Introduction
Bladder cancer has become a global health issue, as
the second most common type of ‘tract cancer’ in the developed
countries, with over 400,000 new cases and 160,000 mortalities
worldwide per year (1). At diagnosis,
~30% of bladder cancer patients have invasive muscle tumor cells,
and 10% of patients have metastatic tumor cells with a poor
prognosis (2). Currently, surgical
resection, radiation and chemotherapy are common therapeutic
strategies used to fight against bladder cancer. However, surgery
and the current cisplatin-based combination therapeutic options are
limited by the side effects associated with treatment and by some
tumor cells eventually developing drug resistance (3). Therefore, it is imperative to develop
novel strategies to overcome bladder cancer, including therapies
that can be applied in combination with current strategies.
Mounting evidence indicates that natural compounds provide a new
window of opportunity due to their safety and potential to overcome
resistance to chemotherapy (4).
Chrysin (5,7-dihydroxyflavone) is a natural flavone
found in multiple plant extracts, such as honey and pollen, that
has a wide range of biological activities such as antibacterial,
antihypertensive, anti-allergic, anti-inflammatory, and antioxidant
effects (5–9). Chrysin is also an active inhibitor of
cancer cells. For example, chrysin can induce cell death in various
cancer types such as hepatocellular, breast, prostate and
pancreatic carcinomas (10,11). In addition, chrysin overcame
chemoresistance to cisplatin and Adriamycin in non-small cell lung
cancer (NSCLC) cells (8). These
findings make chrysin an attractive antitumor agent. However, there
is still little knowledge about the effects of chrysin on bladder
cancer cells.
In the present study, we observed a stronger
cytotoxicity for chrysin against human bladder cancer cells than
against non-malignant immortalized urothelial cells.
Mechanistically, we showed that chrysin induced apoptosis via the
intrinsic pathway. In addition, we demonstrated that chrysin could
induce ER stress and suppress the STAT3 signaling pathway, which
relies on the generation of ROS.
Materials and methods
Cell culture and chemicals
The human bladder cancer cell lines T-24 and 5637
and the non-malignant immortalized urothelial SV-HUC-1 cells were
purchased from the Shanghai Bank of Cell Culture (Shanghai, China).
Cells were cultured in RPMI 1640 medium (Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
plus 100 U/ml penicillin and 100 µg/ml streptomycin (Thermo Fisher
Scientific, Inc.) at 37°C in a humidified atmosphere of 5%
CO2. Chrysin (purity >99%) was purchased from Sigma
(St. Louis, MO, USA) and dissolved in DMSO.
3-(4,5-Dimethylthiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT),
N-acetyl cysteine (NAC) and DMSO were purchased from Sigma.
Z-LEHD-FMK, Z-DEVD-FMK and Z-IETD-FMK were purchased from Abcam
(Cambridge, CA, USA). All other chemicals were obtained from
Sigma.
MTT assay
Cells were cultured in 96-well plates at a density
of 1×104 cells/well and treated with various
concentrations of chrysin. After treatment for the indicated time,
MTT was added, followed by another 4 h incubation. The OD at 570 nm
was read using a microplate reader (Biotek, Winooski, VT, USA).
Experiments were carried out in triplicate and were repeated
separately three times.
Apoptosis assay
Apoptotic cell death was examined by the analysis of
DNA fragmentation as previously described (12). Briefly, cells were collected after
treatment. Nicoletti buffer [sodium citrate 0.1% (w/v) containing
0.1% Triton X-100 (w/v) and propidium iodide 50 ng/ml] was added to
the cell pellets and incubated for 2 h at 4°C in the dark room. The
fluorescence intensity was then measured with a FACSVerse TM flow
cytometer (Beckman Coulter, Fullerton, CA, USA) and analyzed with
the FlowJo software (Tree Star, Inc., Ashland, OR, USA). The
formula to calculate the apoptosis is as follows: (percentage of
treated apoptosis-percentage of spontaneous
apoptosis)/(100-percentage of spontaneous apoptosis) ×100.
Subcellular fraction purification
The cytosolic fraction was isolated using cytosolic
and mitochondrial fraction kits (Beyotime, Shanghai, China)
according to the manufacturer's guide.
Western blot analysis
For each sample, cells were lysed using the CHAPS
buffer provided by Cell Signaling Technology (Danvers, MA, USA).
Protein samples (20 µg) were separated on a 10% SDS-PAGE gel and
transferred onto a PVDF membrane (Millipore, Billerica, MA, USA).
PVDF membranes were then blocked with 5% skimmed milk (BYL40422; BD
Biosciences, Franklin Lakes, NJ, USA). The following primary
antibodies were used at 4°C overnight: PERK (cat. no. ab65142;
dilution, 1:1,000), EIF2α (cat. no. ab5369; dilution, 1:1,000;
Abcam), GAPDH (cat. no. SAB 2100894; dilution, 1:5,000; Sigma),
p-PERK (cat. no. 3179; dilution 1:1,000), p-EIF2α (cat. no. 3398;
dilution, 1:1,000), caspase-3 (cat. no. 9662; dilution 1:1,000),
caspase-8 (cat. no. 9746; dilution 1:1,000), caspase-9 (cat. no.
9502; dilution 1:1,000), Bcl-2 (cat. no. 4223; dilution, 1:1,000),
Mcl-1 (cat. no. 5453; dilution 1:1,000), Bcl-xl (cat. no. 2762;
dilution, 1:1,000), Bax (cat. no. 2774; dilution, 1:1,000), p-STAT3
(cat. no. 9154; dilution, 1:1,000), and STAT3 (cat. no. 4904;
dilution 1:1,000) were purchased from Cell Signaling Technology.
Then anti-rabbit (cat. no. A0545; dilution, 1:5,000) and anti-mouse
(cat. no. M5899; dilution, 1:5,000) HRP-conjucted secondary
antibodies (Sigma) were used at room temperature for 1 h. Signals
were visualized with ECL reagent (Pierce Biotechnology, Inc.,
Rockford, IL, USA). Densitometric analysis was performed with
Quantity One software (Bio-Rad Laboratories, Richmond, CA,
USA).
Caspase activity assays
The activity of caspases was measured using a
caspase activity assay kit purchased from Beyotime (Beijing, China)
according to the manufacturer's protocol. The peptides were DEVD-,
IETD- and LEHD-p-NA for the caspase-3, −8 and −9 assays,
respectively. Briefly, cell lysates were prepared after treatment
and incubated with the supplied reaction buffer and the
colorimetric substrates at 37°C for 2 h in the dark. Then, the
absorbance of the solutions was measured with a Bio Tek microplate
reader at 405 nm.
ROS detection
After treatment with chrysin, cells were collected
and stained with 15 µM DCFH-DA for 30 min, washed with PBS and then
evaluated using a FACSVerseTM flow cytometer. Data were analyzed
using FlowJo V10 (Tree Star, Inc.).
Statistical analysis
All data values were presented as the mean ± SD
(standard deviation). Statistical comparisons were measured using a
one-way analysis of variance (ANOVA) followed by Tukey's test.
Statistical analyses were carried out by using SPSS software (SPSS
Inc., Chicago, IL, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Cytotoxic effect of chrysin on bladder
cancer cells
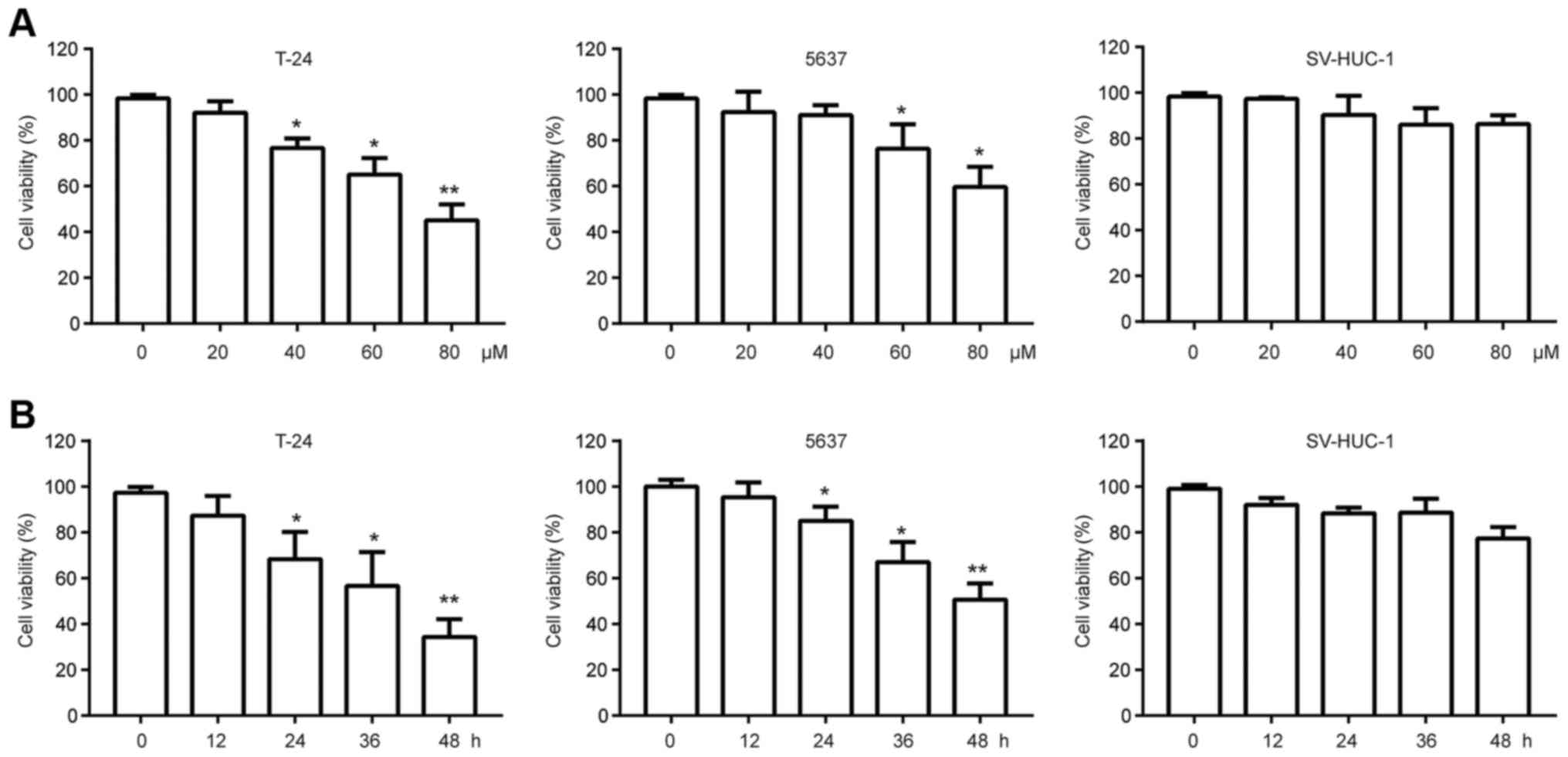
To evaluate the cytotoxic effects of chrysin on
human bladder cancer cells in vitro, we incubated two human
bladder cancer lines, T24 and 5637, with various doses (20, 40, 60,
80 µM) of chrysin for 24 h; cell viability was measured using the
MTT assay. The results showed that chrysin displayed an antitumor
effect in a concentration-dependent manner (Fig. 1A). Then, we treated T24 and 5637 cells
with 40 µM for various times (12, 24, 36 48 h). As shown in
Fig. 1B, chrysin also repressed the
viability of bladder cancer cells in a time-dependent manner. To
examine the cytotoxic effects of chrysin on normal bladder cells,
we incubated SV-HUC-1, which is an immortalized non-malignant
bladder urothelial cell line, with chrysin. Interestingly, chrysin
exerted little toxicity on SV-HUC-1 (Fig.
1A and B). Taken together, these data suggest that chrysin
exerts a selective antitumor effect on bladder tumor cells but not
on normal cells. T24 cells were selected for further analysis since
they were more sensitive to chrysin than the 5637 cells.
Chrysin selectively induces apoptosis
in human bladder cells
Then, we asked whether chrysin could induce
apoptosis in bladder cancer cells. T24 cells were treated with
various doses (20, 40, 80 µM) of chrysin for 24 h, and apoptotic
cells were measured using the apoptosis assay as described earlier
(12). As shown in Fig. 2A, we found that chrysin induced
apoptosis in a dose-dependent manner in T24 cells, while there was
little apoptosis in the non-malignant SV-HUC-1 cells. To further
investigate the apoptosis induced by chrysin, we used western
blotting to measure the protein levels of caspases, which are a
family of protease enzymes that play essential roles in apoptosis.
We found that chrysin triggers the activation of caspase-3/9 but
not caspase-8 in a dose-dependent manner in T24 cells but not in
SV-HUC-1 cells (Fig. 2B). In
addition, the activities of caspases were measured by a commercial
caspase activity assay kit. In line with the western blot results,
the activity of caspase-3/9 but not caspase-8 was significantly
enhanced by chrysin treatment in T24 cells. At the same time, the
activity of caspases could not be detected in the SV-HUC-1 cells
(Fig. 2C).
 | Figure 2.Chrysin induces apoptosis in bladder
cancer cells but not non-malignant cells. (A) Human bladder cancer
T24 and non-malignant SV-HUC-1 cells were treated with various
doses (0, 20, 40, 80 µM) of chrysin for 24 h, then cellular
apoptosis was measured by flow cytometry. (B) Human bladder cancer
cells T24 and SV-HUC-1 cells were treated with various doses (0,
20, 40, 80 µM) of chrysin for 24 h, then cellular lysates were
subjected to western blot analysis. The band densities of western
blots were quantitatively analyzed. (C) Human bladder cancer cells
T24 and SV-HUC-1 cells were treated with various doses (0, 20, 40,
80 µM) of chrysin for 24 h, then cellular lysates were subjected to
caspase activity assays. Data are presented as the mean ± SD (n=3).
*P<0.05, **P<0.01, ***P<0.001 vs. control. |
Chrysin induces apoptosis via the
intrinsic pathway in bladder cancer cells
There are two pathways leading to apoptosis, known
as the intrinsic and extrinsic pathways (13). The intrinsic pathway is initiated by
caspase-9, while the extrinsic pathway is initiated by caspase-8.
Both pathways ultimately rely on the activation of caspase-3. Our
observation that caspase-9/3 but not caspase-8 was activated after
the treatment of chrysin suggested that the induction of apoptosis
was via the intrinsic pathway in T24 cells. To further confirm
this, various specific caspase inhibitors were used. As indicated
in Fig. 3A, the specific caspase-3
inhibitor (Z-DEVD-FMK) and caspase-9 inhibitor (Z-LEHD-FMK) but not
the caspase-8 inhibitor (Z-IETD-FMK) could markedly inhibit both
the apoptosis-induced by chrysin (80 µM) and the chrysin-mediated
repression T24 cell viability (Fig.
3A). The intrinsic apoptotic pathway is known to be regulated
by the Bcl-2 proteins, which can be divided into anti-apoptotic and
pro-apoptotic members (13).
Therefore, we asked whether the levels of Bcl-2 proteins were
affected after the chrysin treatment. We found that the expressions
of the anti-apoptotic proteins Bcl-2, Mcl-1 and Bcl-xl were
repressed, while the pro-apoptotic protein Bax was upregulated, by
chrysin in a dose-dependent manner (Fig.
3B). Furthermore, we also observed an increase in cytochrome c
and Smac/DIABLO in the cytosol in a dose-dependent manner in T24
cells (Fig. 3C). These findings
suggest that chrysin may induce intrinsic apoptosis via the
modulation of Bcl-2 proteins.
 | Figure 3.Chrysin induces apoptosis via the
intrinsic pathway in bladder cancer cells. (A) Human bladder cancer
cells T24 were treated with or without chrysin (80 µM) in
combination with different caspase inhibitors (Z-LEHD-FMK,
Z-IETD-FMK, Z-DEVD-FMK) for 24 h, then cellular apoptosis and
viability were measured with flow cytometry and MTT assay,
respectively. (B) Human bladder cancer cells T24 were treated with
various doses (0, 20, 40, 80 µM) of chrysin for 24 h, and then
cellular lysates were subjected to western blot. (C) Human bladder
cancer cells T24 and SV-HUC-1 cells were treated with various doses
(0, 20, 40, 80 µM) of chrysin for 24 h, then cellular cytosolic
fractions were subjected to western blot analysis. The band
densities of western blots were quantitatively analyzed. Data are
presented as the mean ± SD (n=3). *P<0.05, **P<0.01,
***P<0.001 vs. control. |
Chrysin induces ER stress and inhibits
the STAT3 pathway in bladder cancer cells
Endoplasmic reticulum (ER) stress plays a critical
role in apoptosis induced by different natural flavones or their
derivatives (14–16). Therefore, we suspected that the
activation of ER stress involved in bladder cancer cell apoptosis
may be induced by chrysin. PERK (protein kinase RNA-like
endoplasmic reticulum kinase), a type-I ER transmembrane protein,
can be activated in response to ER stress (17). PERK activation then directly leads to
eIF2α (eukaryotic initiation factor 2α) phosphorylation and
translation inhibition (18). In
addition, ATF4 (activating transcription factor-4) induces the
transcription of various target genes during the ER stress response
(19). Therefore, PERK-eIF2α-ATF4
axis activation is a hallmark of ER stress. We measured the status
of PERK, eIF2α, and ATF4 in T24 cells after chrysin treatment. As
indicated in Fig. 4, chrysin induces
the up-regulation of ATF-4 and the phosphorylation of PERK eIF2α in
T24 cells in a dose-dependent manner.
It has been well documented that STAT proteins are
involved in the process of apoptosis by regulating the expression
of Bcl-2 proteins (20).
Constitutively active STAT3 is found in different human cancers.
Various flavones have also been found to induce apoptosis via
inhibition of STAT3 activation. To further investigate the
potential mechanism of the antitumor effect of chrysin, we examined
the levels of STAT3 protein. We found that treatment with chrysin
reduced the level of phosphorylated STAT3 but not total STAT3 in a
dose-dependent manner.
These results suggest that both ER stress and STAT3
may potentially be involved in chrysin-induced apoptosis in T24
cells.
ROS accumulation is associated with
the antitumor effect of chrysin
Many studies have indicated that ROS accumulation
induced by anti-cancer agents can trigger cell death in different
types of cancers. Therefore, we asked whether ROS generation is
associated with the antitumor effect of chrysin. We detected
intracellular ROS levels after chrysin treatment and found that ROS
generation was increased in a dose-dependent manner (Fig. 5A and B). Moreover, we treated T24
cells with chrysin (80 µM) in the presence of the ROS scavenger NAC
(20 µM) and then assessed intracellular ROS production. As shown in
Fig. 5A and B, NAC abolished
chrysin-induced ROS generation. To further characterize the role of
ROS, we examined the ER stress markers p-STAT3 and Bcl-2 by western
blot. We found that NAC could block the up-regulated expression of
p-PERK, p-EIF2α and ATF4. In addition, NAC could abrogate the
effects of chrysin on p-STAT3 and Bcl-2 family proteins. (Fig. 5C and D). Finally, NAC also abolished
the effects of chrysin on the cell viability and apoptosis
(Fig. 5E and F). Altogether, ROS
generation appears to play an essential role in the antitumor
effects of chrysin.
Discussion
Bladder cancer continues to be a global health
problem. The most common approach for treating early stage bladder
cancer is surgical resection, and chemotherapy is critical for the
eradication of tumors that cannot be removed by surgery. Although
several chemotherapeutic agents for the management of bladder
cancer have been developed in clinics, drug resistance and side
effects remain pivotal issues (3).
Natural products have been found to be valuable sources of
antitumor agents with reduced side effects. A wide variety of
natural products, such as atractylenolide I, curcumin, quercetin
and fucoidan, possess significant cytotoxic activity against
bladder cancer cells (21–24). Chrysin is a flavonoid found in many
plant extracts and possesses various bioactivities, including a
potential antitumor effect in multiple types of human cancers
(11). However, little is known about
the effects of chrysin on bladder cancer cells. In the present
study, we found that chrysin shows promising antitumor effects
against bladder cancer cells in vitro. This finding is
verified by the following evidence: i) chrysin represses the
viability of bladder cancer cells but not normal cells; ii) chrysin
induces apoptosis in bladder cancer cells; iii) chrysin induces ER
stress and inhibits STAT3 signaling in bladder cancer cells; and
iv) chrysin exerts an antitumor effect via the induction of
ROS.
In the present study, we discover that chrysin
selectively induces the suppression of bladder cancer cell growth
in a dose-dependent manner but did not reduce viability in the
normal immortalized urothelial SV-HUC-1 cells. Similarly, it has
been previously reported that non-transformed cells from different
origins, such as fibroblasts and epithelial cells, are much more
resistant to the cytotoxic effect of chrysin than malignant cells
(25,26). These findings indicate the selective
antitumor effect of chrysin in targeting malignant cells and
sparing non-cancer cells.
It is well known that apoptosis plays an essential
role in the antitumor effects of chemotherapeutic agents. The
activation of caspases, a group of cysteine proteases, is a key
intracellular event in apoptosis. In the present study, we found
that chrysin treatment promoted the activation of caspase-3 and
caspase-9 but not caspase-8 in T24 cells. Moreover, specific
inhibitors of caspase-3/9 but not of caspase-8 could block the
antitumor effect of chrysin. We also found that the anti-apoptotic
proteins Bcl-2, Bcl-xl, and Mcl-1 were repressed and that the
pro-apoptotic protein Bax was upregulated after chrysin treatment.
These findings indicate that chrysin selectively induces apoptosis
in bladder cancer cells via the intrinsic apoptotic pathway. Our
findings are similar to previous studies. For example, chrysin
induced apoptosis in human lung cancer cells by activating
caspase-3/9, decreasing Bcl-2 and increasing Bax (27). In a recent study, chrysin was also
found to induce apoptosis in hepatocellular carcinoma cells via the
intrinsic apoptotic pathway and regulation of Bcl-2 family proteins
(28). Interestingly, many other
flavonoids such as quercetin, chalcone and hesperetin exert their
antitumor effects by triggering intrinsic apoptosis (29–31).
Another important finding from our study was that
chrysin activated the ER stress pathway in bladder cancer cells, as
evidenced by increased levels of p-PERK, p-eIF2α and ATF4.
Phosphorylation of PERK is one of the early events in ER stress.
Activated PERK directly phosphorylates eIF-2α which results in the
repression of eIF-2α. On the other hand, eIF-2α increases the
translation of ATF4. These findings are similar to a recent study
in which chrysin also induced ER stress in prostate cancer cells
(32). Targeting ER stress is a
useful strategy against various tumors and our findings provide
further insight into the antitumor effects of chrysin.
STAT3 belongs to a family of cytosolic transcription
factors that are involved in cell proliferation, migration and
differentiation (33). Constitutive
activation of STAT3 has been implicated in various human cancers,
including bladder cancer (33).
Inhibition of constitutively phosphorylated STAT3 leads to growth
inhibition and apoptosis of tumor cells both in vitro and
in vivo (33). Interestingly,
chrysin has been reported to overcome resistance to TRAIL, a TNF
superfamily member that can selectively induce apoptosis in tumor
cells but not normal cells, via inhibition of STAT3 phosphorylation
(34). Therefore, modulation of STAT3
may account for the antitumor effects of chrysin, and this property
may be applied to inhibit STAT3 activation in other carcinomas as
well.
ROS are highly reactive oxygen free radicals and
trigger multiple cellular responses including ER stress and
apoptosis (35). Increasing evidence
indicates that excessive oxidative stress could be used to
eliminate tumor cells (36). Natural
chemicals with the ability to generate ROS, such as costunolide,
pachymic acid and baicalein, have demonstrated potential antitumor
effects in bladder cancer cells (37–39).
Herein, increased ROS production was also detected in the
chrysin-treated T24 cells. Importantly, abrogation of ROS
generation by NAC completely reversed the chrysin-induced ER
stress, inhibition of p-STAT3 and apoptosis, suggesting an
essential role of ROS in the antitumor effects of chrysin.
In summary, we studied the antitumor effects of
chrysin and the potential underlying mechanism of this effect. We
found that chrysin repressed cell viability and induced the
intrinsic pathway of apoptosis, the activation ER stress and the
modulation of STAT3 activity. Moreover, the antitumor effects of
chrysin rely on the generation of ROS. These results are
encouraging, although still preliminary and further investigation
into how chrysin actions is needed. Our findings indicate that
chrysin possesses great potential as a promising candidate for the
treatment of bladder cancer.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arantes-Rodrigues R, Pinto-Leite R,
Fidalgo-Goncalves L, Palmeira C, Santos L, Colaço A and Oliveira P:
Synergistic effect between cisplatin and sunitinib malate on human
urinary bladder-cancer cell lines. Biomed Res Int. 2013:7914062013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nguyen DP and Thalmann GN: Contemporary
update on neoadjuvant therapy for bladder cancer. Nat Rev Urol.
14:348–358. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pratheeshkumar P, Son YO, Korangath P,
Manu KA and Siveen KS: Phytochemicals in cancer prevention and
therapy. Biomed Res Int. 2015:3240212015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Parajuli P, Joshee N, Rimando AM, Mittal S
and Yadav AK: In vitro antitumor mechanisms of various Scutellaria
extracts and constituent flavonoids. Planta Med. 75:41–48. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gresa-Arribas N, Serratosa J, Saura J and
Sola C: Inhibition of CCAAT/enhancer binding protein delta
expression by chrysin in microglial cells results in
anti-inflammatory and neuroprotective effects. J Neurochem.
115:526–536. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Torres-Piedra M, Ortiz-Andrade R,
Villalobos-Molina R, Singh N, Medina-Franco JL, Webster SP, Binnie
M, Navarrete-Vázquez G and Estrada-Soto S: A comparative study of
flavonoid analogues on streptozotocin-nicotinamide induced diabetic
rats: Quercetin as a potential antidiabetic agent acting via
11beta-hydroxysteroid dehydrogenase type 1 inhibition. European
journal of medicinal chemistry. 45:2606–2612. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang J, Qiu J, Dong J, Li H, Luo M, Dai X,
Zhang Y, Leng B, Niu X, Zhao S and Deng X: Chrysin protects mice
from staphylococcus aureus pneumonia. J Appl Microbiol.
111:1551–1558. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Du Q, Gu X, Cai J, Huang M and Su M:
Chrysin attenuates allergic airway inflammation by modulating the
transcription factors T-bet and GATA-3 in mice. Mol Med Rep.
6:100–104. 2012.PubMed/NCBI
|
|
10
|
Huang C, Wei YX, Shen MC, Tu YH, Wang CC
and Huang HC: Chrysin, abundant in morinda citrifolia fruit
water-etoac extracts, combined with apigenin synergistically
induced apoptosis and inhibited migration in human breast and liver
cancer cells. J Agric Food Chem. 64:4235–4245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kasala ER, Bodduluru LN, Madana RMVAK,
Gogoi R and Barua CC: Chemopreventive and therapeutic potential of
chrysin in cancer: Mechanistic perspectives. Toxicol Lett.
233:214–225. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu R, Deedigan L, Albarenque SM, Mohr A
and Zwacka RM: Delivery of sTRAIL variants by MSCs in combination
with cytotoxic drug treatment leads to p53-independent enhanced
antitumor effects. Cell Death Dis. 4:e5032013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Baig S, Seevasant I, Mohamad J, Mukheem A,
Huri HZ and Kamarul T: Potential of apoptotic pathway-targeted
cancer therapeutic research: Where do we stand? Cell Death Dis.
7:e20582016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Singh MP, Han J and Kang SC:
3′,5-dihydroxy-3,4′,7-trimethoxyflavone-induces
ER-stress-associated HCT-116 programmed cell death via redox
signaling. Biomed Pharmacothe. 88:151–161. 2017. View Article : Google Scholar
|
|
15
|
Lin CC, Kuo CL, Lee MH, Lai KC, Lin JP,
Yang JS, Yu CS, Lu CC, Chiang JH, Chueh FS and Chung JG: Wogonin
triggers apoptosis in human osteosarcoma U-2 OS cells through the
endoplasmic reticulum stress, mitochondrial dysfunction and
caspase-3-dependent signaling pathways. Int J Oncol. 39:217–224.
2011.PubMed/NCBI
|
|
16
|
Bhardwaj M, Kim NH, Paul S, Jakhar R, Han
J and Kang SC: 5-hydroxy-7-methoxyflavone triggers
mitochondrial-associated cell death via reactive oxygen species
signaling in human colon carcinoma cells. PloS one.
11:e01545252016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harding HP, Zhang Y and Ron D: Protein
translation and folding are coupled by an
endoplasmic-reticulum-resident kinase. Nature. 397:271–274. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi Y, Vattem KM, Sood R, An J, Liang J,
Stramm L and Wek RC: Identification and characterization of
pancreatic eukaryotic initiation factor 2 alpha-subunit kinase,
PEK, involved in translational control. Mol Cell Biol.
18:7499–7509. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harding HP, Zhang Y, Zeng H, Novoa I, Lu
PD, Calfon M, Sadri N, Yun C, Popko B, Paules R, et al: An
integrated stress response regulates amino acid metabolism and
resistance to oxidative stress. Molecular cell. 11:619–633. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Aggarwal BB and Shishodia S: Molecular
targets of dietary agents for prevention and therapy of cancer.
Biochem Pharmacol. 71:1397–1421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu R, Yu BX, Chen JF, Lv XY, Yan ZJ, Cheng
Y and Ma Q: Anti-tumor effects of Atractylenolide I on bladder
cancer cells. J Exp Clin Cancer Res. 35:402016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong QS, Zheng LD, Lu P, Jiang FC, Chen
FM, Zeng FQ, Wang L and Dong JH: Apoptosis-inducing effects of
curcumin derivatives in human bladder cancer cells. Anticancer
Drugs. 17:279–287. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Su Q, Peng M, Zhang Y, Xu W, Darko KO, Tao
T, Huang Y, Tao X and Yang X: Quercetin induces bladder cancer
cells apoptosis by activation of AMPK signaling pathway. Am J
Cancer Res. 6:498–508. 2016.PubMed/NCBI
|
|
24
|
Park HY, Kim GY, Moon SK, Kim WJ, Yoo YH
and Choi YH: Fucoidan inhibits the proliferation of human urinary
bladder cancer T24 cells by blocking cell cycle progression and
inducing apoptosis. Molecules. 19:5981–5998. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Yang X and Morris ME: Flavonoids
are inhibitors of breast cancer resistance protein (ABCG2)-mediated
transport. Mol Pharmacol. 65:1208–1216. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cardenas M, Marder M, Blank VC and Roguin
LP: Antitumor activity of some natural flavonoids and synthetic
derivatives on various human and murine cancer cell lines. Bioorg
Med Chem. 14:2966–2971. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Samarghandian S, Nezhad MA and Mohammadi
G: Role of caspases, Bax and Bcl-2 in chrysin-induced apoptosis in
the A549 human lung adenocarcinoma epithelial cells. Anticancer
Agents Med Chem. 14:901–909. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Q, Ma S, Liu B, Liu J, Zhu R and Li
M: Chrysin induces cell apoptosis via activation of the
p53/Bcl-2/caspase-9 pathway in hepatocellular carcinoma cells. Exp
Ther Med. 12:469–474. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Srivastava S, Somasagara RR, Hegde M,
Nishana M, Tadi SK, Srivastava M, Choudhary B and Raghavan SC:
Quercetin, a natural flavonoid interacts with dna, arrests cell
cycle and causes tumor regression by activating mitochondrial
pathway of apoptosis. Sci Rep. 6:240492016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ramirez-Tagle R, Escobar CA, Romero V,
Montorfano I, Armisén R, Borgna V, Jeldes E, Pizarro L, Simon F and
Echeverria C: Chalcone-induced apoptosis through caspase-dependent
intrinsic pathways in human hepatocellular carcinoma cells. Int J
Mol Sci. 17:2602016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu D, Zhang J, Wang J, Li J, Liao F and
Dong W: Hesperetin induces apoptosis of esophageal cancer cells via
mitochondrial pathway mediated by the increased intracellular
reactive oxygen species. Tumour Biol. 37:3451–3459. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ryu S, Lim W, Bazer FW and Song G: Chrysin
induces death of prostate cancer cells by inducing ROS and ER
stress. J Cell Physiol. 232:3786–3797. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu H, Lee H, Herrmann A, Buettner R and
Jove R: Revisiting STAT3 signalling in cancer: New and unexpected
biological functions. Nat Rev Cancer. 14:736–746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lirdprapamongkol K, Sakurai H, Abdelhamed
S, Yokoyama S, Athikomkulchai S, Viriyaroj A, Awale S, Ruchirawat
S, Svasti J and Saiki I: Chrysin overcomes TRAIL resistance of
cancer cells through Mcl-1 downregulation by inhibiting STAT3
phosphorylation. Int J Oncol. 43:329–337. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moloney JN and Cotter TG: ROS signalling
in the biology of cancer. Semin Cell Dev Biol. 30383–30384.
2017.
|
|
37
|
Choi EO, Park C, Hwang HJ, Hong SH, Kim
GY, Cho EJ, Kim WJ and Choi YH: Baicalein induces apoptosis via
ROS-dependent activation of caspases in human bladder cancer 5637
cells. Int J Oncol. 49:1009–1018. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rasul A, Bao R, Malhi M, Zhao B, Tsuji I,
Li J and Li X: Induction of apoptosis by costunolide in bladder
cancer cells is mediated through ROS generation and mitochondrial
dysfunction. Molecules. 18:1418–1433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jeong JW, Lee WS, Go SI, Nagappan A, Baek
JY, Lee JD, Lee SJ, Park C, Kim GY and Kim HJ: Pachymic acid
induces apoptosis of EJ bladder cancer cells by DR5 Up-regulation,
ROS generation, modulation of Bcl-2 and IAP family members.
Phytother Res. 29:1516–1524. 2015. View
Article : Google Scholar : PubMed/NCBI
|