Introduction
Lung cancer is the main cause of cancer-related
death worldwide. Non-small cell lung carcinoma (NSCLC), accounting
for approximately 80% of lung cancer, is the most common
histopathological type. Surgery is the only treatment method for
early stages of NSCLC, but the 5-year overall survival rate is only
40% (1). Although remarkable progress
has been made in the past decades, the molecular mechanisms of
NSCLC carcinogenesis are still not clear.
Long non-coding RNAs (lncRNAs), which are non-coding
RNAs with more than 200 nucleotides in length, are frequently
cell-type specific and contribute to the progression of multiple
human tumors (2,3). Moreover, lncRNAs also have the potential
function as diagnostic or prognostic markers of cancer (4–10). Nuclear
enriched abundant transcript 1 (NEAT1) is a nuclear-restricted
lncRNA that has two isoforms: NEAT1-1 and NEAT1-2 (11). It was recently revealed as an
essential architectural component of paraspeckle structure
(12,13). NEAT1 plays a role in several
biological processes, including stress response and cellular
differentiation through paraspeckles pathway (14). NEAT1 is also a marker for poor
survival of breast cancer patients (15). In prostate cancer, NEAT1 was proved to
promote oncogenic growth by altering gene promoters and then
inducing transcription (16). But
until now, the specific mechanism of NEAT1 is still not clear in
NSCLC.
The aim of this study is to explore the function and
mechanism of NEAT1 in NSCLC. We first found the increased
expression of NEAT1 in NSCLC and its correlation with poor
survival. Then, we found that NEAT1 (represented by NEAT1-2)
harbors three conserved miR-377-3p cognate sites, and E2F3 harbors
two conserved miR-377-3p cognate sites using Biological information
technology. At last, our results showed that NEAT1 might be a
crucial oncogenic regulator involved in NSCLC development by
functioning as a ceRNA, which binding to miR-377-3p and then
further activated the E2F3 pathway.
Materials and methods
Tissue collection
All the samples (20 paired NSCLC tissues and
adjacent normal tissues) were obtained from patients suffering from
NSCLC who operated at the second affiliated hospital of Harbin
Medical University between 2011.5 and 2012.8. All these patients
did not recieve any local or systemic treatment before operation.
All collected tissue samples were immediately treated by liquid
nitrogen, then stored at −80°C before RNA extraction. All patients
signed their informed consent. The study has got approval from the
Research Ethics Committee of Harbin Medical University, and thus
meets the standards of the Declaration of Helsinki in its revised
version of 1975 and its amendments of 1983, 1989, and 1996.
Cell lines and culture conditions
Normal human embryonic lung fibroblasts cell line
(HELF) and several NSCLC cancer cell lines, including A549, SPC-A1,
H1299, 95D, SK-MES-1, and NCI-H520, were used in this study. These
cells were cultured in RPMI-1640 (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) or Dulbecco's modified Eagle's medium
(DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
fetal bovine serum (FBS) at 37°C in humidified air with 5%
CO2.
Transfection of cell lines
The siRNA (small interfering RNA) sequences
(GenePharma, Shanghai, China) were shown in Table I. Synthetic sequence-scrambled siRNA
was used as negative controls (NC). Human E2F3 gene was ligated
into pGCMV vector (GenePharma). The empty vector was used as NC.
Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) was used to
complete transfection in A549 and H1299 cells according to the
manufacturer's protocol.
 | Table I.Sequence of NEAT1 siRNA. |
Table I.
Sequence of NEAT1 siRNA.
| NEAT1 siRNA | Sequence |
|---|
| siRNA1 |
5′-GCCAUCAGCUUUGAAUAAAUU-3′ |
| siRNA2 |
5′-GGUGUUAUCAAGUGAAUUAUU-3′ |
| siRNA3 |
5′-GCCUUGUAAAUGCCUAUAUUU-3′ |
Cell proliferation ability
analysis
Different groups of cells were seeded into 6 wells
plate at 1×105 cells per well. The cell number was
counted for three consecutive days and then growth curve was drawn
according to the cell number at 24, 48 and 72 h. 1×106
cells were prepared for cell cycle analysis. First, the cells were
fixed with 70% cold ethanol. After 12 h at 4°C, the cells were
washed with PBS and stained with propidium iodide (PI). Finally,
flow cytometry (BD Biosciences, San Jose, CA, USA) was conducted to
analyze cell cycle.
Luciferase reporter assays
The wild-type and mutant 3′-UTR of human E2F3 or
lncRNA NEAT1 were inserted into the pmiR-RB-REPORT TM (Guangzhou
RiboBio Co., Ltd., Guangzhou, China) through XhoI and NotI sites.
Co-transfected wild-type/mutant reporter plasmid and miR-Ribo TM
negative control into A549 and H1299 cells by Lipofectamine 2000
(Invitrogen), and then measured luciferase activities using the
Dual-Luciferase Reporter Assay system 48 h post-transfection
according to the manufacturer's instructions (Promega Corp.,
Madison, WI, USA).
RNA isolation and quantitative reverse
transcription polymerase chain reaction (qRT-PCR)
Total RNA in cells and tissues was extracted using
Trizol reagent (Invitrogen). RNA was reversed into cDNA through
reverse transcription kits (Guangzhou RiboBio Co., Ltd.). Then the
levels of target mRNAs were detected using qRT-PCR by
SYBR® Premix Ex Taq™II (Takara Bio, Inc., Otsu, Japan)
in the ABI PRISM® 7300 real-time PCR system (Applied
Biosystems, Foster City, CA, USA). GAPDH and U6 were used as
endogenous controls. The primer sequences involved in this study
were shown in Table II.
| Table II.Sequence of primers. |
Table II.
Sequence of primers.
| Gene | Primer sequence |
|---|
| GAPDH | Forward
5′-TGTGGGCATCAATGGATTTGG-3′ |
|
| Reverse
5′-ACACCATGTATTCCGGGTCAAT-3′ |
| NEAT1_2 | Forward
5′-CAGTTAGTTTATCAGTTCTCCCATCCA-3′ |
|
| Reverse
5′-GTTGTTGTCGTCACCTTTCAACTCT-3′ |
| E2F3 | Forward
5′-AGAAAGCGGTCATCAGTACCT-3′ |
|
| Reverse
5′-TGGACTTCGTAGTGCAGCTCT-3′ |
| miR-377-3p | RT:
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACacaa-3′ |
| U6 | RT:
5′-AAAATATGGAACGCTTCACGAATTTG-3′ |
| miR-377-3p | Forward
5′-GGGAGGCAGTGTATTGTTA-3′ |
|
| Reverse
5′-GTCGTATCCAGTGCAGGGTCCGAGGT-3′ |
Western blot analysis
Total protein was obtained from the A549 and H1299
cells after 48 h of transfection using cell lysis reagent. BCA
protein assay reagent kit (Thermo Fisher Scientific, Inc.) was used
to detected the protein concentrations in supernatant. The protein
was separated in 10% SDS-polyacrylamide gels, transferred to PVDF
membranes (Hertfordshire, UK), incubated in 5% milk powder to block
non-specific reaction, and then treated with antibodies overnight
at 4°C, including anti-E2F3 (1:1,000 dilutions; Affinity),
anti-cyclin D1 (1:1,000 dilutions; Affinity), anti-cyclinD2
(1:1,000 dilutions; Affinity), anti-CDK4 (1:1,000 dilutions;
Affinity), anti-p21 (1:1,000 dilutions; Affinity), anti-p57
(1:1,000 dilutions; Affinity) and anti-GADPH (1:5,000 dilutions;
Affinity). At last, the membranes were incubated with second
antibody (conjugated to horseradish peroxidase, 1:12,000 dilutions)
for 2 h and exposed to ECL Advance reagent (GE Healthcare
Biosciences, Buckinghamshire, UK) for 2 min. The band was
visualized with Fluor S Multimager and Quantity One 4.1 (Bio-Rad
Laboratories, Hercules, CA, USA).
Statistical analysis
In the present study, all the data are presented as
means ± SD. A two-tailed paired t-test was used to reflect the
difference between groups. P<0.05 indicated that the difference
was statistically significant.
Results
NEAT1 is up-regulated in NSCLC tissues
and correlates with poor prognosis
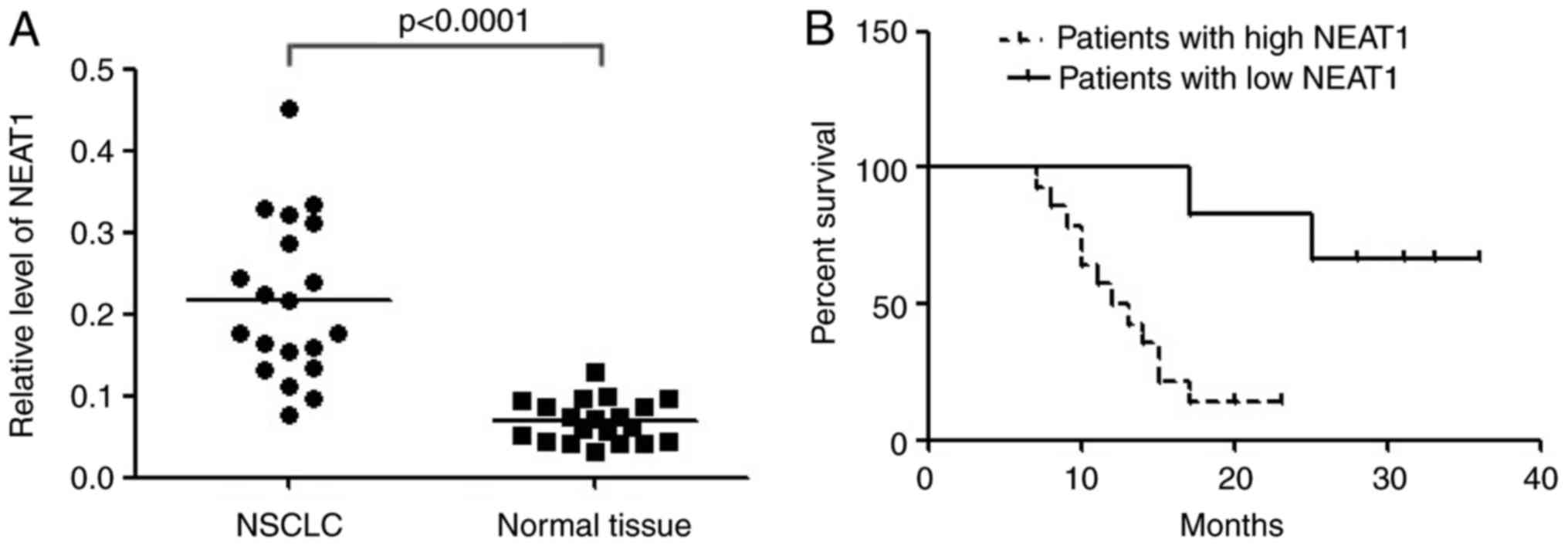
In order to validate the differential expression,
NEAT1 was detected in 20 paired NSCLC tissues and adjacent normal
tissues with qRT-PCR. The result showed that NEAT1 expression was
significantly up-regulated in cancerous tissues than normal
counterparts (Fig. 1A). Compared with
patients whose cancer tissue expressed less NEAT1 (≤2 folds of
increase, n=6), patients whose cancer tissue expressed more NEAT1
(>2 folds of increase, n=14) had shorter overall survival
according to Kaplan-Meier survival analysis (Fig. 1B). The results demonstrated that high
levels of NEAT1 were associated with NSCLC poor prognosis.
Upregulation of NEAT1 might play important roles in NSCLC.
NEAT1 promotes NSCLC cells growth in
vitro
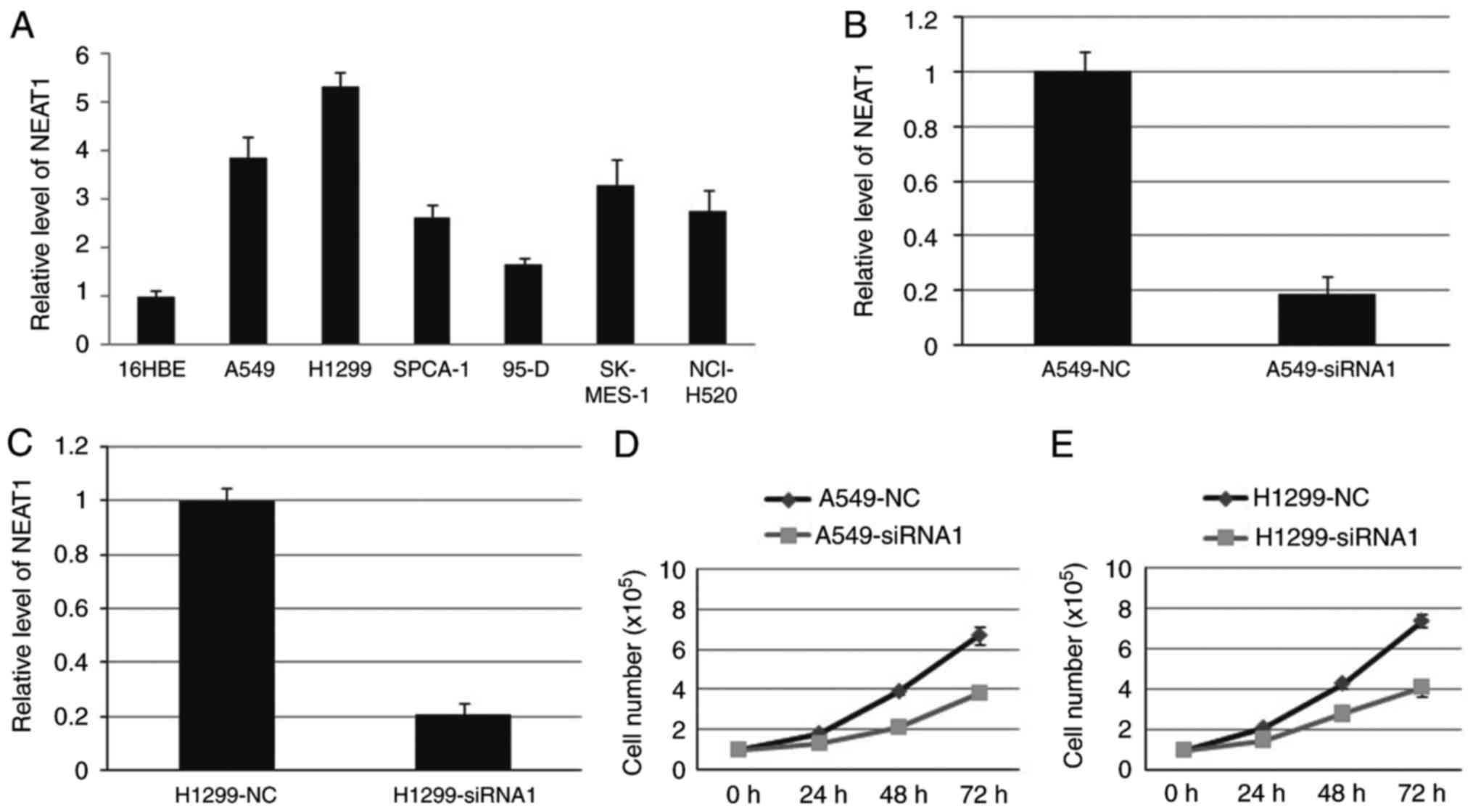
We first detected NEAT1 expression in NSCLC cell
lines, including A549, H1299, SPCA-1, 95-D, SK-MES-1 and NCI-H520
cell line. The result showed that the expression level of NEAT1 in
NSCLC cell lines was higher than that in 16HBE cell line (Fig. 2A). Then, endogenous NEAT1 expression
was down-regulated by approximately 80% through small interfering
RNAs (siRNAs) in A549 and H1299 cells (Fig. 2B and C). Compared with blank cells,
the proliferation ability was decreased in A549-siRNA1 and
H1299-siRNA1 cells (Fig. 2D and E).
Cell cycle analysis indicated that deletion of NEAT1 induced cell
cycle arrest in G1 phase.
NEAT1 promotes E2F3 expression in part
through competitive binding miRNA-377-3p
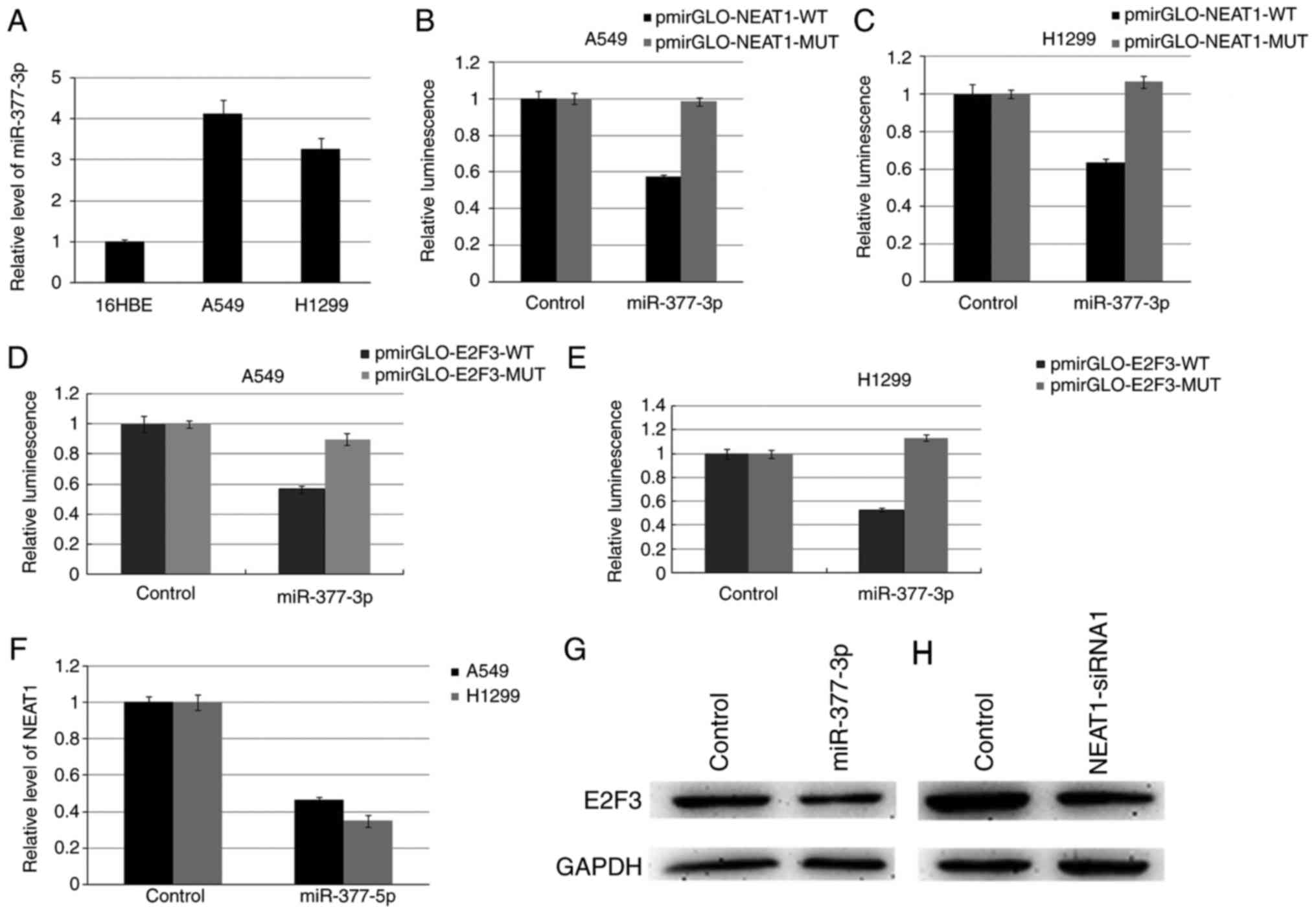
In view of the potential function of lncRNA as a
ceRNA or a molecular sponge of miRNAs, we screened the miRNAs
interacting with NEAT1 using the bioinformatics, starBase2.0. Among
these miRNAs, miR-377-3p (MIMAT0000730) was selected for further
study. We first verified miR-377-3p expression in A549 and H1299
cells, and it is significantly higher than 16HBE cell (Fig. 3A). Dual-luciferase reporter assay
indicated that miR-377-3p could decrease the luciferase activity of
pmirGLO-NEAT1-WT (wild-type), however, it had no affect on the
pmirGLO-NEAT1-MUT (Fig. 3B and C).
Targetscan, miRanda and PicTar were used to predict the possible
targets of miR-377-3p, and found that E2F3 involving in cell cycle
regulation harbors two conserved miR-377-3p cognate sites.
Luciferase reporter assays indicated that luciferase activity was
declined more than 50% after co-transfected with miR-377-3p and the
reporter vector with E2F3-3′-WT (Fig. 3D
and E). Up-regulating miR-377-3p in A549 and H1299 cells
suppressed the expression of NEAT1 and E2F3 (Fig. 3F and G). Inhibition of NEAT1 resulted
in down-regulation of E2F3 protein level (Fig. 3H).
E2F3 promotes NSCLC proliferation
through regulating cell cycle in NSCLC
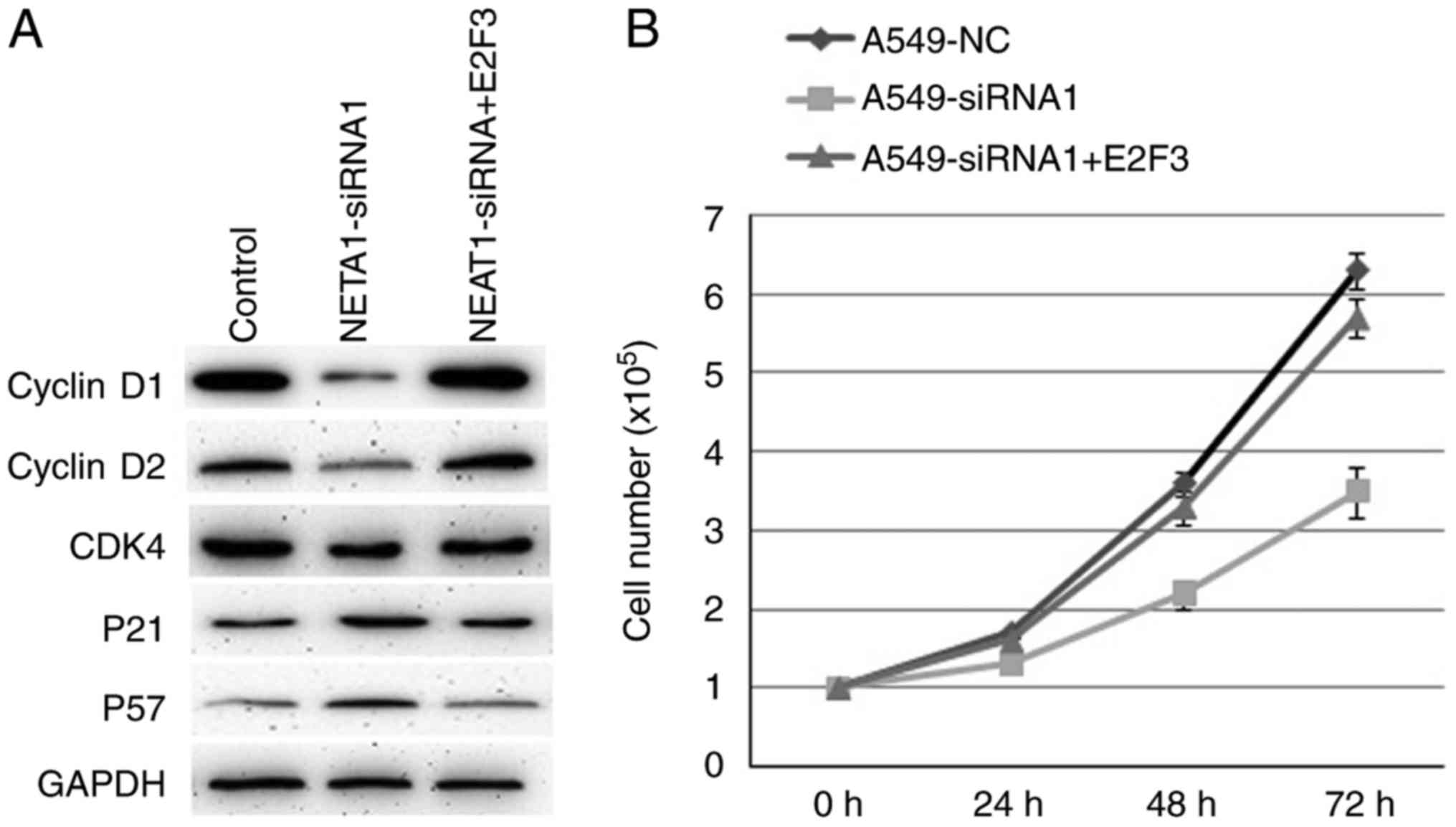
Inhibiting NEAT1 in A549 cells suppressed the
protein expression of cyclinD1, cyclinD2 and CDK4, whereas
increased the protein levels of p21 and p57 (Fig. 4A). Consistently, over-expression of
E2F3 in A549-siRNA1 cells reversed the expression of these proteins
mentioned above (Fig. 4A).
Over-expression of E2F3 also reversed the cell proliferation
ability inhibited by decreased NEAT1 (Fig. 4B). So, E2F3 mediated the cell
proliferation induced by NEAT1 by regulating cell cycle associated
proteins expression.
Discussion
Recent studies have demonstrated lncRNAs play an
important role in human cancers (17–20). Our
research indicated that NEAT1 was up-regulated in NSCLC. NEAT1
acted as an oncogene to promote the proliferation ability of NSCLC
cells, which is largely due to its ceRNAs function to suppress
miR-377-3p and subsequently derepress E2F3, and then activate the
E2F3 signaling pathway. This study is important to illustrate the
relationship between NEAT1 and NSCLC development.
This study indicated that NEAT1 was significantly
up-regulated in NSCLC tissues compared to adjacent normal lung
tissues. Particularly, our study also demonstrated an association
between increased NEAT1 expression levels and NSCLC poor prognosis.
This result indicated that NEAT1 expression may be a helpful
biomarker for doctor to recognize patients with poor prognosis. It
was in accordance with Pan and his colleagues' findings (21). In addition, NEAT1 significantly
improved NSCLC cell proliferative ability. In summary, the results
of this research suggested that NEAT1 may act as an oncogene as
well as contribute significantly to NSCLC development and
progression.
It has been reported that paraspeckles pathway is
one of the molecular mechanisms by which NEAT1 plays roles in
controlling several biological processes (12,13).
Recently, NEAT1 was found to repress the transcription of some
genes by protein sequestration into paraspeckles (22). Here, we discovered that NEAT1 might
function by another potential molecular mechanism in NSCLC
progression-acting as ‘molecular sponges’ to regulate microRNAs.
Many studies have proved that lncRNAs can work as ceRNAs to
regulate microRNAs, such as GAS5 (23). For another instance, CCAT1 can promote
the development of gallbladder cancer by acting as miR-218-5p
sponges (24). In the present study,
we found that NEAT1, acting as a ceRNA, bound miR-377-3p and
restricted its function.
As we have known, miRNA can regulate the expression
of multiple target genes generally. It was reported that miR-377
can inhibit human clear cell renal cell carcinoma by targeting ETS1
(25), and miR-377 can also suppress
cell proliferation and invasion in hepatocellular carcinoma by
inhibiting TIAM1 expression (26).
Additionally, Wen et al revealed miR-377 regulates
mesenchymal stem cell-induced angiogenesis in ischemic hearts by
targeting VEGF (27). But among the
target genes of miR-377, E2F3 transcription factor caught our
attention. The G1/S transition of the cell cycle is regulated in
part by E2Fs transcription factors and is a tightly orchestrated
process that culminates in DNA synthesis (28). E2Fs regulate expression of genes
involved in many cellular processes, including apoptosis, mitosis,
differentiation, and development with E2F1-3 as transcriptional
activators and E2F4-8 as transcriptional repressors of these genes
(29). E2F3 plays an essential role
in normal proliferation (30).
Furthermore, many cancers have been found to be associated with
aberrant E2F expression, including lung cancer, bladder cancer and
prostate cancer (31,32). In our study, we proved that E2F3 can
increase the protein expression of cyclinD1, cyclinD2 and CDK4,
whereas suppressing the protein levels of p21 and p57.
In summary, our results suggested lncRNA NEAT1
promoted the growth of NSLC by competitively binding to miR-377 and
then removing the suppressive effect of miR-377 on E2F3. NEAT1 may
also be seen as a factor that prompt the traces of NSCLC. Our
research has provided a better understanding of the NEAT1 function
in NSCLC, which might facilitate the development of lncRNA-directed
diagnostics and therapeutics against this disease.
Acknowledgements
This work was supported by the Heilongjiang
Provincial Health Bureau (No. 2016-176).
References
|
1
|
Laskin JJ and Sandler AB: State of the art
in therapy for non-small cell lung cancer. Cancer Invest.
23:427–442. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Orom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou M, Sun Y, Sun Y, Xu W, Zhang Z, Zhao
H, Zhong Z and Sun J: Comprehensive analysis of lncRNA expression
profiles reveals a novel lncRNA signature to discriminate
nonequivalent outcomes in patients with ovarian cancer. Oncotarget.
7:32433–32448. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhou M, Guo M, He D, Wang X, Cui Y, Yang
H, Hao D and Sun J: A potential signature of eight long non-coding
RNAs predicts survival in patients with non-small cell lung cancer.
J Transl Med. 13:2312015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou M, Zhao H, Wang Z, Cheng L, Yang L,
Shi H, Yang H and Sun J: Identification and validation of potential
prognostic lncRNA biomarkers for predicting survival in patients
with multiple myeloma. J Exp Clin Cancer Res. 34:1022015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou M, Xu W, Yue X, Zhao H, Wang Z, Shi
H, Cheng L and Sun J: Relapse-related long non-coding RNA signature
to improve prognosis prediction of lung adenocarcinoma. Oncotarget.
7:29720–29738. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou M, Wang X, Shi H, Cheng L, Wang Z,
Zhao H, Yang L and Sun J: Characterization of long non-coding
RNA-associated ceRNA network to reveal potential prognostic lncRNA
biomarkers in human ovarian cancer. Oncotarget. 7:12598–12611.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou M, Zhao H, Xu W, Bao S, Cheng L and
Sun J: Discovery and validation of immune-associated long
non-coding RNA biomarkers associated with clinically molecular
subtype and prognosis in diffuse large B cell lymphoma. Mol Cancer.
16:162017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yuan SX, Yang F, Yang Y, Tao QF, Zhang J,
Huang G, Yang Y, Wang RY, Yang S, Huo XS, et al: Long noncoding RNA
associated with microvascular invasion in hepatocellular carcinoma
promotes angiogenesis and serves as a predictor for hepatocellular
carcinoma patients' poor recurrence-free survival after
hepatectomy. Hepatology. 56:2231–2241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hutchinson JN, Ensminger AW, Clemson CM,
Lynch CR, Lawrence JB and Chess A: A screen for nuclear transcripts
identifies two linked noncoding RNAs associated with SC35 splicing
domains. BMC Genomics. 8:392007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sasaki YT, Ideue T, Sano M, Mituyama T and
Hirose T: MENepsilon/beta noncoding RNAs are essential for
structural integrity of nuclear paraspeckles. Proc Natl Acad Sci
USA. 106:2525–2530. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sunwoo H, Dinger ME, Wilusz JE, Amaral PP,
Mattick JS and Spector DL: MEN epsilon/beta nuclear-retained
non-coding RNAs are up-regulated upon muscle differentiation and
are essential components of paraspeckles. Genome Res. 19:347–359.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Naganuma T and Hirose T: Paraspeckle
formation during the biogenesis of long non-coding RNAs. RNA Biol.
10:456–461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Choudhry H, Albukhari A, Morotti M, Haider
S, Moralli D, Smythies J, Schödel J, Green CM, Camps C, Buffa F, et
al: Tumor hypoxia induces nuclear paraspeckle formation through
HIF-2α dependent transcriptional activation of NEAT1 leading to
cancer cell survival. Oncogene. 34:45462015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chakravarty D, Sboner A, Nair SS,
Giannopoulou E, Li R, Hennig S, Mosquera JM, Pauwels J, Park K,
Kossai M, et al: The oestrogen receptor alpha-regulated lncRNA
NEAT1 is a critical modulator of prostate cancer. Nat Commun.
5:53832014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nie FQ, Zhu Q, Xu TP, Zou YF, Xie M, Sun
M, Xia R and Lu KH: Long non-coding RNA MVIH indicates a poor
prognosis for non-small cell lung cancer and promotes cell
proliferation and invasion. Tumour Biol. 35:7587–7594. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yap KL, Li S, Muñoz-Cabello AM, Raguz S,
Zeng L, Mujtaba S, Gil J, Walsh MJ and Zhou MM: Molecular interplay
of the noncoding RNA ANRIL and methylated histone H3 lysine 27 by
polycomb CBX7 in transcriptional silencing of INK4a. Mol Cell.
38:662–674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang F, Huo XS, Yuan SX, Zhang L, Zhou WP,
Wang F and Sun SH: Repression of the long noncoding RNA-LET by
histone deacetylase 3 contributes to hypoxia-mediated metastasis.
Mol Cell. 49:1083–1096. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou M, Zhong L, Xu W, Sun Y, Zhang Z,
Zhao H, Yang L and Sun J: Discovery of potential prognostic long
non-coding RNA biomarkers for predicting the risk of tumor
recurrence of breast cancer patients. Sci Rep. 6:310382016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pan LJ, Zhong TF, Tang RX, Li P, Dang YW,
Huang SN and Chen G: Upregulation and clinicopathological
significance of long non-coding NEAT1 RNA in NSCLC tissues. Asian
Pac J Cancer Prev. 16:2851–2855. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hirose T, Virnicchi G, Tanigawa A,
Naganuma T, Li R, Kimura H, Yokoi T, Nakagawa S, Bénard M, Fox AH
and Pierron G: NEAT1 long noncoding RNA regulates transcription via
protein sequestration within subnuclear bodies. Mol Biol Cell.
25:169–183. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou M, Diao Z, Yue X, Chen Y, Zhao H,
Cheng L and Sun J: Construction and analysis of dysregulated
lncRNA-associated ceRNA network identified novel lncRNA biomarkers
for early diagnosis of human pancreatic cancer. Oncotarget.
7:56383–56394. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma MZ, Chu BF, Zhang Y, Weng MZ, Qin YY,
Gong W and Quan ZW: Long non-coding RNA CCAT1 promotes gallbladder
cancer development via negative modulation of miRNA-218-5p. Cell
Death Dis. 6:e15832015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang R, Ma Y, Yu D, Zhao J and Ma P:
miR-377 functions as a tumor suppressor in human clear cell renal
cell carcinoma by targeting ETS1. Biomed Pharmacother. 70:64–71.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen G, Lu L, Liu C, Shan L and Yuan D:
MicroRNA-377 suppresses cell proliferation and invasion by
inhibiting TIAM1 expression in hepatocellular carcinoma. PLoS One.
10:e01177142015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wen Z, Huang W, Feng Y, Cai W, Wang Y,
Wang X, Liang J, Wani M, Chen J, Zhu P, et al: MicroRNA-377
regulates mesenchymal stem cell-induced angiogenesis in ischemic
hearts by targeting VEGF. PLoS One. 9:e1046662014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bracken AP, Ciro M, Cocito A and Helin K:
E2F target genes: Unraveling the biology. Trends Biochem Sci.
29:409–417. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Takahashi Y, Rayman JB and Dynlacht BD:
Analysis of promoter binding by the E2F and pRB families in vivo:
Distinct E2F proteins mediate activation and repression. Genes Dev.
14:804–816. 2000.PubMed/NCBI
|
|
30
|
Humbert PO, Verona R, Trimarchi JM, Rogers
C, Dandapani S and Lees JA: E2f3 is critical for normal cellular
proliferation. Genes Dev. 14:690–703. 2000.PubMed/NCBI
|
|
31
|
Foster CS, Falconer A, Dodson AR, Norman
AR, Dennis N, Fletcher A, Southgate C, Dowe A, Dearnaley D, Jhavar
S, et al: Transcription factor E2F3 overexpressed in prostate
cancer independently predicts clinical outcome. Oncogene.
23:5871–5879. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sherr CJ and McCormick F: The RB and p53
pathways in cancer. Cancer Cell. 2:103–112. 2002. View Article : Google Scholar : PubMed/NCBI
|