Introduction
Gastric cancer (GC) is the fourth leading cause of
cancer-associated mortality worldwide, and its poor prognosis
represents a major challenge in the clinical setting, although the
incidence of GC has declined in developing countries due to the
successful reduction of H. pylori infection (1–3).
Approximately half of all patients with gastric tumors are
diagnosed at an advanced stage in western countries (4), when palliative chemotherapies may be
the only therapeutic option (5,6). In
addition, multiple treatments have not only failed to improve the
outcomes of GC but may also cause severe side effects (6). For example, combinations of two chemo
drugs are recommended to treat advanced GC but have more side
effects, including increased chance of infection, fatigue and
shortness of breath (7,8). Thus, novel drug candidates that exert
potent anti-GC effects are urgently required.
There is a well-established association between
inflammation and cancer (9). In
1863, Virchow observed a ‘lymphoreticular infiltrate’ in the tumor
site and proposed a potential association between inflammation and
cancer (10). Over the past decades,
it has become widely accepted that inflammation plays a key role in
tumorigenesis and certain underlying molecular mechanisms have been
described (10,11). For example, inflammation may induce
carcinogenesis by promoting angiogenesis and proliferation of tumor
cells or causing DNA damage (12).
Inflammation associated with tumor progression is caused by a
variety of immune cells, including T lymphocytes, B lymphocytes,
natural killer cells, dendritic cells, neutrophils and macrophages
(13). Nuclear factor (NF)-κB is one
of the key factors driving inflammation in immune cells (14). In the tumor microenvironment, NF-κB
is activated by various stimuli, including pro-inflammatory
cytokines, cellular and environmental stresses, as well as DNA
damage (15). Activated NF-κB
stimulates tumor growth and metastasis by promoting cell
proliferation and tumor angiogenesis, thus preventing cell
apoptosis and remodeling tumor metabolism (14,15).
Therefore, inhibition of NF-κB appears to be a promising approach
to cancer treatment.
Bay11-7082 is a known inhibitor of NF-κB that acts
by blocking tumor necrosis factor-α-induced IκB phosphorylation
(16). Previous studies have
demonstrated that treatment with Bay11-7082 exerts antitumor
effects on different types of cancer, including bladder, breast,
esophageal and lung cancers (17–19).
However, to the best of our knowledge, only a limited number of
studies have investigated the antitumor activity of Bay11-7082 in
GC. Thus, the present study aimed to investigate the antitumor
effects of Bay11-7082 on GC and elucidate its underlying molecular
mechanisms. Furthermore, the present study investigated whether the
effects of Bay11-7082 are mediated via inhibition of GLI Family
Zinc Finger 1 (Gli1), which acts as an oncogene in GC (20).
Materials and methods
Cell lines and cell culture
MKN45 cells were purchased from Fuheng Biotechnology
Co., Ltd. (https://www.fudancell.com), while HGC
cells were purchased from Procell Life Science & Technology
Co., Ltd. All cells were maintained in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine
serum (FBS, Tianhang Biotechnology Co., Ltd., http://zjthsw.foodmate.net), 100 µg/ml streptomycin
and 100 IU/ml penicillin (Beyotime Institute of Biotechnology), at
37°C with 5% CO2.
MTT assay
The effect of Bay11-7082 on the proliferation of
MKN45 and HGC27 cells was assessed via the MTT assay. Cells were
seeded into 96-well plates at a density of 5×104
cells/well and cultured overnight at 37°C. Following incubation,
different concentrations of Bay11-7082 (0.01, 0.10, 1.00, 10.00 and
20.00 µM) were added into each well and incubated for 24, 48 and 72
h, respectively. Subsequently, MTT solution (Beyotime Institute of
Biotechnology) was added into each well and incubated for 4 h at
37°C. Following the MTT incubation, the purple formazan crystals
were dissolved using dimethyl and cell proliferation was
subsequently analyzed at a wavelength of 450 nm.
The half-maximal inhibitory concentration
(IC50) values were calculated based on the percentage of
cell proliferation (vehicle-treated cells were considered as 100%
viable).
Wound healing assay
Following treatment with 10 nM Bay11-7082 for 72 h,
the wound healing assay was performed. Briefly, HGC27 or MKN45
cells were incubated in serum-free medium in the presence of
mitomycin (Sigma-Aldrich; Merck KGaA, 1 µg/ml) for 1 h at 37°C.
Once the cells reached 90% confluence, the monolayers were
scratched using 200 µl pipette tips, and cells were washed with
serum-free medium to remove cell debris. Subsequently, cells were
cultured for another 24 h at 37°C and cell migration was observed
under a confocal microscope (magnification, ×100; Nikon 80i, Nikon
Corporation).
Cell invasion
The cell invasion assay was performed using a
24-well Transwell chamber, which includes a membrane filter (3.0
µm) and inserts coated with 200 mg/ml Matrigel and dried overnight
at 37°C under sterile conditions. Cells (1×104) were
plated in the upper chambers of Transwell plates in serum-free
RPMI-1640 medium, while RPMI-1640 medium supplemented with 10% FBS
was plated in the lower chambers. Following incubation for 24 h at
37°C, the invasive cells were fixed with 4% polyoxymethylene at
25°C for 30 min and subsequently stained with crystal violet (0.5%
v/v in ethanol) at 25°C for 5 min. Stained cells were counted using
a confocal microscope (magnification, ×200; Nikon 80i, Nikon
Corporation).
Reverse transcription-quantitative
(RT-q) PCR
The RNA extraction kit (BioTeke Corporation) was
used to extract RNA from MKN45 and HGC27 cells, according to the
manufacturer's instructions. RT kit and RNase inhibitor were
purchased from BioTeke Corporation. The conditions for RT were as
follows: 70°C for 10 min, 25°C for 10 min, 42°C for 50 min and 80°C
for 10 min. qPCR was subsequently performed using SYBR-Green Master
Mix (Beijing Solarbio Science & Technology Co., Ltd.). The
primer sequences used for qPCR were designed by Wanlei Biotech Co.,
Ltd., (http://www.wanleibio.cn), and the melt
curves were used to analyze the accuracy. The following primer
sequences were used: Gli1 forward, 5′- TTCCTACCAGAGTCCCAAGT-3′ and
reverse, 5′-CCCTATGTGAAGCCCTATTT; p65 forward,
5′-GGGGACTACGACCTGAATG-3′ and reverse, 5′-GGGCACGATTGTCAAAGAT-3′;
and β-actin forward, 5′-GGCACCCAGCACAATGAA-3′ and reverse,
5′-TAGAAGCATTTGCGGTGG-3′. The following thermocycling conditions
were used: 94°C for 5 min, followed by 40 cycles at 94°C for 10
sec, 60°C for 20 sec and at 40°C for 1 min 30 sec. Relative
expression levels were calculated using the 2−ΔΔCq
method (21) and normalized to the
internal reference gene GAPDH.
Western blotting
Total protein was extracted from MKN45 and HGC27
cells, as previously described (22). Briefly, a cold RIPA buffer (Wanlei
Biotech Co., Ltd.) containing protease inhibitor was used to lyse
the cells. Subsequently, the extraction buffer was centrifuged at
13,000 × g for 15 min at 4°C to remove the sample debris and other
insoluble materials. Total protein was quantified via the BCA
protein assay (Biosharp Life Sciences) (23) and 40 µg protein/lane was separated by
10% SDS-PAGE. The separated proteins were subsequently transferred
onto PVDF membranes and blocked with 5% non-fat milk at room
temperature for 2 h. The membranes were incubated with primary
antibodies against NF-κB p65 (1:500; cat. no. WL01980),
phosphorylated (p)-NF-κB p65 (1:500; cat. no. WL02169) and Gli1
(1:3,000; cat. no. 66905-1-lg) overnight at 4°C (all purchased from
Wanlei Biotech Co., Ltd.). Following the primary incubation,
membranes were incubated with goat anti-rabbit secondary IgG
antibodies conjugated with horseradish peroxidase (1:5,000; cat.
no. WLA023) at 37°C for 45 min. β-actin was used as the internal
control (1:1,000, WL01845). The Biorad Gel Imaging System (Bio-Rad
Laboratories, Inc.) was used to detect the expression levels of the
target genes.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8 software (GraphPad Software, Inc.). Data are presented as
the mean ± standard deviation. Unpaired Student's t-test was used
to compare differences between the control and Bay11-7082-treated
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
Cytotoxic effects of Bay11-7082 on
HGC27 and MKN45 cells
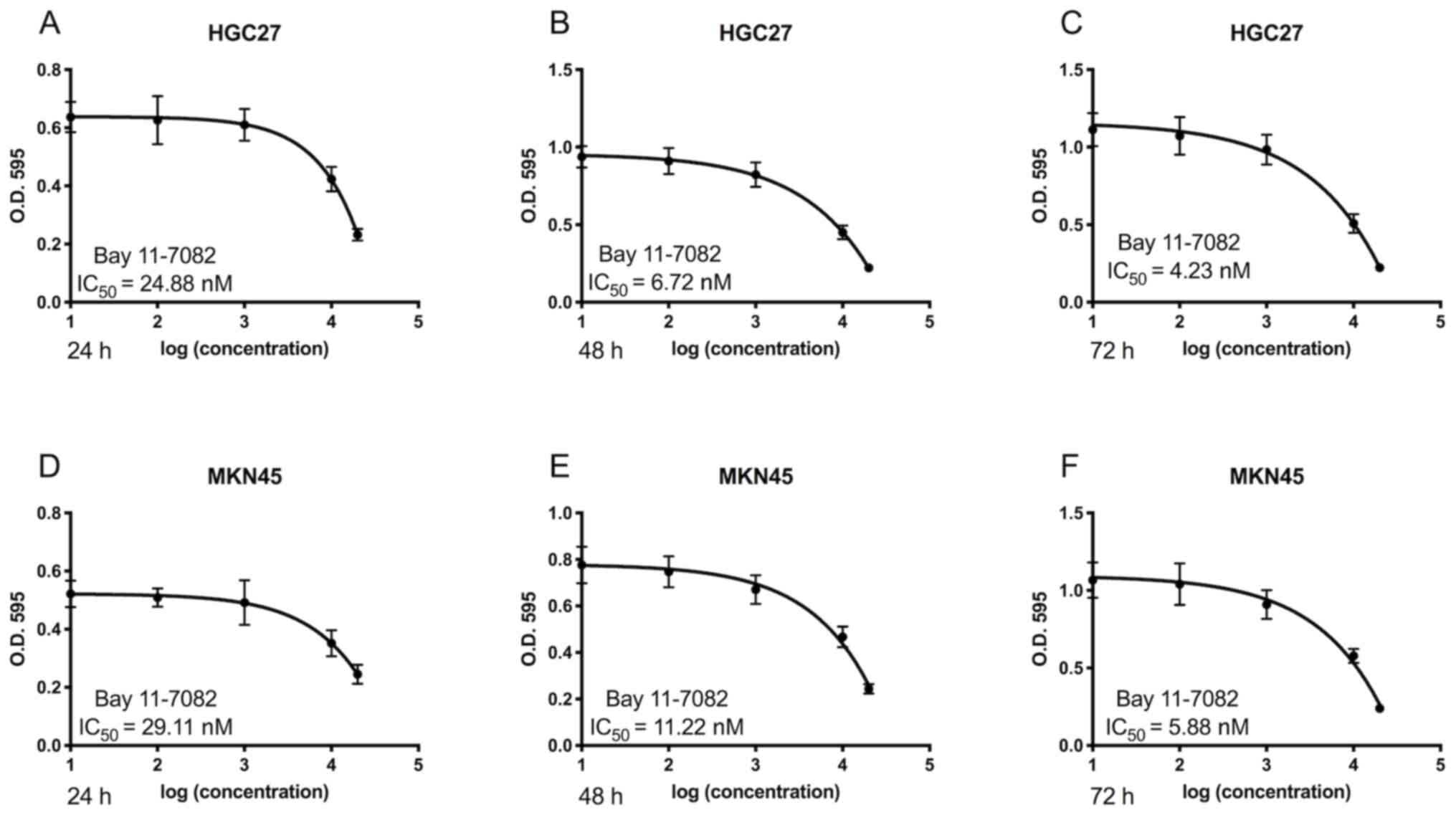
The cytotoxic effects of Bay11-7082 on HGC27 and
MKN45 cells were first determined. As presented in Fig. 1, treatment with Bay11-7082 markedly
suppressed the proliferation of HGC27 and MKN45 cells. Furthermore,
Bay11-7082 inhibited the proliferation of GC cells in dose- and
time-dependent manners. The IC50 values of Bay11-7082 in
HGC27 cells at 24, 48 and 72 h were 24.88, 6.72 and 4.23 nM,
respectively (Fig. 1A-C). The
IC50 values of Bay11-7082 in MKN45 cells at 24, 48 and
72 h were 29.11, 11.22 and 5.88 nM (Fig.
1D-F). Based on these results, 10 nM Bay11-7082 was selected
for further mechanistic studies.
Bay11-7082 suppresses the migratory
and invasive abilities of HGC27 and MKN45 cells
The effect of Bay11-7082 on the migration of HGC27
and MKN45 cells was assessed via the wound healing assay. The
results demonstrated that treatment with Bay11-7082 significantly
decreased the migratory ability of HGC27 cells compared with the
control group (P<0.01; Fig. 2A and
B). Similarly, treatment with Bay11-7082 significantly
decreased the migratory ability of MKN45 cells compared with the
control group (P<0.01; Fig. 2C and
D).
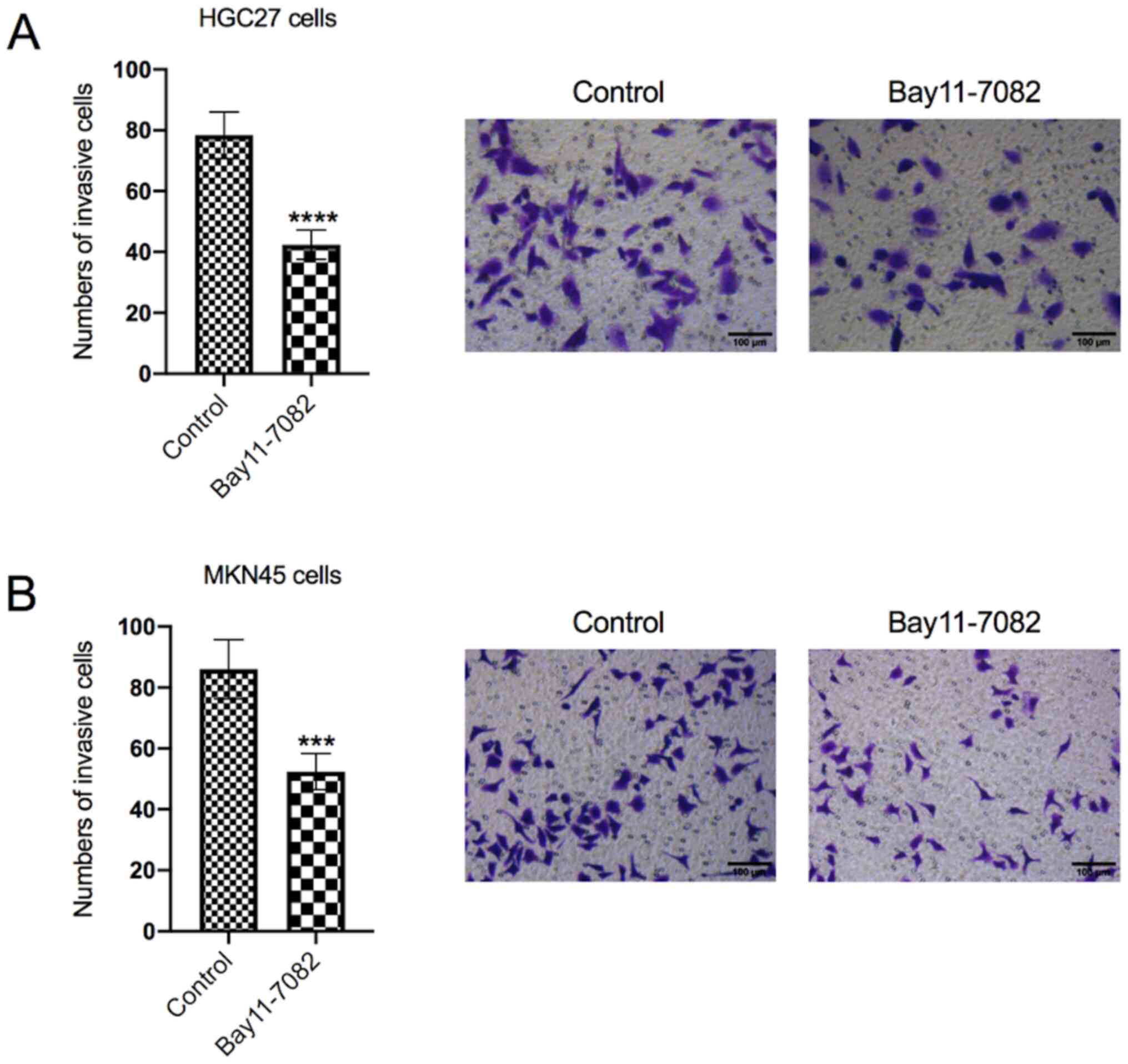
The effect of Bay11-7082 on HGC27 and
MKN45 cell invasion was also determined
As presented in Fig.
3A, treatment with Bay11-7082 significantly decreased the
number of invasive HGC27 cells compared with the control group
(P<0.0001). Similarly, treatment with Bay11-7082 significantly
decreased the numbers of invasive MKN45 cells compared with the
control group (P<0.001; Fig.
3B).
Bay11-7082 inhibits Gli1 mRNA and
protein expression levels in HGC27 and MKN45 cells
The molecular mechanisms underlying the inhibitory
effects of Bay11-7082 on HGC27 and MKN45 cells were investigated.
First, the effects of Bay11-7082 on Gli1 and p65 mRNA expression
levels were assessed. Notably, Bay11-7082 had no inhibitory effect
on p65 mRNA expression (Fig. 4A and
C). Thus, it was inferred that the inhibitory effect of
Bay11-7082 on NF-κB was exerted at the post-transcriptional level.
As expected, the results demonstrated that treatment with
Bay11-7082 decreased the protein expression levels of p-NF-κB p65
in both HGC27 and MKN45 cells (Fig.
4E).
The effect of Bay11-7082 on Gli1 mRNA
expression was investigated
Notably, the results demonstrated that treatment
with Bay11-7082 significantly inhibited Gli1 mRNA expression in
HGC27 cells compared with the control group (P<0.001; Fig. 4B). Similarly, treatment with
Bay11-7082 significantly suppressed Gli1 mRNA expression in MKN45
cells compared with the control group (P<0.0001; Fig. 4D). The effect of Bay11-7082 on Gli1
protein expression was also investigated. The results demonstrated
that treatment with Bay11-7082 significantly suppressed Gli1
protein expression compared with the control group (Fig. 4E).
Discussion
The present study aimed to investigate the effects
of Bay11-7082, a known inhibitor of NF-κB (24), on the GC cell lines, HGC27 and MKN45.
The results demonstrated that treatment with Bay11-7082
significantly inhibited the proliferation, migration and invasion
of HGC27 and MKN45 cells. Notably, in addition to its inhibitory
effects on p-NF-κB p65, Bay11-7082 also exerted inhibitory effects
on Gli1, which is a well-known glioma-associated oncogene (20).
NF-κB is activated by various stimuli, including
pro-inflammatory cytokines, cellular and environmental stressors,
and DNA damage in the tumor tissues (12). Activated NF-κB stimulates tumor
growth and metastasis through a series of processes, including: i)
Promoting cell proliferation and tumor angiogenesis, ii) preventing
cell apoptosis, and iii) remodeling the tumor metabolism (15). Thus, inhibition of NF-κB appears to
be a promising approach to cancer treatment.
Bay11-7082 act as an inhibitor of NF-κB (16). Previous studies have demonstrated
that Bay11-7082 exerts broad inhibitory effects against prostate,
esophageal, lung and colorectal cancers and lymphoma (25–29). For
example, Bay11-7082 has been reported to prevent tumor growth at
the primary site, as well as leukemic cell infiltration in various
organs of NOG mice (24). Notably,
Bay11-7082 does not exert any severe adverse effects on mice during
the treatment period (30). In
addition, Bay11-7082 has been used in the treatment of several
other experimental diseases, such as systemic lupus erythematosus,
stress-induced gastric inflammatory damage, diabetic neuropathy and
endothelin-induced lung edema (31–34).
These studies have demonstrated that apart from inhibiting NF-κB,
Bay11-7082 also exerts diverse effects on other signaling pathways.
For example, Zhang et al (17) demonstrated that the antitumor effect
of Bay11-7082 on bladder cancer is associated with its modulation
of Snail signaling pathways. In addition, Zhang et al
(18) reported that Bay11-7082 also
regulates apoptosis-related genes, including B-cell lymphoma
(Bcl)-2 and Bcl-xL, and the expression of matrix metalloproteinases
in lung cancers. However, few studies have focused on the antitumor
effects of Bay11-7082 in GC (17,24). In
addition, Bay11-7082 has not undergone clinical development.
To the best of our knowledge, the present study was
the first to investigate the effects of Bay11-7082 on two GC cell
lines, namely HGC27 and MKN45, which are commonly used in
pre-clinical studies (35,36). The HGC27 cell line was derived from a
lymph node metastasis of undifferentiated GC (37), while MKN45 is a poorly differentiated
human gastric adenocarcinoma cell line, which was found to be
moderately metastatic (38). In the
present study, parallel studies were performed on both GC cell
lines to determine the effects of Bay11-7082 in GC. The results
demonstrated that treatment with Bay11-7082 significantly inhibited
the proliferation, migration and invasion of HGC27 and MKN45 cells.
Increasing evidence suggest that there is a complex interplay
between NF-κB and Sonic hedgehog (SHH) (39). Cai et al (40) discovered that hedgehog signaling
regulates NF-κB through the classical pathway, SHH/PTCH1/SMO/Gli1,
in multiple myeloma cells. Notably, Wei et al (41) reported that NF-κB and Gli1 form a
positive feedback loop in esophageal cell lines, whereas inhibition
of either NF-κB or Gli1 inhibits cell migration, invasion and
proliferation. The results of the present study demonstrated that
Bay11-7082, in addition to inhibiting NF-κB, also regulated Gli1
expression in the HGC27 and MKN45 cells, indicating a crosstalk
between NF-κB and Gli1. These results support the hypothesis that
Bay11-7082 may be used as a novel therapeutic method for the
treatment of GC. However, in vivo studies are required to further
elucidate the antitumor activity of Bay11-7082 in GC. In addition,
further studies are required to confirm the association between
NF-κB and SHH in GC to support the clinical use of Bay11-7082 for
the treatment of GC.
Hedgehog/Gli signaling recently attracted the
attention of oncologists due to its widespread oncogenic activity
in a variety of human malignancies (42,43).
Hedgehog/Gli signaling has been reported to be associated with
cancer cell proliferation, metastasis, angiogenesis and
self-renewal, making this signaling pathway a promising treatment
target (44). In addition, NF-κB and
the hedgehog/Gli signaling pathway has been implicated in the
complex network of diverse molecular mechanisms leading to GC
(45). The activation process of the
human hedgehog pathway is initiated by the ligand of the hedgehog
pathway, SHH, which results in entry of the nuclear factor Gli
family proteins into the nucleus and initiates the regulation of
downstream target genes (46). The
Gli family includes three transcription factors, Gli1, Gli2 and
Gli3. Gli1 is the only transcriptional activator, whereas Gli2 and
Gli3 act as either positive or negative regulators (47). Thus, Gli1 expression leads to a
positive feedback loop, and it acts as a constitutive activator
(48).
In most cases, overexpression of Gli1 is considered
a symbol of hedgehog signaling pathway activation (49). Thus, the present study aimed to
investigate the effects of NF-κB inhibition on the hedgehog/Gli
signaling pathway. The effect of Bay11-7082 on hedgehog gene
expression was elucidated by assessing Gli1 mRNA and protein
expression levels in the hedgehog signaling pathway. Notably, the
results demonstrated that treatment with Bay11-7082 significantly
decreased Gli1 mRNA and protein expression levels in HGC27 and
MKN45 cells compared with the control group. Taken together, these
results suggest that inhibition of NF-κB also affects the
hedgehog/Gli signaling pathway. However, to elucidate the
association between the effect of Bay11-7082 and the hedgehog
pathway, SHH expression must also be investigated in prospective
studies.
In conclusion, the present study demonstrated that
treatment with Bay11-7082 significantly inhibited the
proliferation, migration and invasion of HGC27 and MKN45 cells.
Furthermore, the mechanistic studies revealed that Bay11-7082
exerted its anticancer effects in part by regulating NF-κB and
Gli1.
Acknowledgements
The authors of the present study would like to thank
Dr Syed Fayaz Hashmi (College of Pharmacy, University of Rhode
Island) for critically assessing the manuscript.
Funding
The present study was largely supported by the
financial support of the Science and Technology Development Project
(grant no. SYSD2020167) Funded by Science and Technology Bureau of
Suzhou City.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
YY and HQ performed most of the experiments,
analyzed the data and drafted the initial manuscript. TZ and YC
were responsible for the experimental design and drafting of the
initial manuscript. YY and HQ confirmed the authenticity of all the
raw data. All authors have read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rawla P and Barsouk A: Epidemiology of
gastric cancer: Global trends, risk factors and prevention. Prz
Gastroenterol. 14:26–38. 2019.PubMed/NCBI
|
|
2
|
Machlowska J, Baj J, Sitarz M, Maciejewski
R and Sitarz R: Gastric Cancer: Epidemiology, risk factors,
classification, genomic characteristics and treatment strategies.
Int J Mol Sci. 21:40122020. View Article : Google Scholar
|
|
3
|
Balar AV, Galsky MD, Rosenberg JE, Powles
T, Petrylak DP, Bellmunt J, Loriot Y, Necchi A, Hoffman-Censits J,
Perez-Gracia JL, et al IMvigor210 Study Group, : Atezolizumab as
first-line treatment in cisplatin-ineligible patients with locally
advanced and metastatic urothelial carcinoma: A single-arm,
multicentre, phase 2 trial. Lancet. 389:67–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Winawer SJ: Gastric cancer: Worldwide
burden and prevention opportunities. Chin J Dig Dis. 6:107–109.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ilson DH: Advances in the treatment of
gastric cancer. Curr Opin Gastroenterol. 33:473–476. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hess LM, Michael D, Mytelka DS, Beyrer J,
Liepa AM and Nicol S: Chemotherapy treatment patterns, costs, and
outcomes of patients with gastric cancer in the United States: A
retrospective analysis of electronic medical record (EMR) and
administrative claims data. Gastric Cancer. 19:607–615. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lordick F and Siewert JR: Recent advances
in multimodal treatment for gastric cancer: A review. Gastric
Cancer. 8:78–85. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zali H, Rezaei-Tavirani M and Azodi M:
Gastric cancer: Prevention, risk factors and treatment.
Gastroenterol Hepatol Bed Bench. 4:175–185. 2011.PubMed/NCBI
|
|
9
|
Liccardi G and Pentimalli F: Cancer,
immunity and inflammation. Report from the CDD Cambridge
Conferences 2018 and 2019. Cell Death Dis. 10:7982019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Diakos CI, Charles KA, McMillan DC and
Clarke SJ: Cancer-related inflammation and treatment effectiveness.
Lancet Oncol. 15:e493–e503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kawanishi S, Ohnishi S, Ma N, Hiraku Y and
Murata M: Crosstalk between DNA Damage and Inflammation in the
Multiple Steps of Carcinogenesis. Int J Mol Sci. 18:18082017.
View Article : Google Scholar
|
|
13
|
Zamarron BF and Chen W: Dual roles of
immune cells and their factors in cancer development and
progression. Int J Biol Sci. 7:651–658. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Del Prete A, Allavena P, Santoro G,
Fumarulo R, Corsi MM and Mantovani A: Molecular pathways in
cancer-related inflammation. Biochem Med (Zagreb). 21:264–275.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dolcet X, Llobet D, Pallares J and
Matias-Guiu X: NF-kB in development and progression of human
cancer. Virchows Arch. 446:475–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao J, Zhang H, Huang Y, Wang H, Wang S,
Zhao C, Liang Y and Yang N: Bay11-7082 attenuates murine lupus
nephritis via inhibiting NLRP3 inflammasome and NF-κB activation.
Int Immunopharmacol. 17:116–122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang Q, Mao Z and Sun J: NF-κB inhibitor,
BAY11-7082, suppresses M2 tumor-associated macrophage induced EMT
potential via miR-30a/NF-κB/Snail signaling in bladder cancer
cells. Gene. 710:91–97. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang L, Xie J, Gan R, Wu Z, Luo H, Chen
X, Lu Y, Wu L and Zheng D: Synergistic inhibition of lung cancer
cells by EGCG and NF-κB inhibitor BAY11-7082. J Cancer.
10:6543–6556. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Viola K, Kopf S, Huttary N, Vonach C,
Kretschy N, Teichmann M, Giessrigl B, Raab I, Stary S, Krieger S,
et al: Bay11-7082 inhibits the disintegration of the
lymphendothelial barrier triggered by MCF-7 breast cancer
spheroids; the role of ICAM-1 and adhesion. Br J Cancer.
108:564–569. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang CT, Liang Q, Yang L, Lin XL, Wu S,
Chen Y, Zhang XT, Gao YJ and Ge ZZ: RAB31 Targeted by miR-30c-2-3p
regulates the GLI1 signaling pathway, affecting gastric cancer cell
proliferation and apoptosis. Front Oncol. 8:5542018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu M, Ren L, Liang F, Zhang Y, Jiang L, Ma
W, Li C, Li X and Ye X: Effect of epiberberine from Coptis
chinensis Franch on inhibition of tumor growth in MKN-45 ×enograft
mice. Phytomedicine. 76:1532162020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang H, Wang J, Fan JH, Zhang YQ, Zhao JX,
Dai XJ, Liu Q, Shen YJ, Liu C, Sun WD, et al: Ilexgenin A exerts
anti-inflammation and anti-angiogenesis effects through inhibition
of STAT3 and PI3K pathways and exhibits synergistic effects with
Sorafenib on hepatoma growth. Toxicol Appl Pharmacol. 315:90–101.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
García MG, Alaniz L, Lopes EC, Blanco G,
Hajos SE and Alvarez E: Inhibition of NF-kappaB activity by BAY
11-7082 increases apoptosis in multidrug resistant leukemic T-cell
lines. Leuk Res. 29:1425–1434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng X, Chang RL, Cui XX, Avila G, Huang
MT, Liu Y, Kong AN, Rabson AB and Conney AH: Inhibition of
NF-kappaB by (E)3-[(4-methylphenyl)-sulfonyl]-2-propenenitrile
(BAY11-7082; BAY) is associated with enhanced
12-O-tetradecanoylphorbol-13-acetate-induced growth suppression and
apoptosis in human prostate cancer PC-3 cells. Int J Oncol.
32:257–264. 2008.PubMed/NCBI
|
|
26
|
Li B, Li YY, Tsao SW and Cheung AL:
Targeting NF-kappaB signaling pathway suppresses tumor growth,
angiogenesis, and metastasis of human esophageal cancer. Mol Cancer
Ther. 8:2635–2644. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xue W, Meylan E, Oliver TG, Feldser DM,
Winslow MM, Bronson R and Jacks T: Response and resistance to NF-κB
inhibitors in mouse models of lung adenocarcinoma. Cancer Discov.
1:236–247. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Scaife CL, Kuang J, Wills JC, Trowbridge
DB, Gray P, Manning BM, Eichwald EJ, Daynes RA and Kuwada SK:
Nuclear factor kappaB inhibitors induce adhesion-dependent colon
cancer apoptosis: Implications for metastasis. Cancer Res.
62:6870–6878. 2002.PubMed/NCBI
|
|
29
|
Keller SA, Hernandez-Hopkins D, Vider J,
Ponomarev V, Hyjek E, Schattner EJ and Cesarman E: NF-kappaB is
essential for the progression of KSHV- and EBV-infected lymphomas
in vivo. Blood. 107:3295–3302. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dewan MZ, Terashima K, Taruishi M,
Hasegawa H, Ito M, Tanaka Y, Mori N, Sata T, Koyanagi Y, Maeda M,
et al: Rapid tumor formation of human T-cell leukemia virus type
1-infected cell lines in novel NOD-SCID/gammac(null) mice:
Suppression by an inhibitor against NF-kappaB. J Virol.
77:5286–5294. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Miyamoto R, Ito T, Nomura S, Amakawa R,
Amuro H, Katashiba Y, Ogata M, Murakami N, Shimamoto K, Yamazaki C,
et al: Inhibitor of IkappaB kinase activity, BAY 11-7082,
interferes with interferon regulatory factor 7 nuclear
translocation and type I interferon production by plasmacytoid
dendritic cells. Arthritis Res Ther. 12:R872010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jia YT, Ma B, Wei W, Xu Y, Wang Y, Tang HT
and Xia ZF: Sustained activation of nuclear factor-kappaB by
reactive oxygen species is involved in the pathogenesis of
stress-induced gastric damage in rats. Crit Care Med. 35:1582–1591.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kumar A, Negi G and Sharma SS: Suppression
of NF-κB and NF-κB regulated oxidative stress and neuroinflammation
by BAY 11-7082 (IκB phosphorylation inhibitor) in experimental
diabetic neuropathy. Biochimie. 94:1158–1165. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Piechota A and Goraca A: Influence of
nuclear factor-κB inhibition on endothelin-1 induced lung edema and
oxidative stress in rats. J Physiol Pharmacol. 62:183–188.
2011.PubMed/NCBI
|
|
35
|
Terzioğlu G, Türksoy Ö and Bayrak ÖF:
Identification of An mtDNA Setpoint Associated with Highest Levels
of CD44 Positivity and Chemoresistance in HGC-27 and MKN-45 Gastric
Cancer Cell Lines. Cell J. 20:312–317. 2018.PubMed/NCBI
|
|
36
|
Peng Y, Liu YM, Li LC, Wang LL and Wu XL:
MicroRNA-503 inhibits gastric cancer cell growth and
epithelial-to-mesenchymal transition. Oncol Lett. 7:1233–1238.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akagi T and Kimoto T: Human cell line
(HGC-27) derived from the metastatic lymph node of gastric cancer.
Acta Med Okayama. 30:215–219. 1976.PubMed/NCBI
|
|
38
|
Busuttil RA, Liu DS, Di Costanzo N,
Schröder J, Mitchell C and Boussioutas A: An orthotopic mouse model
of gastric cancer invasion and metastasis. Sci Rep. 8:8252018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kasperczyk H, Baumann B, Debatin KM and
Fulda S: Characterization of sonic hedgehog as a novel NF-kappaB
target gene that promotes NF-kappaB-mediated apoptosis resistance
and tumor growth in vivo. FASEB J. 23:21–33. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cai K, Na W, Guo M, Xu R, Wang X, Qin Y,
Wu Y, Jiang J and Huang H: Targeting the cross-talk between the
hedgehog and NF-κB signaling pathways in multiple myeloma. Leuk
Lymphoma. 60:772–781. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wei L, Yan N, Sun L, Bao C and Li D:
Interplay between the NF κB and hedgehog signaling pathways
predicts prognosis in esophageal squamous cell carcinoma following
neoadjuvant chemoradiotherapy. Int J Mol Med. 41:2961–2967.
2018.PubMed/NCBI
|
|
42
|
Chen M, Carkner R and Buttyan R: The
hedgehog/Gli signaling paradigm in prostate cancer. Expert Rev
Endocrinol Metab. 6:453–467. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie H, Paradise BD, Ma WW and
Fernandez-Zapico ME: Recent Advances in the Clinical Targeting of
Hedgehog/GLI Signaling in Cancer. Cells. 8:3942019. View Article : Google Scholar
|
|
44
|
Wessler S, Krisch LM, Elmer DP and Aberger
F: From inflammation to gastric cancer - the importance of
Hedgehog/GLI signaling in Helicobacter pylori-induced chronic
inflammatory and neoplastic diseases. Cell Commun Signal.
15:152017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wu WK, Cho CH, Lee CW, Fan D, Wu K, Yu J
and Sung JJ: Dysregulation of cellular signaling in gastric cancer.
Cancer Lett. 295:144–153. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kasper M, Regl G, Frischauf AM and Aberger
F: GLI transcription factors: Mediators of oncogenic Hedgehog
signalling. Eur J Cancer. 42:437–445. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sasaki H, Nishizaki Y, Hui C, Nakafuku M
and Kondoh H: Regulation of Gli2 and Gli3 activities by an
amino-terminal repression domain: Implication of Gli2 and Gli3 as
primary mediators of Shh signaling. Development. 126:3915–3924.
1999.PubMed/NCBI
|
|
48
|
Lauth M and Toftgård R: Non-canonical
activation of GLI transcription factors: Implications for targeted
anti-cancer therapy. Cell Cycle. 6:2458–2463. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Damhofer H, Veenstra VL, Tol JA, van
Laarhoven HW, Medema JP and Bijlsma MF: Blocking Hedgehog release
from pancreatic cancer cells increases paracrine signaling potency.
J Cell Sci. 128:129–139. 2015. View Article : Google Scholar : PubMed/NCBI
|