Introduction
Head and neck cancer is an aggressive malignancy
comprising approximately 6% of all newly diagnosed cancers
(1). 95% of tumors arising in the
head and neck region are squamous cell carcinomas. HPV negative
head and neck squamous cell carcinoma (HNSCC) are most commonly
associated with smoking and alcohol abuse. HNSCC are known to be
infiltrated by various kinds of immune cells such as dendritic
cells (DCs), but efficient immune responses are strongly impaired
(2,3).
DCs are bone marrow derived leukocytes with an antigen-presenting
function, such as B-cells and monocytes and can be divided into
different subgroups (4–6). Human plasmacytoid DCs (pDCs) were first
identified within the T-cell areas of lymphoid organs (7,8), but also
in peripheral blood (9) and they are
the principal source of IFN-α producing cells (10). pDCs are able to recognize CpG motifs
within microbial DNA, which are unmethylated CG dinucleotides in a
certain sequence context and trigger the production of INF-α in
pDCs (11). It has been shown that
CpG DNA can be recognized by TLR9 and thus stimulates the
polyclonal activation of B lymphocytes and the secretion of
proinflammatory cytokines by pDCs and macrophages (12–14).
Cytokines in the HNSCC microenvironment also have a strong
influence on the immune response and thus play a critical role in
tumor aggressiveness and its response to chemo- and radiation
therapy (15,16). The leading cytokines identified in the
HNSCC microenvironment are interleukin (IL)-4, IL-6, IL-8 and
IL-10, granulocyte macrophage-colony-stimulating factor (GM-CSF),
vascular endothelial growth factor (VEGF), prostaglandin
E2 and basic fibroblast growth factor (bFGF) (17–19).
pDCs are considered to be primarily responsible for
the establishment of an adaptive TH1 immune response (20,21). Under
the influence of IL-3 and CD40-ligand they also can adjust the
immune response to a tolerance induction. This happens by
polarising the secretion of cytokines which lead to a TH2 specific
immune response, for example IL-4 and IL-10 (22). In this work we comprehensively
analyzed the capacity of CpG-oligonucleotides to induce IFN-α
secretion in human pDC under the influence of HNSCC.
Materials and methods
Preparation and stimulation of
pDCs
Buffy coats were provided by the blood bank of the
University of Lübeck in an anonymized manner and were used to
isolate human peripheral blood mononuclear cells (PBMCs). Blood
donors were healthy, without medication or symptomatic allergies.
PBMCs were obtained from buffy coats by Ficoll-Hypaque density
gradient centrifugation as described previously (2). pDCs were isolated by magnetically
activated cell sorting using the BDCA-4 DC isolation kit from
Miltenyi Biotec (Bergisch-Gladbach, Germany). Cell numbers were
calculated by light microscopy and cell viability was determined by
trypan blue staining of dead cells as well as using flow
cytometry.
5×104 pDCs were cultured and stimulated
in 96-well round bottom plates in 100 µl of medium, comprising
Dulbecco's modified Eagle's medium (Gibco Life Technologies,
Carlsbad, CA, USA), 10% heat-inactivated FCS (Gibco Life
Technologies), non-essential amino acids (Gibco Life Technologies),
sodium pyruvate (Sigma-Aldrich, St. Louis, MO, USA) and
X-Vivo (Cambrex Bioscience, Rockland, ME, USA). The cells
were stimulated for 12, 24 and 48 h, respectively, with the
following, in endotoxin-free water solved agents: 3 respective 6
µg/ml CpG ODN 2216 (Invivogen, Inc., San Diego, CA, USA; Metabion
International AG, Martinsried, Germany) and HNSCC supernatant
(preparation as described below) in a solution medium/supernatant
of 1:1 or in cell suspension 1:4. Human recombinant IL-10, IL-8 and
IL-6 were purchased from Biosource and added in a concentration of
1, 10 and 100 ng/ml (IL-10), resp. 100 pg/ml (IL-6), resp. 10
pg/ml, 100 pg/ml, 1 ng/ml. The IL-10 receptor antibody was
purchased by R&D Systems, Inc. (Minneapolis, MN, USA) and used
in a concentration of 2,5 µg/ml.
Preparation of HNSCC supernatants
Permanent HNSCC cell lines BHY (DSMZ, Braunschweig,
Germany (23) and PCI-13 (Dr Theresa
Whiteside, Hypopharyngeal cancer, Pittsburgh Cancer Institute,
Pittsburgh, PA, USA) were used to generate HNSCC supernatants.
HNSCC cells were cultured in DMEM-medium (Gibco Life Technologies)
supplemented with 10% FCS, 1 mM glutamine and 0.1 mM sodium
pyruvate. Cell-free supernatants were collected by centrifugation
and filtration after 48 h of cell cultivation and frozen once
(−20°C).
Detection of IFN-α
The IFN-α module set from Bender MedSystems (Vienna,
Austria) was used to detect IFN-α in cell culture supernatants
according to the instructions provided by the manufacturer. The
photometric extinction was converted into pg/ml by inverse
polynomic regression on the basis of the standard curve. The
detection threshold is stated by 3.16 pg/ml in accordance to the
manufacturer.
Statistical analysis
Data are expressed as mean with standard deviation
from three independent experiments. Statistical significance was
evaluated by paired Student's t-test.
Results
IFN-α production is reduced in
response to soluble factors of the HNSCC microenvironment
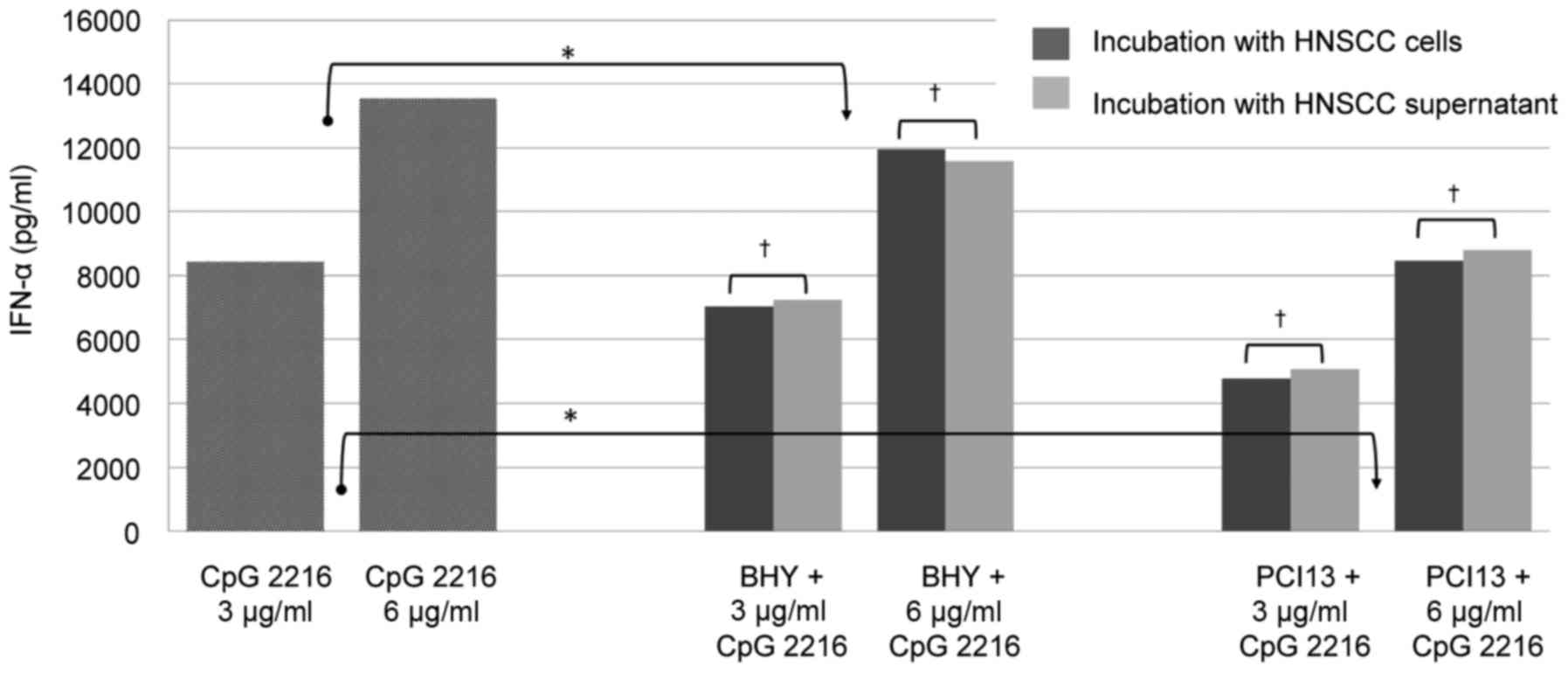
pDCs were simultaneously stimulated with 3 and 6
µg/ml CpG 2216 for 48 h in the presence and absence of HNSCC cells
and supernatant, respectively. Therefore two different permanent
HNSCC cells lines BHY and PCI13 were used. The IFN-α production was
reduced approximately 15% by BHY and 40% by PCI 13 in the mean and
was significant in both cell lines (P<0.05; Fig. 1).
Our data demonstrates that the inhibitory effect of
HNSCC does not depend on the presence of tumor cells. There is no
significant difference between the incubation with HNSCC cells or
supernatant, which underlines that the inhibition is caused by
soluble factors and does not require a direct cell contact.
Pre-incubation with HNSCC has a
negative effect on the IFN-α secretion
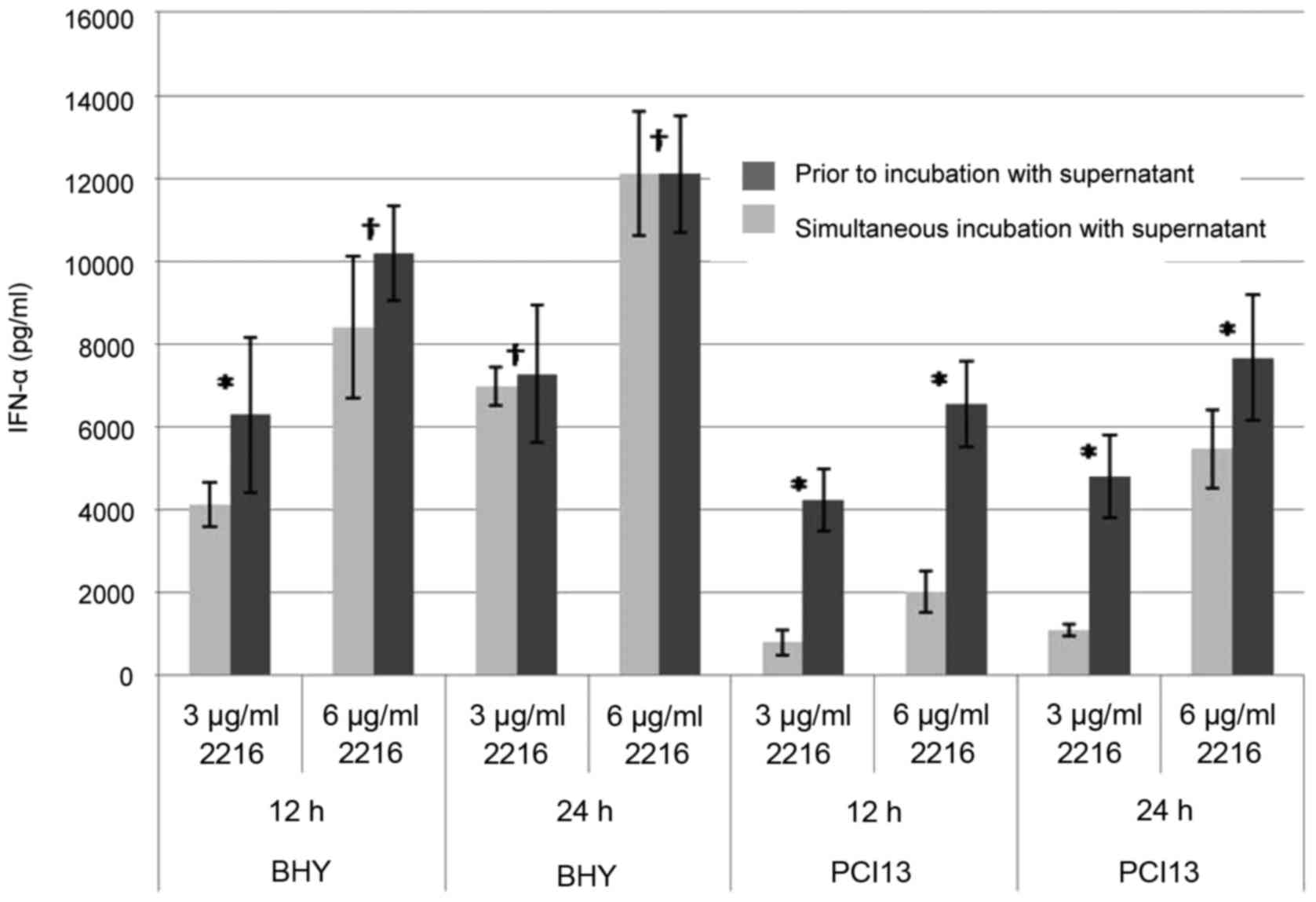
pDCs of healthy donors were pre-incubated with HNSCC
supernatants for 24 or 36 h, respectively. Subsequently CpG ODN was
added for another 12 and 24 h, so that the overall incubation time
was 48 h. As positive control cells of the same donors were
incubated with HNSCC supernatant and CpG simultaneously,
unstimulated pDCs were used as a negative control (data not
shown).
pDCs which were pre-incubated with HNSCC prior to
CpG stimulation showed a stronger impairment in the IFN-α secretion
than those cells incubated simultaneously (Fig. 2). Longer pre-incubation times led to a
stronger impaired IFN-α secretion. After 24 h of incubation with
PCI13 supernatant prior to CpG stimulation, the IFN-α is reduced by
25%; after 36 h incubation by 75% (Fig.
2B). The overall reduction by BHY was almost the same after 24
h regardless the pre-incubation period. Nevertheless, after 36 h of
incubation with supernatant, the impairment is significant
(Fig. 2A).
IL-10 is the main inhibitor of pDC
IFN-α secretion
To further elucidate which component in the
supernatant of HNSCC might be causing the impairment of pDC
function, we exposed native pDC to different concentrations of
cytokines relevant in the micro milieu of HNSCC, such as IL-6, IL-8
and IL-10.
The IL-6 concentrations were defined by the average
serum levels in HNSCC patients (19,5 pg/ml) and the maximum (312
pg/ml) as well as levels in supernatants of immortalized cell lines
(up to 4,000 pg/ml). IL-8 was used in concentrations of 10 pg/ml,
100 pg/ml and 1 ng/ml. IL-10 was used in concentrations of 1, 10
and 100 ng/ml. The concentrations were also defined with respect to
our previous data and in accordance to the literature (19,24–26).
There was no significant effect on the CpG induced
IFN-α secretion in response to the addition of IL-6 and IL-8.
Neither different concentrations nor different combinations of both
cytokines were able to impair the pDC IFN-α secretion (data not
shown).
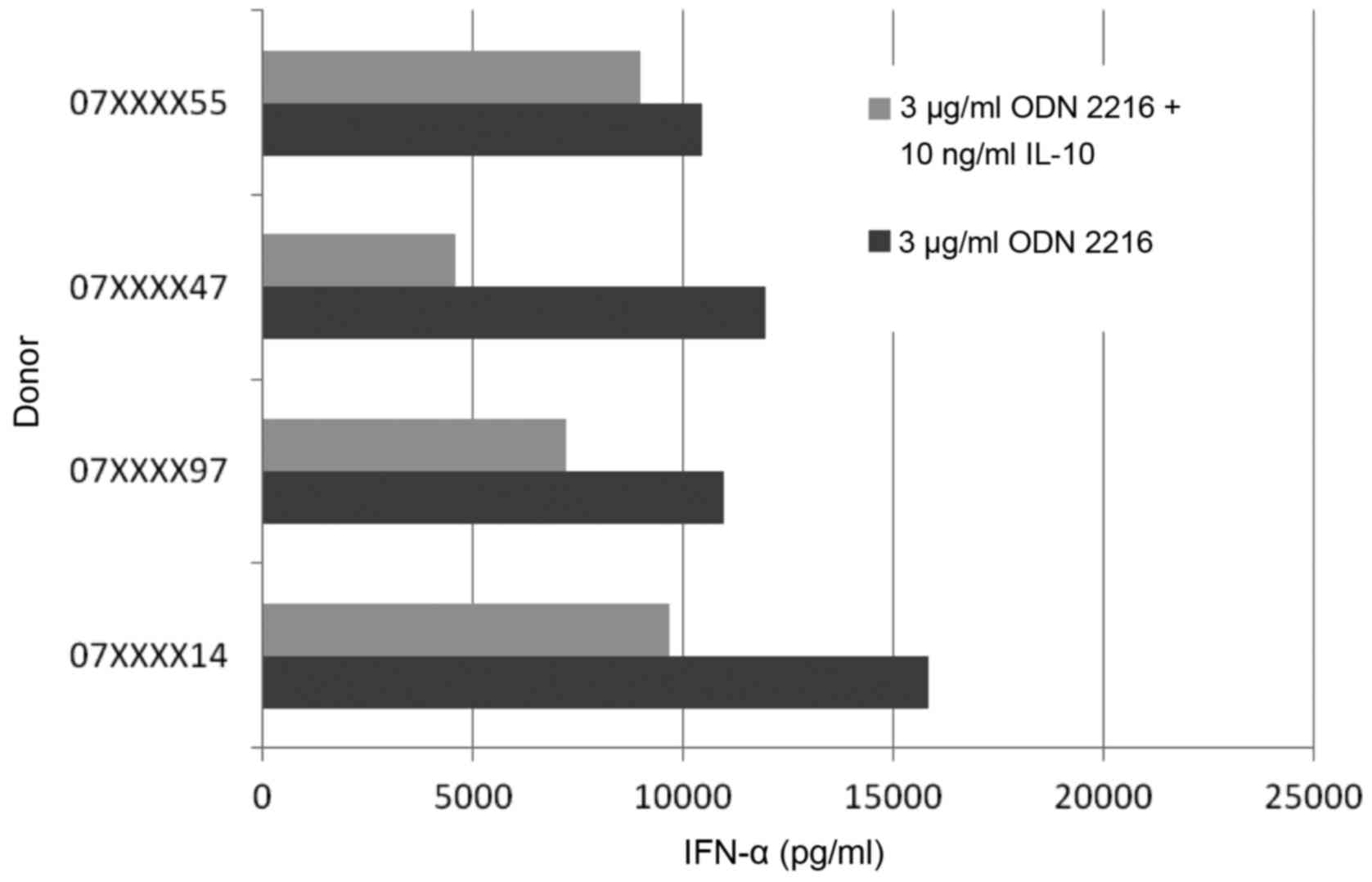
On the contrary, IL-10 showed a significant effect
on the CpG induced IFN-α secretion. These findings correlate with
data from Duramad et al (25)
in 2003. We were able to reproduce these findings in our settings.
Furthermore the CpG induced IFN-α secretion showed more or less a
wide range of variation (Fig. 3). The
four tested pDCs of healthy donors showed a fluctuation margin of
approximately 45%. The decrease of the IFN-α secretion compared to
positive control was 14, 62, 52 and 39%.
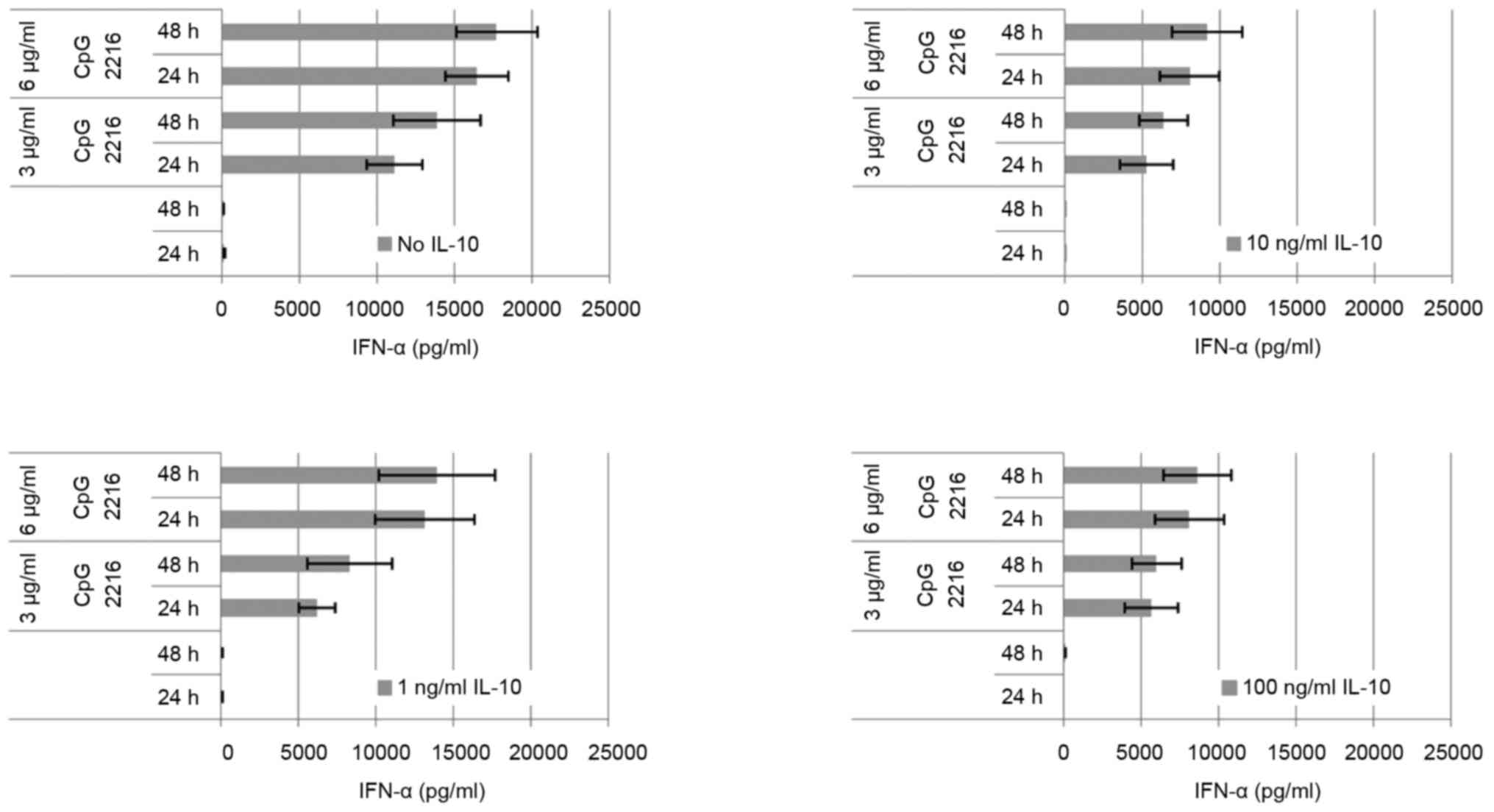
The IL-10 induced reduction on the CpG induced IFN-α
secretion in pDC is dose-dependent (Fig.
4). The maximum is reached at 10 ng/ml. At a concentration of 1
ng/ml IL-10, the suppression of the IFN-α secretion can be
outweighed by elongation of incubation and dose escalation. At a
concentration of 10 ng/ml IL-10 this is no longer the case, and the
difference between 3 and 6 µg/ml ODN 2216 is minor. The effect of 1
ng/ml IL-10 is already higher at 3 µg/ml ODN than at 6 µg/ml. The
amount of secreted cytokine on stimulation by CpG changes only
slightly at IL-10 concentration of 10 ng/ml. The dose escalation up
to 100 ng/ml has no additional significant effect on the secreted
amount of IFN-α.
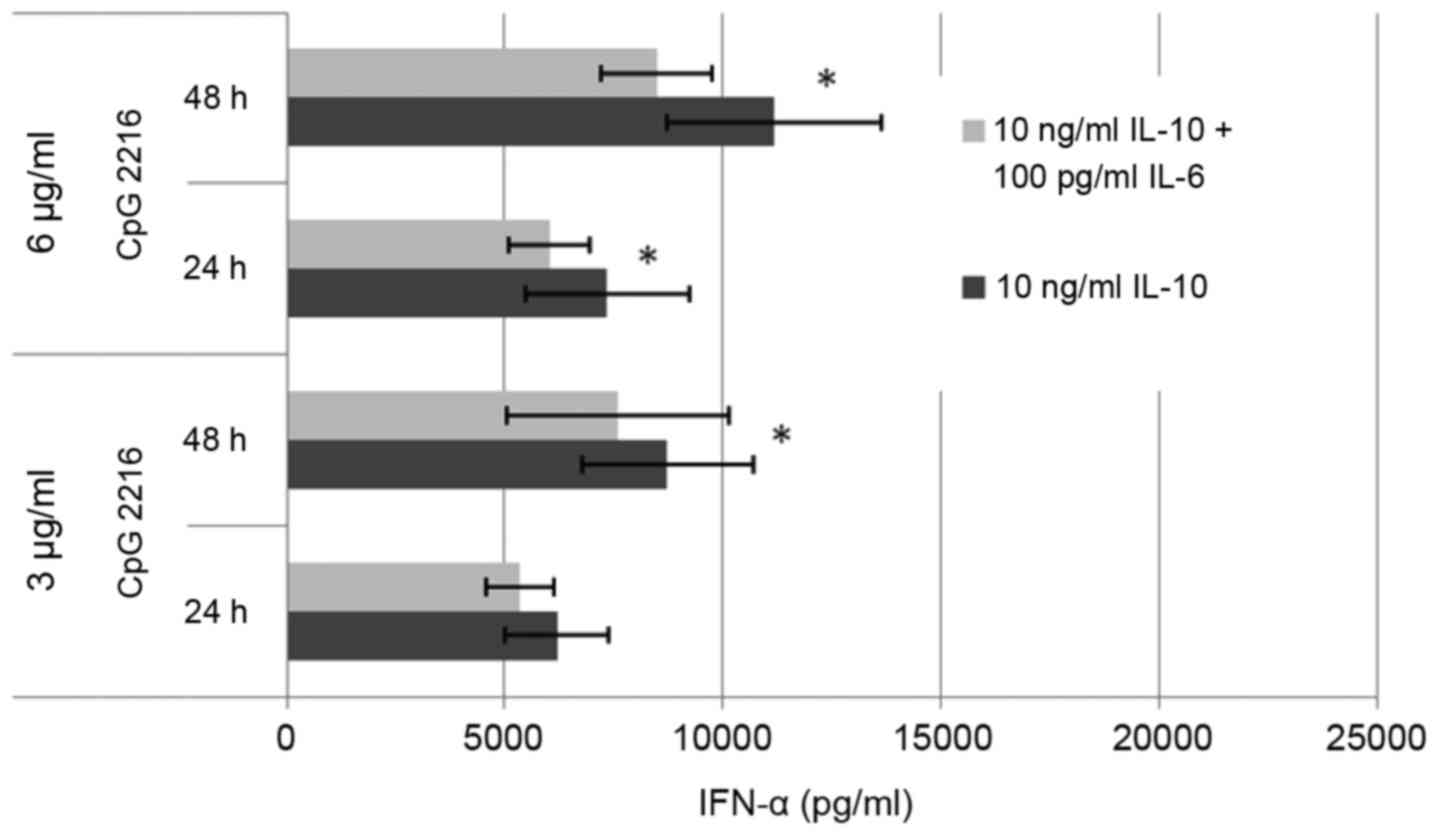
We also were able to show a synergistic effect of
IL-10 and IL-6 (Fig. 5). The
combination of IL-10 with IL-8 had no additional effect (data not
shown). The combination of IL-10 and IL-6 raised the suppressive
effect on the IFN-α secretion by 13% in average.
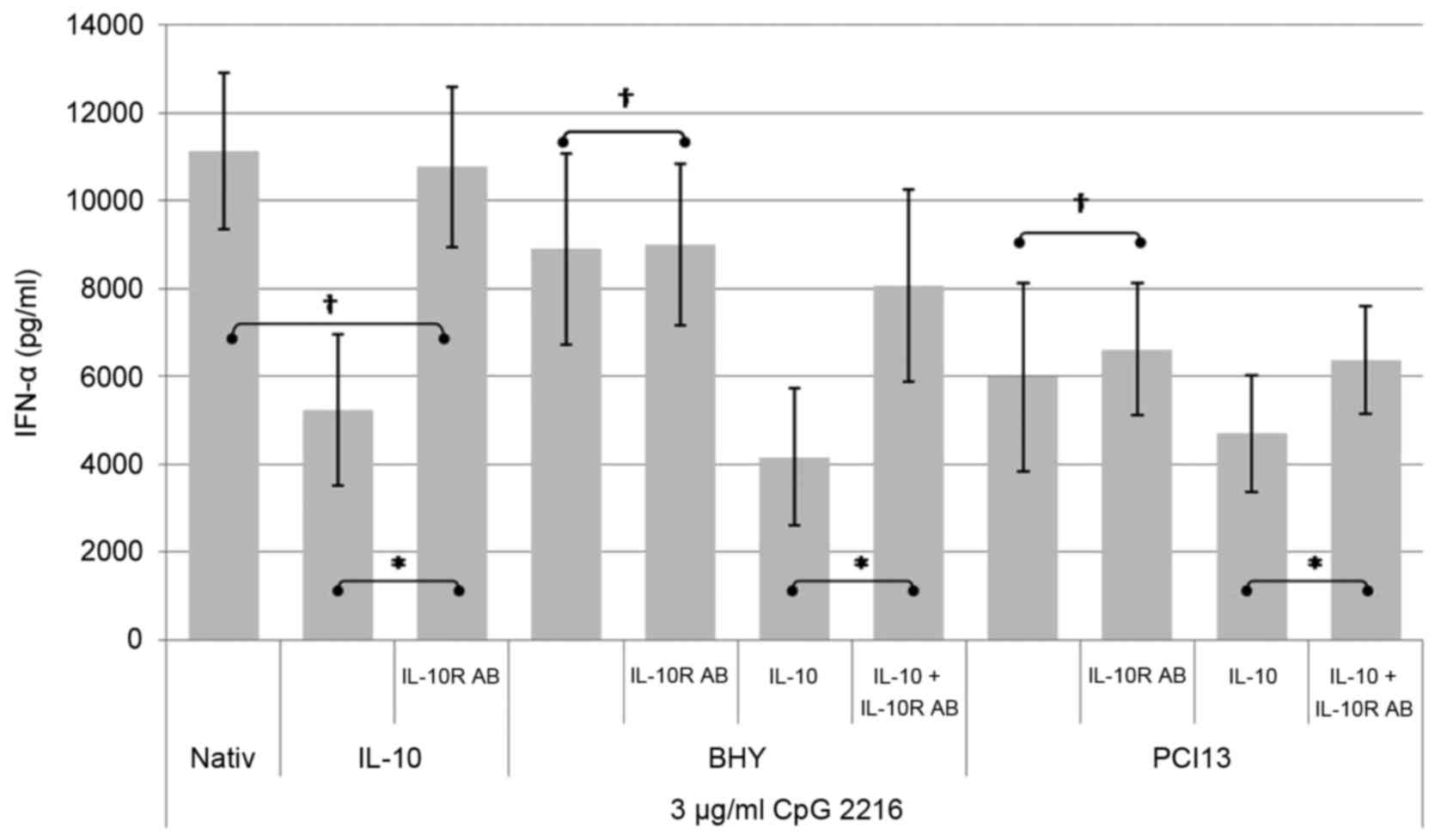
The inhibitory effect of HNSCC can be
antagonized by antibodies to the IL-10 receptor
PDCs were incubated with HNSCC supernatant and/or
IL-10 in the presence and absence of blocking antibodies to the
IL-10-receptor. The IFN-α concentration was detected by ELISA after
24 h. The negative control were cells incubated with medium and
IL-10 (data not shown). The positive control pDCs were incubated
with CpG ODN 2216.
As shown in Fig. 6,
the administration of a blocking antibody to the IL-10 receptor is
not able to neutralize the suppressive effect of the complete HNSCC
supernatant on the CpG induced IFN-α secretion completely. However
pDCs incubated under the influence of IL-10 can be fully restored
by adding the IL-10 receptor blocking antibody, which clearly
documents that various other factors play a role in the IFN alpha
suppressing mechanism.
The suppressive influence of HNSCC supernatant can
be additionally boosted by adding IL-10. Antibodies to the IL-10
receptor can diminish this additional effect but are not able to
restore the full functionality.
Discussion
The HNSCC micro milieu severely impairs the CpG ODN
induced IFN-α secretion in pDCs in vitro up to 40%. The
intensity of inhibition varies among different HNSCC cell lines
according to incubation time and CpG dosage. BHY and PCI13 show
different concentrations of soluble factors. BHY supernatant
contains high amounts of IL-6 (3,750 pg/ml) and a lower
concentration of IL-8 (820 pg/ml). The supernatant of PCI13
contains a low concentration of IL-6 (620 pg/ml) but nearly the
same amount of IL-8 (780 pg/ml) like BHY. The cytokines IL-4 and
IL-10 were measured in much lower concentrations in the supernatant
of both cell lines. The immunosuppressive IL-10 and also IL-1 are
not primarily secreted by the tumor itself. The micro milieu of the
tumor induces secretion in mDC and other cells of its environment.
IL-1 stimulates the increased secretion of the tumor stimulating
and -relevant cytokines IL-4, IL-6 and GM-CSF. The different
concentrations of the cytokines explain the varying influence of
HNSCC on the CpG induced IFN-α secretion.
With HNSCC supernatant, pre-incubated pDCs prior to
CpG stimulation show a more efficient impairment of the INF-α
secretion than those cells simultaneously incubated with CpG and
HNSCC (Fig. 2). Longer pre-incubation
times lead to a stronger impairment of IFN-α secretion than in the
case of PCI13 which can not be compensated by an extended
incubation time with CpG or a higher dosage of CpG.
Furthermore our data revealed that the inhibitory
effect of HNSCC depends on soluble factors and cell to cell contact
is not required (Fig. 1).
Our study was able to identify IL-10 as one soluble
factor in the HNSCC micro milieu that markedly reduces the IFN-α
secretion of pDCs.
The presumption that this would be key to the
explanation of the mechanism was deceptive. As shown in Fig. 6 IL-10R antibody can not abolish the
effect of HNSCC on the CpG induced IFN-α secretion entirely. The
ability of pDCs incubated with mere IL-10 can be restored almost
entirely by adding IL-10R antibody. Combined with the HNSCC
supernatant, the ability can only be retrieved by a smaller level.
This leads to the assumption that there are other factors in the
HNSCC supernatant besides IL-10 that lead to a reduction of the CpG
induced IFN-α secretion. Waibler et al had stated IL-10
before as a negative regulator regarding the GpG induced IFN-α
secretion, but we were able to show that it is not the only soluble
factor in the HNSCC micro milieu.
In literature, a cytotoxic effect of IL-10 for
CpG-activated pDCs is stated, nevertheless in our work we were not
able to reproduce this statement by FACS Annexin and PI staining
(data not shown) [Duramad et al (25)].
We were able to show a synergism of IL-6 and IL-10
(Fig. 5). Mere IL-6 has no influence
on the CpG induced IFN-α secretion but in combination with IL-10 it
alters the secretion.
It is therefore most likely that even more factors
have an influence on this mechanism.
More factors which have been described in the HNSCC
supernatant are for example the VEGF family as multifunctional
factors in angiogenesis, tumor progression, immunosuppression and
immunotolerance (27). A synergism of
IL-6, IL-1 and GM-CSF has already been described in matters of down
regulation of CD80 in tumor cells, which can be reversed by IFN-γ
(28).
The important role of IL-6 was shown in clinical
studies by Riedel et al (29)
and Duffy et al (30), and
discussed as a biomarker. The important role of IL-6 within the
Stat3 signaling in tumor proliferation has also been shown. When
Stat3 is missing in this cascade, the tumor growth is reduced
(31).
IL-8 plays a role in terms of proliferation and cell
survival in HNSCC. We were not able to show an influence on the CpG
induced IFN-α secretion in our work.
That cytokines have a strong influence of the
pathogenesis in cancer has been shown in several studies. Patients
with advanced disease show a shifted immune profile toward TH2
compared to patients with less advanced disease.
Elevations of IL-10 have been detected in diseases
like cancer and chronic infection. The strong TH1-priming ability
of CpG is the basis for future clinical trials to revert this
immune tolerating status in infectious disease, cancer, asthma and
allergic rhinitis.
Acknowledgements
We thank Brigitte Wollmann for skilful support in
some parts of this study. We are grateful to all members of the
Department of Otorhinolaryngology for helpful discussions and a
comfortable atmosphere. This study was supported by grants of the
Mildred-Scheel-Stiftung (Deutsche Krebshilfe), the Werner and Klara
Kreitz-Stiftung, the Monika-Kutzner-Stiftung and the
Rudolf-Bartling-Stiftung.
References
|
1
|
Liu Z, Yang L, Cui Y, Wang X, Guo C, Huang
Z, Kan Q, Liu Z and Liu Y: Il-21 enhances NK cell activation and
cytolytic activity and induces Th17 cell differentiation in
inflammatory bowel disease. Inflamm Bowel Dis. 15:1133–1144. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hartmann E, Wollenberg B, Rothenfusser S,
Wagner M, Wellisch D, Mack B, Giese T, Gires O, Endres S and
Hartmann G: Identification and functional analysis of
tumor-infiltrating plasmacytoid dendritic cells in head and neck
cancer. Cancer Res. 63:6478–6487. 2003.PubMed/NCBI
|
|
3
|
Reichert TE, Rabinowich H, Johnson JT and
Whiteside TL: Mechanisms responsible for signaling and functional
defects. J Immunother. 21:295–306. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Steinman RM: The dendritic cell system and
its role in immunogenicity. Annu Rev Immunol. 9:271–296. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hart DN: Dendritic cells: Unique leukocyte
populations which control the primary immune response. Blood.
90:3245–3287. 1997.PubMed/NCBI
|
|
6
|
Cella M, Sallusto F and Lanzavecchia A:
Origin, maturation and antigen presenting function of dendritic
cells. Curr Opin Immunol. 9:10–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Müller-Hermelink HK, Stein H, Steinmann G
and Lennert K: Malignant lymphoma of plasmacytoid T-cells.
Morphologic and immunologic studies characterizing a special type
of T-cell. Am J Surg Pathol. 7:849–862. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vollenweider R and Lennert K: Plasmacytoid
T-cell clusters in non-specific lymphadenitis. Virchows Arch B Cell
Pathol Incl Mol Pathol. 44:1–14. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galy A, Christopherson I, Ferlazzo G, Liu
G, Spits H and Georgopoulos K: Distinct signals control the
hematopoiesis of lymphoid-related dendritic cells. Blood.
95:128–137. 2000.PubMed/NCBI
|
|
10
|
Kirkwood J: Cancer immunotherapy: The
interferon-alpha experience. Semin Oncol. 29 3 Suppl 7:S18–S26.
2002. View Article : Google Scholar
|
|
11
|
Krug A, Rothenfusser S, Hornung V,
Jahrsdörfer B, Blackwell S, Ballas ZK, Endres S, Krieg AM and
Hartmann G: Identification of CpG oligonucleotide sequences with
high induction of IFN-alpha/beta in plasmacytoid dendritic cells.
Eur J Immunol. 31:2154–2163. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bauer S and Wagner H: Bacterial CpG-DNA
licenses TLR9. Curr Top Microbiol Immunol. 270:145–154.
2002.PubMed/NCBI
|
|
13
|
Hemmi H, Takeuchi O, Kawai T, Kaisho T,
Sato S, Sanjo H, Matsumoto M, Hoshino K, Wagner H, Takeda K and
Akira S: A Toll-like receptor recognizes bacterial DNA. Nature.
408:740–745. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Krieg AM, Yi AK, Matson S, Waldschmidt TJ,
Bishop GA, Teasdale R, Koretzky GA and Klinman DM: CpG motifs in
bacterial DNA trigger direct B-cell activation. Nature.
374:546–549. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chin D, Boyle GM, Theile DR, Parsons PG
and Coman WB: Molecular introduction to head and neck cancer
(HNSCC) carcinogenesis. Br J Plast Surg. 57:595–602. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Whiteside TL: Immune suppression in
cancer: Effects on immune cells, mechanisms and future therapeutic
intervention. Semin Cancer Biol. 16:3–15. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pries R and Wollenberg B: Cytokines in
head and neck cancer. Cytokine Growth Factor Rev. 17:141–146. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mann EA, Spiro JD, Chen LL and Kreutzer
DL: Cytokine expression by head and neck squamous cell carcinomas.
Am J Surg. 164:567–573. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen Z, Malhotra PS, Thomas GR, Ondrey FG,
Duffey DC, Smith CW, Enamorado I, Yeh NT, Kroog GS, Rudy S, et al:
Expression of proinflammatory and proangiogenic cytokines in
patients with head and neck cancer. Clin Cancer Res. 5:1369–1379.
1999.PubMed/NCBI
|
|
20
|
Liu YJ: IPC: Professional type 1
interferon-producing cells and plasmacytoid dendritic cell
precursors. Annu Rev Immunol. 23:275–306. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cella MF, Facchetti F, Lanzavecchia A and
Colonna M: Plasmacytoid dendritic cells activated by influenza
virus and CD40L drive a potent TH1 polarization. Nat Immunol.
1:305–310. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rissoan MC, Soumelis V, Kadowaki N,
Grouard G, Briere F, de Waal Malefyt R and Liu YJ: Reciprocal
control of T helper cell and dendritic cell differentiation.
Science. 283:1183–1186. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kawamata H, Nakashiro K, Uchida D, Harada
K, Yoshida H and Sato M: Possible contribution of active MMP2 to
lymph-node metastasis and secreted cathepsin L to bone invasion of
newly established human oral-squamous-cancer cell lines. Int J
Cancer. 70:120–127. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Beckebaum S, Zhang X, Chen X, Yu Z,
Frilling A, Dworacki G, Grosse-Wilde H, Broelsch CE, Gerken G and
Cicinnati VR: Increased levels of interleukin-10 in serum from
patients with hepatocellular carcinoma correlate with profound
numerical deficiencies and immature phenotype of circulating
dendritic cell subsets. Clin Cancer Res. 10:7260–7269. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Duramad O, Fearon KL, Chan JH, Kanzler H,
Marshall JD, Coffman RL and Barrat FJ: IL-10 regulates plasmacytoid
dendritic cell response to CpG-containing immunostimulatory
sequences. Blood. 102:4487–4492. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pries R, Thiel A, Brocks C and Wollenberg
B: Secretion of tumor-promoting and immune suppressive cytokines by
cell lines of head and neck squamous cell carcinoma. In Vivo.
20:45–48. 2006.PubMed/NCBI
|
|
27
|
Strauss L, Volland D, Kunkel M and
Reichert TE: Dual role of VEGF family members in the pathogenesis
of head and neck cancer (HNSCC): Possible link between angiogenesis
and immune tolerance. Med Sci Monit. 11:BR280–BR292.
2005.PubMed/NCBI
|
|
28
|
Thomas GR, Chen Z, Leukinova E, Van Waes C
and Wen J: Cytokines IL-1 alpha, IL-6, and GM-CSF constitutively
secreted by oral squamous carcinoma induce down-regulation of CD80
costimulatory molecule expression: Restoration by interferon gamma.
Cancer Immunol Immunother. 53:33–40. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Riedel F, Zaiss I, Herzog D, Götte K, Naim
R and Hörmann K: Serum levels of interleukin-6 in patients with
primary head and neck squamous cell carcinoma. Anticancer Res.
25:2761–2765. 2005.PubMed/NCBI
|
|
30
|
Duffy SA, Taylor JM, Terrell JE, Islam M,
Li Y, Fowler KE, Wolf GT and Teknos TN: Interleukin-6 predicts
recurrence and survival among head and neck cancer patients.
Cancer. 113:750–757. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bollrath J, Phesse TJ, von Burstin VA,
Putoczki T, Bennecke M, Bateman T, Nebelsiek T, Lundgren-May T,
Canli O, Schwitalla S, et al: gp130-mediated Stat3 activation in
enterocytes regulates cell survival and cell-cycle progression
during colitis-associated tumorigenesis. Cancer Cell. 15:91–102.
2009. View Article : Google Scholar : PubMed/NCBI
|