Introduction
Gastric cancer (GC) is one of the most common
malignancies of the digestive tract and the second leading cause of
cancer-related death worldwide (1,2). The
incidence of GC in China is in the second place of malignant
tumors, only to lung cancer. Especially in the rural areas of
China, the annual incidence of GC is about 36.2/10 million, which
located in the first place of a variety of malignant tumors
(3,4).
The wall of the stomach is composed of four layers, an outer
fibrous membrane called the serosa, a three-ply layer of muscle, a
submucous layer, and a mucous layer called the gastric mucosa.
serosa invasion means subserosa (5,6). GC
usually occurs in the mucosal layer of the stomach wall, and it can
be removed with surgery. However, in cases of metastasis to other
organs, surgical methods are not suitable for the treatment of GC
metastasis (7). The metastasis of GC
include the following forms: i) direct invasion, invasion of the
lower end of the esophagus, duodenum, omentum, colon, liver,
spleen, pancreas and other adjacent organs according to the
different growth sites of GC; ii) hematogenous metastasis, the
common metastasis organs are liver, lung, pancreas, bone and so on,
and hepatic metastasis is commonly seen in the blood route
metastasis of GC iii) peritoneal metastasis iv) lymph node
metastasis. Lymph node metastasis is the main route of metastasis
of GC. The rate of advanced GC lymph node metastasis is ~70%. There
also will occur lymph node metastasis in early GC. The lymph node
metastasis rate of GC is positively related to the depth of tumor
invasion. There were 16 groups of regional lymph nodes draining the
stomach, which can be divided into 3 stations according to their
distance from the stomach. GC is metastasis from primary site to
lymph node through lymphatic network to the first station, then,
the cancer cells with vascular innervating the stomach, transfer to
the second station disposition along the blood vessels surrounding
lymph nodes, and the lymph node metastasis to the distant third
station, can be regarded as a distant metastasis (5,6,8–11).
Therefore, it is of great importance in the early diagnosis and
treatment of GC. In the research field of molecular biology, it is
important to search the suitable molecular markers for GC, and to
provide theoretical basis for clinical treatment.
Receptor of activated protein kinase C (RACK1), a
36-kilodalton (kDa) protein with a propeller-like structure of
seven WD40 (Trp-Asp) motifs, was originally identified on the basis
of its ability to bind the activated form of protein kinase C
(PKC), because it has homology with G protein beta subunit, also
known as guanine nucleotide binding protein (G protein) beta
polypeptide 2-like 1 (GNB2L1), and it is highly conserved in
eukaryotes (12–14). RACK1 is a cellular shuttle protein,
which can be located in cytoplasm, mitochondria, endoplasmic
reticulum and nucleus. As a scaffold protein, it provides a
platform for the interaction of a variety of proteins, thus
integrating inputs from distinct signaling pathways. RACK1
interacts with PKC, phosphodiesterase4D5 (PDE4D5), tyrosine
kinases/phosphatases, and signal transducers and activators of
transcription 3 (STAT3) to regulate a multitude of cellular actions
(15–17). For example, RACK1 interacts with
activated PKC to regulate its intracellular localization (18). RACK1 regulates the stability of JNK or
HIF1α protein as an anchored protein (19). RACK1 interacts with ribosomal proteins
to regulate the translation of intracellular proteins (20). RACK1 combines with signal molecules
from different transduction pathway and plays a key role in a
variety of mammalian animal development (21). Therefore, RACK1 is a multifunctional
scaffold protein, involving in regulating various biological
processes, including signal transduction, immune response, cell
growth, migration, differentiation, angiogenesis, tumor growth,
neuronal response, apoptosis, chromatin remodeling and normal
function of clock (22–23). In recent years, RACK1 is considered to
be an important protein in regulating multiple signaling pathways
and many biological functions of tumor such as proliferation,
apoptosis, migration, especially its role in tumor invasion and
metastasis. RACK1 promotes the invasion and metastasis of tumor and
many kinds of cell function by activating PKC (24). RACK1 combines with PKC to regulate
ribosome translation and promotes the expression of invasion and
metastasis of related factors (25).
RACK1 is highly expressed in breast cancer, colon
cancer, pancreatic ductal adenocarcinoma, melanoma, esophageal
squamous cell carcinoma, lung cancer and oral squamous cell
carcinoma and other tumors, and is considered to be a good marker
(26). For example, RACK1 is an even
superior predictor of breast cancer prognosis compared with
commonly used diagnostic biomarkers (27). The high expression level of RACK1 is
closely related to late clinical status, and silencing of RACK1
inhibits the tumorigenicity of epithelial ovarian cancer in
vitro and in vivo (28).
The high expression of RACK1 is correlated to the pathological
stage and tumor size of lung adenocarcinoma, and is also a
potential marker for clinical diagnosis (29). The expression of RACK1 in oral
squamous cell carcinoma was significantly increased, and the
expression level was negatively correlated with the prognosis of
patients (30). In GC research, RACK1
suppresses the gastric tumorigenesis by negatively regulating Wnt
signaling pathway through stabilizing the β-catenin destruction
complex and act as a tumor suppressor in GC cells (31). Downregulation of RACK1 resulted in
enhance of GC cell metastasis, via promoting the autocrine of
interleukin (IL)-8 in vitro and in vivo (32). RACK1 inhibits GC progression through
the NF-κB pathway (33). However, it
is not clear whether RACK1 plays a tumor-suppressive role in GC
cells through unknown mechanisms. Recent studies have indicated
that RACK1 plays an important role in cell cycle progression, and
it has attracted much attention. Genetic analysis of yeast (pombe
S.) showed that RACK1/Cpc2 regulates cell cycle progression, and
negatively regulates WEE1 homolog (S. pombe) (WEE1) protein levels
and thus regulates mitosis (34).
However, how RACK1 and WEE1 interact to regulate the occurrence and
development of GC is still under investigation.
In the present study, the expression level of RACK1
is decreased in GC and was correlated to TNM stage, tumor
differentiation and lymph node metastasis. In GC cells HGC27, the
mRNA and protein levels of RACK1 were significantly reduced, and
overexpression of RACK1 downregulated WEE1 protein expression, thus
inhibits the growth and proliferation of HGC27 cells.
Mechanistically, RACK1 and WEE1 interacted in HGC27 cells and
co-located in the cytoplasm of HGC27 cells. Our results suggest
that the abnormal expression of RACK1 in the tissues of GC was
involved in the occurrence and development of GC. RACK1 and WEE1
interact to regulate the growth and proliferation of GC cells.
Materials and methods
Patient samples
All 70 tumors were diagnosed as GC and selected to
ensure a broad range of clinical behavior (Table I). GC tissue specimens were obtained
after written informed consent from patients undergoing GC surgery
at the First Affiliated Hospital of Jinzhou Medical University
(Jinzhou, China) during 2012–2013. All patients had not received
chemotherapy and radiotherapy before operation. The study was
approved by the Regional Ethics Committee of Jinzhou Medical
University. Another 30 cases of normal GC adjacent to the edge of
the cancer tissue were selected as the control. Samples of tumor
and pericarcinous tissues were cut from the surgical specimens
immediately fixed in buffered formalin for 48 h, embedded in
paraffin, and sectioned before immunohistochemical staining. All
biopsies were examined and classified by two histopathologist (Jing
Y and Miao G) according to the World Health Organization (WHO)
criteria.
 | Table I.Clinicopathological features of 70
cases of GC. |
Table I.
Clinicopathological features of 70
cases of GC.
| Clinical
characteristic | n (%) |
|---|
| Sex |
|
|
Male | 43 (61.43) |
|
Female | 27 (38.57) |
| Age (years) |
|
|
≥60 | 26 (37.14) |
|
<60 | 44 (62.86) |
| Tumor size
(cm) |
|
| ≤5 | 28 (40) |
|
>5 | 42 (60) |
| Infiltrate
depth |
|
| Mucous
membrane | 0 (0) |
|
Submucosa | 3 (4.29) |
|
Muscular layer | 9 (12.86) |
| Fibrous
membrane | 47 (67.14) |
|
Outside | 11 (15.71) |
| Lymph node
metastasis |
|
| ≤6 | 44 (62.85) |
|
7–14 | 18 (25.71) |
|
≥15 | 8 (11.43) |
| Distant
metastasis |
|
|
Yes | 33 (47.14) |
| No | 37 (52.86) |
| Differentiation
level |
|
|
High-Middle | 30 (42.86) |
|
Low | 40 (57.14) |
| TNM stage |
|
|
I–II | 41 (58.57) |
|
III–IV | 29 (41.43) |
Immunohistochemical staining
Ten-micrometer-thick consecutive sections were cut
and mounted on glass slides. After deparaffinizing, rehydrating,
antigen retrieval, and blocking endogenous peroxidases, the
sections were washed three times in 0.01 mol/l phosphate-buffered
saline (PBS) (8 mmol/l Na2HPO4, 2 mmol/l
NaH2PO4, and 150 mmol/l NaCl) for 5 min each
and blocked for 1 h in PBS supplemented with 0.3% Triton X-100 and
5% normal goat serum, followed by incubation of mouse monoclonal
anti-human RACK1 antibody (610177; 1:200 dilution; BD Biosciences,
San Jose, USA) at 4°C overnight. After brief washes in PBS,
sections were exposed for 2 h to Polink-2 plus® Polymer
HRP Detection System (PV-9002; ZSGB-BIO, Beijing, China) followed
by development with 0.003% H2O2 and 0.03%
3,3′-diaminobenzidine in 0.05 mol/l Tris-HCl (pH 7.6). All sections
were counterstained with hematoxylin.
The immunohistochemical evaluation was performed
according to Xie lab (30) and
slightly modified. The German semiquantitative scoring system was
used, considering the staining intensity and area extent.
Generally, each specimen was assigned a score according to the
percentage of stained cells (0, <5%; 1, 5–25%; 2, 26–50%; 3,
51–75%; 4, 76–100%) and the intensity of the staining (0, no
staining; 1, weak staining; 2, moderate staining and 3, strong
staining). The final immunoreactive score was determined by
multiplying the intensity score by the extent of score of stained
cells. As a result, 9 grades were scored as 0, 1, 2, 3, 4, 6, 8, 9,
and 12. When evaluating the protein expression of RACK1, we defined
a score of 0–9 as low and 12 as high, respectively.
Construction of pcDNA3.1A-flag-RACK1
plasmid
Total RNA was extracted from the human embryonic
kidney (HEK) 293 cells using TRIzol reagent (Invitrogen, Carlsbad,
CA) according to the procedure supplied by the manufacturer.
Extracted RNA (1 µg) was used for cDNA synthesis using the
PrimeScript® RT reagent kit (Takara, Dalian, China). The
reaction system was prepared in a total volume of 20 µl containing
12.5 µl RNA primer mix, 4 µl 5xRT reaction buffer, 2 µl dNTPs, 1 µl
RevertAid reverse transcriptase, 0.5 µl RiboLock RNase inhibitor
and ddH2O up to 20 µl. A pair of primers was designed
based on the RACK1 mRNA sequence (Genebank ID: NM_006098.4):
kpnI (Takara) tailed forward
(5′-ggcggGGTACCatgactgagcagatgacccttcg-3′) and XbaI (Takara)
tailed reverse (5′-ggc ggT CTA GAT TAC TTG TCA TCG TCG TCC TTG TAG
TCg cgt gtg cca atg gtc acc-3′) primers (restriction sites are
underlined). The length of the amplification segment was 3,765 bp.
The PCR mixture was mixed in a total volume of 50 µl containing1 µl
cDNA, 1 µl each primer (20 µmol/l), 5 µl10x EasyPfu Buffer
(Mg2+), 0.5 µl EasyPfu DNA Polymerase, 4 µl dNTP mix
(2.5 mmol/l) and ddH2O up to 50 µl. The PCR program was
started at 94°C for 7 min, followed by 35 cycles at 94°C for 45
sec, 55°C for 30 sec, 72°C for 3 min and completed with a final
extension at 72°C for 10 min. The final PCR products were separated
by electrophoresis using 1% polyacrylamide gels, and the target
fragment was purified and recovered using agarose gel extraction
kit (Axygen, Hangzhou, China). Double restriction enzyme digestion
was applied to the purified target fragments and eukaryotic
expression vector pcDNA3.1A-myc-plus(+), respectively. The enzyme
reaction contained 3 µl target gene fragment or vector
pcDNA3.1A-myc-plus(+), 5 µl 10x Fast Digest buffer, 3 µl KpnI, 3 µl
XbaI and ddH2O up to 50 µl. Under the guidance of
the T4 DNA ligase system instructions (Takara), the purified target
fragment of the RACK1 was directionally ligated into
pcDNA3.1A-myc-plus (+) vector in a 20 µl reaction system containing
15 µl target fragment, 2 µl pcDNA3.1A-myc-plus(+), 1 µl T4 DNA
ligase, 2 µl 10xT4 buffer. The reactants were mixed at 16°C for 2
h, then the ligation was transformed into competent E. coli
DH5a cells and inoculated into Luria-Bertani culture media
containing 100 µg/ml ampicillin. After amplification by shaking the
culture overnight at 37°C, the target plasmids were extracted from
the bacterial liquid according to the instructions for the EndoFree
Maxi Plasmid kit (QIAGEN, Duesseldorf, Germany). The resulting
recombinant eukaryotic expression vector was named
pcDNA3.1A-flag-RACK1. The recombinant plasmids was digested with
KpnI and XbaI, and then evaluated by agarose gel
electrophoresis. The recombinant plasmids was further sequenced to
confirm its sequence by Beijing dingguochangsheng Biotechnology
Co., Ltd. (Beijing, China).
Cell culture and transfection
The gastric epithelial cell line GES-1 (31) and GC cell line HGC27 were used in this
study. The GES-1 cells and HGC27 cells were maintained and cultured
in Dulbecco's modified Eagle medium (Invitrogen) and RPMI-1640
(Invitrogen) medium supplemented with 10% fetal bovine serum (FBS;
PAA Laboratories, Pasching, Austria), 100 U/ml penicillin and 50
µg/ml streptomycin (Biochrom KG, Berlin, Germany) at 37°C in a
humidified 5% CO2 incubator, respectively. The cells
were transiently transfected with the plasmids
pcDNA3.1A-flag-RACK1or pcDNA3.1A using Lipofectamine 2000 reagent
(Invitrogen) following the manufacturer's instructions.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA of each sample was extracted using TRIzol
reagent (Invitrogen) following the manufacturer's protocol. Equal
amounts of RNAs (1 µg) were used as templates in each reaction (50
µl total volume) with the one-step RNA PCR kit (TakaRa, Kyoto,
Japan). The nucleotide sequences of the sense and antisense primers
used for RACK1 (Genebank ID: NM_006098.4) amplification were
5′-GGGGTCACTCCCACTTTGTT-3′ and 5′-AATCTGCCGGTTGTCAGAGG-3′,
respectively (263 bp). The primers for β-actin (Genebank ID:
NM_001101.1) amplification were 5′-TGACGGGGTCACCCACACTGTGCCCATCT-3′
and 5′-CTAGAAGCATTTGCGGTGGACGATGGAGGG-3′ (223 bp). The RT-PCR for
RACK1 and β-actin included one round of reverse transcription at
50°C for 30 min, and 30 cycles of PCR amplification with 94°C for
45 sec, 55°C for 30 sec and 72°C for 1 min. The RT-PCR products
were analyzed on 1% agarose gels and viewed under ultraviolet light
(Bio-Rad, Hercules, CA).
Protein isolation and
immunoblotting
Total protein lysates were prepared in 50 mM
Tris-HCl (pH 7.5) containing 150 mM NaCl and 0.1% NP-40, supplied
with protease inhibitor cocktail (Roche Molecular Biochemical,
Indianapolis, USA). Cell debris was removed by centrifugation for
15 min at 13,000 g at 4°C. Protein concentration was measured with
Pierce™ Bicinchoninic acid (BCA) Protein Assay Kit (Fisher
Scientific). Equal amounts of total protein samples were separated
on 10% sodiumdodecyl sulphate (SDS)-polyacrylamide gels (PAGE)
(Life Technologies, Grand Island, NY, USA) with electrophoresis,
and separated proteins were transferred onto 0.45 µm polyvinylidene
fluoride (PVDF) membranes (Bio-Rad, Hercules, CA, USA) blocked with
Tris-Buffered Saline and Tween-20 (TBST; 20 mM Tris-HCl pH 7.5, 150
mM NaCl and 1% Tween-20) containing 5% fat free dry milk for 2 h
and incubated for 16 h with anti-RACK1 antibody (dilution,
1:1,000), rabbit polyclonal anti-human WEE1 antibody (ab203236;
Abcam, CA, USA) (dilution, 1:1,000) and mouse anti-β ACTIN
monoclonal antibody (TA-09; ZSGB-BIO, Beijing, China) (dilution,
1:1,000) in TBST. After primary antibody incubation, membranes were
washed three times in TBST, followed by incubation with secondary
antibodies cross-linked with horseradish peroxidase (HRP)
(dilution, 1:5,000). Immunoreactive proteins were visualized with
an enhanced chemiluminescence (ECL) detection system (Beyotime,
Jiangsu, China). The relative expression of the target protein was
calculated as the gray value ratio of target protein content to
β-ACTIN content (target protein/β-ACTIN) using Image J software
analysis.
Cell viability analysis
The cell viability measurements were carried out
using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium
bromide (MTT) assay. The total cell number was quantified at 24 h
intervals up to 96 h. Approximately 5×103 of HGC27 cells were
seeded into 96-well plates, washed twice with PBS and 20 µl MTT (5
mg/ml with PBS, pH 7.4) was added to each well. Then, the cells
were incubated at 37°C for 4 h and 150 µl dimethylsulfoxide (DMSO;
Sigma-Aldrich, St. Louis, USA) was added to dissolve the formazan
crystals. After shaking the plate for 10 min, cell viability was
obtained by measuring the absorbance at 490 nm wavelength with
enzyme-labeling instrument (Bio-Tek ELX800, Winooski, VT, USA),
this assay was done six times. The proliferation rate was
calculated according to the following formula: cell viability rate
(%)=average absorbance of experimental group/average absorbance of
blank control group ×100%.
Immunoprecipitation and
immunoblotting
HGC27 cells were collected after transfection for 48
h, washed with PBS (pH 7.4), and lysed in modified RIPA buffer (50
mM Tris (pH 7.8), 150 mM NaCl, 5 mM EDTA, 15 mM MgCl2,
1% NP-40, 0.5% sodium deoxycholate, 1 mM DTT, and 20 mM
N-ethylmaleimide) supplemented with complete protease inhibitor
cocktail. Lysates were cleared by centrifugation at 12,000 g for 15
min at 4°C and quantified protein according to BCA kit. The same
amount of protein precipitation was taken and added 20 µl lysate
and 5 µl 5xSDS sample buffer, boiled for 5 min and preserved at
−20°C. The precipitation was added 20 µl precold mixed suspension
of protein A/G agarose beads (Santa Cruz Biotechnology, Santa Cruz,
USA) and 1 µg IgG and the mixtures were centrifuged at 12,000 g for
30 min at 4°C. The supernatant was incubated on ice for 2 h with
l-2 µg RACK1 antibody, then 50 µl protein A/G agarose
immunoprecipitation reagent was added to each lysate and incubated
with rotation for overnight at 4°C. The beads were retrieved by
centrifugation and washed five times with RIPA buffer and once with
PBS. Protein bound to the beads were eluted by boiling in 2xSDS
sample buffer, separated by SDS-PAGE, transferred protein to PVDF
membrane and blocked as described above. WEE1 antibody were
incubated overnight at 4°C and washed 3 times in TBST followed by
incubation with HRP conjugated goat anti rabbit IgG (dilution ratio
1:5,000) for 2 h. The signals were detected by ECL detection
system.
Immunofluorescence analysis
HGC27 cells seeded on 6-well chamber slides were
fixed in 4% paraformaldehyde, permeabilized in 0.2% Triton X-100
for 5 min and blocked in 1% BSA for 1 h. Protein levels were
detected using RACK1 and WEE1 antibodies overnight at 4°C, The
cells were washed with PBS for 5 min three times followed by
incubation for 45 min at 37°C with Cy3-conjugated or
FITC-conjugated secondary antibodies (Amersham Biosciences). The
coverslips were washed with PBS, stained nucleus with DAPI
(Sigma-Aldrich, St. Louis, USA) for 5 min at room temperature, and
mounted in PBS containing 50% glycerol, and viewed on a Leica laser
scanning confocal microscope equipped with a Photometrics Cool
SnapES N&B camera driven by MetaMorph software (Universal
Imaging Corporation, Downingtown, USA).
Statistical analysis
All data were representative of at least three
independent experiments with similar results. Data were presented
as mean ± SD. Graphpad prism 5 software was used for all
statistical analysis. A student's t test was used to determine
significant differences (two-tailed, P<0.05). Pearson's
correlation coefficients were used to determine whether two
prognosis-related factors were correlated to each other over all
cases.
Results
RACK1 proteins were lowly expressed in
GC patients
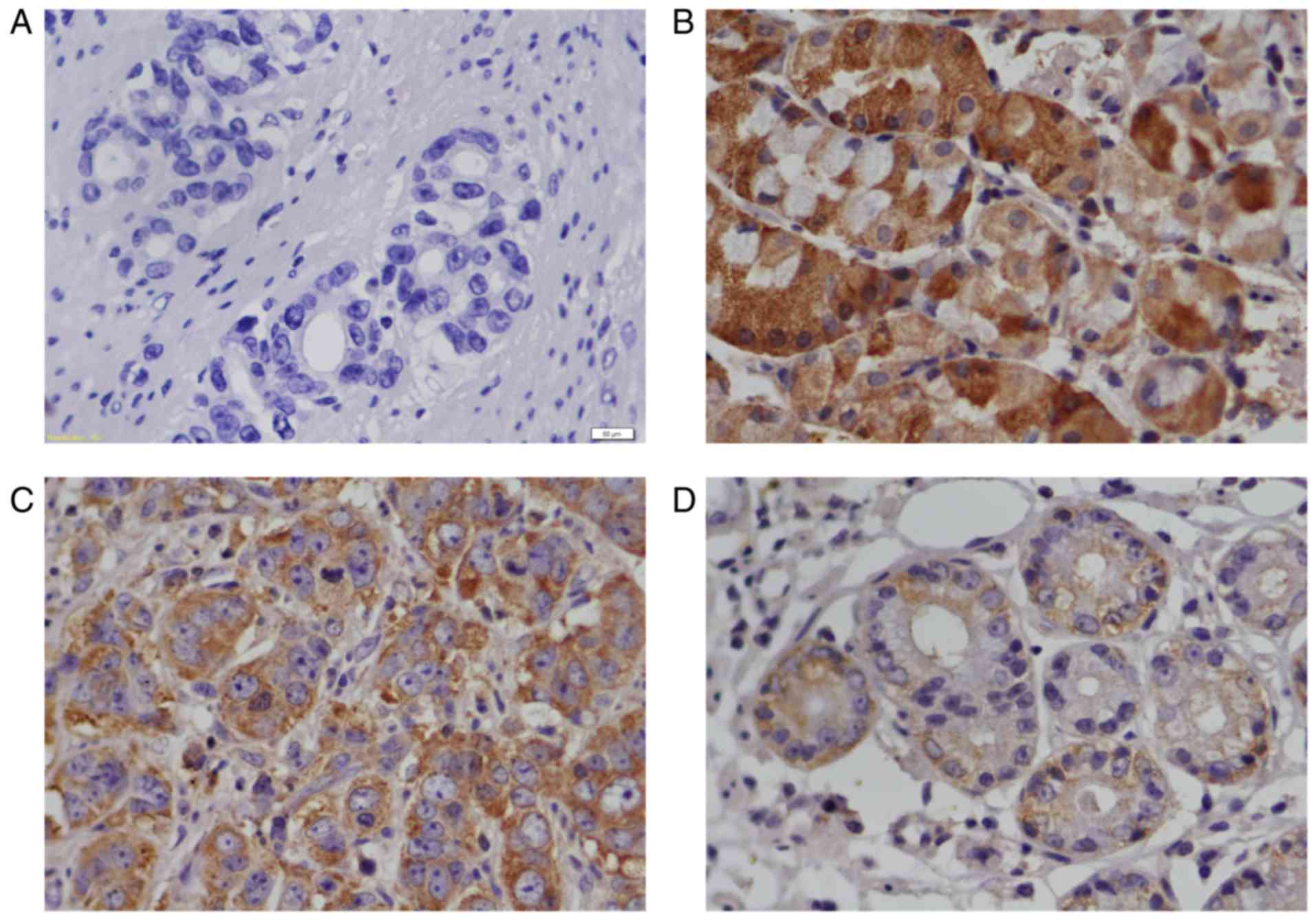
Fig. 1 presents the
results of the immunostaining assay on RACK1 protein expression in
GC tissues and adjacent gastric tissues. The immunohistochemical
staining showed a significant decrease of RACK1 protein in the GC
tissues (Fig. 1C and D, Table II) compared with pericarcinous
tissues (Fig. 1B), and the expression
of RACK1 in high and middle differentiation of GC tissues (Fig. 1C) are higher than that in poorly
differentiated GC tissues (Fig. 1D).
The positive expression of RACK1 protein was located in the
cytoplasm of GC tissues and adjacent gastric tissues (Fig. 1B-D).
| Table II.The protein expression of RACK1 in GC
tissues (n=70) and pericarcinous tissues (n=30). |
Table II.
The protein expression of RACK1 in GC
tissues (n=70) and pericarcinous tissues (n=30).
|
|
| RACK1
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Histological
type | n | High (stage
III–IV) | Low (stage
I–II) | Ratio (%) | P-value |
|---|
| Pericarcinous
tissues | 30 | 26 | 4 | 86.67 | 0.032 |
| GC tissues | 70 | 24 | 46 | 34.29 |
|
The low expression of RACK1 correlated
with pathological parameters in GC patients
The protein expression of RACK1 in stage I–II of GC
tissues was higher than that in stage III–IV of GC tissues
(P<0.01), and the protein expression of RACK1 in the high-middle
differentiated GC tissues was higher than that in the low
differentiated GC tissues (P<0.01), and the decreased expression
of RACK1 was associated withlymph node metastasis (P<0.05). The
protein expression level of RACK1 in GC was related to TNM stage,
tumor differentiation, and lymph node metastasis, while it has no
correlation with age, sex, tumor size and depth of tumor invasion
(Table III).
| Table III.The relationship between expression
of RACK1 in GC tissues and clinical pathological parameters. |
Table III.
The relationship between expression
of RACK1 in GC tissues and clinical pathological parameters.
|
|
| RACK1
expression |
|
|---|
|
|
|
|
|
|---|
| Clinical
features | n | Low | High | P-value |
|---|
| Sex |
|
|
|
|
|
Male | 43 | 31 | 12 | 0.156 |
|
Female | 27 | 15 | 12 |
|
| Age |
|
|
|
|
|
≥60 | 26 | 15 | 11 | 0.227 |
|
<60 | 44 | 31 | 13 |
|
| Tumor Size |
|
|
|
|
| ≤5 | 28 | 18 | 10 | 0.837 |
|
>5 | 42 | 28 | 14 |
|
| Differentiation
level |
|
|
|
|
|
High-Middle | 30 | 12 | 18 | <0.001 |
|
Low | 40 | 34 | 6 |
|
| Infiltrate
depth |
|
|
|
|
|
Submucosa | 3 | 2 | 1 | 0.923 |
|
Muscular layer | 9 | 5 | 4 |
|
| Fibrous
membrane | 47 | 32 | 15 |
|
|
Outside | 11 | 7 | 4 |
|
| Lymph node
metastasis |
|
|
|
|
| ≤6 | 44 | 28 | 16 | 0.028 |
|
7–14 | 18 | 12 | 6 |
|
|
≥15 | 8 | 5 | 3 |
|
| TNM stage |
|
|
|
|
|
StageI–II | 41 | 20 | 21 | <0.001 |
| Stage
III–IV | 29 | 26 | 3 |
|
Downregulation of RACK1 expression in
GC cell lines
Agarose gel electrophoresis analysis showed that the
RACK1 mRNA expression in normal gastric mucosal cells GES-1 was
higher than that in GC cells HGC27 (Fig.
2A). The protein expression of RACK1 in HGC27 cells was lower
than that in GES-1 cells (Fig.
2B).
Overexpression of RACK1 inhibited
tumor growth in vivo
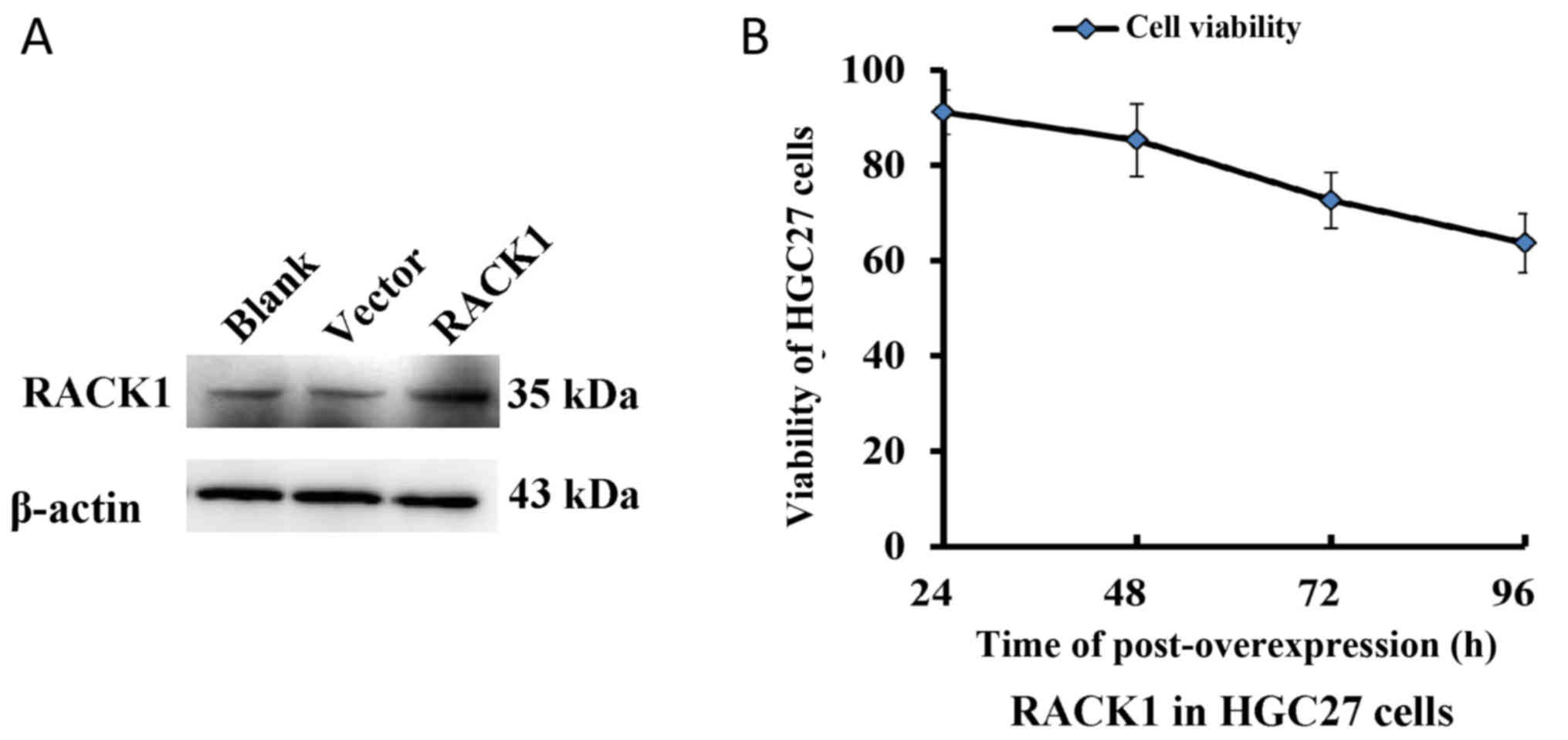
The protein expression of RACK1 was significantly
increased in the HGC27 cells transfected with pcDNA3.1-RACK1
compared with the cells transfected with or without pcDNA3.1 vector
(Fig. 3A). The survival rate of HGC27
cells transfected with pcDNA3.1-RACK1 was significantly reduced at
72 h and 96 h (P<0.01, Fig. 3B),
which showed that the overexpression of RACK1 could significantly
inhibit the growth of HGC27 cells.
The interaction between RACK1 and WEE1
in HCG27 cells
The protein expression of WEE1 was significantly
decreased in HGC27 cells transfected with pcDNA3.1-RACK1 compared
with cells with or without pcDNA3.1 vector, and there was no
significant difference between two control groups (Fig. 4A). In order to explore possible
mechanism(s) underlying RACK1 regulation, interactions between
RACK1 and WEE1 using HGC27 cells via immunoprecipitation and
immunofluorescence analyses were performed. Co-immunoprecipitation
was investigated in HGC27 cells incubated with RACK1 antibody and
detected with WEE1 antibody. As shown in Fig. 4B, RACK1 was co-immunoprecipitated with
WEE1. This interaction was further confirmed to detect endogenous
RACK1 and WEE1 in HGC27 cells. Immunofluorescence analysis revealed
that RACK1 co-localized with WEE1 mainly in the cytoplasm of HGC27
cells (Fig. 4C).
Discussion
RACK1 has been identified as an anchoring or adaptor
protein in multiple intracellular signal transduction pathways and
showed heterogeneity in different tumors (32–36). It
was found that RACK1 is overexpressed in several types of cancers
such as breast, colon, melanomas and lung (36), suggesting that RACK1 is involved in
the occurrence and development of tumor as an oncogene. On the
other hand, recent studies have reported that RACK1 is expressed
lowly in GC tissues and cells, suggesting that RACK1 plays a tumor
suppressor role in the development of GC (32–35).
In order to verify the function of RACK1 in GC
tissues and cells, in this study, firstly, we found that RACK1 was
downregulated in GC tissues using immunohistochemical staining,
research on clinicopathological characteristics of these patients
indicated that RACK1 expression was significantly correlated with
TNM stage, tumor differentiation and lymph node metastasis,
suggesting that the expression level of RACK1 is negatively
regulated the development and metastasis of GC. The study is
consistent with results of Deng et al (32). Secondly, we detected that the mRNA and
protein level of RACK1 in HGC27 cells was significantly lower than
that of GES-1 cells, which is consistent with the result of GC cell
line SGC 7901 (19). Then,
upregulation of RACK1 inhibits the proliferation of HGC27 cells,
which is consistent with the findings of Deng et al and
Yong-Zheng et al (32,34), suggesting that RACK1 negatively
regulates the process of GC cells.
WEE1 is a member of the serine/threonine protein
kinase family involved in terminal phosphorylation and inactivation
of cyclin dependent kinase 1 (CDK1) and is a key regulator of cell
cycle progression (7). Some studies
have found that WEE1 is highly expressed in malignant melanoma,
breast cancer, osteosarcoma and glioma (37–40). Kim
HY reported that high expression of WEE1 is associated with poor
prognosis in male GC patients with lymph node metastasis, and WEE1
expression was detected in 12 GC cell lines, 7 strains with high
WEE1 expression, 5 strains with little or no WEE1 expression, but
there is no information about HGC27 cells (7). Normal cells repair damaged DNA during
G1-arrest, however cancer cells often have deficient G1-arrest and
largely depend on G2-arrest. Thus, cancer cells have increased DNA
damage at the G2-checkpoint compared to normal cells. The molecular
switch for the G2/M transition is held by WEE1 and is pushed
forward by Cell division cycle 25 (CDC25) (7). To study the possible mechanism of
overexpressed RACK1 inhibits the growth and proliferation of GC
cells, we detected the down expression level of WEE1 in HGC27
cells. Therefore, the overexpression of RACK1 in HGC27 cells
destroyed the balance of G2/M checkpoint and inhibited cell
proliferation. To further study the functional relationship between
RACK1 and WEE1 in GC cells, we found RACK1 interacted with WEE1 by
immuneprecipitation and both were co-localized in the cytoplasm by
immunofluorescence using HGC27 cells. Therefore, the interaction
between RACK1 and WEE1 is one of the molecular mechanism in
regulating the growth and proliferation of GC cells.
In summary, the abnormal expression of RACK1 is
involved in the occurrence and development of GC, and negatively
regulate the process of GC cells. The interaction of RACK1 and WEE1
is one of the molecular mechanisms in regulating development of GC.
However, in this study, we verified the interaction and
localization of RACK1 and WEE1 in HGC27 cells with antibody, the
result may be affected by antibody quality, protein molecular
weight, therefore, exogenous plasmids were transfected into the
cells for further testing. Furthermore, how RACK1 and WEE1 interact
to regulate the molecular mechanism(s) of GC is still under
investigation.
Acknowledgments
The authors thank Professor Li Feng of China Medical
University for gifting gastric epithelial cell line GES-1 and
gastric cancer cell line HGC27. The authors also thank Dr. Luan
Zhidong of Jinzhou Medical University for giving
pcDNA3.1A-myc-plus(+) plasmid. The authors acknowledge grant
supports received from the National Nature Science Foundation of
China (grant nos. 31371173, 81270698, and 81401199) and the Project
of Science and Technology Plan of Liaoning Province (grant no.
2015020697).
Glossary
Abbreviations
Abbreviations:
|
RACK1
|
receptor of activated C Kinase 1
|
|
GC
|
gastric cancer
|
|
GNB2L1
|
guanine nucleotide binding protein
beta polypeptide 2-like 1
|
|
WEE1
|
WEE1 homolog (S. pombe)
|
References
|
1
|
McLean MH and El-Omar EM: Genetics of
gastric cancer. Nat Rev Gastroenterol Hepatol. 11:664–674. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Badgwell B: Multimodality therapy of
localized gastric adenocarcinoma. J Natl Compr Canc Netw.
14:1321–1327. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fang M, Wu J, Lai X, Ai H, Tao Y, Zhu B
and Huang L: CD44 and CD44v6 are correlated with gastric cancer
progression and poor patient prognosis: Evidence from 42 studies.
Cell Physiol Biochem. 40:567–578. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang J, Fei X, Wu W, Chen X, Su L, Zhu Z
and Zhou Y: SLC7A5 Functions as a Downstream Target Modulated by
CRKL in Metastasis Process of Gastric Cancer SGC-7901 Cells. PLoS
One. 11:e01661472016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lawrence W. Way and Gerard M. Doherty:
Current surgical diagnosis & treatment. 11th. McGraw-Hill
Companies, Inc.; New York: pp. 285–287. 2003
|
|
6
|
Japanese Gastric Cancer Association:
Japanese classification of gastric carcinoma: 3rd English edition.
Gastric Cancer. 14:101–112. 2001.
|
|
7
|
Kim HY, Cho Y, Kang H, Yim YS, Kim SJ,
Song J and Chun KH: Targeting the WEE1 kinase as a molecular
targeted therapy for gastric cancer. Oncotarget. 7:49902–49916.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amin MB, Edge S, Greene F, Byrd DR,
Brookland RK, Washington MK, Gershenwald JE, Compton CC, Hess KR,
Sullivan DC, et al: AJCC Cancer Staging Manual. 8th. Springer; New
York: pp. 93–99. 2017
|
|
9
|
Roder JD, Böttcher K, Busch R, Wittekind
C, Hermanek P and Siewert JR: Classification of regional lymph node
metastasis from gastric carcinoma. German Gastric Cancer Study
Group. Cancer. 82:621–631. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lim DH, Kim HS, Park YS, Lee J, Park SH,
Lim HY, Ji SH, Park MJ, Yi SY, An JY, et al: Metastatic lymph node
in gastric cancer; is it a real distant metastasis? BMC Cancer.
10:252010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Akagi T, Shiraishi N and Kitano S: Lymph
Node Metastasis of Gastric Cancer. Cancers (Basel). 3:2141–2159.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li JJ and Xie D: RACK1, a versatile hub in
cancer. Oncogene. 34:1890–1898. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gandin V, Senft D, Topisirovic I and Ronai
ZA: RACK1 function in cell motility and protein synthesis. Genes
Cancer. 4:369–377. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gibson TJ: RACK1 research-ships passing in
the night? FEBS Lett. 586:2787–2789. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Robles MS, Boyault C, Knutti D,
Padmanabhan K and Weitz CJ: Identification of RACK1 and protein
kinase Calpha as integral components of the mammalian circadian
clock. Science. 327:463–466. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bird RJ, Baillie GS and Yarwood SJ:
Interaction with receptor for activated C-kinase 1 (RACK1)
sensitizes the phosphodiesterase PDE4D5 towards hydrolysis of cAMP
and activation by protein kinase C. Biochem J. 432:207–216. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang W, Zong CS, Hermanto U,
Lopez-Bergami P, Ronai Z and Wang LH: RACK1 recruits STAT3
specifically to insulin and insulin-like growth factor 1 receptors
for activation, which is important for regulating
anchorage-independent growth. Mol Cell Biol. 26:413–424. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Haberman Y, Alon LT, Eliyahu E and Shalgi
R: Receptor for activated C kinase (RACK) and protein kinase C
(PKC) in egg activation. Theriogenology. 75:80–89. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li M, Liu Y, Jin F, Sun X, Li Z, Liu Y,
Fang P, Shi H and Jiang X: Endothelin-1 induces hypoxia inducible
factor 1α expression in pulmonary artery smooth muscle cells. FEBS
Lett. 586:3888–3893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu M, Peng P, Wang J, Wang L, Duan F, Jia
D, Ruan Y and Gu J: RACK1-mediated translation control promotes
liver fibrogenesis. Biochem Biophys Res Commun. 463:255–261. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng D, Zhu X, Barchiesi F, Gillespie DG,
Dubey RK and Jackson EK: Receptor for activated protein kinase C1
regulates cell proliferation by modulating calcium signaling.
Hypertension. 58:689–695. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Su J, Xu J and Zhang S: RACK1, scaffolding
a heterotrimeric G protein and a MAPK cascade. Trends Plant Sci.
20:405–407. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia X, Zhang L and Mao X: S-propranolol
protected H9C2 cells from ischemia/reperfusion-induced apoptosis
via downregultion of RACK1 Gene. Int J Clin Exp Patho.
l8:10335–10344. 2015.
|
|
24
|
Grosso S, Volta V, Sala LA, Vietri M,
Marchisio PC, Ron D and Biffo S: PKCbetaII modulates translation
independently from mTOR and through RACK1. Biochem J. 415:77–85.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ruan Y, Sun L, Hao Y, Wang L, Xu J, Zhang
W, Xie J, Guo L, Zhou L, Yun X, et al: Ribosomal RACK1 promotes
chemoresistance and growth in human hepatocellular carcinoma. J
Clin Invest. 122:2554–2566. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Campagne C, Julé S, Alleaume C, Bernex F,
Ezagal J, Château-Joubert S, Estrada M, Aubin-Houzelstein G,
Panthier JJ and Egidy G: Canine melanoma diagnosis: RACK1 as a
potential biological marker. Vet Pathol. 50:1083–1090. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao XX, Xu JD, Liu XL, Xu JW, Wang WJ, Li
QQ, Chen Q, Xu ZD and Liu XP: RACK1: A superior independent
predictor for poor clinical outcome in breast cancer. Int J Cancer.
127:1172–1179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lin Y, Cui M, Teng H, Wang F, Yu W and Xu
T: Silencing the receptor of activated C-kinase 1 (RACK1)
suppresses tumorigenicity in epithelial ovarian cancer in vitro
in vivo. Int J Oncol. 44:1252–1258. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi YY, Lee SY, Lee WK, Jeon HS, Lee EB,
Lee HC, Choi JE, Kang HG, Lee EJ, Bae EY, et al: RACK1 is a
candidate gene associated with the prognosis of patients with early
stage non-small cell lung cancer. Oncotarget. 6:4451–4466. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang X, Liu N, Ma D, Liu L, Jiang L, Zhou
Y, Zeng X, Li J and Chen Q: Receptor for activated C kinase 1
(RACK1) promotes the progression of OSCC via the AKT/mTOR pathway.
Int J Oncol. 49:539–548. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li X, Ke Q, Li Y, Liu F, Zhu G and Li F:
DGCR6L, a novel PAK4 interaction protein, regulates PAK4-mediated
migration of human gastric cancer cell via LIMK1. Int J Biochem
Cell Biol. 42:70–79. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Deng YZ, Yao F, Li JJ, Mao ZF, Hu PT, Long
LY, Li G, Ji XD, Shi S, Guan DX, et al: RACK1 suppresses gastric
tumorigenesis by stabilizing the β-catenin destruction complex.
Gastroenterology. 142:812–823.e15. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen L, Min L, Wang X, Zhao J, Chen H, Qin
J, Chen W, Shen Z, Tang Z, Gan Q, et al: Loss of RACK1 promotes
metastasis of gastric cancer by inducing a miR-302c/IL8 signaling
loop. Cancer Res. 75:3832–4381. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yong-Zheng X, Wan-Li M, Ji-Ming M and
Xue-Qun R: Receptor for activated protein kinase C 1 suppresses
gastric tumor progression through nuclear factor-kB pathway. Indian
J Cancer. 52 Suppl 3:E172–E175. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Núñez A, Franco A, Soto T, Vicente J,
Gacto M and Cansado J: Fission yeast receptor of activated C kinase
(RACK1) ortholog Cpc2 regulates mitotic commitment through Wee1
kinase. J Biol Chem. 285:41366–41373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang F, Osawa T, Tsuchida R, Yuasa Y and
Shibuya M: Downregulation of receptor for activated C-kinase 1
(RACK1) suppresses tumor growth by inhibiting tumor cell
proliferation and tumor-associated angiogenesis. Cancer Sci.
102:2007–2013. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mueller S, Hashizume R, Yang X, Kolkowitz
I, Olow AK, Phillips J, Smirnov I, Tom MW, Prados MD, James CD, et
al: Targeting Wee1 for the treatment of pediatric high-grade
gliomas. Neuro Oncol. 16:352–360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Magnussen GI, Holm R, Emilsen E, Rosnes
AK, Slipicevic A and Flørenes VA: High expression of wee1 is
associated with poor disease-free survival in malignant melanoma:
Potential for targeted therapy. PLoS One. 7:e382542012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kreahling JM, Foroutan P, Reed D, Martinez
G, Razabdouski T, Bui MM, Raghavan M, Letson D, Gillies RJ and
Altiok S: Wee1 inhibition by MK-1775 leads to tumor inhibition and
enhances efficacy of gemcitabine in human sarcomas. PLoS One.
8:e575232013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Murrow LM, Garimella SV, Jones TL, Caplen
NJ and Lipkowitz S: Identification of WEE1 as a potential molecular
target in cancer cells by RNAi screening of the human tyrosine
kinome. Breast Cancer Res Treat. 122:347–357. 2010. View Article : Google Scholar : PubMed/NCBI
|