Introduction
The effect of cytotoxic treatment in testicular
tumors is mainly moderated by the induction of apoptosis (1). Due to the rapid response after
exposure to chemotherapeutic agents an intact and effective
apoptotic pathway in most testicular cancers is assumed (2). Nevertheless, various patients show
complex clinical courses, including tumors with chemoresistence and
thus lower apoptosis rates (3).
Recent studies have shown that the elimination of cells may be also
conducted using the alternative cell death pathway ʻautophagyʼ
(4,5).
Autophagy is an evolutionarily conserved, pervasive
and multi-step ʻself-eatingʼ process, by which cytosolic material
is sequestered in a double-lipid membrane, delivered to the
lysosome for degradation and digested to provide energy and to
build the foundation for cell-survival (6). While autophagy has for long been
considered to be a bulk degradation process, recent discoveries
show that it rather displays a sophisticated portfolio of
selectivity provided by different molecular strategies such as the
detection of several autophagy receptor proteins, including
pacemaker molecules such as p62 (SQSTM1) and BCL2-associated
athanogene 3 (BAG3) (7–9). To the best of our knowledge, no study
has been published to date that examined the expression of the
cohesive autophagy proteins BAG3 and p62 in testicular cancer. The
aim of our study was to get first evidence for the potential role
of autophagy-associated factors in the development and progression
of testicular cancers. Therefore, we performed immunohistochemistry
and western blot analyses for p62 and BAG3 of testicular cancer
specimens of different entities and corresponding normal appearing
testis tissue.
Materials and methods
Patient selection and clinicopathological
analysis
Orchiectomy specimens of 84 testicular cancer
patients from the Goethe University Hospital Frankfurt, Germany
from 2002 to 2011 were evaluated. All patients provided written
informed consent for the use of their tissues, and the study was
approved by the Ethics Committee of the Goethe University
Frankfurt/Main, Germany. All patients revealed adequate clinical
follow-up data.
Tissue microarray
All tissue samples were retrieved from orchiectomy
specimens, fixed in 10% buffered formalin and embedded in paraffin
at time of surgery. For tissue microarray (TMA) construction
hematoxylin and eosin-stained slides of human testicular germ cell
tumors (TGCTs) were reviewed by a certified pathologist. The most
representative areas of each tumor sample, and assigned
corresponding biopsies of the transitional zone and the unaffected
sites of the orchiectomy specimens, were marked and adequate cores
were removed from the formalin-fixed paraffin-embedded (FFPE) block
in triplicates. These samples were assembled in the array. The TMA
location number was linked to the database including the
clinicopathological data.
Immunohistochemistry
An automated immunostainer (Ventana, Strasbourg,
France) using standard protocols was used for immunohistochemical
staining. Briefly, 4-µm samples were heated to 100°C,
incubated with Inhibitor D (Ventana), and then incubated with the
primary antibody (antibodies: p62, monoclonal mouse anti-p62 Ick
ligand; dilution: 1:100; BD Transduction Laboratories, Franklin
Lakes, NJ, USA; BAG3, polyclonal rabbit antibody; dilution, 1:100;
Abnova, Taipei City, Taiwan; cleaved caspase 3, polyclonal rabbit
antibody; dilution: 1:100; Cell Signaling, Danvers, MA, USA). The
secondary antibody solution was incubated after rinsing, followed
by sequential incubation with Blocker D and SA-HRP D (both from
Ventana). Visualization was accomplished using diaminobenzidine
(DAB) D and DAB H2O2 D (Ventana). Finally,
the slides were counterstained with hemalaun and mounted. The
detailed immunohistochemical staining procedure was previously
described (10). Notably, not every
core of the TMA was evaluable for each protein (p62/BAG3) due to
technical reasons, resulting in variations of numbers of the
analyzed tissue specimens.
Scoring
The stained TMAs were evaluated with an Olympus BX50
light microscope. For semi-quantitative evaluation of the slides, a
multi-score of staining frequency and intensity was applied.
Staining frequency was assessed as follows: 0 (0–1%), 1 (1–10%), 2
(10–25%), 3 (25–50%), and 4 (>50%). The staining intensity was
rated as follows: no staining, 0; weak, 1; moderate, 2; and strong,
3. This scoring system was previously described (11).
SDS-PAGE and western blotting
Frozen tissue of 3 tumors were available for
analysis. Tumor and normal tissue were snap-frozen in liquid
nitrogen, homogenized and incubated in lysis buffer (20 mM Tris-HCl
buffer, pH 7.4; 150 mM NaCl; 1.5 mM ethylenediaminetetraacetic acid
(EDTA); 3% glycerol; 1 mg/ml BSA; 1% NP-40; 1 mg/ml leupeptin; 5
mg/ml aprotinin; 2 mg/ml pepstatin A) for 60 min at 0°C. Protein
lysates were loaded onto 12 or 15% SDS-polyacrylamide gels.
Proteins were separated at 135 V and then blotted to nitrocellulose
membranes (Protran BA83; 2 ml; Schleicher und Schuell, Dassel,
Germany) in Towbin buffer [25 mM Tris, 192 mM glycine and 20%
methanol (v/v)] at 15 V for 35 min. The blots were blocked in
blocking buffer (5% BSA, 50 mM Tris-HCl pH 7.5, 150 mM NaCl and
0.05% Tween-20) at 20°C for 1 h. The resulting blots were probed
with a rabbit monoclonal anti-BAG3 antibody diluted 1:3,000
(Biozol, Abnova), a mouse monoclonal anti-p62 antibody diluted
1:1,000 (BD Biosciences), a rabbit monoclonal anti-caspase 3
antibody diluted 1:1,000 (Cell Signaling) and a mouse monoclonal
anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibody
(Calbiochem) diluted 1:10,000.
Statistical analysis
The semi-quantitative scores were assigned as
ordinal scale response variable and analyzed along with nominal
variables (tissue/tumor type). Non-parametric Wilcoxon test was
used for statistical analyses. Corresponding normal and tumor
tissues were assessed using a matched-pairs analyses. A
significance level α=0.05 was selected for all tests. Statistical
analysis was performed using JMP 11.0.0 software (SAS, Cary, NC,
USA).
Results
BAG3, but not p62, is significantly
upregulated in seminoma as compared to both non-seminoma and normal
testicular tissue
Immunohistochemical analyses for p62 (Fig. 1A–C) revealed that p62 protein was
mainly detected within the parenchyma of normal testicular tissue
(Fig. 1A), which was resected
adjacent to non-seminomas (Fig. 1B)
or classical seminomas (Fig. 1C).
Notably, in both normal (Fig. 1A)
and neoplastic (Fig. 1B and C)
testicular tissue, stromal tissue components remained largely
negative for p62. In general staining scores for p62 were rather
low reaching a median expression score of 3 in both normal (range
1–6) and neoplastic (range 0–9) testicular tissue.
Only slightly higher expression values were obtained
for BAG3. While normal testicular tissue (Fig. 1D) displayed a median expression
score of 4 (range 0–9), non-seminomas (Fig. 1E) and seminomas (Fig. 1F) showed a median expression score
of 3.5 (range 0–9) and 6 (range 1–12), respectively. In contrast to
p62, BAG3 was also weakly expressed in stromal tissue of both
normal (Fig. 1D) and neoplastic
(Fig. 1E and F) specimens.
Statistical analyses of the immunohistochemical
staining revealed no significant differences in p62 expression
between normal and neoplastic testicular tissue (Fig. 2A) or between non-seminoma and
seminoma (Fig. 2B).
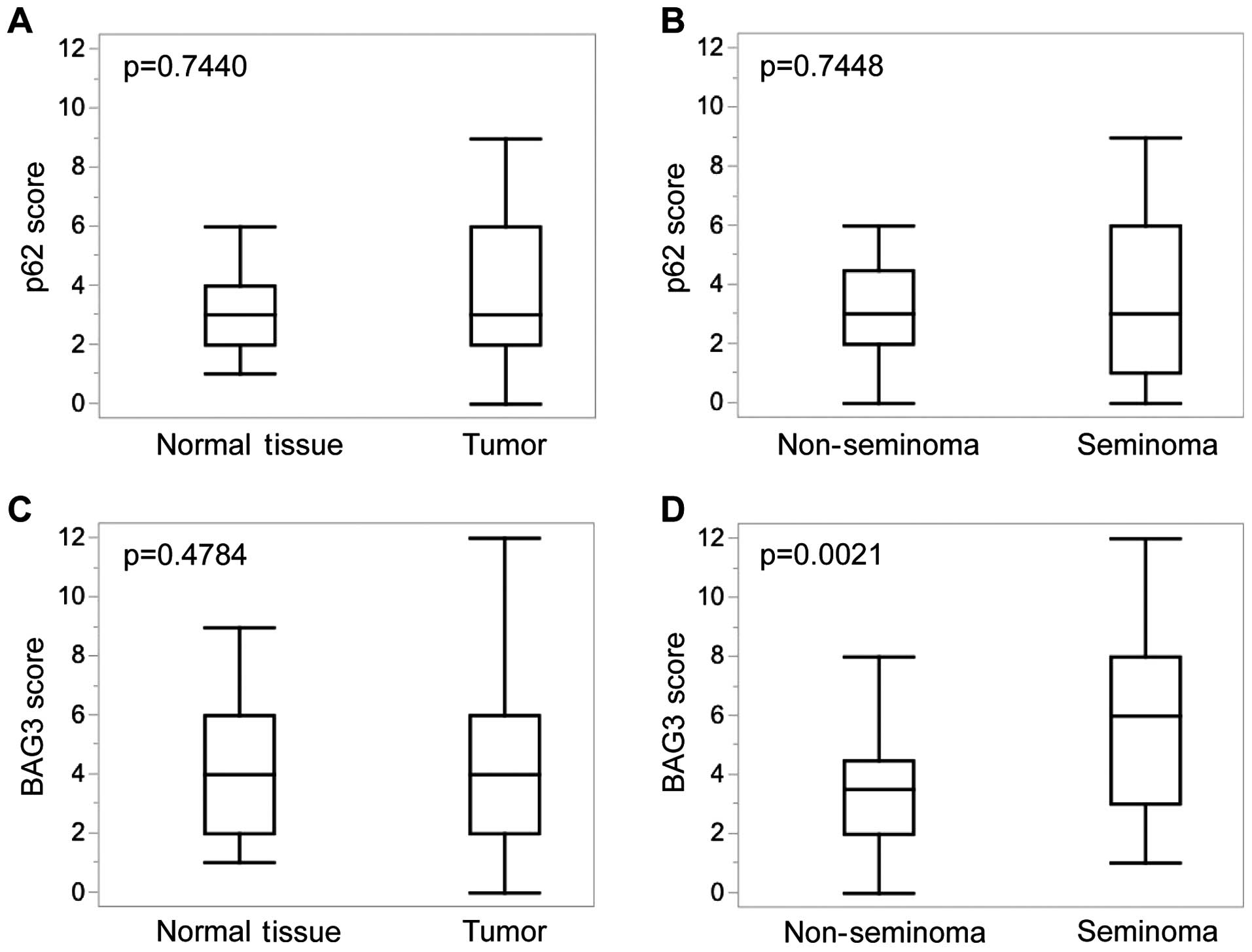 | Figure 2Statistical analysis of
immunohistochemical stainings of TMA. Immunohistochemical staining
scores are depicted for (A) p62 expression in 19 punches of normal
appearing testicular tissue (median, 3; range, 1–6) and 66 punches
of tumor tissue (median, 3; range; 0–9). (B) p62 expression in
tumor tissue punches of 34 non-seminoma (median, 3; range, 0–9) and
32 seminoma specimens (median; 3; range, 0–9). (C) BAG3 expression
in 23 normal appearing testicular tissue (median, 4; range, 0–9)
and 73 punches of tumor tissue (median, 4; range, 0–12). (D) BAG3
expression in tumor tissue punches of 42 non-seminoma (median, 3.5;
range, 0–9) and 31 seminoma specimens (median, 6; range, 1–12).
p-values were obtained using non-parametric Wilcoxon test. |
While no significant differences were detected for
BAG3, expression in normal testicular tissues when compared to
testicular tumor tissues in general (Fig. 2C), seminomas displayed significantly
higher BAG3 expression levels (p=0.0021) when compared to their
non-seminomatous counterparts (Fig.
2D).
A matched-pairs analysis for p62 and BAG3 expression
in non-neoplastic testicular tissues and their corresponding
adjacent neoplastic tissues did not reveal any significant
differences between the tissues (data not shown).
In contrast to the autophagy markers investigated in
our study, apoptosis rates were significantly higher in testicular
tumors as compared to tumor-adjacent normal testicular tissue while
no differences in the apoptosis levels were seen in the comparison
between seminomas and non-seminomas (Fig. 3). It is worthwhile mentioning that
all investigated testicular tumors were chemonaive indicating that
the detected apoptosis levels in our cohort reflect the endogenous
rate of type-I cell death. Western blot analysis demonstrated that
in contrast to unequivocally increased apoptosis levels in TGCT
tissues, similarly or even slightly higher expression levels of
BAG3 and/or p62 were detected in normal testicular control tissue
as compared to corresponding TGCT samples (Fig. 4).
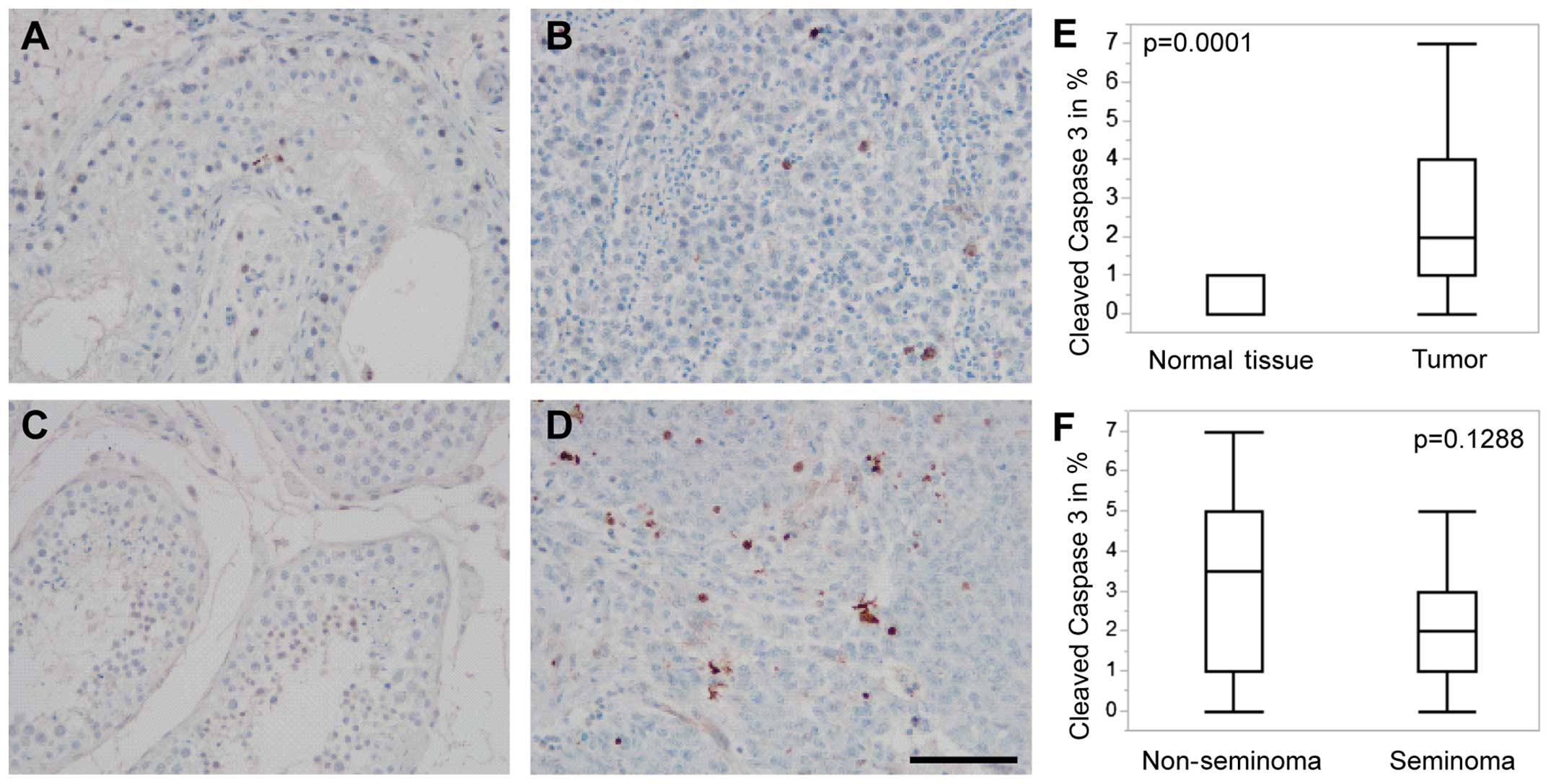 | Figure 3Cleaved caspase 3 expression in
testicular cancer. (A) Cleaved caspase 3 immunohistochemistry in
normal appearing testicular tissue (seminoma patient). (B) Cleaved
caspase 3 immunohistochemistry in a seminoma (A and B, tissue from
the same patient). (C) Cleaved caspase 3 immunohistochemistry in
normal appearing testicular tissue (non-seminoma patient). (D)
Cleaved caspase 3 immunohistochemistry, empryonal carcinoma (C and
D, tissue from the same patient) (scale bar, 100 µm for all
images). (E) Cleaved caspase 3-positive cells in % in 21 punches of
normal appearing testis tissue (mean, 1%; range 0–1%) and 50
punches of tumor tissue (median, 2%; range, 0–7%). (F) Cleaved
caspase 3-positive cells in % in 20 punches of non-seminomas
(median, 3.5%; range, 0–7%) and 30 seminomas (median, 2%; range,
0–7%); p-values were obtained using non-parametric Wilcoxon
test. |
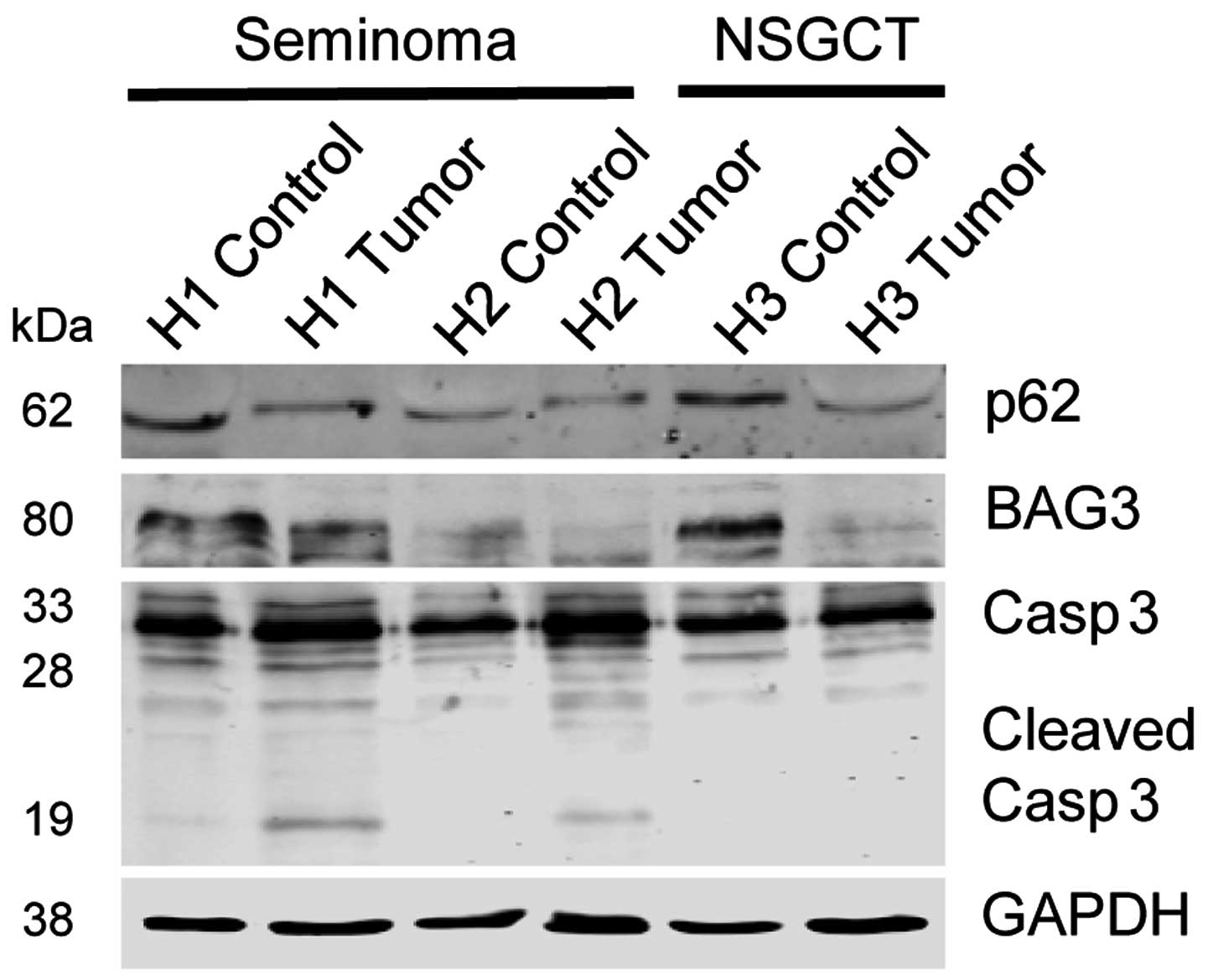 | Figure 4Protein biochemical analyses of
autophagy- and apoptosis-associated molecules in normal and
neoplastic testicular tissues. Western blot analysis of p62, BAG3
and cleaved caspase 3. GAPDH served as a loading control. H1 is a
specimen of a seminoma pT2, L1, V0 and R0. H2 represents tissue of
a seminoma pT3, L1, V0 and R0. H3 is an NSGCT, mixed malignant germ
cell tumor consisting of a malignant teratoma (>50%), embryonic
carcinoma (~30%) with a yolk sac tumor component, pT2, L1, V1 and
R0. |
p62 and BAG3 display distinct
loco-regional expression patterns in normal and neoplastic human
testicular tissues
Although, except for BAG3 expression in seminomatous
vs. non-seminomatous tumors, statistical comparisons for p62 or
BAG3 expression did not reveal significant differences between
normal and neoplastic testicular tissues or between the distinct
seminomatous and non-seminomatous tumors, we aimed at taking a
closer look at the immunohistochemically stained specimens with
regard to potential expression differences in areas with distinct
microenvironmental properties. In some perivascular areas in
seminomas, both p62 (Fig. 5A and
Aa) and BAG3 (Fig. 5B and Ba)
expression was considerably stronger as compared to more remote
tumor regions (Fig. 5Ab and
Bb).
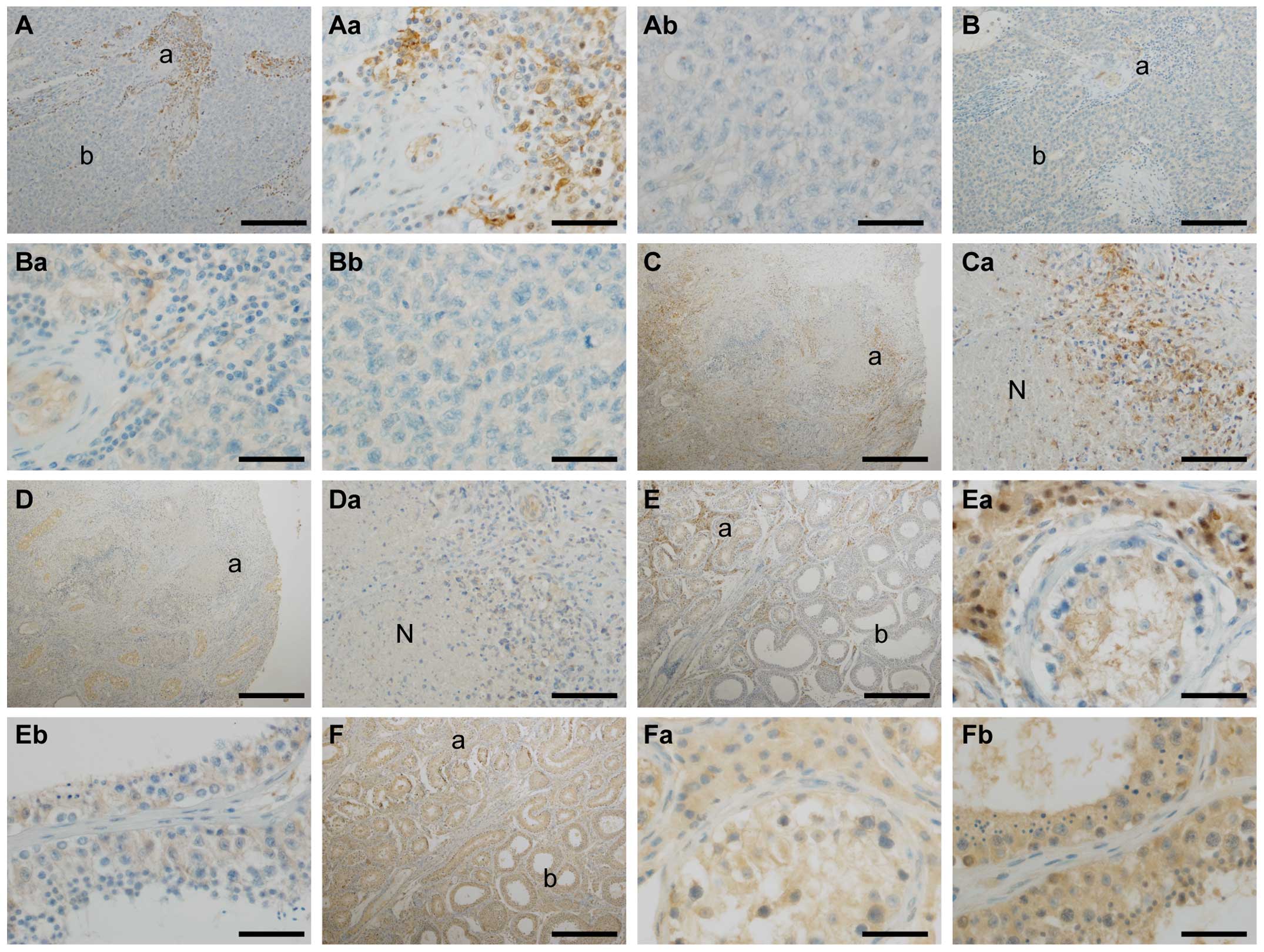 | Figure 5p62 and BAG3 protein expression
display distinct loco-regional patterns. (A) p62
immunohistochemistry, seminoma, scale bar, 200 µm; (a)
location of the zoomed-in image (Aa); (b) location of the zoomed-in
image (Ab); (Aa) cut-out of (A), scale bar, 50 µm; (Ab)
cut-out of (A), scale bar, 50 µm. (B) BAG3
immunohistochemistry, seminoma, scale bar, 200 µm; (a)
location of the zoomed-in image (Ba); (b) location of the zoomed-in
image (Bb); (Ba) cut-out of (B), scale bar, 50 µm; (Bb)
cut-out of (B), scale bar, 50 µm. (C) p62
immunohistochemistry, testicular embryonal carcinoma with necrosis;
(a) location of the zoomed-in image (Ca), scale bar, 500 µm;
(Ca) cut-out of (C), 'N' indicating necrosis, scale bar, 100
µm. (D) BAG3 immunohistochemistry, testicular embryonal
carcinoma; (a) location of the zoomed-in image (Da), scale bar: 500
µm; (Da) cut-out of (D), 'N' indicating necrosis, scale bar,
100 µm. (E) p62 immunohistochemistry, overview of a border
zone of a seminoma, scale bar, 500 µm; (a) ocation of the
zoomed-in image (Ea); (b) location of the zoomed-in image (Eb);
(Ea) cut-out of (E), scale bar, 50 µm; (Eb) cut-out of (E),
scale bar, 50 µm. (F) BAG3 immunohistochemistry, overview of
a border zone of a seminoma, scale bar, 500 µm; (a) location
of the zoomed-in image (Fa); (b) location of the zoomed-in image
(Fb); (Fa) cut-out of (F), scale bar, 50 µm; (Fb) cut-out of
(F), scale bar, 50 µm. |
Another repeatedly observed distinct staining
pattern was found in perinecrotic areas of higher malignant
testicular tumors such as embryonal carcinomas (Fig. 5C and D). Both p62 (Fig. 5C) and BAG3 (Fig. 5D) were considerably upregulated in
close proximity to necrotic foci, however, decreasing again at a
distance of only a few micrometers from the border of the necrotic
zone.
Finally, a slightly stronger protein expression of
p62 (Fig. 5E) and BAG3 (Fig. 5F) was observed at the border zone
between normal and neoplastic testicular tissue. Although the
immunohistochemical results did not reach significance when the
whole specimens were taken into account, these loco-regional
staining differences may provide a more sophisticated view on
autophagy-associated molecules in normal and neoplastic testicular
tissues.
Discussion
Autophagy is highly contextual in regard to tumor
cell death and survival decisions. In general autophagy acts as a
pro-survival stress response, but it can also trigger
cytodestructing effects (12–15).
In our study we investigated the expression profiles of BAG3 and
p62 in different types of testicular cancer and normal appearing
testis tissue to gain insight into a potential in vivo
relevance of autophagy in the development and progression of
testicular cancer. Both p62 and BAG3 play a crucial role in the
sophisticated portfolio of autophagy (16).
BAG3 was shown to sustain cell survival and it
underlies resistance mechanisms to chemotherapy in several human
neoplasms in vitro (17–19).
Protein homeostasis in aged (or stressed) cells is driven by an
increased BAG3 expression, regulated by the transcription factor
heat shock factor 1 (HSF1) (20)
and stabilized by the recruitment of the macro-autophagy pathway
involving HSP70, SQSTM1/p62 and LC3 (7,21,22).
It is also known that BAG3 mitigates proteotoxicity via selective
autophagy (23).
The cargo receptor p62 functionally interfaces to
the autophagic machinery through degradation of ubiquitin-tagged
proteins. p62 recruits protein aggregates as substrate to
autophagic membranes to form autophagosomes and thus promotes their
degradation (24). In addition, p62
itself is a substrate for autophagic degradation and accumulates
upon inhibition of autophagy (24,25).
We noted that BAG3 and p62 are not upregulated in
testicular cancers as compared to normal testicular tissue in both
immunohistochemical and immunoblotting analyses. The normal tissue
even shows slightly higher expression levels of BAG3 and/or p62.
The expansive growing tumor may induce pressure on the normal
tissue particularly in a cloistered organ such as the testis. The
resulting insufficient supply of e.g. oxygen and changes in
microenvironmental nutrition may induce autophagy in order to
mitigate the adverse condition and then avoid cell damage or even
cell death (26). Our findings
implicate that microscopically normal appearing testicular tissue
may be stressed at the molecular level due to the neighboring
pathologies and thus induces protective mechanisms.
In regards to BAG3 these results are contrary to
previous studies which detected substantial BAG3 expression in
several types of human cancers, such as glioblastoma, pancreatic
carcinoma, leukemia and thyroid carcinoma, compared with very low
basal levels of BAG3 in non-malignant cells (18,27,28).
Other studies have reported that an increased expression of BAG3
was shown to induce the formation of p62 bodies (9) and thus promoting selective autophagy
via its interaction with p62. In our case, p62 may not be
upregulated, hypothetically due to the fact that p62 itself is a
substrate for autophagic degradation (9). We observed that in both normal and
neoplastic testicular tissue, stromal tissue components remained
largely negative for p62. In contrast, previous studies have shown
that stromal p62 expression was associated with the histological
grade and molecular subtype (29,30).
Our results demonstrated that there is no significant difference in
autophagy induction in neoplastic tissue when compared to normal
testicular tissue. Contrary to these findings, apoptotic activity
is significantly increased in testicular tumor, but not in normal
testicular tissue, as proliferative tumor tissue of a certain size
usually becomes hypoxic, eventually necrotic and thus enhanced
apoptosis can be detected particularly at the margins of hypoxic or
necrotic tumor areas (31,32). Up to now the loco-regional
expression patterns of autophagy in TGCTs have not been a subject
of research. We believe that a ʻtopographicalʼ point-of-view may be
a determinant for the interpretation of our results. In our case,
areas of crucial relevance for local and systemic tumor progression
and survival show increased expression patterns of p62 and BAG3.
When having a holistic view on the staining a slightly stronger
protein expression for p62 and BAG3 was noted at the border zone
where tumor pins protrude into the normal testicular tissue, a fact
that may represent a protective barrier, in order to protect the
tumor.
Another repeatedly observed distinct staining
pattern of BAG3 and p62 was found in perinecrotic (where presumably
hypoxia is maximal) areas of higher malignant testicular tumors
such as embryonal carcinomas. Autophagy induction in cancer cells
may occur due to hypoxia (13,32),
and it has been previously demonstrated that expression of
hypoxia-inducible factor 1α (HIF-1α) downstream targets is
associated with a higher histologic grade in fibroepithelial tumors
of the breast (33). In this
scenario autophagy provides the nutrients needed for cancer
survival and stabilization of HIF-1α allows for instance that
higher grade phyllode tumors adapt to hypoxia (30). In another clinical setting, the
cardiac infarction, this hypothesis has been verified. Autophagy is
more prominent in cardiomyocytes in the border zone than in those
in more remote areas, and the size of the autophagosomes was
exceedingly large in the former. Previously, Kanamori et al
suggested a protective role during post-infarction cardiac
remodeling (34). Surprisingly
contrary to these findings, in some perivascular areas in seminoma,
presumably a site with optimal oxygenation and nutrient status,
both p62 and BAG3 expression was considerably stronger as compared
to more remote tumor regions.
Up to now there are no studies that have explored
the relationship between seminoma and non-seminoma in respect to
autophagy. We observed that BAG3 is significantly upregulated in
seminoma compared to non-seminoma. This difference may be due to
the heterogeneity of testicular germ cell tumors (TGCT). As stated
by Elstrand et al, discrepancies may be due to the very
complex types of intracellular molecular interaction of the BAG3
proteins (i.e. with HSP proteins and Bcl-2 members) in tumors of
various origin and differentiation (22,27,35,36).
While seminoma and non-seminoma show great similarity in etiology,
precursor lesions and chromosome 12p alterations, different TGCT
subtypes display distinct tumor genetics that may explain their
different histology and biological behavior. Studies have revealed
differences between seminoma and non-seminoma e.g. for KRAS, NRAS
and BRAF (37). Different genetic
factors may also influence the autophagic behavior of the
testicular tumor entities. Further investigations are necessary to
determine the exact expression profile of autophagy proteins in
different types of testicular cancer.
The limitations of the present study are defined
through the fact that autophagy is a dynamic, multistep process and
the accurate measurement of autophagy involves the quantification
of autophagic flux (38).
Therefore, the identification of proteins associated with autophagy
or those that are degraded by autophagy through quantitative
methods such as immunohistochemistry (IHC)/tissue microarray (TMA)
may be problematic. These methods provide a snapshot of the
identified proteins at a specific point in time and are actually
unable to reflect the activity of autophagy.
In conclusion, even though autophagy markers display
a loco-regional expression pattern in normal and neoplastic human
testicular tissue, our findings imply that in general apoptosis,
but not autophagy induction differs between normal and neoplastic
testis tissue. The precise differences and interrelationships of
autophagy in respect to seminoma and non-seminoma have to be
further investigated with an autophagy-related molecular
calibration of these two distinct types of testicular cancer.
References
|
1
|
Mayer F, Stoop H, Scheffer GL, Scheper R,
Oosterhuis JW, Looijenga LH and Bokemeyer C: Molecular determinants
of treatment response in human germ cell tumors. Clin Cancer Res.
9:767–773. 2003.PubMed/NCBI
|
|
2
|
Spierings DC, de Vries EG, Vellenga E and
de Jong S: The attractive Achilles heel of germ cell tumours: An
inherent sensitivity to apoptosis-inducing stimuli. J Pathol.
200:137–148. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lian J, Wu X, He F, Karnak D, Tang W, Meng
Y, Xiang D, Ji M, Lawrence TS and Xu L: A natural BH3 mimetic
induces autophagy in apoptosis-resistant prostate cancer via
modulating Bcl-2-Beclin1 interaction at endoplasmic reticulum. Cell
Death Differ. 18:60–71. 2011. View Article : Google Scholar
|
|
5
|
Voss V, Senft C, Lang V, Ronellenfitsch
MW, Steinbach JP, Seifert V and Kögel D: The pan-Bcl-2 inhibitor
(−)-gossypol triggers autophagic cell death in malignant glioma.
Mol Cancer Res. 8:1002–1016. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakatogawa H, Suzuki K, Kamada Y and
Ohsumi Y: Dynamics and diversity in autophagy mechanisms: Lessons
from yeast. Nat Rev Mol Cell Biol. 10:458–467. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Amaravadi RK, Lippincott-Schwartz J, Yin
XM, Weiss WA, Takebe N, Timmer W, DiPaola RS, Lotze MT and White E:
Principles and current strategies for targeting autophagy for
cancer treatment. Clin Cancer Res. 17:654–666. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gamerdinger M, Hajieva P, Kaya AM, Wolfrum
U, Hartl FU and Behl C: Protein quality control during aging
involves recruitment of the macroautophagy pathway by BAG3. EMBO J.
28:889–901. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baumgarten P, Harter PN, Tönjes M, Capper
D, Blank AE, Sahm F, von Deimling A, Kolluru V, Schwamb B,
Rabenhorst U, et al: Loss of FUBP1 expression in gliomas predicts
FUBP1 mutation and is associated with oligodendroglial
differentiation, IDH1 mutation and 1p/19q loss of heterozygosity.
Neuropathol Appl Neurobiol. 40:205–216. 2014. View Article : Google Scholar
|
|
11
|
Harter PN, Bunz B, Dietz K, Hoffmann K,
Meyermann R and Mittelbronn M: Spatio-temporal deleted in
colorectal cancer (DCC) and netrin-1 expression in human foetal
brain development. Neuropathol Appl Neurobiol. 36:623–635. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Baehrecke EH: Autophagy: Dual roles in
life and death? Nat Rev Mol Cell Biol. 6:505–510. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Degenhardt K, Mathew R, Beaudoin B, Bray
K, Anderson D, Chen G, Mukherjee C, Shi Y, Gélinas C, Fan Y, et al:
Autophagy promotes tumor cell survival and restricts necrosis,
inflammation, and tumorigenesis. Cancer Cell. 10:51–64. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roy S and Debnath J: Autophagy and
tumorigenesis. Semin Immunopathol. 32:383–396. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Galluzzi L, Pietrocola F, Bravo-San Pedro
JM, Amaravadi RK, Baehrecke EH, Cecconi F, Codogno P, Debnath J,
Gewirtz DA, Karantza V, et al: Autophagy in malignant
transformation and cancer progression. EMBO J. 34:856–880. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Johansen T and Lamark T: Selective
autophagy mediated by autophagic adapter proteins. Autophagy.
7:279–296. 2011. View Article : Google Scholar :
|
|
17
|
Chen HY, Liu P, Sun M, Wu LY, Zhu HY, Qiao
C, Dong HJ, Zhu DX, Xu W and Li JY: Bag3 gene expression in chronic
lymphocytic leukemia and its association with patients' prognosis.
Zhongguo Shi Yan Xue Ye Xue Za Zhi. 18:838–842. 2010.In Chinese.
PubMed/NCBI
|
|
18
|
Liao Q, Ozawa F, Friess H, Zimmermann A,
Takayama S, Reed JC, Kleeff J and Büchler MW: The anti-apoptotic
protein BAG-3 is overexpressed in pancreatic cancer and induced by
heat stress in pancreatic cancer cell lines. FEBS Lett.
503:151–157. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boiani M, Daniel C, Liu X, Hogarty MD and
Marnett LJ: The stress protein BAG3 stabilizes Mcl-1 protein and
promotes survival of cancer cells and resistance to antagonist
ABT-737. J Biol Chem. 288:6980–6990. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rosati A, Ammirante M, Gentilella A,
Basile A, Festa M, Pascale M, Marzullo L, Belisario MA, Tosco A,
Franceschelli S, et al: Apoptosis inhibition in cancer cells: A
novel molecular pathway that involves BAG3 protein. Int J Biochem
Cell Biol. 39:1337–1342. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Behl C: BAG3 and friends: Co-chaperones in
selective autophagy during aging and disease. Autophagy. 7:795–798.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X, Colvin T, Rauch JN, Acosta-Alvear D,
Kampmann M, Dunyak B, Hann B, Aftab BT, Murnane M, Cho M, et al:
Validation of the Hsp70-Bag3 protein-protein interaction as a
potential therapeutic target in cancer. Mol Cancer Ther.
14:642–648. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rapino F, Jung M and Fulda S: BAG3
induction is required to mitigate proteotoxicity via selective
autophagy following inhibition of constitutive protein degradation
pathways. Oncogene. 33:1713–1724. 2014. View Article : Google Scholar
|
|
24
|
Yao TP: The role of ubiquitin in
autophagy-dependent protein aggregate processing. Genes Cancer.
1:779–786. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hua F, Li K, Yu JJ, Lv XX, Yan J, Zhang
XW, Sun W, Lin H, Shang S, Wang F, et al: TRB3 links insulin/IGF to
tumour promotion by interacting with p62 and impeding
autophagic/proteasomal degradations. Nat Commun. 6:79512015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Filomeni G, De Zio D and Cecconi F:
Oxidative stress and autophagy: The clash between damage and
metabolic needs. Cell Death Differ. 22:377–388. 2015. View Article : Google Scholar :
|
|
27
|
Romano MF, Festa M, Petrella A, Rosati A,
Pascale M, Bisogni R, Poggi V, Kohn EC, Venuta S, Turco MC, et al:
BAG3 protein regulates cell survival in childhood acute
lymphoblastic leukemia cells. Cancer Biol Ther. 2:508–510. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Festa M, Del Valle L, Khalili K, Franco R,
Scognamiglio G, Graziano V, De Laurenzi V, Turco MC and Rosati A:
BAG3 protein is overexpressed in human glioblastoma and is a
potential target for therapy. Am J Pathol. 178:2504–2512. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choi J, Jung W and Koo JS: Expression of
autophagy-related markers beclin-1, light chain 3A, light chain 3B
and p62 according to the molecular subtype of breast cancer.
Histopathology. 62:275–286. 2013. View Article : Google Scholar
|
|
30
|
Kim JY, Jung WH and Koo JS: Expression of
autophagy-related proteins according to androgen receptor and HER-2
status in estrogen receptor-negative breast cancer. PLoS One.
9:e1056662014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qiu Y, Li P and Ji C: Cell death
conversion under hypoxic condition in tumor development and
therapy. Int J Mol Sci. 16:25536–25551. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kuijper A, van der Groep P, van der Wall E
and van Diest PJ: Expression of hypoxia-inducible factor 1 alpha
and its downstream targets in fibroepithelial tumors of the breast.
Breast Cancer Res. 7:R808–R818. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kanamori H, Takemura G, Goto K, Maruyama
R, Tsujimoto A, Ogino A, Takeyama T, Kawaguchi T, Watanabe T,
Fujiwara T, et al: The role of autophagy emerging in postinfarction
cardiac remodelling. Cardiovasc Res. 91:330–339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Elstrand MB, Kleinberg L, Kohn EC, Tropé
CG and Davidson B: Expression and clinical role of antiapoptotic
proteins of the bag, heat shock, and Bcl-2 families in effusions,
primary tumors, and solid metastases in ovarian carcinoma. Int J
Gynecol Pathol. 28:211–221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Franceschelli S, Rosati A, Lerose R, De
Nicola S, Turco MC and Pascale M: Bag3 gene expression is regulated
by heat shock factor 1. J Cell Physiol. 215:575–577. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sheikine Y, Genega E, Melamed J, Lee P,
Reuter VE and Ye H: Molecular genetics of testicular germ cell
tumors. Am J Cancer Res. 2:153–167. 2012.
|
|
38
|
Mizushima N, Yoshimori T and Levine B:
Methods in mammalian autophagy research. Cell. 140:313–326. 2010.
View Article : Google Scholar : PubMed/NCBI
|















