Introduction
Glioblastoma is the most common and fatal primary
tumor in the central nervous system. A combination of surgery,
radiotherapy and chemotherapy is widely used to treat gliomas,
particularly malignant gliomas. However, the prognosis of patients
suffering from malignant glioma remains poor, with a median
survival in the range of 12–15 months (1). Thus, the development of efficient
treatment therapies that specifically target glioma cells is
required.
MicroRNAs (miRNAs) are small, non-coding RNAs that
are ~18–24 nucleotides in length and are predicted to regulate the
expression of approximately one-third of all human genes at the
post-transcriptional and translational levels (2). One miRNA may target several genes, and
one gene may be targeted by multiple miRNAs, indicating that miRNAs
are involved in the modulation of a wide range of biological
processes, such as apoptosis (3).
Dysfunction of miRNAs has been reported to commonly occur in
several human cancers (4,5), including glioma and its aggressive
glioblastoma subtype (6,7). Modulating miRNA activities may provide
exciting opportunities for cancer therapy (8).
The miR-34 family (miR-34a, miR-34b and miR-34c),
which is described as a p53 effector, has antiproliferative and
pro-apoptotic functions (9,10). In particular, downregulated miR-34c
is a critical factor that contributes to malignancy in human
laryngeal carcinoma, while its overexpression inhibits cell
proliferation and induces apoptosis via targeting of c-Met
(11). miR-34c has two identified
mature miRNAs: miR-34c-3p and miR-34c-5p (12). Although miR-34c-3p and miR-34c-5p
have been established as tumor suppressors in a variety of tumors
(13,14), their targets and functions in glioma
are largely unknown.
In the present study, we analyzed the function of
miR-34c-3p and miR-34c-5p in human glioblastoma. U251 and U87
glioblastoma cell lines were transfected with miR-34c-3p or
miR-34c-5p mimics. The biological characteristics of U251 and U87
cells were evaluated after enhancing the expression of miR-34c-3p
or miR-34c-5p. Additionally, it was investigated whether the effect
of miR-34c-3p or miR-34c-5p was associated with the downstream gene
Notch2.
Materials and methods
Human sample
Formalin-fixed paraffin-embedded human glioma tissue
samples (n=18) and normal brain tissues (n=5) were obtained from
the Neurosurgery Department of The Second Hospital of Hebei Medical
University (Shi Jiazhuang, China). The glioma tissue samples were
divided into three groups according to the malignant grade,
including Grade II (n=6), Grade III (n=6) and Grade IV (n=6). Data
collection and analysis were approved by the ethics committee of
Hebei Medical University.
Cells and cell culture
Human malignant glioma cell lines, U87 and U251,
were purchased from the Chinese Academy of Sciences Cell Bank
(Shanghai, China). The human normal glial cell line HEB, originally
established by Kumar et al(15), was purchased from the Guangzhou
Institute of Biomedicine and Health, Chinese Academy of Sciences.
U251, U87 and HEB cell lines were grown in Dulbecco’s modified
Eagle’s medium (DMEM; Hyclone, Logan, UT, USA) enriched with 10%
fetal bovine serum (Hyclone) in a 37°C, 5% CO2
incubator.
Transient miRNA transfection
Synthetic, chemically modified miRNA mimics were
designed and synthesized by RiboBio (Guangzhou, China).
Double-stranded scrambled RNA was used as the negative control
(NC). For transfection, 2×105 cells were seeded into
each well of 24-well plates and grown for 24 h until they were
30–50% confluent. Cells were washed, placed in Opti-MEM and
transfected with oligonucleotides using Lipofectamine™ RNAiMAX
Transfection Reagent (Invitrogen Life Technologies, Carlsbad, CA,
USA) according to the manufacturer’s instructions. After 4 h, the
medium was changed to DMEM/F12 or DMEM-high glucose, respectively,
and cells were cultured at 37°C in 5% CO2.
Real-time PCR analysis
Total RNA was extracted using TRIzol (Invitrogen),
according to the manufacturer’s instructions. The first strand cDNA
was generated by reverse transcription, and then PCR amplification
was performed using a real-time PCR cycler (ABI PRISM®
7500 Sequence Detection System; Applied Biosystems, Foster City,
CA, USA). Expression of miR-34c-3p and miR-34c-5p were quantified
by miR-qRT PCR using SYBR Green RealtimePCR master mix (Toyobo Co.,
Ltd., Osaka, Japan). U6 was used as an internal control to
normalize RNA input. Primers for miR-34c-3p and miR-34c-5p were
designed as previously described (14). The results of real-time PCR were
analyzed by the ΔΔCT method: ΔCT=CTselected
gene-CTU6, ΔΔCT=ΔCTtherapy
group-ΔCTcontrol group, RQ (Relative
Quantitation)therapy group=2−ΔΔCT,
RQcontrol group=1.
MTS assay
The cell survival rate was evaluated using the MTS
assay (Promega, Madison, WI, USA). Transfected and control cells in
the log phase of growth were seeded in 96-well plates at a cell
density of 1×104/well in 100 μl DMEM/F12 or DMEM-high
glucose supplemented with 10% fetal bovine serum. For four
consecutive days, 10 μl MTS solution was added to each well and
mixed, the cells were incubated at 37°C in the 5% CO2
incubator for an additional 4 h, and the reaction was stopped by
lysing the cells with 200 μl DMSO for 20 min. Absorbance at 490 nm
was measured with a Multiskan FC Microplate Photometer (Thermo
Fisher Scientific, Logan, UT, USA) and data are expressed as the
percentage of the control. All MTS assays were performed in
triplicate for each group and experiments were repeated at least
twice to confirm the consistency of results.
Cell-cycle analysis
For cell cycle analysis, transfected and control
cells in the log phase of growth were harvested by trypsinization
48 h post-transfection, washed with PBS twice, fixed in 70% ethanol
overnight at 4°C and then incubated with 100 μg/ml DNase-free RNase
A (Sigma-Aldrich, St. Louis, MO, USA), 0.2% Triton X-100 and
stained with 50 μg/ml propidium iodide (PI; Sigma-Aldrich) at 4°C
for 30 min. A total of 104 nuclei were examined in a FACSCaliber
flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and DNA
histograms were analyzed by ModFit software (BD Biosciences).
Experiments were performed in triplicate.
Apoptosis assay
Apoptosis analysis was performed in transfected and
control cells by staining with the Annexin V-FITC Apoptosis
Detection kit (Abcam, Cambridge, MA, USA). In brief, cells were
harvested at a density of 1×106 cells/ml in 1X binding
buffer and stained with FITC-labeled Annexin V for 15 min at room
temperature. Cells were then resuspended in 0.5 ml in 1X binding
buffer and stained with 10 μl PI after centrifugation for 5 min at
1,000 × g. Samples were immediately analyzed using the FACSCaliber
flow cytometer (BD Biosciences). Data were analyzed by Cell Quest
software (BD Bioscience).
Transwell invasion assay
For the transwell invasion assay, we prepared 8
μm-pore Transwell polycarbonate insert chambers (BD Biosciences)
coated with 40 μl Matrigel (BD Biosciences). Following 2 h
incubation at 37°C, the Matrigel solidified. Then cells of the
transfected and control groups (1×105) in 100 μl of
serum-free DMEM/F12 or DMEM-high glucose were added into the upper
compartment of the chambers, and 600 μl conditioned medium from
U251 and U87 cells was used as the chemoattractant and placed in
the bottom chambers. Following 24 h of incubation at 37°C and 5%
CO2, the medium was removed from the upper chamber. The
non-invaded cells on the upper chamber were scraped off with a
cotton swab. The invaded cells on the lower membrane were fixed
with 4% paraformaldehyde and stained with 0.1% crystal violet
(Invitrogen Life Technologies) for 10 min. The number of invaded
cells was counted from three randomly selected visual fields, each
from the central and peripheral portion of the membrane, using an
inverted microscope (CKX41; Olympus, Tokyo, Japan) at ×200
magnification. Each experiment was repeated in triplicate.
Human Notch2 transcript
The human Notch2 transcript contains a 3751-bp
3′UTR. To explore the possible regulation of Notch2 by miRNAs,
in silico analysis of miRNAs predicted to target the 3′UTR
of its transcript was performed. Several online softwares,
including miRanda, TargetScan and PICTAR, predicted that the
sequence between nucleotides 2045 and 3073 is likely targeted by
miR-34c-3p and miR-34c-5p.
Western blot analysis
U251 and U87 cells were lysed in 1X cell lysis
buffer (R&D systems, Minneapolis, MN, USA) 48 h following
transfection. Homogenates were clarified by centrifugation at
14,000 × g for 15 min at 4°C, and protein concentration was
measured by NanoDrop (Thermo Scientific). Subsequently, 10% sodium
dodecyl sulfate polyacrylamide was used for gel electrophoresis,
then gels were transferred to polyvinylidene fluoride membranes
(Millipore, Billerica, MA, USA). After permeabilizing and blocking,
the membranes were incubated with primary antibody against Notch2
(1:1,000 dilution, Cell Signaling Technology, Inc., Danvers, MA,
USA), followed by incubation with HRP-conjugated secondary antibody
(1:1,000 dilution, Zymed, South San Francisco, CA, USA). Reactions
were developed with ECL or ECL plus (GE Healthcare, Amersham,
UK).
Statistical analysis
SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA)
was used for statistical analysis. Spearman’s rank correlation test
was used for association analysis between endogenous miR-34c-3p and
miR-34c-5p levels and pathological grading. Differences in means
were analyzed by the two-tailed t-test. All data are expressed as
the mean ± standard deviation. P<0.05 was considered to indicate
a statistically significant difference.
Results
miR-34c-3p and miR-34c-5p are
downregulated in malignant glioma
The miR-34c-3p and miR-34c-5p expression levels in
malignant glioma were measured. All human glioma tissues exhibited
lower levels of miR-34c-3p and miR-34c-5p as compared with normal
brain tissues, respectively. The level of miRNAs was negatively
correlated with the pathological grading of glioblastoma (Fig. 1A). Similarly, U251 and U87 cell
lines also exhibited a small amount of endogenous miR-34c-3p or
miR-34c-5p (Fig. 1B), indicating an
etiological role of miR-34c-3p and miR-34c-5p reduction in glioma
progression. Transfection of either pre-miR-34c-3p or
pre-miR-34c-5p mimics in U251 and U87 cells significantly increased
their respective levels, while transfection with scrambled miRNA
negative control (NC) had no effect (Fig. 1B).
miR-34c-3p inhibits U251 and U87 cell
proliferation, but miR-34c-5p only affects U251 cells
Glioblastoma cell proliferation was quantified in
vitro by MTS assay. Transfection of U251 and U87 cells with
miR-34c-3p mimics resulted in 78.42 and 72.37% growth inhibition,
compared with normal and NC groups (Fig. 2; P<0.05; 96 h after
transfection). However, transfection with miR-34c-5p only inhibited
the cell proliferation to 80.84% in U251 cell lines (P<0.05 vs.
normal; P<0.05 vs. NC), but not in U87 cell lines (P>0.05 vs.
normal; P>0.05 vs. NC). These results implied that miR-34c-3p
and miR-34c-5p may function as tumor suppressors in glioma cells
in vitro. A significant difference in proliferation
inhibition was only observed in U87 cells transfected with
pre-miR-34c-3p (72.37%) against cells transfected with
pre-miR-34c-5p (94.89%), suggesting alternative regulatory pathways
are involved (Fig. 2).
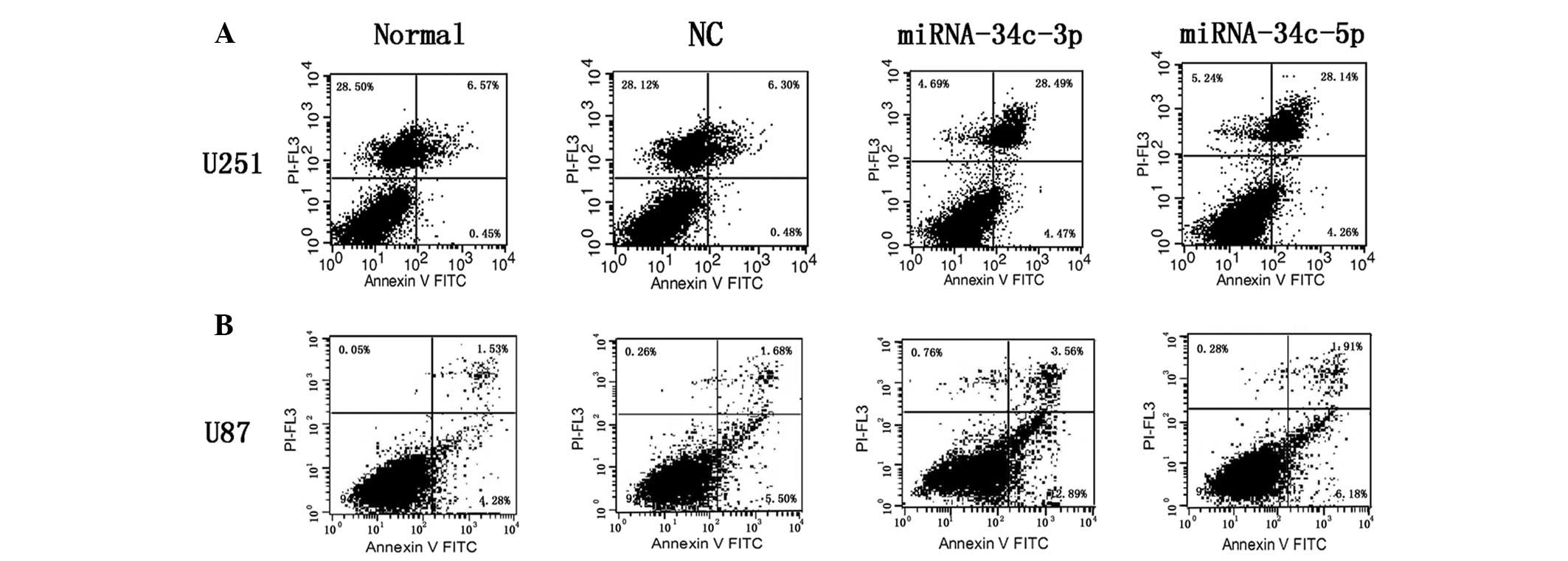
miR-34c-3p and miR-34c-5p overexpression
inducesapoptosis in glioma cell lines
To confirm apoptosis following miR-34c-3p or
miR-34c-5p overexpression in glioma cell lines, transfected cells
were analyzed for Annexin V expression by flow cytometry.
Statistically significant increases in Annexin V+ and
PI+ apoptotic cells were observed in miR-34c-3p (28.49%)
and miR-34c-5p (28.14%) mimic treatment groups in U251 glioblastoma
cells, compared with normal (6.57%) or NC (6.3%) groups (Fig. 3A). However, only miR-34c-3p-treated
cells underwent apoptosis in U87 cell lines (3.56%), but no
apoptosis was observed in miR-34c-5p-treated U87 (1.91%), normal
(1.53%) or NC (1.68%) cells (Fig.
3B).
Effect of miR-34c-3p and miR-34c-5p on
the cell cycle of U251 and U87 cell lines
The cell cycle distributions of normal, NC and
transfected cells were analyzed by flow cytometry. In U251 cells,
as shown in Fig. 4A, the G0/G1
phase fraction of normal and NC cells was 72.7 and 73.44%,
respectively. Administration of miR-34c-3p and miR-34c-5p mimics
decreased the percentage of cells in G0/G1 phase to 44.24 and
43.58%, respectively. However, the G2/M phase fraction in normal
and NC cells were 4.4 and 4.31%, respectively, while miR-34c-3p and
miR-34c-5p overexpression increased this fraction to 21.6 and
21.83%, respectively. Similarly, the percentages of cells in S
phase in normal and NC cells were lower than those of the
transfected cells.
In U87 cells, also shown in Fig. 4B, the G0/G1 phase fraction of normal
and NC cells was 69.3 and 75.62%, respectively. While miR-34c-3p
overexpression decreased the G0/G1 phase to 44.05%, transfected
miR-34c-5p did not affect the G0/G1 phase (79.07%). The S phase
fraction of normal and NC cells was 20.66 and 13.61%, respectively,
and this increased significantly with miR-34c-3p overexpression to
48.83%; however, this effect did not occur with miR-34c-5p
overexpression (11.04%). The G2/M phase fraction showed no
significant difference among the four groups.
These results suggested that transfection with
miR-34c-3p or miR-34c-5p in U251 cells and with miR-34c-3p in U87
cells produced S phase arrest with G0/G1 reduction. Although, no
significant changes were observed with miR-34c-5p transfection in
U87 cells or in the normal or NC groups. Cell cycle changes partly
explained the differences in cell proliferation, revealing
regulatory pathways involving cell division. Notably, miR-34c-3p
and miR-34c-5p treatment produced significant levels of apoptosis
relative to those of the controls, while no significant sub-G0
population was observed.
Cell invasive ability is depressed by
miR-34c-3p and miR-34c-5p mimic transfection
To evaluate the impact of miR-34c-3p and miR-34c-5p
expression on cell invasion, U251 and U87 cells were treated with
oligonucleotides and placed on 8 μm-pore size insert chambers
coated with a mixture of extracellular matrix proteins. The number
of U251 cells invading through the Matrigel following miR-34c-3p
and miR-34c-5p mimic treatment was 66.75±10.63 and 60.63±15.32
cells per field respectively, which was lower than that of the
normal (90.88±10.38 cells per field) and the NC (104.75±12.06 cells
per field) groups (Fig. 5A).
Similar results were observed in U87 cells, where invasive activity
was decreased to 10.63±2.19 and 11.63±4.75 cells per field in
miR-34c-3p and miR-34c-5p mimic-treated cells compared with the
normal (57.75±15.55 cells per field) or NC (44.25±19.78 cells per
field) cells (Fig. 5B). Our
experiments demonstrated that miR-34c-3p or miR-34c-5p
overexpression inhibited the invasive ability of U251 and U87
glioblastoma cells in vitro.
miR-34c-3p overexpression reduces
expression of Notch2, but miR-34c-5p overexpression does not
miR-34c-3p mimics reduced Notch2 expression by ~15%
compared to normal or NC cells (Fig.
6A) in U251 and U87 cell lines. By contrast, Notch2 expression
was not downregulated following transfection with miR-34c-5p mimics
(Fig. 6B). These data suggested
that the tumor suppressor activity of miR-34c-3p in glioblastoma
cells may be regulated by the Notch pathway, but that of miR-34c-5p
is not.
Discussion
It is well-documented that the mature miRNA-34
family, as tumor suppressors, shows a global decrease in expression
in many different human cancers, including laryngeal carcinoma
(11), prostate cancer (16) and cervical carcinoma (17). Although Luan et al have
demonstrated that miR-34a overexpression inhibited cell migration
and invasion in the glioma cell line, U251 (18), the effect of miR-34c on gliomas is
unknown.
In the present study, we profiled miR-34c-3p and
miR-34c-5p expression in glioblastoma and normal brain tissue. Our
data revealed that the level of these two miRNAs was significantly
decreased in glioblastoma compared to normal brain tissue.
Moreover, the miRNA values were negatively correlated with
aggressive behavior of glioblastoma. Overexpression of either
miR-34c-3p or miR-34c-5p in U251 cells caused inhibition of
proliferation, cell apoptosis, S phase arrest and cell invasion
suppression. However, this was not the case in U87 cells. The
proliferation inhibition effect of miR-34c-5p was not observed.
Furthermore, miR-34c-3p expression clearly produced cell apoptosis
and S phase arrest, while no apoptosis or major cell cycle changes
were observed with miR-34c-5p. These findings suggested that loss
of miR-34c-3p or miR-34c-5p expression may be critical in
glioblastoma pathogenesis. miR-34c-3p and miR-34c-5p may target
different mRNAs and thus display different results through
dissimilar pathways in U87 cells.
Many investigators have demonstrated that expression
of the miR-34 family resulted in G0/G1 cell cycle arrest in diverse
cellular contexts (19,20). The typical sub-G0 population would
occur as well as cell apoptosis. However, the present study showed
that expression of miR-34c-3p or miR-34c-5p in U251 and U87 cells
caused S phase arrest, indicating that the function of miR-34c was
different but complementary to that of miR-34a.
Previous studies have demonstrated that miR-34
family overexpression regulated proliferation and senescence
through inhibition of E2F3, BCL-2 and MYC (16,21,22).
Transient expression of miR-34a markedly inhibited glioma growth
in vivo by targeting c-Met and Notch (23). In the present study, we also
predicted that Notch2 was the precise intracellular target of
miR-34c by using miRanda, TargetScan and PICTAR databases, which
was different from that in a previous study (14). Notch signaling is key in the
regulation of brain tumor cell proliferation (24). In GBM and astrocytoma, Notch1 and
Notch2 are expressed at a high level, while the frequency and the
intensity of Notch2 expression is higher than that of Notch1
(25,26). In the present study, it was found
that the value of Notch2 was downregulated after transfection with
miR-34c-3p, indicating that the effect of miR-34c-3p on cell
proliferation inhibition occurred via the Notch2 pathway. However,
the level of Notch2 was not significantly different between the
miR-34c-5p and control groups. Hence, we hypothesize that the
mechanism of miR-34c-5p may be different from that of
miR-34c-3p.
To the best of our knowledge, this study provided
the first evidence of miR-34c-3p and miR-34c-5p values in glioma
patients’ tissues. Transfection with these two miRNAs inhibited the
cell proliferation, cell cycle changes, apoptosis and cell
invasion. Our findings suggest important roles of miR-34c-3p and
miR-34c-5p in glioma etiology and provide potential candidates for
treating malignant glioma. More experiments are required to further
validate the effect of miR-34c-3p on the Notch2 pathway.
References
|
1
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fu LL, Wen X, Bao JK and Liu B:
MicroRNA-modulated autophagic signaling networks in cancer. Int J
Biochem Cell Biol. 44:733–736. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lima RT, Busacca S, Almeida GM, Gaudino G,
Fennell DA and Vasconcelos MH: MicroRNA regulation of core
apoptosis pathways in cancer. Eur J Cancer. 47:163–174. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nan Y, Han L, Zhang A, et al: MiRNA-451
plays a role as tumor suppressor in human glioma cells. Brain Res.
1359:14–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sana J, Hajduch M, Michalek J, Vyzula R
and Slaby O: MicroRNAs and glioblastoma: roles in core signalling
pathways and potential clinical implications. J Cell Mol Med.
15:1636–1644. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho WC: MicroRNAs in cancer - from
research to therapy. Biochim Biophys Acta. 1805:209–217.
2010.PubMed/NCBI
|
|
9
|
Corney DC, Flesken-Nikitin A, Godwin AK,
Wang W and Nikitin AY: MicroRNA-34b and MicroRNA-34c are targets of
p53 and cooperate in control of cell proliferation and
adhesion-independent growth. Cancer Res. 67:8433–8438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He L, He X, Lim LP, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cai KM, Bao XL, Kong XH, et al:
Hsa-miR-34c suppresses growth and invasion of human laryngeal
carcinoma cells via targeting c-Met. Int J Mol Med. 25:565–571.
2010.PubMed/NCBI
|
|
12
|
Landgraf P, Rusu M, Sheridan R, et al: A
mammalian microRNA expression atlas based on small RNA library
sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cannell IG, Kong YW, Johnston SJ, et al:
p38 MAPK/MK2-mediated induction of miR-34c following DNA damage
prevents Myc-dependent DNA replication. Proc Natl Acad Sci USA.
107:5375–5380. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
López JA and Alvarez-Salas LM:
Differential effects of miR-34c-3p and miR-34c-5p on SiHa cells
proliferation apoptosis, migration and invasion. Biochem Biophys
Res Commun. 409:513–519. 2011.PubMed/NCBI
|
|
15
|
Kumar B, Hovland AR, Prasad JE, et al:
Establishment of human embryonic brain cell lines. In Vitro Cell
Dev Biol Anim. 37:259–262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hagman Z, Larne O, Edsjö A, et al: miR-34c
is downregulated in prostate cancer and exerts tumor suppressive
functions. International journal of cancer. Int J Cancer.
127:2768–2776. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pang RT, Leung CO, Ye TM, et al:
MicroRNA-34a suppresses invasion through downregulation of Notch1
and Jagged1 in cervical carcinoma and choriocarcinoma cells.
Carcinogenesis. 31:1037–1044. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luan S, Sun L and Huang F: MicroRNA-34a: a
novel tumor suppressor in p53-mutant glioma cell line U251. Arch
Med Res. 41:67–74. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ji Q, Hao X, Zhang M, et al: MicroRNA
miR-34 inhibits human pancreatic cancer tumor-initiating cells.
PloS One. 4:e68162009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tarasov V, Jung P, Verdoodt B, et al:
Differential regulation of microRNAs by p53 revealed by massively
parallel sequencing: miR-34a is a p53 target that induces apoptosis
and G1-arrest. Cell Cycle. 6:1586–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Christoffersen NR, Shalgi R, Frankel LB,
et al: p53-independent upregulation of miR-34a during
oncogene-induced senescence represses MYC. Cell Death Differ.
17:236–245. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kumamoto K, Spillare EA, Fujita K, et al:
Nutlin-3a activates p53 to both down-regulate inhibitor of growth 2
and up-regulate mir-34a, mir-34b, and mir-34c expression, and
induce senescence. Cancer Res. 68:3193–3203. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Guessous F, Zhang Y, Kofman A, et al:
microRNA-34a is tumor suppressive in brain tumors and glioma stem
cells. Cell Cycle. 9:1031–1036. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Solecki DJ, Liu XL, Tomoda T, Fang Y and
Hatten ME: Activated Notch2 signaling inhibits differentiation of
cerebellar granule neuron precursors by maintaining proliferation.
Neuron. 31:557–568. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sivasankaran B, Degen M, Ghaffari A, et
al: Tenascin-C is a novel RBPJkappa-induced target gene for Notch
signaling in gliomas. Cancer Res. 69:458–465. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fan X, Mikolaenko I, Elhassan I, et al:
Notch1 and notch2 have opposite effects on embryonal brain tumor
growth. Cancer Res. 64:7787–7793. 2004. View Article : Google Scholar : PubMed/NCBI
|