Introduction
Exosomes are bioactive vesicles derived from the
endosomal membrane system of the cell, followed by secretion into
the surrounding body fluids, with diameters ranging from 40–100 nm
and a density from 1.13–1.19 g/ml. Exosomes were officially named
by Johnstone et al in 1987, who found that microvesicles can
assist maturing erythrocytes to eliminate the transferring
receptors as well as other dumped proteins (1). However, numerous studies have
identified that exosomes can be produced by various types of cells,
including different types of epithelial and nerve cells (2), and have even been detected in various
body fluids (3–5). Exosomes are saucer-like vesicles under
electron microscopy, which contain special proteins, lipids, RNA
and micro-RNAs (6,7). They can be identified by Alix, Tsg101,
heat shock protein 70 (Hsp70), and cluster of differentiation 63
(CD63), CD81 and CD9, and they play different roles in various
pathological conditions. Thus far, in addition to participating in
metabolizing products, it has been demonstrated that exosomes
possess a number of biological functions, including immune
regulation, communication of cells, matrix remodeling, signaling
pathways activating through transferring growth factor or receptor,
intercellular exchanging of oncoprotein and oncogene, and induction
of angiogenesis and regulation of the treatment reaction. In
particular, their significant role in intercellular communication
has gained more and more attention in studies (1,8–11).
Currently, there are three main methods for extraction of exosomes:
i) Classic ultracentrifugation (12); ii) density gradient centrifugation,
which is widely used at present (13); and iii) the technology of
immunomagnetic capture (14–16).
Due to the high cost and lack of specific antigens, the certain
advantages of magnetic bead technology have resulted in its limited
application. At present, the other two methods have a lack of a
systematic comparison. The aim of the present study was to
demonstrate that the Tca8113 human tongue squamous cell carcinoma
cell line can secrete exosomes, and to perform an extensive
evaluation of the methods widely used for extracting the exosomes,
including density gradient centrifugation and
ultracentrifugation.
Materials and methods
Cell culture and collection of required
culture medium
The human tongue squamous cell carcinoma cell line,
Tca8113, was obtained from the Cell Institute, Chinese Academy of
Sciences (Shanghai, China) and cultured in Dulbecco’s modified
Eagle’s medium (DMEM; Hyclone Laboratories, Inc., Logan, UT, USA)
containing 10% fetal bovine serum (FBS; Gibco-BRL, Carlsbad, CA,
USA) with 5% CO2 at 37°C, and cells were passaged when
they were 90–100% confluent. Tca8113 cells were washed twice with
10 ml phosphate-buffered saline (PBS) and cultured for 48 h in 5 ml
DMEM media with 10% FBS, which was previously centrifuged at
100,000 × g for 70 min to eliminate bovine-derived exosomes.
Subsequently, ~50 ml culture medium (CM) was collected and stored
at −20°C for later use.
Ultracentrifugation exosome (UC-Exo)
isolation
Exosomes were isolated from the required cell
culture supernatant as described previously (12). Briefly, the cell culture supernatant
was centrifuged (300 × g for 10 min; 2,000 × g for 20 min to
eliminate dead cells; and 10,000 × g for 30 min to remove debris)
and then pelleted by ultracentrifugation at 100,000 × g for 70 min
at 4°C. The pellet was resuspended in 1 ml PBS and re-centrifuged
(100,000 × g, 70 min), as aforementioned. The products (UC-Exo)
were resuspended in 200 μl PBS and passed through 0.22-μm
microcentrifuge filters (Sigma-Aldrich, St. Louis, MO, USA) prior
to being stored at −80°C.
Density gradient exosome (DG-Exo)
isolation
Exosomes were isolated using a method described by
Lamparski et al (17).
Briefly, the collected CM was centrifuged at 2000 × g for 30 min to
remove cellular debris. The processed supernatant was concentrated
by centrifugation for 50 min at 1,000 × g in a 100 kDa molecular
weight cut-off hollow-fibre membrane (Millipore, Bedford, MA, USA).
The concentrated products were collected and added to an
ultracentrifuge tube with a 30% sucrose/D2O cushion
(density, 1.210 g/cm3) at the bottom of the tube,
followed by ultracentrifugation (Sorvall Ultra Pro 80; Kendro
Laboratory Products Ltd., Newtown, CT, USA) in a Surespin 630
swinging bucket (Thermo Fisher Scientific, Rockford, IL, USA) at
100,000 × g at 4°C for 70 min. Subsequently, the cushion was
collected, followed by being washed and concentrated twice with PBS
by centrifuging for 50 min at 1,000 × g in the aforementioned
capsule. The following steps were consistent with the former
method.
Analysis of the laser particle size
The analysis of the particle size was carried out
and repeated three times with a laser diffraction instrument
(Malvern Zetasizer Nano ZS90; Malvern Instruments Ltd., Malvern,
UK). A 30-μl aliquot of exosomes isolated respectively from the two
types of methods was diluted with PBS to 1 ml and transferred to
the specific tube subsequent to repeatedly blowing. The particle
size was measured every 0.5 min for the duration of the test and
the results were automatically recorded.
Transmission electron microscopy
(TEM)
The process of observation through TEM were
performed as previously described (16) with slight alterations. Briefly, a 20
μl aliquot of exosome preparations were placed onto formvar-coated
200-mesh copper grids (ProSciTech, Queensland, Australia) for 1 min
at room temperature and allowed to dry through filter paper. The
grids were subsequently washed twice with water for 5 min and
stained with 20 g/l uranyl acetate in water (ProSciTech) for 1 min.
The grids were examined at an acceleration voltage of 100 kV using
a JEOL JEM-2100 TEM (JEOL USA, Inc., Peabody, MA, USA).
Protein extraction and concentration
determination
As a result of the pre-experiment, the highest
concentration of protein appeared in the certain instances when 100
μl lysis buffer was added to the same volume of samples. According
to the total protein extraction kit instructions (Bi Yuntian
Biological Technology Institution, Shanghai, China), 100 μl lysis
buffer was added to the same volume of samples on ice, followed by
shaking wildly on the horizontal shaking platform for 15 min and
centrifuging at 15,558 × g at 4°C for another 15 min. Subsequently,
the supernatant was obtained. Finally, a 20-μl aliquot of the
supernatants was used for the determination of the protein
concentration with the bicinchoninic acid (BCA) Protein Assay kit
(Bi Yuntian Biological Technology Institution).
SDS-page gel electrophoresis with
Coomassie brilliant blue staining and western blotting
Total cellular proteins were loaded and run on 10%
SDS gels (Beyotime Institute of Biotechnology, Shanghai, China) and
then either transferred onto polyvinylidene fluoride membranes
(Bio-Rad, Hercules, CA, USA) or stained with Coomassie brilliant
blue (Beyotime Institute of Biotechnology). The membranes were
blocked in 5% (w/v) skimmed milk in Tris-buffered saline with Tween
20 and incubated at 4°C with primary antibodies against monoclonal
mouse anti-human Alix (1:1,000; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) or Hsp70 (1:1,000; Santa Cruz Biotechnology,
Inc.) overnight. The polyclonal rabbit anti-goat IgG-horseradish
peroxidase secondary antibodies (Wuhan Boster Biological
Technology, Ltd., Wuhan, China) were incubated at room temperature
for 1 h and the membranes were visualized by the Amersham ECL
Select detection system (Amersham Pharmacia Biotech, Little
Chalfont, UK).
Statistical analysis
All data were performed using SPSS 13.0 (SPSS, Inc.,
Chicago, IL, USA) statistical software. A paired t-test was used to
compare the protein concentration between the two methods.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Particle size distribution of the
exosomes
The particle size of the exosomes isolated
respectively from the two types of method were measured by a laser
diffraction instrument. The results revealed that the range of the
particle size of UC-Exo (Fig. 1A)
was wider than that of DG-Exo (Fig.
1B), even >200 nm, whilst the diameters of DG-Exo were
mostly in a uniform range from 30–120 nm. The values of the
polydispersity index were 0.387 and 0.481 for UC-Exo and DG-Exo,
respectively, and were within the normal range.
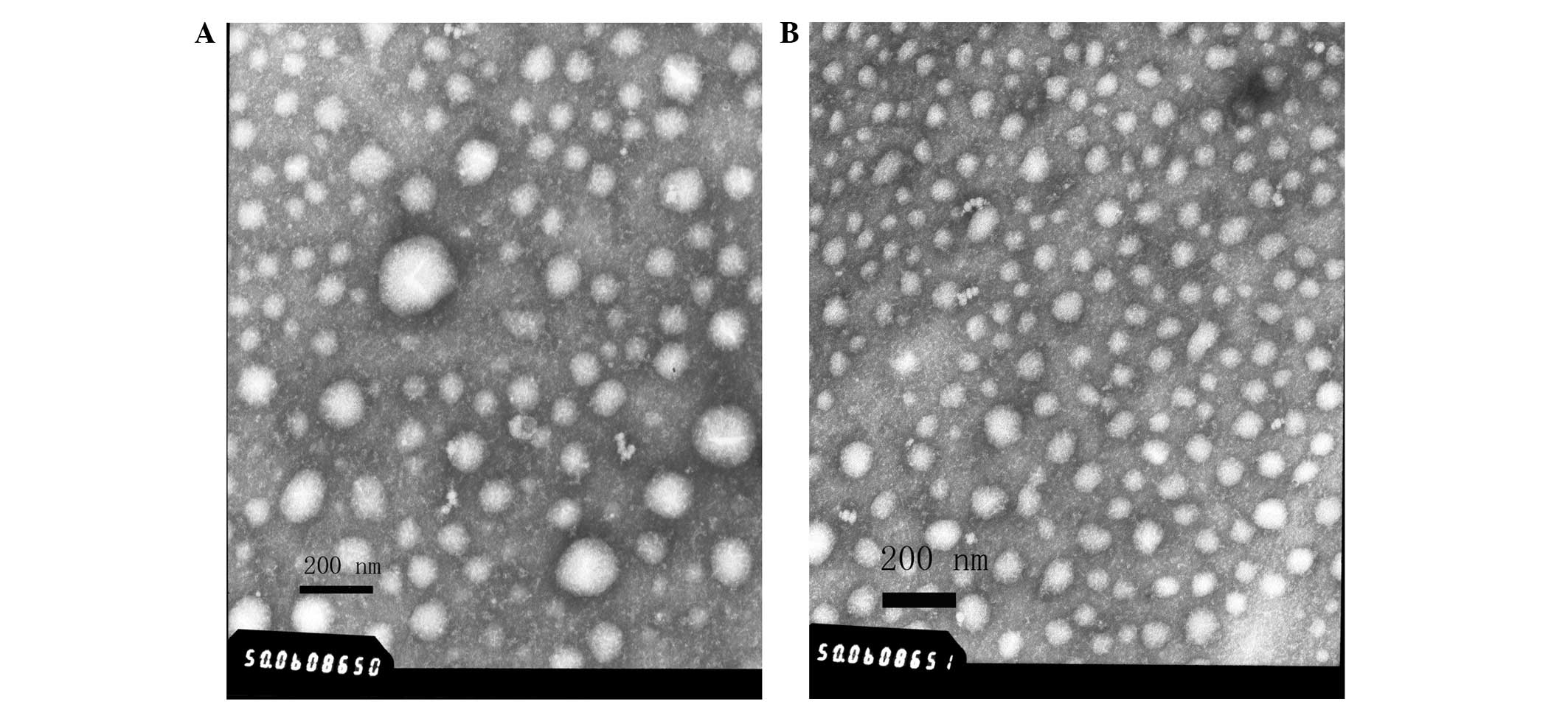
Morphology observation of the
exosomes
Morphological analysis of the UC-Exo and DG-Exo
samples using TEM revealed the same results that the vesicles were
comprising round-shaped 30–150-nm diameter vesicles, which is
consistent with the aforementioned exosomes (Fig. 2), while the differences with the
traditional cup-shape may be associated with the preparation of the
samples. However, certain UC-Exo (Fig.
2A) samples contained vesicles that were >200 nm in
diameter, which could even be suspected to be a class of apoptosis
body, while the DG-Exo were well-distributed (Fig. 2B).
Determination of the protein
concentration of the exosomes
The determination of the total protein concentration
of the UC-Exo and DG-Exo samples were compared using the BCA
method. The measurement included 12 separate samples from UC-Exo
and DG-Exo (Table I) and the
correlation coefficient was 0.99. Fig.
3 shows the curve comparison chart generated by Table I, with a paired t-test, P<0.05,
which indicated that the difference had a statistical
significance.
 | Table IDetermination of protein concentration
of exosomes purified by two different methods. |
Table I
Determination of protein concentration
of exosomes purified by two different methods.
| Sample | UC-Exo concentration,
μg/ml | DG-Exo concentration,
μg/ml |
|---|
| 1 | 174.957 | 380.910 |
| 2 | 132.555 | 467.076 |
| 3 | 75.361 | 364.452 |
| 4 | 179.888 | 288.936 |
| 5 | 64.325 | 410.922 |
| 6 | 103.051 | 471.916 |
| 7 | 135.000 | 535.814 |
| 8 | 165.013 | 410.922 |
| 9 | 163.076 | 411.892 |
| 10 | 132.096 | 427.382 |
| 11 | 133.064 | 329.598 |
| 12 | 252.146 | 327.622 |
Protein composition of the exosomes
The proteins were separated by SDS-PAGE gel
electrophoresis and stained by Coomassie brilliant blue. The
results showed that the bands of the two samples were almost the
same and were mainly distributed between 40–130 kDa with obvious
separated strips (Fig. 4).
Identification of the molecular
markers
The molecular phenotype of the exosomes was typical
of exosomes from other sources. In the present study, it was found
that Hsp70 and Alix, the protein markers of exosomes, were detected
similarly in the purified exosomes with various methods (Fig. 5).
Discussion
Exosomes may participate in cell communication by
delivering proteins, RNA and miRNA (18–20)
and has the ability to induce or suppress the immune system
(21). Exosomes are capable of
promoting angiogenesis, remodelling the microenvironment and
promoting tumor growth (22,23).
Their use in diagnosis and treatment have been confirmed (3,4,24).
However, the current area of study offering the most promise lies
with isolating and extracting high quality exosomes.
The significant reference standard to judge exosomes
purification methods is based on the ability to remove other
membranous particles and concentrate protein. Although
ultracentrifugation is the most widely used method for exosomes
isolation, it is limiting in these respects, so a more specific
method is required. In the present study, two strategies were
compared for purifying the human tongue cancer cell line
Tca8113-derived exosomes; ultracentrifugation and density gradient
separation. The efficacy of the two strategies was judged by TEM,
particle size distribution and cursory proteome profiling of the
enrichment of typical exosomal markers, including Alix and Hsp70,
which was confirmed by western blot analysis and protein
concentration.
Experimental results show that the exosomes
extracted from the two methods, comprising round-shaped 30–150-nm
diameter vesicles, are consistent with exosomes reported previously
(16,25). However, the coexistence of the
larger vesicles with UC-Exo is unknown and presumably not due to
the vesicles being slightly clumped together or due to other mixed
impurities. Another study has noted that steps, including freezing
and thawing, or multiple centrifugal steps do not affect exosome
size and shape (17). Western blot
analysis revealed the presence of the exosome markers Alix and
Hsp70 in both methods, which showed that both of the methods can
extract the exosomes. In addition, the results of laser particle
size measurement revealed that UC-Exo had a wider range of diameter
distribution and a larger proportion of vesicles with diameters
>200 nm, while the DG-Exo was more uniform and the majority of
diameters were distributed between 30–150 nm with an average of
87.3 nm. This shows that the latter samples have a higher purity.
Through SDS-PAGE gel electrophoresis and Coomassie brilliant blue
staining, the bands of the two samples were found to be almost the
same and mainly distributed between 40–130 kDa with obvious
separated strips, while the 170 kDa was not obvious. However, by
comparing the two methods with protein concentration and western
blot semi-quantitative analysis, it was found that the total
protein concentration of DG-Exo was higher and had a higher
proportion of target protein, which indicated that density gradient
centrifugation has a higher extraction efficiency. Additionally,
the density gradient centrifugation used a 30% sucrose/heavy water
cushion, which has a density distribution range that is consistent
with exosomes and is a type of variable purification, and has been
used widely to purify exosomes (26). The repeated purification with
ultrafiltration also produces samples with a higher purity and
improved quality. Therefore, density gradient centrifugation is
more comprehensive and more efficient than ultracentrifugation in
the extraction of exosomes. However, there are specific problems
attached, including the higher cost, the higher requirements of
equipment and technical ability, the time required is longer and
the separation and extraction of heavy water at a high-quality is
difficult. In addition, future studies could be conducted to
identify whether the cell damage from heavy water can be simply
eliminated by repeated dilution of the purification and whether
this will effect the subsequent experiment, whether the material
existing in the extraction of the sample whose diameter is >200
nm can be further purified or excluded and whether one or more
specific markers and extraction methods for exosomes will be a
problem that requires solving. In conclusion, both preparations
contained vesicles with sizes of 30–150 nm and expression of the
exosome markers, Alix and HSP70. However, density gradient
separation was considered to be the efficient method to isolate
exosomes, as it was able to enrich exosome markers, and
exosome-associated proteins by at least two-fold more than the
other methods studied. Protein, lipid, mRNA and microRNA analyses
of highly-purified vesicles will lead to significant advances in
exosome characterization, and facilitate a deeper understanding of
their biological functions.
Acknowledgements
This study was supported by the National Natural
Science Foundation (grant no. 81172578).
References
|
1
|
Johnstone RM, Adam M, Hammond JR, et al:
Vesicle formation during reticulocyte maturation. Association of
plasma membrane activities with released vesicles (exosomes). J
Biol Chem. 262:9412–9420. 1987.
|
|
2
|
Cocucci E, Racchetti G and Meldolesi J:
Shedding microvesicles: artefacts no more. Trends Cell Biol.
19:43–51. 2009.
|
|
3
|
Gonzales PA, Pisitkun T, Hoffert JD,
Tchapyjnikov D, Star RA, Kleta R, et al: Large-scale proteomics and
phosphoproteomics of urinary exosomes. J Am Soc Nephrol.
20:363–379. 2009.
|
|
4
|
Nilsson J, Skog J, Nordstrand A, Baranov
V, Mincheva-Nilsson L, Breakefield XO and Widmark A: Prostate
cancer-derived urine exosomes: a novel approach to biomarkers for
prostate cancer. Br J Cancer. 100:1603–1607. 2009.
|
|
5
|
Michael A, Bajracharya SD, Yuen PS, Zhou
H, Star RA, Illei GG and Alevizos I: Exosomes from human saliva as
a source of microRNA biomarkers. Oral Dis. 16:34–38. 2010.
|
|
6
|
Skog J, Würdinger T, van Rijn S, et al:
Glioblastoma microvesicles transport RNA and proteins that promote
tumour growth and provide diagnostic biomarkers. Nat Cell Biol.
10:1470–1476. 2008.
|
|
7
|
Hong BS, Cho JH, Kim H, et al: Colorectal
cancer cell-derived microvesicles are enriched in cell
cycle-related mRNAs that promote proliferation of endothelial
cells. BMC Genomics. 10:5562009.
|
|
8
|
Zhang HG and Grizzle WE: Exosomes and
cancer: a newly described pathway of immune suppression. Clin
Cancer Res. 17:959–964. 2011.
|
|
9
|
Keller S, Sanderson MP, Stoeck A and
Altevogt P: Exosomes: from biogenesis and secretion to biological
function. Immunol Lett. 107:102–108. 2006.
|
|
10
|
Simons M and Raposo G: Exosomes -
vesicular carriers for intercellular communication. Curr Opp Cell
Biol. 21:575–581. 2009.
|
|
11
|
Mathivanan S, Ji H and Simpson RJ:
Exosomes: extracellular organelles important in intercellular
communication. J Proteomics. 73:1907–1920. 2010.
|
|
12
|
Raposo G, Nijman HW, Stoorvogel W,
Liejendekker R, Harding CV, Melief CJ and Geuze HJ: B lymphocytes
secrete antigen-presenting vesicles. J Exp Med. 183:1161–1172.
1996.
|
|
13
|
Ostrowski M, Carmo NB, Krumeich S, et al:
Rab27a and Rab27b control different steps of the exosome secretion
pathway. Nat Cell Biol. 12:19–30. 2010.
|
|
14
|
Rabinowits G, Gerçel-Taylor C, Day JM,
Taylor DD and Kloecker GH: Exosomal microRNA: a diagnostic marker
for lung cancer. Clin Lung Cancer. 10:42–46. 2009.
|
|
15
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008.
|
|
16
|
Mathivanan S, Lim JW, Tauro BJ, Ji H,
Moritz RL and Simpson RJ: Proteomics analysis of A33
immunoaffinity-purified exosomes released from the human colon
tumor cell line LIM1215 reveals a tissue-specific protein
signature. Mol Cell Proteomics. 9:197–208. 2010.
|
|
17
|
Lamparski HG, Metha-Damani A, Yao JY,
Patel S, Hsu DH, Ruegg C and Le Pecq JB: Production and
characterization of clinical grade exosomes derived from dendritic
cells. J Immunol Methods. 270:211–226. 2002.
|
|
18
|
Chairoungdua A, Smith DL, Pochard P, Hull
M and Caplan MJ: Exosome release of beta-catenin: a novel mechanism
that antagonizes Wnt signalling. J Cell Biol. 190:1079–1091.
2010.
|
|
19
|
Verweij FJ, van Eijndhoven MA, Hopmans ES,
et al: LMP1 association with CD63 in endosomes and secretion via
exosomes limits constitutive NF-kappaB activation. EMBO J.
30:2115–2129. 2011.
|
|
20
|
Ohshima K, Inoue K, Fujiwara A, et al:
Let-7 microRNA family is selectively secreted into the
extracellular environment via exosomes in a metastatic gastric
cancer cell line. PloS One. 5:e132472010.
|
|
21
|
Clayton A, Al-Taei S, Webber J, Mason MD
and Tabi Z: Cancer exosomes express CD39 and CD73, which suppress T
cells through adenosine production. J Immunol. 187:676–683.
2011.
|
|
22
|
Xiang X, Poliakov A, Liu C, et al:
Induction of myeloid-derived suppressor cells by tumor exosomes.
Int J Cancer. 124:2621–2633. 2009.
|
|
23
|
Castellana D, Zobairi F, Martinez MC, et
al: Membrane microvesicles as actors in the establishment of a
favorable prostatic tumoral niche: a role for activated fibroblasts
and CX3CL1-CX3CR1 axis. Cancer Res. 69:785–793. 2009.
|
|
24
|
Xiao WH, Sanren GW, Zhu JH, et al: Effect
of 5-aza-2′-deoxycytidine on immune-associated proteins in exosomes
from hepatoma. World J Gastroenterol. 16:2371–2377. 2010.
|
|
25
|
Regev-Rudzki N, Wilson DW, Carvalho TG, et
al: Cell-cell communication between malaria-infected red blood
cells via exosome-like vesicles. Cell. 153:1120–1133. 2013.
|
|
26
|
Lamparski HG, Metha-Damani A, Yao JY,
Patel S, Hsu DU, Ruegg C and LePecq JB: Production and
characterization of clinical grade exosomes derived from dendritic
cells. J Immunol Methods. 270:211–226. 2002.
|