Introduction
Prostate cancer (PC) is a type of malignant cancer
that affects male prostate tissue, and is a result of abnormal and
disordered acinar cell growth (1).
Currently, PC is the second most frequently observed cancer
worldwide and its morbidity rate is increasing annually (2). According to the World Health
Organization (WHO), there were 899,000 new PC cases in 2008, which
accounted for ~14% of new cancer cases in men worldwide (3). According to a prediction by the WHO, the
number of patients with PC will have increased to 1.7 million by
2030 (4). Epidemiological studies
have reported that the morbidity rate of PC is dependent on
geography and ethnicity, and is more prevalent in developed
countries (5,6).
Cofilin-1 is an actin-associated protein with a low
molecular weight that is present in eukaryotic cells (7). The genes encoding cofilin-1 are located
on chromosome 11q13 and are expressed in various types of
non-muscle tissue, including the liver and brain (8). The remodeling of the actin cytoskeleton
serves an important role in the processes of invasion and
metastasis of cancer cells (9). The
regulation of actin and actin-associated proteins,
includingcofilin-1, is important in the formation of pseudopodium
(8). Cofilin-1 is an essential
regulatory factor for cancer cell metastasis and invasion, and its
overexpression leads to increased cancer cell migration (10). Therefore, cofilin-1 inhibition may
significantly reduce the invasiveness of cancer cells (11). In melanoma cells, exogenous cofilin-1
signaling may upregulate the expression of matrix
metalloproteinases 2 and 14 (11).
Cofilin-1 is therefore able to promote matrix degradation,
facilitating pseudopodium-mediated invasion.
Mammalian target of rapamycin (mTOR) is an important
signal transduction molecule, which regulates transcription and
translation through the integration of extracellular signals
(12). mTOR complex 1 (mTORC1) is the
primary form of mTOR and has an important role in cell growth,
proliferation, differentiation and apoptosis (12–14).
mTORC1 consists of mTOR, TORC subunit LST8 and
regulatory-associated protein of mTOR (RPTOR) (14). A previous study demonstrated that the
dysregulation of elements of the mTOR signaling pathway was
associated with the occurrence, development and metastasis of
various types of cancer (15).
Cucurbitacin E is a member of the cucurbitacin
family, which are a group of tetracyclic triterpenoids extracted
from cucurbitaceous plants (16).
Studies have demonstrated that cucurbitacin E possesses a number of
pharmacological functions, including anticancer, anti-viral and
anti-inflammatory effects (17,18).
Cucurbitacin E is able to inhibit the activation of signal
transducer and activator of transcription 3 (STAT3)-associated
transcription factors, and disrupt the mitogen-activated protein
kinase signaling pathway (19).
Cucurbitacin E induces G2/M cell cycle arrest and apoptosis in
cancer cells, and subsequent cytoskeletal changes (17). Therefore, the present study aimed to
investigate whether cucurbitacin E induces the apoptosis of human
PC through cofilin-1 and mTORC1 signaling.
Materials and methods
Materials
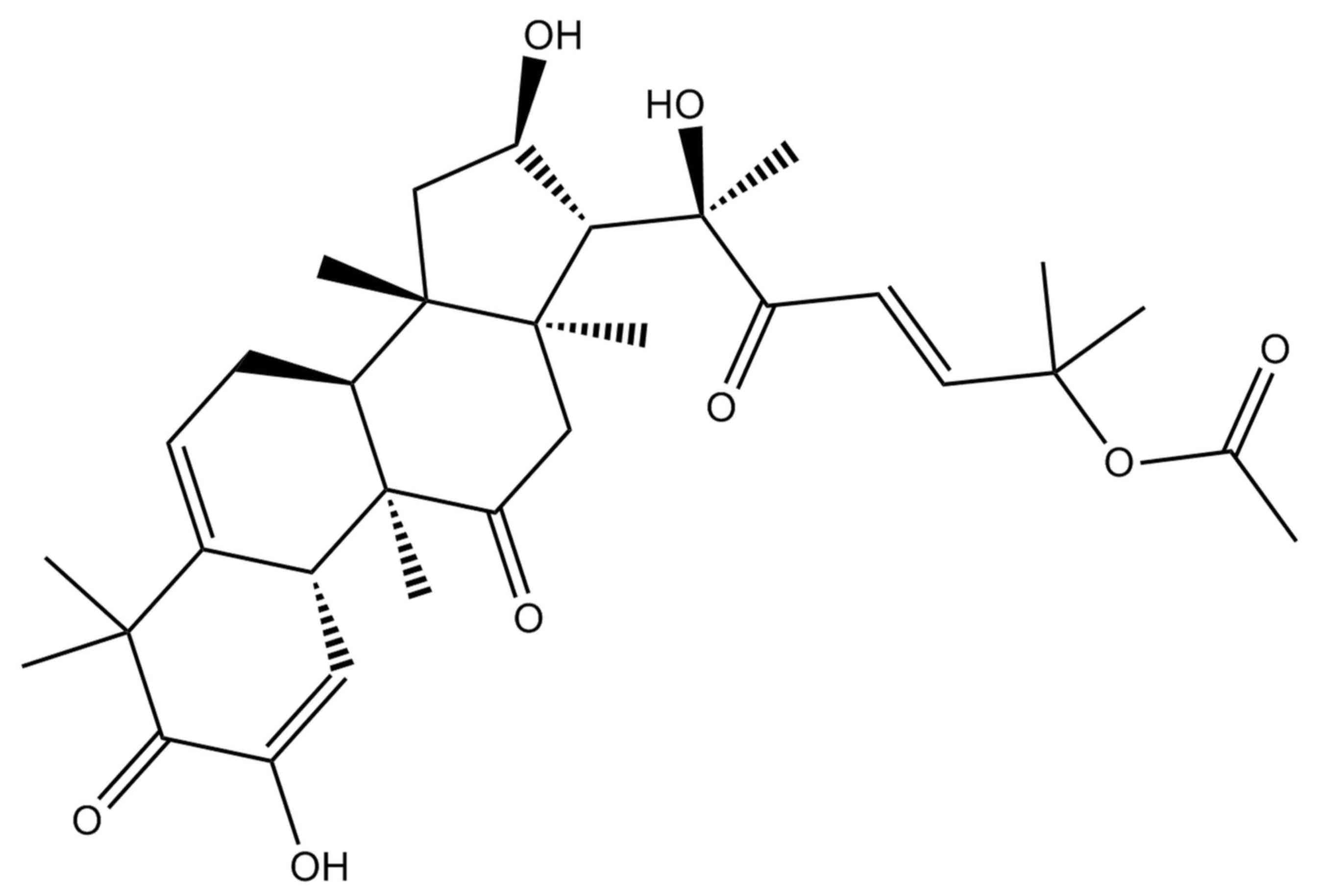
Cucurbitacin E was obtained from Sigma-Aldrich
(Merck Millipore, Darmstadt, Germany) and its formula is shown in
Fig. 1. Dimethyl sulfoxide (DMSO),
SDS, MTT and lactate dehydrogenase (LDH) were also obtained from
Sigma-Aldrich (Merck Millipore). Dulbecco's modified Eagle's medium
(DMEM), fetal bovine serum (FBS), penicillin and streptomycin were
obtained from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
The Cell Death Detection ELISA kit was obtained from Roche
Diagnostics (Indianapolis, IN, USA). Polyvinylidene difluoride
(PVDF) membranes were obtained from GE Healthcare Life Sciences
(Chalfont, UK).
Cell culture
The human PC cell line, LNCaP, was obtained from the
Experiment Center of Yan'an University (Yan'an, China), and
cultured in DMEM supplemented with 10% FBS, 100 U/ml penicillin and
100 µg/ml streptomycin. Cells were incubated at 37°C in a
humidified atmosphere containing 5% CO2.
Cell viability assay
Cell viability was determined using the MTT assay,
as previously described (15).
Briefly, LNCaP cells were seeded in 96-well plates at a density of
3,300 cells/well and incubated overnight at 37°C, prior to
treatment with 0, 0.01, 0.1, 1, 10 and 100 µM cucurbitacin E. At 24
and 48 h following treatment with cucurbitacin E, 20 µl MTT
solution (0.1 mg/ml) was added to the cells and the plates were
subsequently incubated at 37°C for 4 h. Following incubation, 150
µl DMSO was added for 20 min to dissolve the formazan crystals. The
absorbance at 570 nm was measured using a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA) and the cell viability was
subsequently calculated using the following formula:
(A570sample - A570blank)/(A570control -
A570blank) × 100.
Cytotoxicity assay
Cytotoxicity was determined using the LDH assay, as
previously described (15). Briefly,
LNCaP cells were seeded in 96-well plates at a density of 3,300
cells/well and incubated overnight at 37°C prior to treatment with
0, 0.1, 1 and 10 µM cucurbitacin E. After 24 h, 100 µl LDH solution
was added to the cells and the plates were incubated at 37°C for 30
min. The absorbance at 490 nm was measured using a microplate
reader.
Detection of apoptosis
LNCaP cells were seeded into 6-well plates at a
density of 1×105 cells/well, in triplicate, prior to
treatment with 0, 0.1, 1 and 10 µM cucurbitacin E for 24 h.
Apoptotic nucleosomes were measured using the Cell Death Detection
kit (KeyGen, Shanghai, China) with 10 µg Annexin V-fluorescein
isothiocyanate and 5 µg propidium iodide for 15 min in darkness
according to the manufacturer's protocol. Apoptosis rate was
analyzed by flow cytometry (Epics XL Flow Cytometer; Beckman
Coulter, Inc., Brea, CA, USA). The data were analyzed using the
ModFit LT program (Verity Software House, Topsham, ME, USA).
Western blot analysis
LNCaP cells were seeded into 6-well plates at a
density of 1×105 cells/well, in triplicate, prior to
treatment with 0, 0.1, 1 and 10 µM cucurbitacin E for 24 h. Cells
were lysed on ice in radioimmunoprecipitation assay lysis buffer
(Beyotime Institute of Biotechnology, Haimen, China) and
homogenized. Total protein concentration was subsequently assessed
using a Bradford Assay kit (Beyotime Institute of Biotechnology).
Equal amounts of protein (50 µg) were resolved on a 12% w/v gel
using SDS-PAGE and transferred onto a PVDF membrane. The membrane
was blocked using 5% milk in TBS containing Tween-20 at 37°C for 1
h and incubated with the following primary antibodies overnight at
4°C: Mouse anti-cofilin-1 (1:500, sc-53934); mouse phosphorylated
(p)-anti-mTOR (1:500, sc-293132); anti-AMP-activated protein kinase
(AMPK) (1:400, sc-25792); mouseanti-P53 (1:2,000; sc-6243), mouse
anti-caspase-9 (1:400, sc-47698; all Santa Cruz Biotechnology,
Inc., Dallas, TX, USA) and mouse anti-β-actin (AF0003, 1:500;
Beyotime Institute of Biotechnology). The membrane was then
incubated with horseradish peroxidase-conjugated goat anti-rabbit
or anti-mouse secondary antibodies (40,000, A0208 or A02581;
Beyotime Institute of Biotechnology) for 2 h at room temperature.
β-actin was used as the loading control. Protein bands were
visualized using New-SUPER ECL (cat. no. KGP1127; Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China) and the band intensity was
quantified using ImageJ software (version 3.0; National Institutes
of Health, Bethesda, MA, USA).
Statistical analysis
Values are presented as the mean ± standard error of
the mean. Statistical analysis was performed using GraphPad Prism
4.0 (GraphPad Software Inc., La Jolla, CA, USA), and the results
were analyzed using analysis of variance and the Bonferroni post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Treatment with cucurbitacin E
suppresses LNCaP cell viability
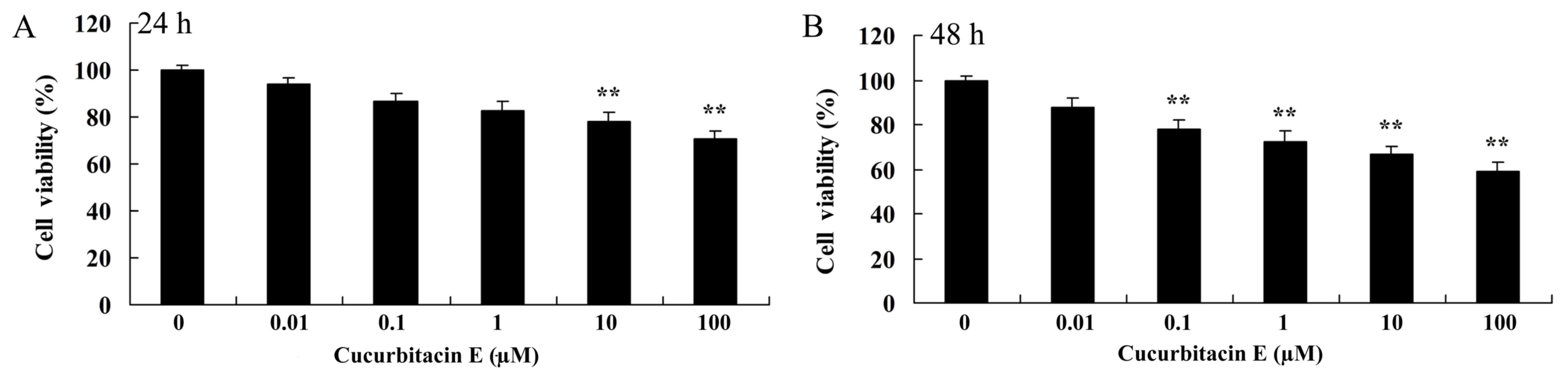
The MTT assay was performed to demonstrate that
treatment with cucurbitacin E reduces the viability of human PC
cells. Treatment with cucurbitacin E was shown to suppress LNCa
cell viability in a dose- and time-dependent manner (Fig. 2). Treatment with 10 and 100 µM
cucurbitacin E for 24 h significantly decreased LNCaP cell
viability compared with the untreated cells (P=0.0092 and 0.0081,
respectively; Fig. 2A). Treatment
with 0.1–100 µM cucurbitacin E for 48 h significantly decreased
LNCaP cell viability compared with the untreated cells (P=0.0079,
0.0063, 0.0055 and 0.0041 for 0.1, 1, 10 and 100 µM, respectively;
Fig. 2B).
Cucurbitacin E exhibits cytotoxicity
in LNCaP cells
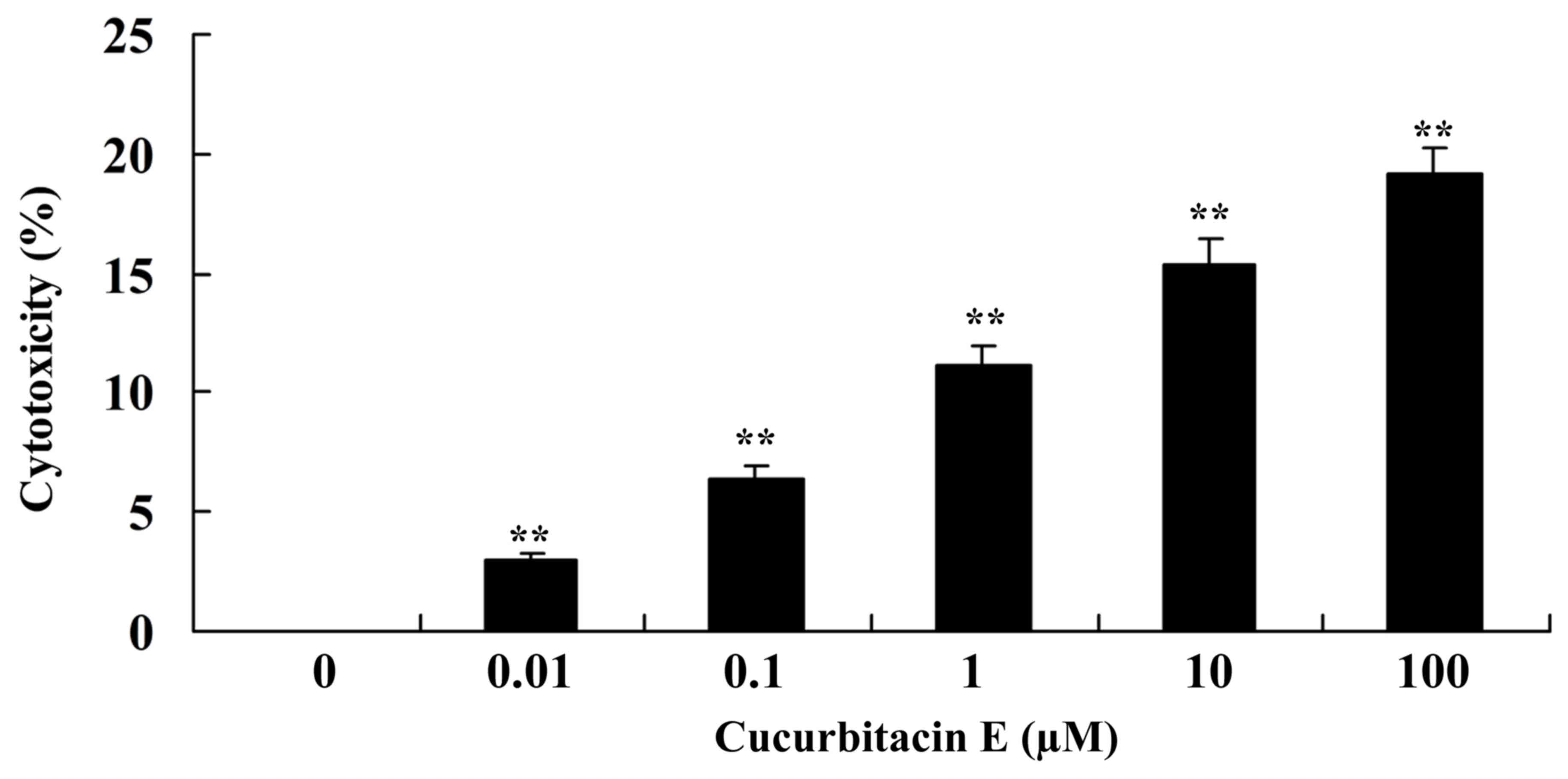
The LDH assay was performed to analyze the
cytotoxicity of cucurbitacin E on human PC cells. Cucurbitacin E
significantly exhibited marked cytotoxicity on LNCaP cells in a
dose-dependent manner (P=0.00092, 0.0079, 0.0065, 0.0050 and
0.00021 for 0.01, 0.1, 1, 10 and 100 µM, respectively; Fig. 3).
Treatment with cucurbitacin E induces
apoptosis of LNCaP cells
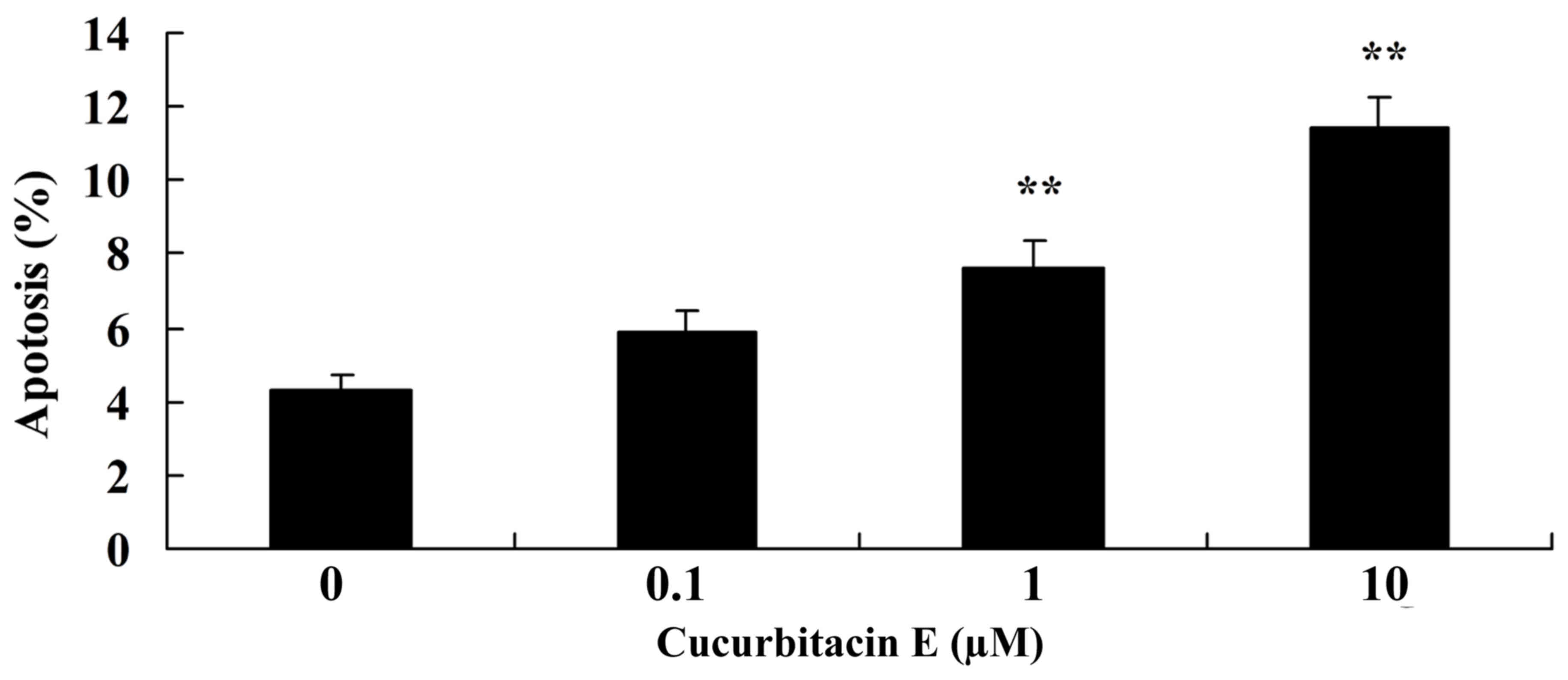
The Cell Death Detection ELISA kit was used to
measure the effect of cucurbitacin E on the apoptosis of human PC
cells. Treatment with 1 and 10 µM cucurbitacin E for 24 h led to
significantly increased apoptosis in LNCaP cells, as compared with
the untreated cells (P=0.0076 and 0.0045, respectively; Fig. 4).
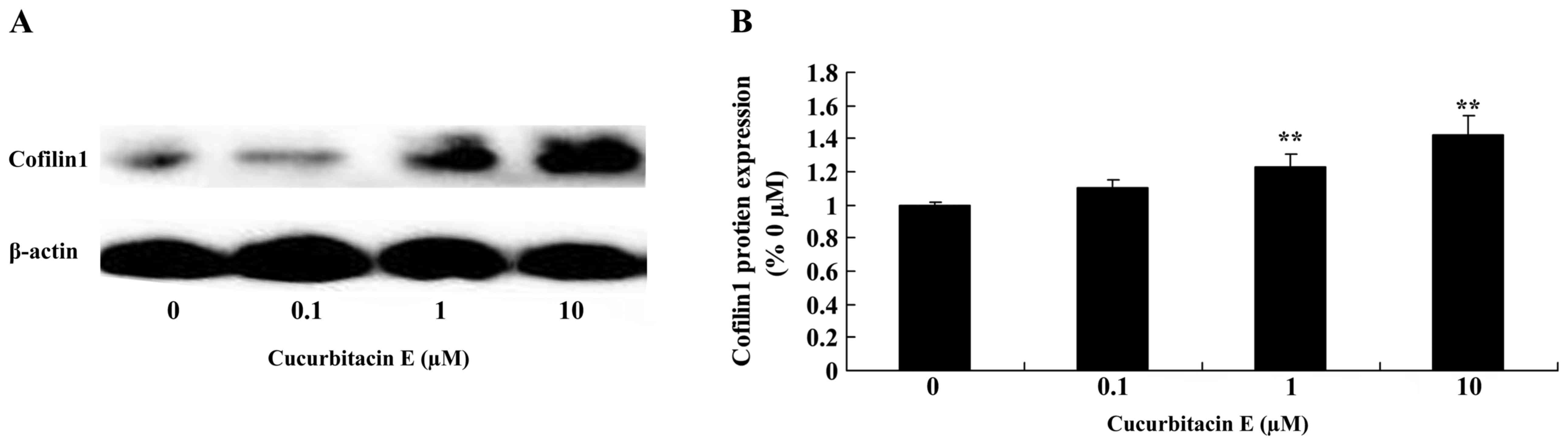
Treatment with cucurbitacin E induces
cofilin-1 protein expression
The effect of treatment with 0, 0.1, 1 and 10 µM
cucurbitacin E for 24 h on cofilin-1 protein expression in LNCaP
cells was evaluated. Cofilin-1 protein expression was significantly
increased following treatment with 1 and 10 µM cucurbitacin E
compared with the untreated cells (P=0.0076 and 0.0045,
respectively; Fig. 5A and B).
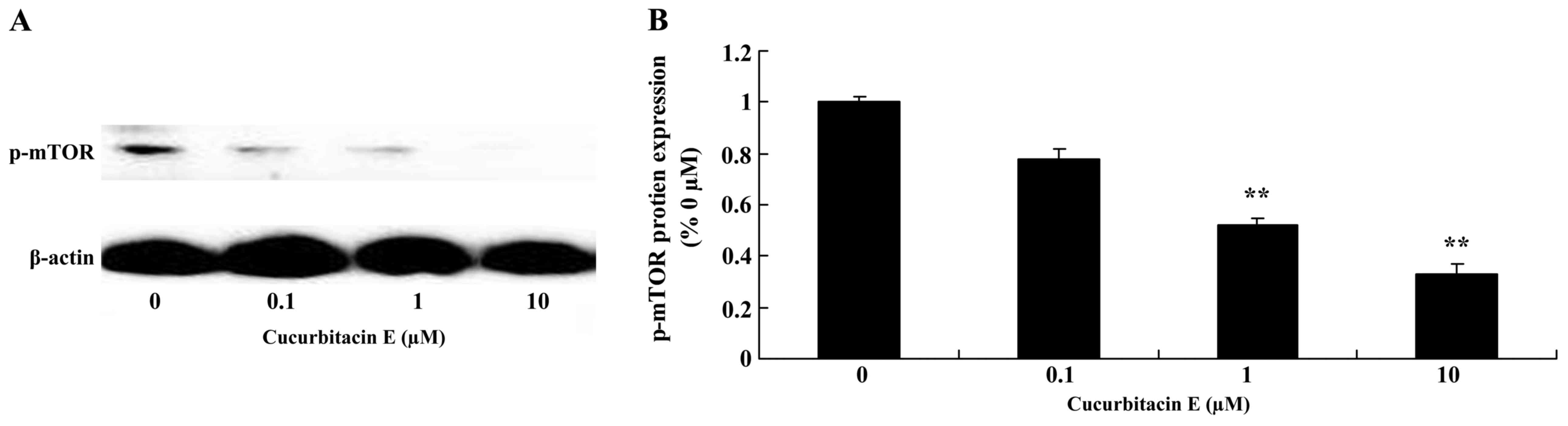
Treatment with cucurbitacin E induces
mTOR protein expression
To determine whether the induction of cofilin-1
expression impacted on the expression of other proteins, mTOR
protein expression was evaluated using western blotting. mTOR
protein expression was significantly increased following treatment
with 1 and 10 µM cucurbitacin E compared with the untreated cells
(P=0.0030 and 0.0009, respectively; Fig.
6A and B).
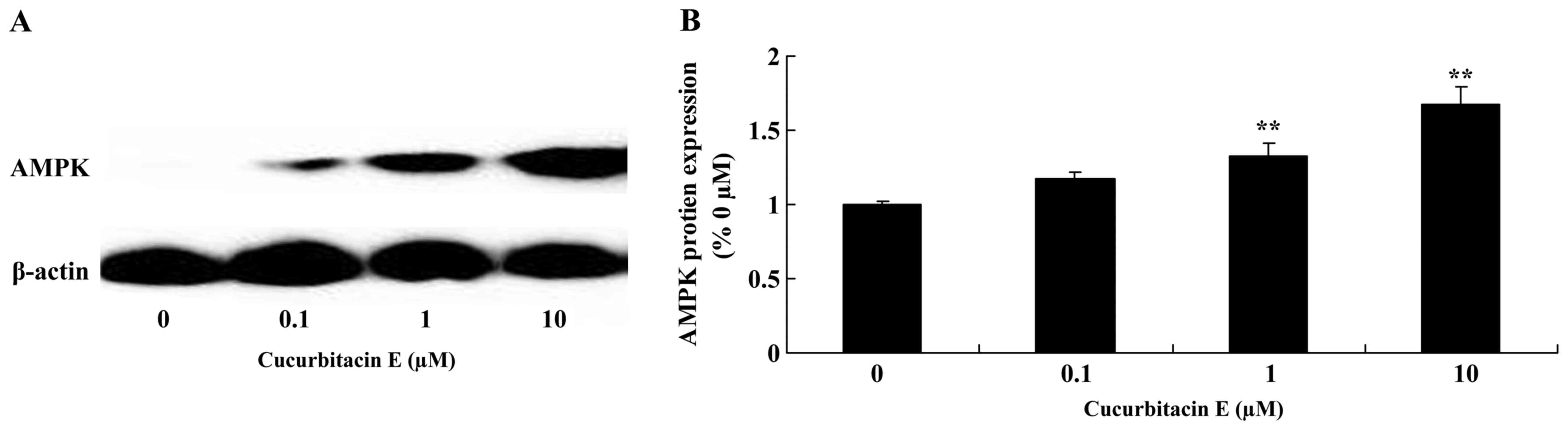
Treatment with cucurbitacin E induces
AMPK protein expression
To determine changes in the expression of apoptotic
proteins in LNCaP cells following treatment with cucurbitacin E,
AMPK protein expression was evaluated using western blotting. AMPK
protein expression was significantly increased following treatment
with 1 and 10 µM cucurbitacin E compared with the untreated cells
(P=0.0011 and P<0.0001, respectively; Fig. 7A and B).
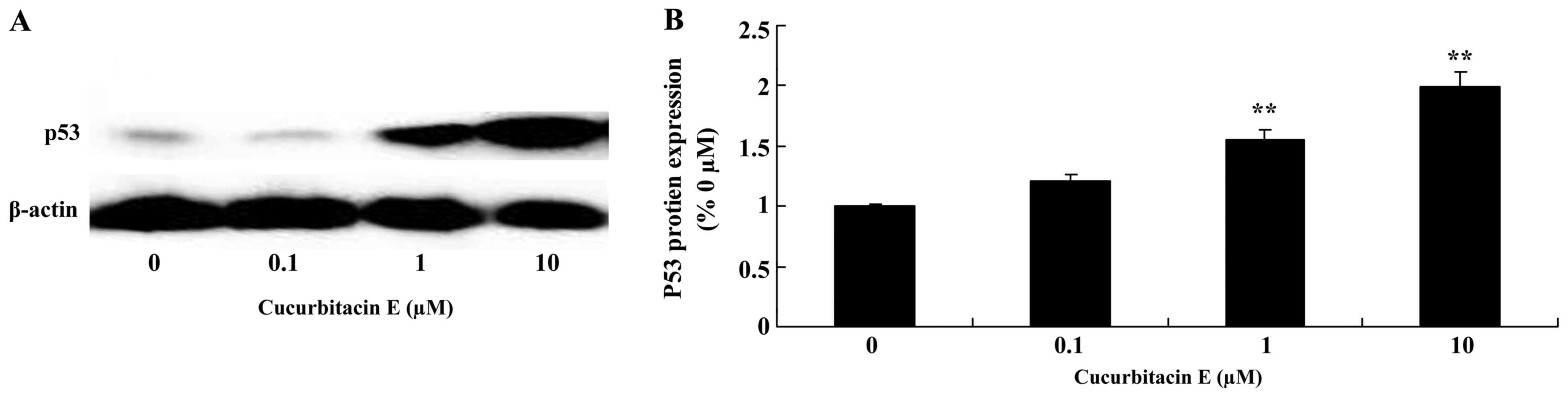
Treatment with cucurbitacin E induces
p53 protein expression
The effect of treatment with 0, 0.1, 1 and 10 µM
cucurbitacin E for 24 h on p53 protein expression in LNCaP cells
was evaluated. p53 protein expression was significantly increased
following treatment with 1 and 10 µM cucurbitacin E compared with
the untreated cells (P=0.0021 and P<0.0001, respectively;
Fig. 8A and B).
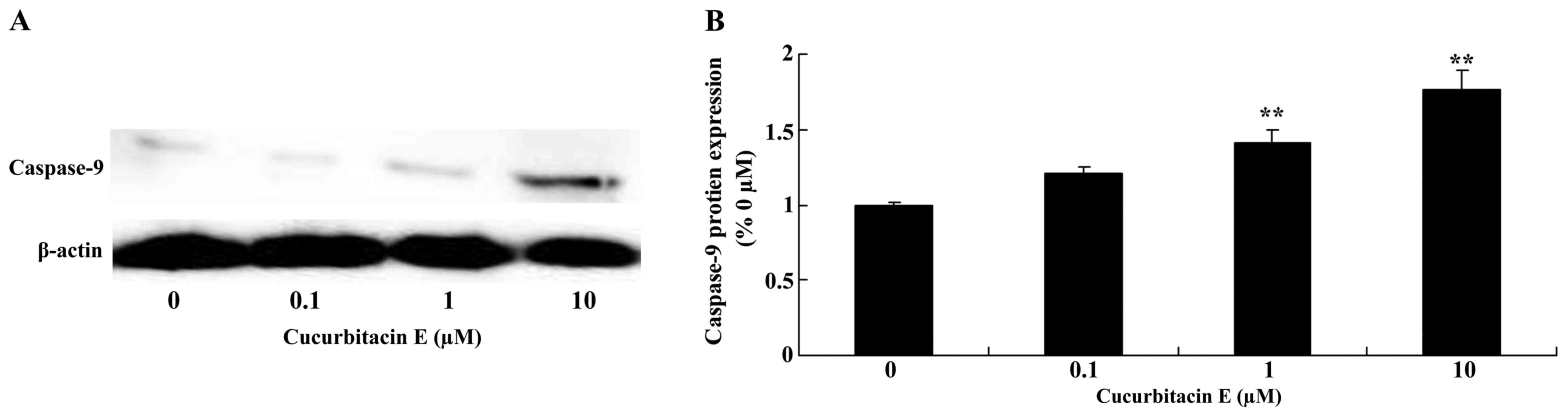
Treatment with cucurbitacin E
inducescaspase-9 protein expression
The effect of treatment with 0, 0.1, 1 and 10 µM
cucurbitacin E for 24 h on caspase-9 protein expression in LNCaP
cells was evaluated. Caspase-9 protein expression was significantly
increased following treatment with 1 and 10 µM cucurbitacin E
compared with the untreated cells (P=0.0050 and 0.0023,
respectively; Fig. 9A and B).
Discussion
PC is the most frequently occurring cancer in
Western countries and a primary cause of cancer-associated
mortality (4). Although castration is
effective in patients with early stage PC, other therapeutic
regimes are limited by the development of hormonal-refractory or
hormone-resistant PC (20). The
present study demonstrated that cucurbitacin E exerted cytotoxic
effects on LNCaP cells, decreasing the cell viability and inducing
apoptosis. Previous studies demonstrated that treatment with
cucurbitacin E induced apoptosis in triple-negative breast cancer,
ovarian cancer and PC cells (21–23).
Cofilin-1 is a low molecular weight polypeptide
expressed universally in eukaryotes (9). Cofilin-1 expression has been shown to be
dysregulated in various types of cancer tissue, including bladder,
breast and esophageal cancer, and affects the occurrence,
progression, diagnosis, prognosis and treatment of cancer (8). The results of the present study
demonstrated that treatment with cucurbitacin E induced cofilin-1
protein expression in LNCaP cells. Nakashima et al (24) suggested that cucurbitacin E inhibits
cofilin-1 phosphorylation in human leukemia cells.
mTORC1 is an important regulatory factor of cell
growth, proliferation and cell cycle progression by regulating the
phosphorylation of its downstream proteins (25). RPTOR interacts with mTOR and mediates
trophic signal transduction to regulate cell size and
mTOR-associated protein expression (26). The mTOR signaling pathway is an
important signaling pathway; its overactivation is able to promote
the growth and proliferation of cancer cells, inhibit apoptosis and
autophagy, and promote cell migration, invasion, metastasis and
angiogenesis (27). mTOR is closely
associated with the genesis and development of cancer. Previous
studies have demonstrated that mTOR protein expression is
significantly upregulated in various types of cancer tissue,
including PC, liver cancer, cervical cancer, colorectal cancer,
lung adenocarcinoma, esophageal squamous carcinoma, non-small cell
lung cancer and extra hepatic bile duct carcinoma (25,28). The
results of the present study demonstrated that treatment with
cucurbitacin E induced mTOR protein expression inhuman PC cells.
Zha et al (29) suggested that
cucurbitacin E induces autophagy through AMPK and mTORC1
signaling.
AMPK regulates the formation of the apoptosome
through activation of the p53 signaling pathway and caspase-9
precursors (26). AMPK activates a
number of proteases and endonucleases, and inhibits DNA repairase,
which leads to the damage of cytoskeletal proteins and
nucleoproteins, and subsequent apoptosis (30). The results of the present study
demonstrated that treatment with cucurbitacin E induced AMPK, p53
and caspase-9 protein expression in LNCaP cells. Sun et al
(31) reported that cucurbitacin E
treatment may be effective against pancreatic cancer by activating
p53 and STAT3 signaling. Huang et al (18) demonstrated that treatment with
cucurbitacin E induced G2/M cell cycle arrest through the
activation of caspase-8 and −9. The present study demonstrated that
treatment with cucurbitacin E induced apoptosis in human PC cells
via cofilin-1 and mTORC1 signaling. These results suggested that
cucurbitacin E may be a potential therapeutic candidate for the
treatment of PC.
References
|
1
|
Sidaway P: Prostate cancer: New biomarker
panel prognosticates patient survival. Nat Rev Clin Oncol.
12:3102015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee BH, Kibel AS, Ciezki JP, Klein EA,
Reddy CA, Yu C, Kattan MW and Stephenson AJ: Are biochemical
recurrence outcomes similar after radical prostatectomy and
radiation therapy? Analysis of prostate cancer-specific mortality
by nomogram-predicted risks of biochemical recurrence. Eur Urol.
67:204–209. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chambers SK, Newton RU, Girgis A, Nielsen
L, Lepore S, Mihalopoulos C, Gardiner R, Galvão DA and Occhipinti
S: Living with prostate cancer: Randomised controlled trial of a
multimodal supportive care intervention for men with prostate
cancer. BMC Cancer. 11:3172011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kawakami J, Cowan JE, Elkin EP, Latini DM,
DuChane J and Carroll PR: CaPSURE Investigators:
Androgen-deprivation therapy as primary treatment for localized
prostate cancer: Data from cancer of the prostate strategic
urologic research endeavor (CaPSURE). Cancer. 106:1708–1714. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Habl G, Uhl M, Katayama S, Kessel KA,
Hatiboglu G, Hadaschik B, Edler L, Tichy D, Ellerbrock M, Haberer
T, et al: Acute toxicity and quality of life in patients with
prostate cancer treated with protons or carbon ions in a
prospective randomized phase II Study-The IPI Trial. Int J Radiat
Oncol Biol Phys. 95:435–443. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Petersen SE, Bentzen L, Emmertsen KJ,
Laurberg S, Lundby L and Hoyer M: Development and validation of a
scoring system for late anorectal side-effects in patients treated
with radiotherapy for prostate cancer. Radiother Oncol. 111:94–99.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu B, Fukada K, Zhu H and Kyprianou N:
Prohibitin and cofilin are intracellular effectors of transforming
growth factor beta signaling in human prostate cancer cells. Cancer
Res. 66:8640–8647. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu LI, Fu NI, Luo XU, Li XY and Li XP:
Overexpression of cofilin 1 in prostate cancer and the
corresponding clinical implications. Oncol Lett. 9:2757–2761.
2015.PubMed/NCBI
|
|
9
|
Sundram V, Chauhan SC, Ebeling M and Jaggi
M: Curcumin attenuates β-catenin signaling in prostate cancer cells
through activation of protein kinase D1. PLoS One. 7:e353682012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dang D, Bamburg JR and Ramos DM:
Alphavbeta3 integrin and cofilin modulate K1735 melanoma cell
invasion. Exp Cell Res. 312:468–477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Z, Li Y, Banerjee S, Kong D, Ahmad A,
Nogueira V, Hay N and Sarkar FH: Down-regulation of Notch-1 and
Jagged-1 inhibits prostate cancer cell growth, migration and
invasion, and induces apoptosis via inactivation of Akt, mTOR, and
NF-kappaB signaling pathways. J Cell Biochem. 109:726–736.
2010.PubMed/NCBI
|
|
12
|
Mirkheshti N, Park S, Jiang S, Cropper J,
Werner SL, Song CS and Chatterjee B: Dual targeting of androgen
receptor and mTORC1 by salinomycin in prostate cancer. Oncotarget.
7:62240–62254. 2016.PubMed/NCBI
|
|
13
|
Li CM, Narayanan R, Lu Y, Hurh E, Coss CC,
Barrett CM, Miller DD and Dalton JT:
2-Arylthiazolidine-4-carboxylic acid amides (ATCAA) target dual
pathways in cancer cells: 5′-AMP-activated protein kinase
(AMPK)/mTOR and PI3K/Akt/mTOR pathways. Int J Oncol. 37:1023–1030.
2010.PubMed/NCBI
|
|
14
|
Deeb D, Gao X, Dulchavsky SA and Gautam
SC: CDDO-me induces apoptosis and inhibits Akt, mTOR and NF-kappaB
signaling proteins in prostate cancer cells. Anticancer Res.
27:3035–3044. 2007.PubMed/NCBI
|
|
15
|
Lavie D: The functional groupings of
alpha-elaterin (cucurbitacin E). J Pharm Pharmacol. 10:7821958.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hsu YC, Huang TY and Chen MJ: Therapeutic
ROS targeting of GADD45γ in the induction of G2/M arrest in primary
human colorectal cancer cell lines by cucurbitacin E. Cell Death
Dis. 5:e11982014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan M, Zhu L and Yang Q: Infection of
porcine circovirus 2 (PCV2) in intestinal porcine epithelial cell
line (IPEC-J2) and interaction between PCV2 and IPEC-J2
microfilaments. Virol J. 11:1932014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang WW, Yang JS, Lin MW, Chen PY, Chiou
SM, Chueh FS, Lan YH, Pai SJ, Tsuzuki M, Ho WJ and Chung JG:
Cucurbitacin E Induces G(2)/M Phase Arrest through STAT3/p53/p21
signaling and provokes apoptosis via Fas/CD95 and
mitochondria-dependent pathways in human bladder cancer T24 cells.
Evid Based Complement Alternat Med. 2012:9527622012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li J, Xiong M, Nadavaluru PR, Zuo W, Ye
JH, Eloy JD and Bekker A: Dexmedetomidine attenuates neurotoxicity
induced by prenatal propofol exposure. J Neurosurg Anesthesiol.
28:51–64. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Henning SM, Wang P, Said JW, Huang M,
Grogan T, Elashoff D, Carpenter CL, Heber D and Aronson WJ:
Randomized clinical trial of brewed green and black tea in men with
prostate cancer prior to prostatectomy. Prostate. 75:550–559. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kong Y, Chen J, Zhou Z, Xia H, Qiu MH and
Chen C: Cucurbitacin E induces cell cycle G2/M phase arrest and
apoptosis in triple negative breast cancer. PLoS One.
9:e1037602014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Duncan KL, Duncan MD, Alley MC and
Sausville EA: Cucurbitacin E-induced disruption of the actin and
vimentin cytoskeleton in prostate carcinoma cells. Biochem
Pharmacol. 52:1553–1560. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shan XL, Zhou XY, Yang J, Wang YL, Deng YH
and Zhang MX: Inhibitory effect of cucurbitacin E on the
proliferation of ovarian cancer cells and its mechanism. Chin J
Cancer. 29:20–24. 2010.(In Chinese). View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nakashima S, Matsuda H, Kurume A, Oda Y,
Nakamura S, Yamashita M and Yoshikawa M: Cucurbitacin E as a new
inhibitor of cofilin phosphorylation in human leukemia U937 cells.
Bioorg Med Chem Lett. 20:2994–2997. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu W and Li Y: Salinomycin suppresses LRP6
expression and inhibits both Wnt/β-catenin and mTORC1 signaling in
breast and prostate cancer cells. J Cell Biochem. 115:1799–1807.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yun SM, Jung JH, Jeong SJ, Sohn EJ, Kim B
and Kim SH: Tanshinone IIA induces autophagic cell death via
activation of AMPK and ERK and inhibition of mTOR and p70 S6K in
KBM-5 leukemia cells. Phytother Res. 28:458–464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brown RE, Zotalis G, Zhang PL and Zhao B:
Morphoproteomic confirmation of a constitutively activated mTOR
pathway in high grade prostatic intraepithelial neoplasia and
prostate cancer. Int J Clin Exp Pathol. 1:333–342. 2008.PubMed/NCBI
|
|
28
|
Evren S, Dermen A, Lockwood G, Fleshner N
and Sweet J: Immunohistochemical examination of the mTORC1 pathway
in high grade prostatic intraepithelial neoplasia (HGPIN) and
prostatic adenocarcinomas (PCa): A tissue microarray study (TMA).
Prostate. 70:1429–1436. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zha QB, Zhang XY, Lin QR, Xu LH, Zhao GX,
Pan H, Zhou D, Ouyang DY, Liu ZH and He XH: Cucurbitacin E Induces
Autophagy via downregulating mTORC1 signaling and upregulating AMPK
activity. PLoS One. 10:e01243552015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han MA, Min KJ, Woo SM, Seo BR and Kwon
TK: Eupafolin enhances TRAIL-mediated apoptosis through cathepsin
S-induced down-regulation of Mcl-1 expression and AMPK-mediated Bim
up-regulation in renal carcinoma Caki cells. Oncotarget.
7:65707–65720. 2016.PubMed/NCBI
|
|
31
|
Sun C, Zhang M, Shan X, Zhou X, Yang J,
Wang Y, Li-Ling J and Deng Y: Inhibitory effect of cucurbitacin E
on pancreatic cancer cells growth via STAT3 signaling. J Cancer Res
Clin Oncol. 136:603–610. 2010. View Article : Google Scholar : PubMed/NCBI
|