Introduction
Acute graft-versus-host disease (aGVHD) is a notable
complication following allogeneic hematopoietic stem cell
transplantation (allo-HSCT) (1).
Abnormally activated T cells may be the initial factor for the
occurrence of aGVHD (2). Activated T
cells promote the secretion of cytokines, interferon-γ (IFN-γ) and
interleukin-2 (IL-2), which can migrate to tissues, including in
the lungs, liver and gut, and then result in organ dysfunction
(2,3). Therefore inhibition of T cell
activation may effectively alleviate the incidence of aGVHD.
Cytotoxic T lymphocyte antigen-4 (CTLA-4) is an inhibitory receptor
following T cell activation. CTLA-4 negatively regulates T cell
activity and deficient expression of CTLA-4 leads to autoimmune
diseases (4,5). It has been suggested that the
expression of CTLA-4 is negatively associated with the severity of
aGVHD (6). CTLA-4 can induce the
phosphorylation of STAT3 and negatively modulate the proliferation
and apoptosis of CD4+ T cells, as well as the induction
of T helper (Th)1 cells (6).
However, to the best of our knowledge, the molecule that acts
upstream of CTLA-4 regulation in aGVHD has not been reported. As an
upstream regulatory molecule of CTLA-4 (7–9), T cell
immune response cDNA 7 (TIRC7), a seven transmembrane domain G
protein, was first identified to play a role in graft rejection in
1998 (10). TIRC7 is expressed in
immune organs, such as the spleen, and in T and B lymphocytes,
playing a vital role in kidney and cardiac transplantation, as well
as in collagen-induced arthritis (8,9,11,12). Our
previous study revealed that TIRC7 may be involved in the
pathogenesis of immune thrombocytopenia (ITP) and may serve as an
indicator for evaluating the efficacy of ITP treatment (13). In addition, TIRC7 plays a central
role in allograft rejection and inflammation (14). Therefore, these findings demonstrated
that TIRC7 is closely associated with the immune response. Further
studies demonstrated that TIRC7 is essential in T cell activation
(15,16), and a specific antibody against TIRC7
can prevent immune activation via selective inhibition of Th1 and
Th17 cell-associated cytokine expression of IL-2 and IFN-γ,
indicating the critical role of TIRC7 in Th1 and Th17 cells
(7,17). In our previous study, increased TIRC7
expression was observed in patients with aGVHD (18). In addition, in lymphocytes obtained
from TIRC7-deficient mice, the intracellular and cell surface
expression of CTLA-4 was found to be markedly reduced compared with
that in wild-type lymphocytes before and after activation (19). However, to the best of our knowledge,
the mechanism by which TIRC7 influences the occurrence and
development of aGVHD, and whether TIRC7 plays a regulatory role in
aGVHD via CTLA-4 remain poorly understood. It was hypothesized that
TIRC7 may positively regulate the function of CTLA-4 and inhibit T
cell activation, thus attenuating the development and progression
of aGVHD.
Although blockade of CTLA-4 signaling has previously
been evaluated in GVHD, and early blocking of CTLA-4 signaling is
expected to make T cells more aggressive and increase the severity
of GVHD, as shown by Fevery et al (5) and Li et al (20), the findings of the present study are
contradictory. Therefore, these conclusions are controversial.
In the present study, the role of TIRC7 in the
regulation of aGVHD was investigated, and the liver, lung and
intestine pathology were assessed. The results demonstrated that
compared with the control and other experimental groups, the
pathology scores of liver, lung and intestine of the aGVHD models
in the group treated with a combination of CTLA-4 and TIRC7
monoclonal antibodies (mAbs) were the lowest.
Materials and methods
Materials
TIRC7 and CTLA-4 mAbs were custom generated by Wuhan
GeneCreate Biological Engineering Co., Ltd., and the dilutions used
of the anti-TIRC and anti-CTLA-4 mAbs were 0.25 and 0.40 mg/ml
(dilution buffer, 0.01 mmol/l PBS pH 7.4), respectively.
Animals
Specific-pathogen-free (SPF) male C57BL/6 mice
(H-2Kb; age, 8–12 weeks; weight, 23–28 g; 20 mice) were
used as the donor mice and SPF female BALB/c mice
(H-2Kd; age, 8–12 weeks; weight, 24–26 g; 220 mice) were
used as the recipient mice. Recipient mice were sacrificed by
cervical dislocation at days 7, 14, 21, 28 and 35
post-transplantation, and the endpoint of the animal experiments
was day 40 post-transplantation, when the remaining mice were all
sacrificed by cervical dislocation. The animals were purchased from
Shanghai SLAC Laboratory Animal Co., Ltd., were housed in
sterilized microisolator cages and were maintained in the
individually ventilated cage room of the Experimental Animal Center
of Xuzhou Medical University (Xuzhou, China). The temperature and
relative humidity of the room were 19–21°C and ~50%, respectively.
Animals were maintained on a 12:12-h light/dark cycle. Water was
autoclaved, and feed was purchased from Shanghai SLAC Laboratory
Animal Co. Ltd. Food and water were provided ad libitum. For
the week prior to transplantation and the week following
transplantation, the mice were provided autoclaved acidified water
(pH 2.5) without any special requirements for feed. After 2 weeks
of acclimation to the laboratory conditions, the animal experiments
commenced. All the procedures involving animals performed in the
present study were in accordance with the Institutional Animal Care
and Use Committee guidelines. The experimental protocols were
approved by the Animal Ethics Committee of Xuzhou Medical
University (Xuzhou, China).
Mouse model of aGVHD
The mouse model of aGVHD was established according
to the previous articles published by our lab (21,22). The
recipient mice were exposed to a preconditioning dose of 7.5 Gy
irradiation on the day of the transplantation (0.66 Gy/min), and
within 4 h post-irradiation, different numbers of bone marrow cells
(5×106/mouse) and splenic lymphocytes
(5×105/mouse for mild-moderate aGVHD groups;
5×106/mouse for severe aGVHD groups) obtained from the
donor mice were transfused into the recipient mice. C57BL/6 mice
were sacrificed by cervical dislocation, immersed in iodine volts
for 5 min, and the tibia and femur were aseptically separated.
After removing the attached muscles and fascia, the metaphysis was
cut open. The bone marrow cavity was washed with PBS. Subsequently,
a single cell suspension was produced by filtering through a
220-mesh stainless steel strainer. The bone marrow cells were
prepared following centrifugation at 4°C and 800 × g for 5 min and
suspended in PBS buffer. The spleen of C57BL/6 mice was separated
and cut into small pieces of 5–10 mm3 and placed in a
200-mesh stainless steel filter. Subsequently, the spleen was
gently squeezed with the rubber tip of a sterile syringe core.
After washing with the mouse lymphocyte separation solution (cat.
no. DKW33-R0100; Dakewe Biotech Co., Ltd.) the cell suspension was
prepared and transferred to the centrifuge tube. An equal volume of
PBS buffer was slowly added and then the mixed liquor was
centrifuged at room temperature at 800 × g for 20 min.
Subsequently, the buffy coat layer was aspirated, and 3–4 ml PBS
buffer was added again (the centrifuge index was room temperature,
800 × g, 3 min, twice). Finally, the splenic lymphocytes were
suspended in PBS buffer. For the control groups, the BALB/c mice
were randomly divided into the following three groups (n=20 mice
per group): i) Blank control group [no total body irradiation
(TBI), with normal saline infusion); ii) bone marrow
transplantation (BMT) control group (after TBI, with bone marrow
cells infusion, not splenic lymphocytes); and iii) TBI control
group (only TBI without cell infusion). The mice in the TBI control
group were all dead before day 15 post-transplantation, while the
mice in the blank control group were all alive and there were 3
recipient mice in the BMT control group dead after transplantation
(data not shown); therefore, in the results section, only the blank
control group was presented as the control group. For experimental
groups, preconditioned BALB/c mice were randomly divided into two
groups (n=80 per group): i) Group A, a BMT mouse model with
mild-moderate aGVHD; and ii) group B, a BMT mouse model with severe
aGVHD. The degree of aGVHD in the two groups was induced using
different infusions of splenic lymphocytes, with
5×105/mouse to induce mild-moderate aGVHD and
5×106/mouse for severe aGVHD. Furthermore, according to
the different administrations of CTLA-4 and TIRC7 mAbs, the A and B
groups were randomly divided into four groups (A1-A4 and B1-B4
groups; n=20 mice per group). The CTLA-4 or TIRC7 mAbs were
intraperitoneally injected into the mice. The optimal dosing time
of CTLA-4 antibody was day 0 post-transplantation and its optimal
dose was 40 µg/mouse; the optimal dose and dosing times of TIRC7
antibody were 25 µg/mouse and days 0, 1, 2, 3, 4 and 7
post-transplantation, respectively. Details of the groups are
presented in Tables I and II.
 | Table I.Control groups of mice. |
Table I.
Control groups of mice.
| Control group (n=20
per group) | Treatment |
|---|
| Blank control
group | Mice were
transfused physiological saline via the tail vein |
| BMT control
group | Mice were
transfused 5×106 donor bone marrow cells via the tail
vein |
| TBI control
group | TBI only |
| Table II.Experimental groups of mice. |
Table II.
Experimental groups of mice.
|
|
Treatmenta |
|---|
|
|
|
|---|
| Experimental group
(n=20 per group) | Splenic
lymphocytes | CTLA-4
(µg/day) | TIRC7 (µg/day) |
|---|
| A1 group |
5×105 | 0 | 0 |
| A2 group |
5×105 | 40 | 0 |
| A3 group |
5×105 | 0 | 25 |
| A4 group |
5×105 | 40 | 25 |
| B1 group |
5×106 | 0 | 0 |
| B2 group |
5×106 | 40 | 0 |
| B3 group |
5×106 | 0 | 25 |
| B4 group |
5×106 | 40 | 25 |
Histopathological analyses
The health and behavior of the mice were monitored
daily. Recipient mice were sacrificed by cervical dislocation at
different time points [days 7, 14, 21, 28 and 35
post-transplantation according to the previous study (23)] during a total research period of 40
days post-transplantation, according to our previous study
(22). A total of 3 mice per
experimental group were sacrificed at each time point. Overall, 20
mice per group were used in order to prevent insufficient numbers
of recipient mice due to the death of mice after transplantation
for reasons other than aGVHD, and the remaining mice beyond the
research period were sacrificed by cervical dislocation. Liver,
lung and intestine tissue samples were then isolated. The tissues
were fixed with formaldehyde solution at room temperature for 24–48
h, dehydrated, embedded into paraffin at 60°C and sliced into
4-µm-thick sections using an RM2126 microtome (Leica Microsystems
GmbH). The histological sections were then stained by hematoxylin
and eosin staining at room temperature for 8–10 min and 4–5 sec,
respectively. Next, the pathological changes were evaluated using a
light microscope (Olympus Corporation; magnification, ×20), and the
number of slides examined per tissue sample was three. The images
presented in Figs. 1–6 are representative of each tissue sample
and were selected from the three slides examined per sample with
the help from a pathologist.
 | Figure 1.Pathological changes of the liver in
the control and A groups (magnification, ×20) on day 21
post-allogeneic hematopoietic stem cell transplantation. A large
number of lobulated nuclear cells and lymphocytes infiltrated the
portal vein of the liver and local necrosis occurred (red arrows);
in addition, hepatic sinusoids and central vein were dilated and
blood stasis occurred (green arrows). Control group, blank control
group; A1 group, mild-moderate aGVHD group; A2 group, mild-moderate
aGVHD group with CTLA-4 intraperitoneal injection; A3 group,
mild-moderate aGVHD group with TIRC7 intraperitoneal injection; A4
group, mild-moderate aGVHD group with CTLA-4 and TIRC7
intraperitoneal co-injection. Scale bar, 50 µm. aGVHD, acute
graft-versus-host disease; TIRC7, T cell immune response cDNA 7;
CTLA-4, cytotoxic T lymphocyte antigen-4. |
 | Figure 6.Pathological changes of the intestine
in the control and B groups (magnification, ×20) on day 14
post-allogeneic hematopoietic stem cell transplantation. Intestinal
mucosa necrosis occurred in crypts (red arrows) and most
inflammatory cells were infiltrated (green arrows). Control group,
blank control group; B1 group, severe GVHD group; B2 group, severe
GVHD group with CTLA-4 intraperitoneal injection; B3 group, severe
GVHD group with TIRC7 intraperitoneal injection; B4 group, severe
GVHD group with CTLA-4 and TIRC7 intraperitoneal co-injection.
Scale bar, 50 µm. aGVHD, acute graft-versus-host disease; TIRC7, T
cell immune response cDNA 7; CTLA-4, cytotoxic T lymphocyte
antigen-4. |
Pathology scores following
transplantation
Histopathological changes were examined with a light
microscope (Olympus Corporation; magnification, ×20) and the liver,
lung and intestine tissue samples were scored using the aGVHD
pathology score methods formulated by Blazar et al (24) and Kaplan et al (25). According to the degree of
infiltration of inflammatory cells around blood vessels in the
liver, lung and intestine, the pathology scores of each aGVHD model
were determined, with a final score of 0–8 assigned per tissue
sample. For example, intestinal GVHD was scored on the basis of
crypt apoptosis (0, rare or none; 1, occasional apoptotic bodies
per 10 crypts; 2, few apoptotic bodies per 10 crypts; 3, the
majority of crypts contain an apoptotic body; 4, the majority of
crypts contain >1 apoptotic bodies) and inflammation (0, none;
1, mild; 2, moderate; 3, severe without ulceration; 4, severe with
ulceration), and these scores were added to obtain a final score of
0–8; so if the intestine had few apoptotic bodies per 10 crypts but
severe inflammation without ulceration, the score was 5; if the
intestine had the majority of crypts containing >1 apoptotic
bodies and severe inflammation with ulceration, the score was 8
(25).
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS 16.0 software (SPSS, Inc.) was used for all statistical
analyses. Kruskal-Wallis test and Dunn's test were employed to
compare factors in different groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Pathological changes of aGVHD-targeted
organs (liver, lung and intestine)
As shown in Fig. 1,
the manifestations of hepatic aGVHD were that a large number of
lobulated nuclear cells and lymphocytes infiltrated the portal vein
of the liver and local necrosis occurred; in addition, hepatic
sinusoids and central vein were dilated and blood stasis occurred.
The A4 group appeared to have fewer pathological changes in the
liver compared with the other groups. The pathological changes of
the lung and intestine at days 21 and 14 post-transplantation,
respectively, are shown in Figs. 2
and 3. Inflammatory cells
infiltrated around the blood vessels and part of the lung
bronchiole structure was destroyed, and the intestinal mucosa
exhibited partial shedding, and necrosis and inflammatory cell
infiltration were observed. Figs.
4–6 demonstrated the
pathological changes of liver, lung and intestine, respectively, in
the blank control and B groups at different time points (days 21,
21 and 14, respectively) post-transplantation. The pathological
changes in the liver in B groups (mainly B1-B3) included severe
bile duct injury and lymphocyte infiltration (Fig. 4). Fig.
5 manifested that the lung bronchiole structure was destroyed
and the inflammatory cells were infiltrated around the blood
vessels. Fig. 6 exhibited that
intestinal mucosa necrosis occurred in crypts and that most
inflammatory cells were infiltrated.
 | Figure 2.Pathological changes of the lungs in
the control and A groups (magnification, ×20) on day 21
post-allogeneic hematopoietic stem cell transplantation.
Inflammatory cells infiltrated around the blood vessels (red
arrows) and part of the lung bronchiole structure was destroyed
(green arrows). Control group, blank control group; A1 group,
mild-moderate aGVHD group; A2 group, mild-moderate aGVHD group with
CTLA-4 intraperitoneal injection; A3 group, mild-moderate aGVHD
group with TIRC7 intraperitoneal injection; A4 group, mild-moderate
aGVHD group with CTLA-4 and TIRC7 intraperitoneal co-injection.
Scale bar, 50 µm. aGVHD, acute graft-versus-host disease; TIRC7, T
cell immune response cDNA 7; CTLA-4, cytotoxic T lymphocyte
antigen-4. |
 | Figure 3.Pathological changes of the intestine
in the control and A groups (magnification, ×20) on day 14
post-allogeneic hematopoietic stem cell transplantation. The
intestinal mucosa exhibited partial shedding (red arrows), and
necrosis and inflammatory cell infiltration were observed (green
arrows). Control group, blank control group; A1 group,
mild-moderate aGVHD group; A2 group, mild-moderate aGVHD group with
CTLA-4 intraperitoneal injection; A3 group, mild-moderate aGVHD
group with TIRC7 intraperitoneal injection; A4 group, mild-moderate
aGVHD group with CTLA-4 and TIRC7 intraperitoneal co-injection.
Scale bar, 50 µm. aGVHD, acute graft-versus-host disease; TIRC7, T
cell immune response cDNA 7; CTLA-4, cytotoxic T lymphocyte
antigen-4. |
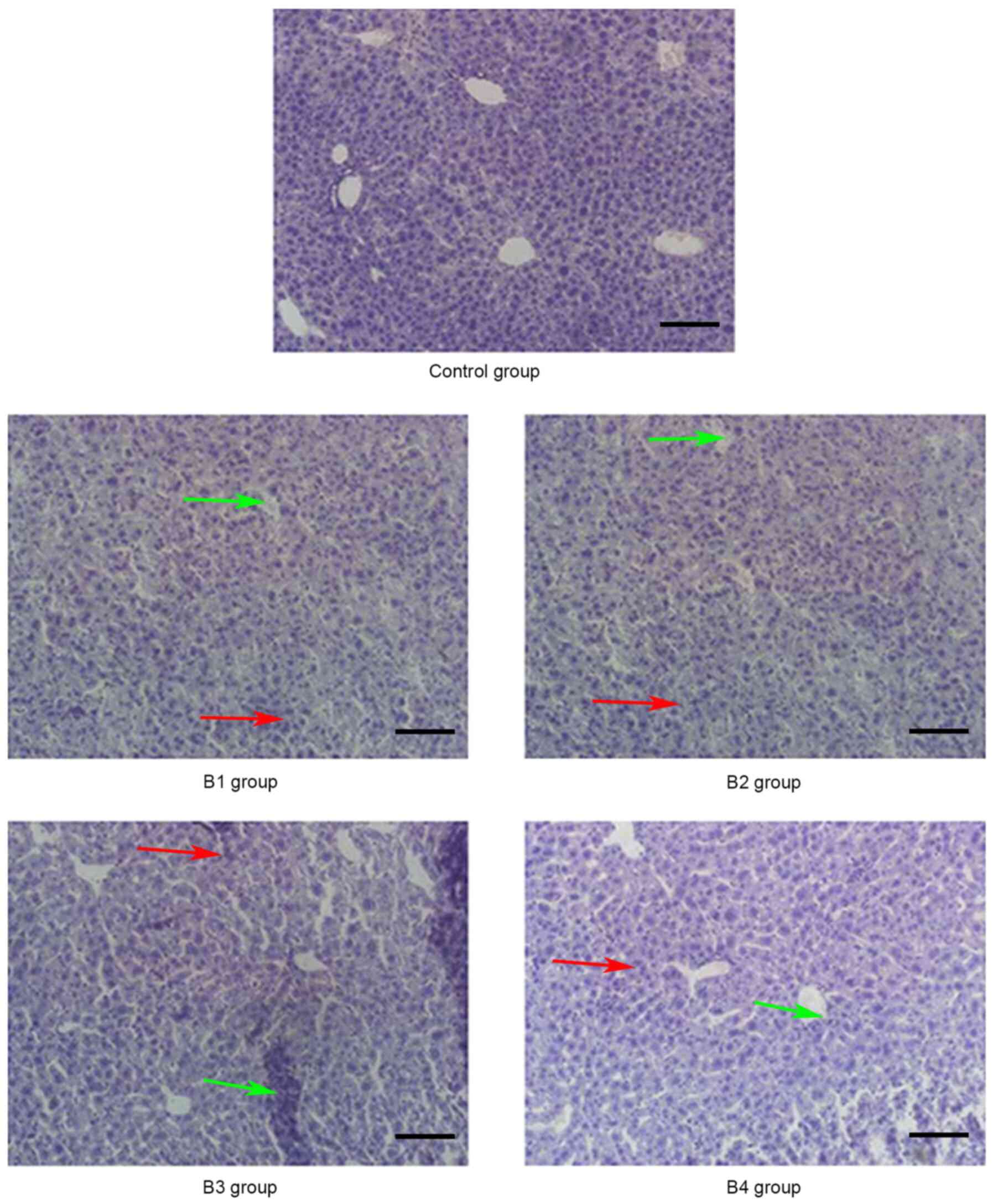 | Figure 4.Pathological changes of the liver in
the control and B groups (magnification, ×20) on day 21
post-allogeneic hematopoietic stem cell transplantation displaying
severe bile duct injury (red arrows) and lymphocyte infiltration
(green arrows). Control group, blank control group; B1 group,
severe GVHD group; B2 group, severe GVHD group with CTLA-4
intraperitoneal injection; B3 group, severe GVHD group with TIRC7
intraperitoneal injection; B4 group, severe GVHD group with CTLA-4
and TIRC7 intraperitoneal co-injection. Scale bar, 50 µm. aGVHD,
acute graft-versus-host disease; TIRC7, T cell immune response cDNA
7; CTLA-4, cytotoxic T lymphocyte antigen-4. |
 | Figure 5.Pathological changes of the lung in
the control and B groups (magnification, ×20) on day 21
post-allogeneic hematopoietic stem cell transplantation. The lung
bronchiole structure was destroyed (red arrows) and inflammatory
cells were infiltrated around the blood vessels (green arrows).
Control group, blank control group; B1 group, severe GVHD group; B2
group, severe GVHD group with CTLA-4 intraperitoneal injection; B3
group, severe GVHD group with TIRC7 intraperitoneal injection; B4
group, severe GVHD group with CTLA-4 and TIRC7 intraperitoneal
co-injection. Scale bar, 50 µm. aGVHD, acute graft-versus-host
disease; TIRC7, T cell immune response cDNA 7; CTLA-4, cytotoxic T
lymphocyte antigen-4. |
Pathological scores of aGVHD-targeted
organs (liver, lung and intestine)
As presented in Fig.
7, the pathological scores of liver in all experimental groups
gradually increased until they peaked on day 21
post-transplantation, following which they gradually declined.
Similar changes were observed in the lung and intestine, with a
peak on days 21 and 14 post-transplantation, respectively (Figs. 8 and 9). From Figs.
7–9, it was observed that at
days 14, 21, 28 and 35 post-transplantation, the pathological
scores of liver, lung and intestine post-transplantation in the A4
group were significantly lower than those of the A1, A2 and A3
groups (P<0.05), and the pathological scores of liver, lung and
intestine in the A2 and A3 groups were also significantly lower
than those of the A1 group (P<0.05). Similarly, at days 14, 21,
28 and 35 post-transplantation, the clinical scores of mice (liver,
lung and intestine) among the B groups was lowest in group B4, and
the scores in the B2 and B3 groups were also lower than those in
the B1 group (P<0.05). By comparing the experimental groups, it
was noted that the pathological scores of liver, lung and intestine
in the A4 group were not significantly different from those in the
B4 group on days 14, 21, 28 and 35 post-transplantation (data not
shown). This suggested that there was no difference in the
pathological changes between the mild-moderate and severe aGVHD
groups after combined use of TIRC7 and CTLA-4 mAbs.
Discussion
At present, to the best of our knowledge, the
pathogenesis of aGVHD remains unclear. Numerous studies have
reported that activation of T lymphocytes is the initial factor of
aGVHD (2,26). A number of membrane proteins and
their ligands have been found on the surface of T cells, which can
be divided into three categories: i) The CD28 family, including
CD28 and CTLA-4, the latter of which has homology with the CD28
molecule and binds to its ligand; ii) the costimulatory signal
molecules associated with tumor necrosis factor and its receptor;
and iii) the structural costimulatory factors associated with
signaling lymphocyte activation molecules (27). A previous study has demonstrated that
T cell activation can be enhanced by downregulating the expression
of CTLA-4 following aGVHD (28). Yoo
et al (29) found that the
apoptosis of T cells is increased and the severity of aGVHD is
decreased after overexpression of CTLA-4 in T cells, indicating
that CTLA-4 may play a negative regulatory role in aGVHD.
Meanwhile, our previous study also found that CTLA-4 levels were
decreased in patients with aGVHD before aGVHD treatment
(corticosteroids as the first-line standard treatment for II–IV
aGVHD) (18), whereas they were
increased in patients with aGVHD after treatment (6). However, TIRC7, a novel membrane
molecule, has been shown to be an essential molecule in the
regulation of lymphocyte activation, and its expression is
increased prior to aGVHD treatment (corticosteroids as the
first-line standard treatment for II–IV aGVHD) but decreased
following treatment (6,18,30).
A previous study demonstrated that TIRC7 plays an
important regulatory function in both T and B cell responses
(19). Meanwhile, as an upstream
regulatory molecule of CTLA-4, TIRC7 inhibits T cell proliferation
by modulation of CTLA-4 (16).
Anti-TIRC7 mAb enhances the upregulation of CTLA-4 expression but
suppresses the upregulation of CD25 in stimulated lymphocytes in
vitro and in vivo (7).
Antibodies against extracellular domains of TIRC7 prolonged
allograft survival in rat and mouse transplantation models, and the
prevention of rejection was mediated at least partially via
induction of CTLA-4 in T cells (31). Our group found that the expression of
CTLA-4 is negatively associated with the severity of aGVHD and
inhibits Th1 cells by increasing STAT3 expression in aGVHD
(6). Nevertheless, there is little
knowledge regarding the exact mechanism by which TIRC7 is
associated with aGVHD.
Based on these results, it was hypothesized that
TIRC7 may inhibit T cell function via CTLA-4 to reduce the severity
of aGVHD. Therefore, the present study evaluated and compared the
effect of CTLA-4 and/or TIRC7 on the development of aGVHD in mice
following HSCT and demonstrated that treatment with CTLA-4 and
TIRC7 mAbs could reduce the degree of the aGVHD.
According to the in vivo results, anti-CTLA-4
mAb administered by intraperitoneal injection could effectively
alleviate the severity of aGVHD, which was consistent with a
previous study that reported a novel mechanism able to effectively
downregulate allogeneic T cell responses through deliberate
ligation of CTLA-4 in vivo (32). Anti-TIRC7 mAb could also effectively
mitigate the severity of aGVHD, which supports the data published
by Kumamoto et al (7), which
demonstrated that targeting TIRC7 with anti-TIRC7 mAb diminishes
lymphocyte infiltration into grafts and delays morphological graft
damage. Notably, combined administration of anti-TIRC7 and
anti-CTLA-4 antibodies had an additive effect in the present study.
The optimal doses of TIRC7 and CTLA-4 mAbs to treat aGVHD in mouse
models have not yet been reported. Referring to the available
literature (7,33) and our preliminary results (data not
shown), the optimal administration time, dose and route of CTLA-4
mAb were determined to be at day 0 within 4 h after irradiation, 40
µg per mouse and intraperitoneal injection, respectively. The
optimal administration time, dose and route of TIRC7 mAb were
determined to be days 0, 1, 3, 4 and 7 post-irradiation, 25 µg per
mouse and intraperitoneal injection, respectively. It was
hypothesized that intraperitoneal injection of both TIRC7 and
CTLA-4 mAbs into recipient mice could activate the immunoreceptor
tyrosine-based inhibitory motif of the intracellular domain of two
protein molecules, negatively regulating the function of T
lymphocytes, and thereby effectively mitigating the severity of
aGVHD. In the present study, TIRC7 and CTLA-4 mAbs exhibited an
additive effect and the combined use could reduce the degree of
aGVHD more effectively compared with TIRC7 or CTLA-4 mAb alone. Due
to the focus of the current study on the aspect of pathology, the
data of the cytokine expression were not presented; however,
compared with the control group, increased levels of IFN-γ, IL-17
and IL-22, and decreased IL-4 levels were observed in groups A and
B, indicating an imbalance of Th1/17/22 and Th2 cells in the
pathogenesis of aGVHD (data not shown). The histopathological
scores of mice with aGVHD in the experimental group supported this
view. Notably, the degree of aGVHD manifestation in the
mild-moderate aGVHD group was less, for example, the aGVHD scores
of the liver, lung and intestine were lower compared with those in
the severe aGVHD group; however, there were no differences in
pathological changes between the mild-moderate and severe GVHD
groups after combined use of TIRC7 and CTLA-4 mAbs (data not
shown). The present results provide a preliminary theoretical basis
for the prevention and treatment of aGVHD in patients with
allo-HSCT by using TIRC7 and CTLA-4 mAbs in the future. TIRC7 and
CTLA-4 mAbs could reduce the severity of aGVHD in mice; however,
the current pathogenesis of whether the two mAbs have a synergistic
effect in aGVHD is not yet clear and needs to be further explored
in future studies.
In conclusion, TIRC7 may positively regulate the
function of CTLA-4, inhibit T cell activation and the secretion of
cytokines, and downregulate the progression of aGVHD. Furthermore,
combined administration of CTLA-4 and TIRC7 mAbs could reduce GVHD
more effectively in mouse models, which suggests that their
combined application may be a novel therapy for preventing and
treating GVHD.
Acknowledgements
The authors would like to thank Miss Xiangbu Wang
(Department of Pathology, The Affiliated Hospital of Xuzhou Medical
University, Xuzhou, China) for her assistance in the selection of
the pathological images of the present study.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81270637, 81300377 and
81600145), the Natural Science Foundation of Jiangsu Province
(grant nos. BK20160226 and BK20160232), the China Postdoctoral
Science Foundation (grant no. 2016M590508), the Foundation of
Jiangsu Province Six Talents Peak (grant no. 2015-wsw-058) and the
Foundation of Jiangsu Province Six-one Project (grant no.
LGY2018084).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FZhu, YX and XF designed the experiments and wrote
the first draft of the manuscript. FZha and DW primarily performed
the experiments, wrote the manuscript and prepared the figures. JQ
and SZ were involved in performing the experiments. KZ, BP and CC
made substantial contributions to the acquisition and analysis of
data. LZ, ZL and KX made contributions to the analysis and
interpretation of data and revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
All the procedures involving animals performed in
the present study were in accordance with the Institutional Animal
Care and Use Committee guidelines. The experimental protocols were
approved by the Animal Ethics Committee of Xuzhou Medical
University (Xuzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ammer J, Prantl L, Holler B, Holler K,
Landfried D, Wolff S, Karrer R, Andreesen R and Holler E:
Successful treatment of a refractory skin ulcer in chronic
cutaneous GvHD after allogeneic HSCT with split-thickness skin
allografting from the stem cell donor. Bone Marrow Transplant.
47:1368–1369. 2002. View Article : Google Scholar
|
|
2
|
Ferrara JL, Levine JE, Reddy P and Holler
E: Graft-versus-host disease. Lancet. 373:1550–1561. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lai HY, Chou TY, Tzeng CH and Lee OK:
Cytokine profiles in various graft-versus-host disease target
organs following hematopoietic stem cell transplantation. Cell
Transplant. 21:2033–2045. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tivol EA, Borriello F, Schweitzer AN,
Lynch WP, Bluestone JA and Sharpe AH: Loss of CTLA-4 leads to
massive lymphoproliferation and fatal multiorgan tissue
destruction, revealing a critical negative regulatory role of
CTLA-4. Immunity. 3:541–547. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fevery S, Billiau AD, Sprangers B,
Rutgeerts O, Lenaerts C, Goebels J, Landuyt W, Kasran A, Boon L,
Sagaert X, et al: CTLA-4 blockade in murine bone marrow chimeras
induces a host-derived antileukemic effect without
graft-versus-host disease. Leukemia. 21:1451–1459. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhu F, Zhong XM, Qiao J, Liu Q, Sun HY,
Chen W, Zhao K, Wu QY, Cao J, Sang W, et al: Cytotoxic T Lymphocyte
Antigen-4 Down-Regulates T Helper 1 cells by increasing expression
of signal transducer and activator of transcription 3 in Acute
Graft-versus-Host Disease. Biol Blood Marrow Transplant.
22:212–219. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kumamoto Y, Tomschegg A, Bennai-Sanfourche
F, Boerner A, Kaser A, Schmidt-Knosalla I, Heinemann T, Schlawinsky
M, Blumberg RS, Volk HD and Utku N: Monoclonal antibody specific
for TIRC7 induces donor-specific anergy and prevents rejection of
cardiac allografts in mice. Am J Transplant. 4:505–514. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Morgun A, Shulzhenko N, Diniz RV, Almeida
DR, Carvalho AC and Gerbase-DeLima M: Cytokine and TIRC7 mRNA
expression during acute rejection in cardiac allograft recipients.
Transplant Proc. 33:1610–1611. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kumamoto Y, Tamura A, Volk HD, Reinke P,
Lohler J, Tullius SG and Utku N: TIRC7 is induced in rejected human
kidneys and anti-TIRC7 mAb with FK506 prolongs survival of kidney
allografts in rats. Transpl Immunol. 16:238–244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Utku N, Heinemann T, Tullius SG, Bulwin
GC, Beinke S, Blumberg RS, Beato F, Randall J, Kojima R, Busconi L,
et al: Prevention of acute allograft rejection by antibody
targeting of TIRC7, a novel T cell membrane protein. Immunity.
9:509–518. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Utku N, Heinemann T, Winter M, Bulwin GC,
Schlawinsky M, Fraser P, Nieuwenhuis EE, Volk HD and Blumberg RS:
Antibody targeting of TIRC7 results in significant therapeutic
effects on collagen-induced arthritis in mice. Clin Exp Immunol.
144:142–151. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shulzhenko N, Morgun A, Rampim GF, Franco
M, Almeida DR, Diniz RV, Carvalho AC and Gerbase-DeLima M:
Monitoring of intragraft and peripheral blood TIRC7 expression as a
diagnostic tool for acute cardiac rejection in humans. Hum Immunol.
62:342–347. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu F, Qiao JL, Wu QY, Cao J, Zeng LY, Li
ZY and Xu KL: Elevated levels of T-cell immune response cDNA 7 in
patients with immune thrombocytopenia. Hematology. 19:477–482.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Utku N, Heinemann T and Milford EL: T-cell
immune response cDNA 7 in allograft rejection and inflammation.
Curr Opin Investig Drugs. 8:401–410. 2007.PubMed/NCBI
|
|
15
|
Heinemann T, Bulwin GC, Randall J,
Schnieders B, Schnieders K, Volk HD, Milford E, Gullans SR and Utku
N: Genomic organization of the gene coding for TIRC7, a novel
membrane protein essential for T cell activation. Genomics.
57:398–406. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bulwin GC, Heinemann T, Bugge V, Winter M,
Lohan A, Schlawinsky M, Schulze A, Walter S, Sabat R, Schulein R,
et al: TIRC7 inhibits T cell proliferation by modulation of CTLA-4
expression. J Immunol. 177:6833–6841. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Frischer JM, Reindl M, Kunz B, Berger T,
Schmidt S, Milford EL, Knosp E, Lassmann H and Utku N: TIRC7 and
HLA-DR axis contributes to inflammation in multiple sclerosis. Mult
Scler. 20:1171–1181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu F, Qiao J, Chen W, Pan B, Wu QY, Cao
J, Sang W, Yan ZL, Zeng LY, Li ZY and Xu KL: Increased expression
of T cell immune response cDNA 7 in patients with acute
graft-versus-host disease. Ann Hematol. 94:1025–1032. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Utku N, Boerner A, Tomschegg A,
Bennai-Sanfourche F, Bulwi GC, Heinemann T, Loehler J, Blumberg RS
and Volk HD: TIRC7 deficiency causes in vitro and in vivo
augmentation of T and B cell activation and cytokine response. J
Immunol. 173:2342–2352. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li J, Semple K, Suh WK, Liu C, Chen F,
Blazar BR and Yu XZ: Roles of CD28, CTLA4, and inducible
costimulator in acute graft-versus-host disease in mice. Biol Blood
Marrow Transplant. 17:962–969. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen W, Li M, Li Z, Yan Z, Cheng H, Pan B,
Cao J, Chen C, Zeng L and Xu K: CXCR4-transduced mesenchymal stem
cells protect mice against graft-versus-host disease. Immunol Lett.
143:161–169. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao K, Zhao D, Huang D, Song X, Chen C,
Pan B, Wu Q, Cao J, Yao Y, Zeng L and Xu K: The identification and
characteristics of IL-22-producing T cells in acute
graft-versus-host disease following allogeneic bone marrow
transplantation. Immunobiology. 218:1505–1513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu S, Shi P, Lv C, Li H, Pan B, Chen W,
Zhao K, Yan Z, Chen C, Loake GJ, et al: Loss of NLRP3 function
alleviates murine hepatic graft-versus-host disease. Biol Blood
Marrow Transplant. 24:2409–2417. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Blazar BR, Taylor PA, McElmurry R, Tian L,
Panoskaltsis-Mortari A, Lam S, Lees C, Waldschmidt T and Vallera
DA: Engraftment of severe combined immune deficient mice receiving
allogeneic bone marrow via In utero or postnatal transfer. Blood.
92:3949–3959. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kaplan DH, Anderson BE, McNiff JM, Jain D,
Shlomchik MJ and Shlomchik WD: Target antigens determine
graft-versus-host disease phenotype. J Immunol. 173:5467–5475.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qian L, Wu Z and Shen J: Advances in the
treatment of acute graft-versus-host disease. J Cell Mol Med.
17:966–975. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alegre ML, Frauwirth KA and Thompson CB:
T-cell regulation by CD28 and CTLA-4. Nat Rev Immunol. 1:220–228.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng J, Liu Y, Liu Y, Liu M, Xiang Z, Lam
KT, Lewis DB, Lau YL and Tu W: Human CD8+ regulatory T
cells inhibit GVHD and preserve general immunity in humanized mice.
Sci Transl Med. 5:168ra92013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoo JS, Lee YJ, Yoon JW, Hyunget KE and
Hwang KW: CTLA-4-Tg/CD-28-KO mice exhibit reduced t cell
proliferation in vivo compared to CD-28-KO mice in a
graft-versus-host disease model. Korean J Physiol Pharmacol.
16:349–353. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu F, Qiao J, Zhong XM, Wu QY, Chen W,
Yao Y, Niu MS, Fu CL, Zeng LY, Li ZY and Xu KL: Antithymocyte
globulin combined with cyclosporine A down-regulates T helper 1
cells by modulating T cell immune response cDNA 7 in aplastic
anemia. Med Oncol. 32:1972015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tamura A, Milford EL and Utku N: TIRC7
pathway as a target for preventing allograft rejection. Drug News
Perspect. 18:103–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hwang KW, Sweatt WB, Brown IE, Blank C,
Gajewski TF, Bluestone JA and Alegre ML: Cutting edge: Targeted
ligation of CTLA-4 in vivo by membrane-bound anti-CTLA-4 antibody
prevents rejection of allogeneic cells. J Immunol. 169:633–637.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Krummel MF, Sullivan TJ and Allison JP:
Superantigen responses and co-stimulation: CD28 and CTLA-4 have
opposing effects on T cell expansion in vitro and in vivo. Int
Immunol. 8:519–523. 1996. View Article : Google Scholar : PubMed/NCBI
|

















