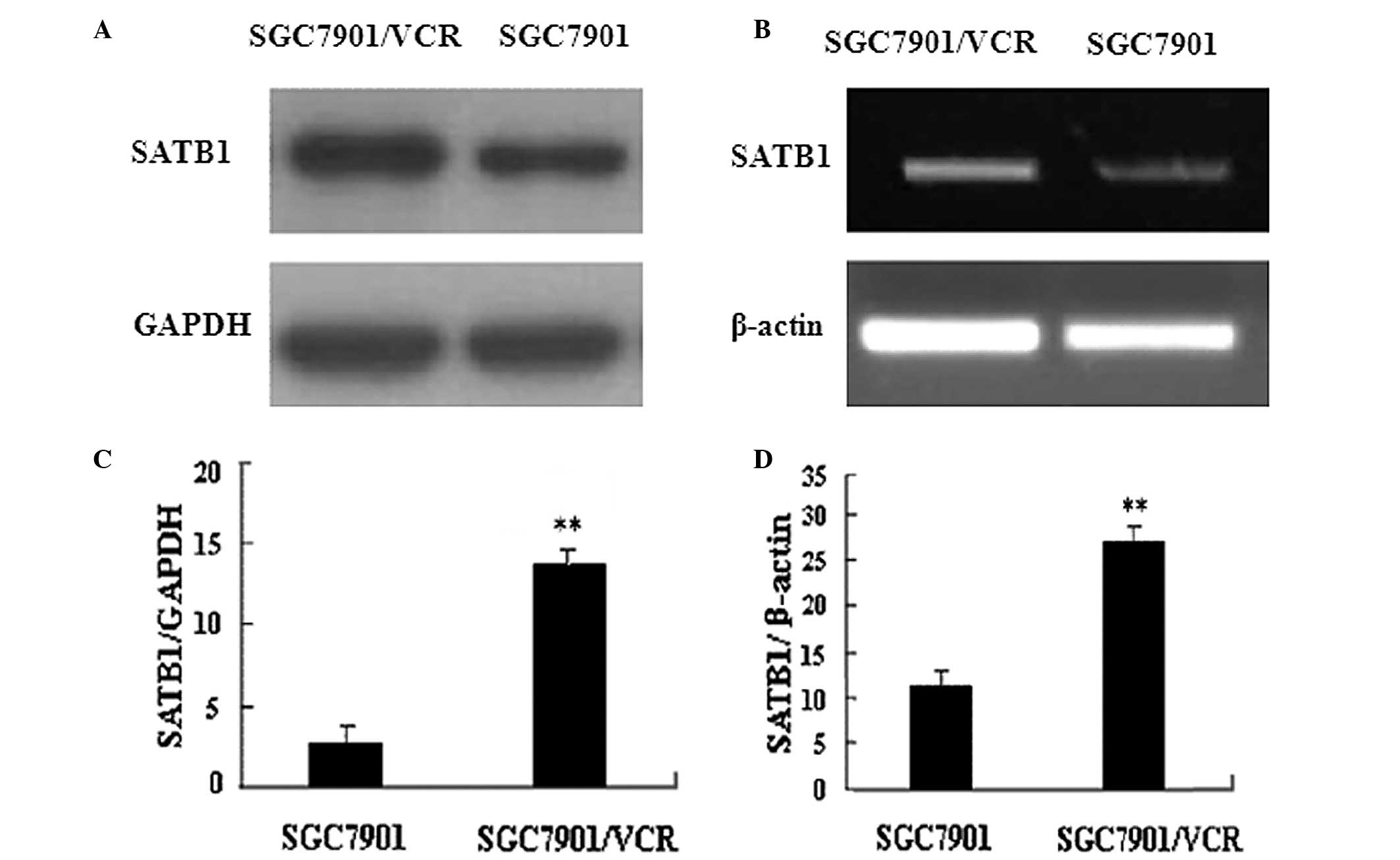
This study compared SATB1 expression in
VCR-resistant SGC7901/VCR cells and parent SGC7901 cells. RT-PCR
and western blot analysis results revealed that the expression of
SATB1 was higher in SGC7901/VCR cells than that in SGC7901 cells at
the mRNA and protein levels (Fig.
1). These data suggest that SATB1 is associated with the
development of MDR in gastric cancer cells.
SATB1 regulates MDR in gastric cancer
cells
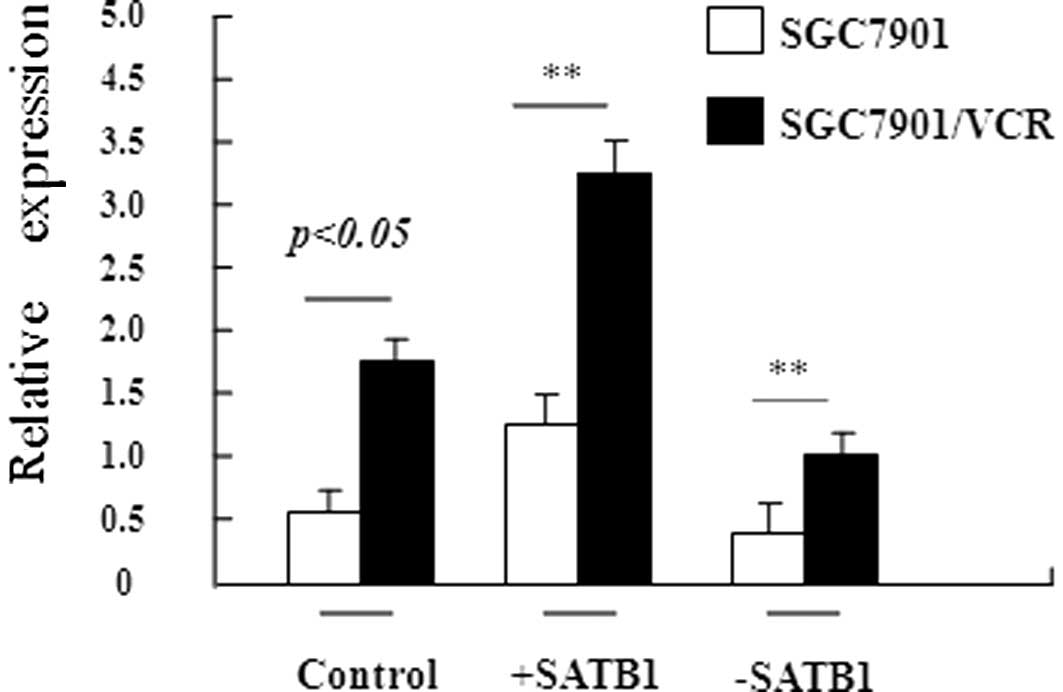
To confirm that SATB1 is involved in the development
of MDR in gastric cancer, stable gastric cancer cell lines were
constructed in which SATB1 was overexpressed (SGC7901-SATB1 and
SGC7901/VCR-SATB1 cells) or depleted (SGC7901-siRNASATB1 and
SGC7901/VCR-siRNASATB1 cells). RT-PCR revealed that the expression
of SATB1 at the mRNA level was high, as expected in these stable
cell lines (Fig. 2). The
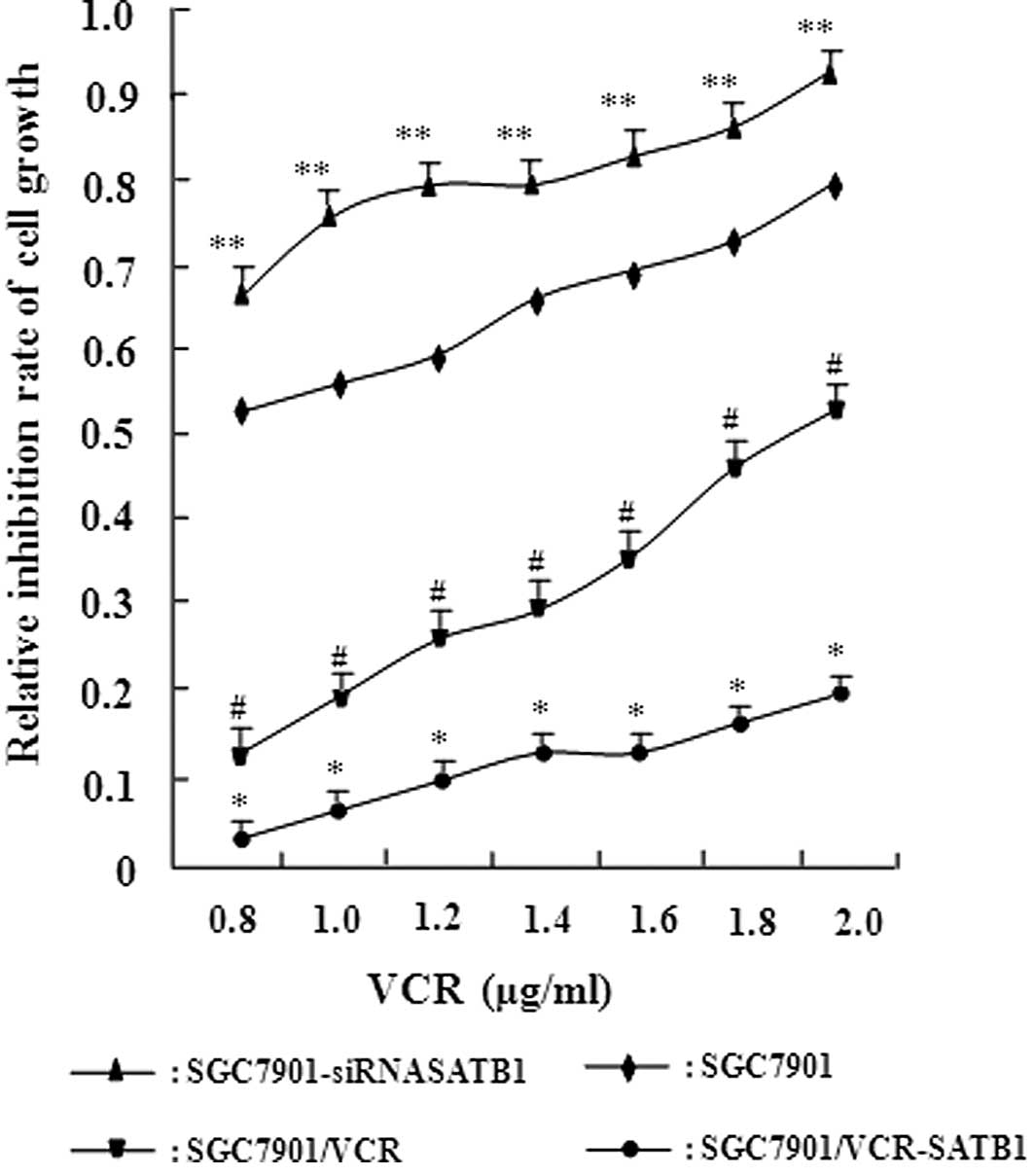
sensitivity of these cell lines to VCR was examined using the in
vitro MTT assay (Fig. 3).
Table I shows the IC50
values of these cells to VCR based on the MTT assay. Compared with
SGC7901 cells, SGC7901-siRNASATB1 cells exhibited a significantly
increased IC50 value for VCR (P<0.05). A significant
discrepancy in the IC50 value was also observed between
SGC7901 and its MDR SGC7901/VCR cells (P<0.01). Moreover, a
higher SATB1 expression in these cells was associated with a higher
IC50 value. These results demonstrate the positive
correlation between the level of SATB1 expression and drug
resistance capacity in gastric cancer, suggesting that SATB1
contributes to MDR in gastric cancer.
 | Table IIC50 values indicating the
sensitivity of the gastric cancer cell line SGC7901 and its
variants to VCR. |
Table I
IC50 values indicating the
sensitivity of the gastric cancer cell line SGC7901 and its
variants to VCR.
| Cell lines | IC50 value
(μg/ml; mean ± SD) |
|---|
| SGC7901 | 0.85±0.11 |
| SGC7901-SATB1 | 1.53±0.45a |
|
SGC7901-siRNASATB1 | 0.31±0.11 |
| SGC7901/VCR | 3.15±0.37a |
|
SGC7901/VCR-SATB1 | 4.66±0.38 |
|
SGC7901/VCR-siRNASATB1 | 1.61±0.15b |
SATB1 expression level is negatively
correlated with intracellular VCR accumulation in gastric cancer
cells
To explore the mechanism by which SATB1 contributes
to MDR in gastric cancer, the effect of SATB1 on intracellular VCR
accumulation and retention was examined. Flow cytometry was
performed to determine the fluorescence intensity of intracellular
VCR in SGC7901 and its variant cell lines which expressed different
levels of SATB1. The results showed that intracellular VCR
accumulation in SGC7901/VCR cells was significantly less than that
in the SGC7901 cells (P<0.01, Table
II). Moreover, the concentration of VCR in the
SGC7901/VCR-SATB1 cells was one-seventh of that in the SGC7901
cells, while the concentration of VCR was significantly increased
in SGC7901-siRNASATB1 cells compared with SGC7901 cells
(P<0.05). Taken together, these data show that the SATB1
expression level is negatively correlated with intracellular VCR
accumulation in gastric cancer cells, which may explain how SATB1
contributes to MDR in gastric cancer.
| Table IIIntracellular VCR accumulation and
retention in the gastric cancer cell line SGC7901 and its
variants. |
Table II
Intracellular VCR accumulation and
retention in the gastric cancer cell line SGC7901 and its
variants.
| Cell lines | Accumulation | Retention |
|---|
| SGC7901 | 74.48±5.54 | 52.71±6.59 |
|
SGC7901-siRNASATB1 | 86.32±4.48a | 33.75±3.41a |
| SGC7901-SATB1 | 64.25±3.60 | 33.75±3.41 |
| SGC7901/VCR | 28.14±1.58c | 15.85±1.70c |
|
SGC7901/VCR-siRNASATB1 | 33.56±5.87 | 18.97±1.20 |
|
SGC7901/VCR-SATB1 | 20.56±2.31b | 8.54±1.05b |
SATB1 protects gastric cancer cells from
VCR-induced apoptosis
It is widely accepted that protection against
drug-induced apoptosis contributes to the development of MDR in
cancer. Therefore, the effect of SATB1 on VCR-induced apoptosis was
investigated in SGC7901 and SGC7901/VCR cells and their variants.
Apoptotic cells were quantified as those that had a DNA content of
<2 N (sub-G1 DNA content) (19). Results of the flow cytometric
analysis revealed that SGC7901 cells had a higher apoptotic index
than SGC7901/VCR cells (P<0.05). In addition, compared with
SGC7901 and SGC7901/VCR cells, the apoptotic index was decreased in
the cells which overexpressed SATB1 but increased in the cells in
which SATB1 expression was depleted by siRNA (P<0.05, Fig. 4). These results indicate that SATB1
was able to inhibit the VCR-induced apoptosis of gastric cancer
cells.
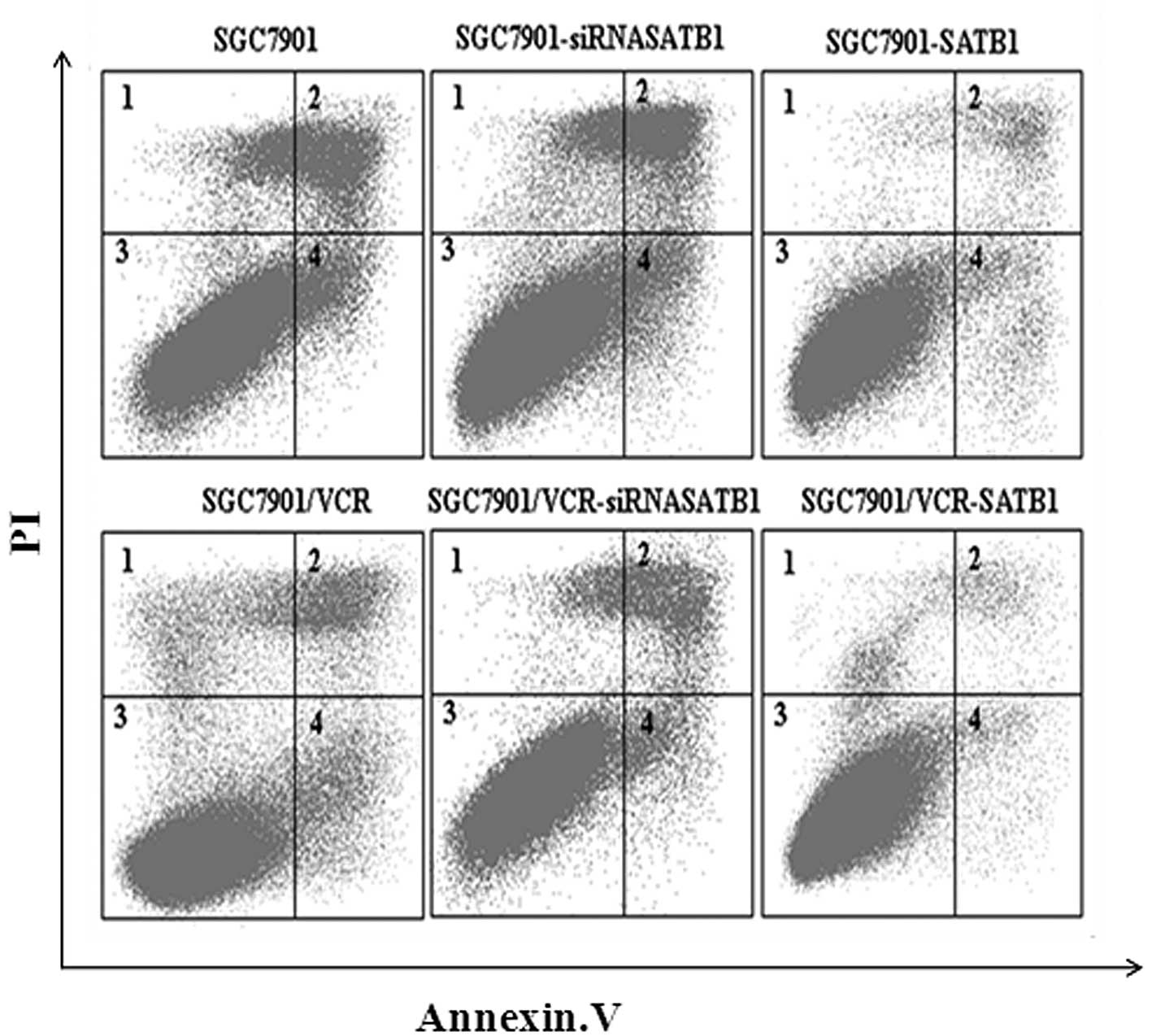 | Figure 4SATB1 protects gastric cancer cells
from VCR-induced apoptosis. The cells were incubated with 1 μg/ml
VCR for 36 h and the apoptotic index was calculated based on flow
cytometry and Annexin V/PI staining. The apoptotic index was 19.8,
35.0, 8.6, 10.6, 16.8 and 4.7%, for SGC7901, SGC7901-siRNASATB1,
SGC7901-SATB1, SGC7901/VCR, SGC7901/VCR-siRNASATB1 and
SGC7901/VCR-SATB1 cells, respectively. SATB1, special AT-rich
sequence binding protein 1; VCR, vincristine; PI, propidium
iodide. |
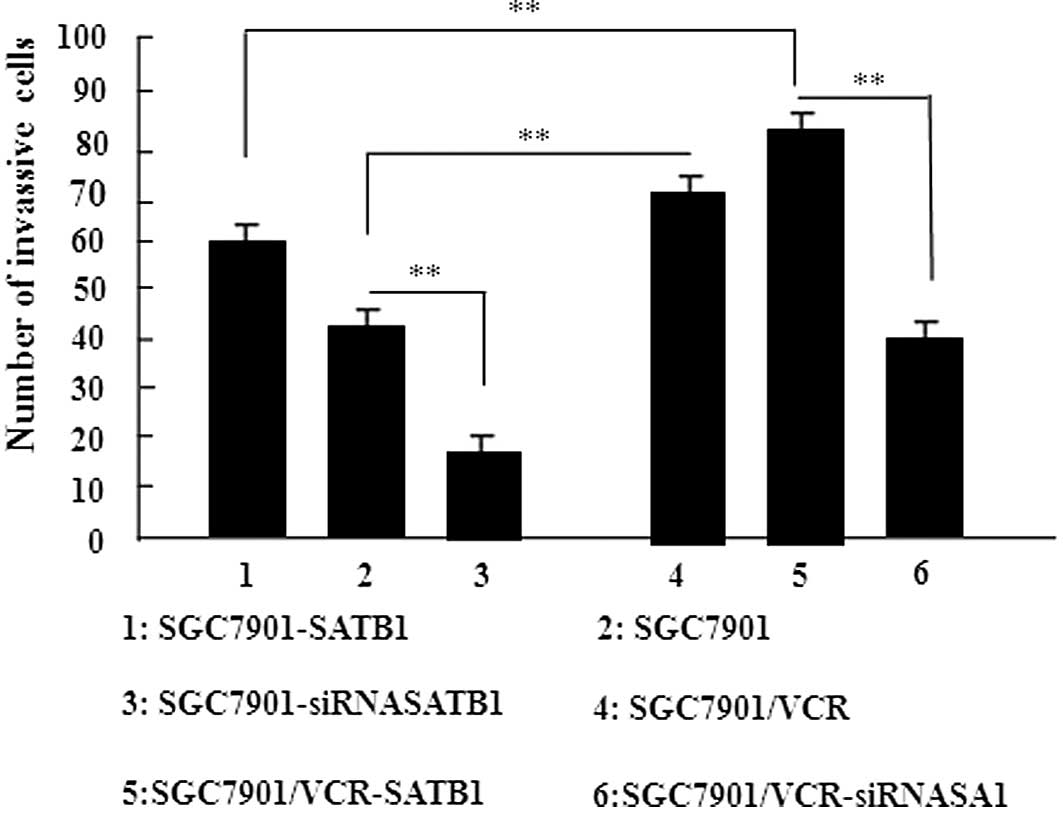
SATB1 promotes the invasion of gastric
cancer cells
An in vitro invasion assay was performed to
investigate the role of SATB1 in gastric cancer invasion. The
results of this study revealed that the overexpression of SATB1
significantly promoted the migration of cells into the lower
transwell chamber while siRNA-mediated depletion of SATB1
significantly inhibited cell migration into the lower chamber,
compared with the control cells (P<0.05, Fig. 5). These data demonstrate that SATB1
promotes the invasion of gastric cancer cells.
Discussion
SATB1 was originally isolated as a nuclear factor
that specifically binds to the BUR sequence (9). More recent studies have revealed that
SATB1 is involved in the reconstruction of global genomes and
regulates the expression of more than 1,000 genes. There is
evidence to suggest that SATB1 modulates cell proliferation and
lineage development and SATB1 is significant in breast cancer
(7,11,12,20).
The mechanism by which SATB1 contributes to the malignant
phenotypes of tumor cells is poorly understood.
MDR is known to be a significant aspect of malignant
cancer and remains the major obstacle to the efficacy of cancer
chemotherapy. Extensive research over the past decades has improved
the understanding of the development of MDR in cancer cells. For
example, over 60 genes have been identified as being upregulated in
the drug-resistant human gastric cancer cell lines SGC7901/VCR and
SGC7901/ADR (adriamycin) (21).
However, no further studies have provided the mechanistic insight
into the regulation of these genes.
Considering that SATB1 is a global regulator of gene
expression, we speculate that the role of SATB1 is to regulate MDR
in gastric cancer. The results of the present study support the
role of SATB1 in MDR in gastric cancer. Western blotting and RT-PCR
analysis were performed and we observed that SATB1 is overexpressed
in VCR-resistant SGC7901 cells. Moreover, it was observed that the
level of SATB1 expression was positively correlated with drug
resistance in SGC7901 cells. Collectively, these findings led to
the conclusion that SATB1 contributes to the development of MDR in
gastric cancer.
To explore the mechanism by which SATB1 contributes
to MDR in gastric cancer, the effect of SATB1 on intracellular VCR
accumulation and retention was examined. MDR has been investigated
extensively for a number of years and a variety of mechanisms for
MDR have been proposed, including increased drug efflux, decreased
drug influx, DNA repair activation, detoxification and blockage of
apoptosis. These mechanisms are not exclusive and may contribute to
MDR in combination. Among them, the blockage of apoptosis and
increased drug efflux are regarded as the most significant
(22). Therefore, in this study,
flow cytometry was performed to analyze the apoptotic rate in
SGC7901-derived cells and it was observed that the expression level
of SATB1 was negatively correlated with the apoptotic index in
gastric cancer cells. Notably, VCR-induced apoptosis was largely
eliminated by SATB1 overexpression in SGC7901 cells. These results
are consistent with previous studies which revealed that SATB1
inhibited apoptosis in Sézary and breast cancer cells (14,23),
suggesting that the SATB1-mediated inhibition of apoptosis may
contribute to MDR in gastric cancer.
Enhanced outward transport and reduced intracellular
accumulation of anticancer drugs have been identified as major
causes of MDR in certain tumors (24,25).
The present study revealed that VCR accumulation and retention
occurred less frequently in SGC7901/VCR than in SGC7901 cells.
Moreover, compared with the parental cells, VCR accumulation was
inhibited in SATB1 overexpressing cells but was promoted in cells
that were depleted of SATB1. A similar finding has been reported in
breast cancer cells (25). ABC
transporters usually function as pumps that remove toxins and drugs
from the cells and p-glycoprotein (p-gp) and MDR-1 are the most
characterized ABC transporters (26). Studies should therefore be conducted
to investigate the effect of SATB1 on the expression of p-gp and
MDR-1, in order to elucidate whether SATB1 contributes to MDR by
modulating the expression of ABC transporters and the intracellular
accumulation of drugs.
Notably, a previous study revealed that the
multidrug transporter p-gp was involved in the migration and
invasion of resistant melanoma cells, indicating a potential link
between MDR and the invasive behavior of cancer cells (27). Therefore, the effect of SATB1 on the
invasive behavior of gastric cancer cells was further investigated.
An in vitro invasion assay demonstrated that gain or loss of
SATB1 function was associated with the increased or decreased
invasion activity of SGC7901 cells, providing complementary
evidence that SATB1 promotes the invasion of gastric cancer cells.
However, to provide further insight into the role of SATB1 in
cancer invasion and metastasis, the role of SATB1 in modulating the
expression of major genes involved in tumor invasion, including
matrix metalloproteinases, requires examination.
In conclusion, the results of the present study
reveal the upregulation of SATB1 in drug-resistant gastric cancer
cells and that SATB1 contributes to MDR by inhibiting the
accumulation of VCR in gastric cancer cells and protecting the
cells from VCR-induced apoptosis. Addtionally, the significant role
of SATB1 in the development of MDR in gastric cancer has been
elucidated. SATB1 was also able to promote the invasion of gastric
cancer cells. SATB1 may therefore be a promising target for the
therapy of drug-resistant and invasive gastric cancer.
Acknowledgements
This study was supported by grants from the National
Nature Science Foundation of China (no. 30901418 and no. 81172186).
We would like to thank all members of our laboratory for their
useful discussion.
References
|
1
|
Han HJ, Russo J, Kohwi Y and
Kohwi-Shigematsu T: SATB1 reprogrammes gene expression to promote
breast tumor growth and metastasis. Nature. 452:187–193. 2008.
View Article : Google Scholar
|
|
2
|
Parker B and Sukumar S: Distant metastasis
in breast cancer: molecular mechanisms and therapeutic targets.
Cancer Biol Ther. 2:14–21. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hao ZH, Li X, Qiao T, Du R, Hong L and Fan
D: CIAPIN1 congers multidrug resistance by upregulating the
expression of MDR-1 and MRP-1 in gastric cancer cells. Cancer Biol
Ther. 5:261–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ozben T: Mechanisms and strategies to
overcome multiple drug resistance in cancer. FEBS Lett.
580:2903–2909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pienta KJ and Smith DC: Advances in
prostate cancer chemotherapy: a new era begins. CA Cancer J Clin.
55:300–318. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spector DL: The dynamics of chromosome
organization and gene regulation. Annu Rev Biochem. 72:573–608.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cai S, Han HJ and Kohwi-Shigematsu T:
Tissue-specific nuclear architecture and gene expression regulated
by SATB1. Nature Genet. 34:42–51. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yasui D, Miyano M, Cai S, Varga-Weisz P
and Kohwi-Shigematsu T: SATB1 targets chromatin remodelling to
regulate genes over long distances. Nature. 419:641–645. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dickinson LA, Joh T, Kohwi Y and
Kohwi-Shigematsu T: A tissue-specific MAR/SAR DNA-binding protein
with unusual binding site recognition. Cell. 70:631–645. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Agrelo R, Souabni A, Novatchkova M,
Haslinger C, Leeb M, Komnenovic V, Kishimoto H, Gresh L,
Kohwi-Shigematsu T, Kenner L and Wutz A: SATB1 defines the
developmental context for gene silencing by Xist in lymphoma and
embryonic cells. Dev Cell. 16:507–516. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nakayama Y, Mian IS, Kohwi-Shigematsu T
and Ogawa T: A nuclear targeting determinant for SATB1, a genome
organizer in the T cell lineage. Cell Cycle. 4:1099–1106. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alvarez JD, Yasui DH, Niida H, Joh T, Loh
DY and Kohwi-Shigematsu T: The MAR-binding protein SATB1
orchestrates temporal and spatial expression of multiple genes
during T-cell development. Genes Dev. 14:521–535. 2000.PubMed/NCBI
|
|
13
|
Zheng J: Is SATB1 a master regulator in
breast cancer growth and metastasis? Womens Health (Lond Engl).
4:329–332. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li QQ, Chen ZQ, Xu JD, Cao XX, Chen Q, Liu
XP and Xu ZD: Overexpression and involvement of special AT-rich
sequence binding protein 1 in multidrug resistance in human breast
carcinoma cells. Cancer Sci. 101:80–86. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu X, Cheng C, Zhu S, Yang Y, Zheng L,
Wang G, Shu X, Wu K, Liu K and Tong Q: SATB1 is an independent
prognostic marker for gastric cancer in a Chinese population. Oncol
Rep. 24:981–987. 2010.PubMed/NCBI
|
|
16
|
Cheng GZ, Chan J, Wang Q, Zhang W, Sun CD
and Wang LH: Twist transcriptionally up-regulates AKT2 in breast
cancer cells leading to increased migration, invasion, and
resistance to paclitaxel. Cancer Res. 67:1979–1987. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li QQ, Wang WJ, Xu JD, Cao XX, Chen Q,
Yang JM and Xu ZD: Involvement of CD147 in regulation of multidrug
resistance to P-gp substrate drugs and in vitro invasion in breast
cancer cells. Cancer Sci. 98:1064–1069. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi Y, Zhai H, Wang X, Han Z, Liu C, Lan
M, Du J, Guo C, Zhang Y, Wu K and Fan D: Ribosomal proteins S13 and
L23 promote multidrug resistance in gastric cancer cells by
suppressing drug-induced apoptosis. Exp Cell Res. 296:337–346.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maxwell SA and Davis GE: Biological and
molecular characterization of an ECV-304-derived cell line
resistant to p53-mediated apoptosis. Apoptosis. 5:277–290. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wen J, Huang S, Rogers H, Dickinson LA,
Kohwi-Shigematsu T and Noguchi CT: SATB1 family protein expressed
during early erythroid differentiation modifies globin gene
expression. Blood. 105:3330–3339. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao Y, You H, Liu F, An H, Shi Y, Yu Q
and Fan D: Differentially expressed gene profiles between multidrug
resistant gastric adenocarcinoma cells and their parental cells.
Cancer Lett. 185:211–218. 2002. View Article : Google Scholar
|
|
22
|
Gottesman MM, Fojo T and Bates SE:
Multidrug resistance in cancer: role of ATP-dependent transporters.
Nat Rev Cancer. 2:48–58. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Su M, Zhou LL, Tu P, Zhang X,
Jiang X and Zhou Y: Deficiency of SATB1 expression in Sezary cells
causes apoptosis resistance by regulating FasL/CD95L transcription.
Blood. 117:3826–3835. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gorlick R and Bertino JR: Drug resistance
in colon cancer. Semin Oncol. 26:606–611. 1999.
|
|
25
|
Naito S, Yokomizo A and Koga H: Mechanisms
of drug resistance in chemotherapy for urogenital carcinoma. Int J
Urol. 6:427–439. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dong X and Mumper RJ: Nanomedicinal
strategies to treat multidrug-resistant tumors current progress.
Nanomedicine (Lond). 5:597–615. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Colone M, Calcabrini A, Toccacieli L,
Bozzuto G, Stringaro A, Gentile M, Cianfriglia M, Ciervo A,
Caraglia M, Budillon A, et al: The multidrug transporter
P-glycoprotein: a mediator of melanoma invasion? J Invest Dermatol.
128:951–971. 2008. View Article : Google Scholar
|