Introduction
Breast cancer and esophageal squamous cell carcinoma
(ESCC) are well known to be sensitive to chemotherapy and/or
radiotherapy, and their combination with surgery has been proven to
have clinical benefits (1–6). A multidisciplinary procedure including
chemotherapy, radiotherapy and surgery is regarded as standard care
for breast cancer and ESCC patients. In clinical practice, in such
patients it is generally accepted that there are responders and
non-responders to chemotherapy with or without radiotherapy, and
limited information exists describing the mechanisms and biomarkers
to predict the responders.
Although chemo-radiotherapy (CRT) is aimed at
directly inducing apoptosis or necrosis, there is accumulating
evidence to support the novel concept that CRT may induce the
immunogenic tumor cell death (ICD) of tumor cells (7–18). CRT
could trigger uptake of antigenic components by dendritic cells
(DCs) and transfer antigenic signals to T-cell-mediated immunity,
resulting in the expansion of antigen-specific cytotoxic T
lymphocytes and production of tumor-specific monoclonal antibodies
(mAbs). We and others have shown that high-mobility group box 1
protein (HMGB1) and calreticulin induced by cytotoxic stresses such
as CRT are important mediators to induce ICD (19). However, whether ICD can be induced
by chemotherapy alone in the clinical setting remains unclear.
Here, we evaluated i) whether expression of HMGB1
and calreticulin correlates with clinical outcomes in response to
chemotherapy, ii) whether the number of CD8-positive
(CD8+) T cells correlates with expression of HMGB1 and
calreticulin, and iii) whether chemotherapy alone upregulates HMGB1
and calreticulin in clinical and in vitro settings.
Materials and methods
Patients and samples
The expression levels of HMGB1, calreticulin, and
CD8 were evaluated by immunohistochemistry in pre-treatment biopsy
specimens and surgically resected specimens obtained from breast
cancer patients (n=52) and ESCC patients (n=8) who had undergone
treatment with neoadjuvant chemotherapy (NAC) between 2005 and 2015
at the Department of Organ Regulatory Surgery, Fukushima Medical
University Hospital. Clinical and pathological information was
retrospectively obtained by review of the medical records, with the
last follow-up being in February 2016. Overall survival (OS) was
defined as the period from the date of surgery to the date of
death. The median follow-up time was 51.0 months. The study was
conducted in accordance with the Declaration of Helsinki and was
approved by the Institutional Review Board of Fukushima Medical
University (reference 2329 for ESCC and 2444 for breast
cancer).
Cell lines
Breast cancer cell lines, MDA-MB-231 [estrogen
receptor (ER)-negative and HER2-negative], MCF-7 (ER-positive and
HER2-negative) and SK-BR-3 (ER-negative and HER2-positive) were
obtained as previously described (20), and cultured in RPMI-1640 medium with
10% fetal calf serum (FCS), 50 U/ml penicillin and 50 µg/ml
streptomycin.
Immunohistochemistry
HMGB1, calreticulin and CD8 immunostaining was
conducted using the avidin-biotin-peroxidase complex method. Each
paraffin section was dewaxed, followed by antigen retrieval with
Target Retrieval Solution (10 mmol citrate buffer at pH 6.0; Dako,
Glostrup, Denmark) in an autoclave (121°C, 15 min). The sections
were cooled at room temperature for 30 min and endogenous
peroxidase was blocked with 3% hydrogen peroxide. Thereafter, the
sections were incubated with diluted normal blocking serum for 20
min and incubated with one of the following: mouse anti-human HMGB1
mAb (cat. no. SAB1403925, clone 2F6, 3 mg/ml; Sigma-Aldrich, Tokyo,
Japan) overnight at 4°C; mouse anti-human calreticulin mAb (cat.
no. ab22683, 5 mg/ml; Abcam, Cambridge, UK) for 2 h at 37°C; or
mouse anti-human CD8 mAb (cat. no. M7103, 1.6 µg/ml; Dako)
overnight at 4°C. Thereafter, the EnVision kit (cat. no. K4001;
Dako) was used for anti-mouse secondary antibody staining according
to the manufacturers protocol and 3,3-diaminobenzidine was used to
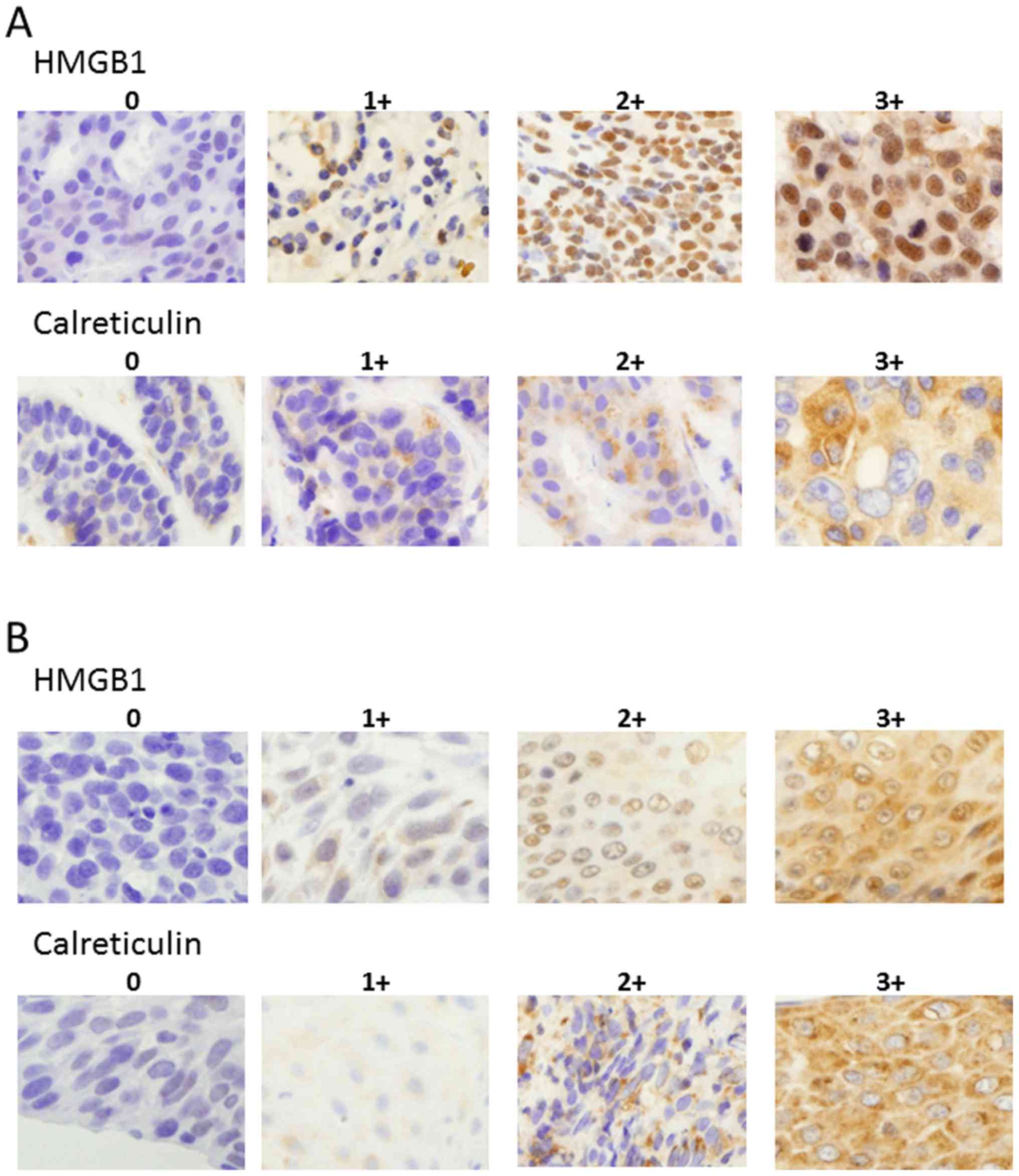
detect specific bindings. The grade of HMGB1 and calreticulin
expression was scored as 0 (0–10% positive), 1+ (>10–30%
positive), 2+ (>30–80% positive), or 3+ (> 80% positive) in
tumor cells (Fig. 1) in serial
sections using five randomly selected areas at a magnification of
×400. The CD8+ T cells were expressed as the mean values
in four randomly selected areas at a magnification of ×400.
Microscopic analyses were evaluated independently by two
investigators (K.A. and K.K.) who had no prior knowledge of the
clinical data.
In vitro treatments of breast cancer
cell lines with chemotherapeutic drugs
Three breast cancer cell lines, MDA-MB-231, MCF-7,
and SK-BR-3, were incubated with RPMI-1640 medium in 6-well plates.
Tumor cells were grown to subconfluency and treated with
chemotherapeutic drugs, paclitaxel (0.1–1 µM) or doxorubicin (0.1–1
µM), in serum-free medium (AIM V®; Thermo Fisher
Scientific, Inc., Wilimington, DE, USA) on day 0. Dying cells were
analyzed by Annexin V and 7-aminoactinomycin D (7-AAD) (both from
BD Pharmingen, San Jose, CA, USA) by flow cytometry, and the
proportion of dying cells was determined using either Annexin
V-positive or 7-AAD-positive cells. Supernatants of treated breast
cancer cell line cultures were measured for HMGB1 contents by ELISA
(Shino-Test Corporation, Tokyo, Japan), and cell surface expression
of calreticulin was evaluated by flow cytometry with
R-phycoerythrin-conjugated anti-calreticulin mAb (Enzo Life
Sciences, Farmingdale, NY, USA).
Statistical analysis
A paired t-test was used to determine the
differences in the HMGB1 score, calreticulin score, and the number
of CD8+ T cells before and after NAC. A Chi-square test
was used for the evaluation between chemo-response and HMGB1 score,
calreticulin score, and the number of CD8+ T cells, as
well as between the number of CD8+ T cells and both
HMGB1 and calreticulin scores. An unpaired t-test was used to
determine HMGB1 and calreticulin expression between the control
cell lines and the target cell lines treated with chemotherapeutic
drugs. Cumulative survival was estimated by the Kaplan-Meier
method, and the differences between the two groups were analyzed by
a log-rank test. All statistical analyses were two-sided and
conducted using Graphpad Prism v6.0 (Graphpad Software, Inc., La
Jolla, CA, USA). P-values <0.05 were considered statistically
significant.
Results
HMGB1 and calreticulin expression
before and after NAC
To evaluate HMGB1 and calreticulin expression within
the tumor microenvironment induced by chemotherapy alone,
immunohistochemical analysis was conducted in pre-treatment biopsy
specimens and surgically resected specimens obtained from
NAC-treated breast cancer and ESCC patients. Concerning NAC, breast
cancer patients (92%, n=48) were treated with
5-fluorouracil-epirubicin-cyclophosphamide (FEC or CEF)
with/without docetaxel or paclitaxel, and trastuzumab was added for
those with HER2-overexpressing tumors (Table I). All ESCC patients were treated
with 5-fluorouracil + cisplatin. In order to semi-quantitatively
evaluate HMGB1 and calreticulin expression, we classified the
patients into 4 grades (0, 1+, 2+ and 3+), as described in
Materials and methods section. The representative immunostainings
using anti-HMGB1 and anti-calreticulin mAbs are shown in Fig. 1.
 | Table I.Clinical course and prognosis of the
breast cancer cases depending on the chemotherapy regimen. |
Table I.
Clinical course and prognosis of the
breast cancer cases depending on the chemotherapy regimen.
| Regimen | No. of
patients | No. of patients
with recurrence | Relapse-free
survival [mean (range) months] | No. of deaths | Overall survival
[mean (range) months] |
|---|
| FEC +
docetaxel | 14 | 2 | 32
(8–108) | 1 | 34
(1–114) |
| FEC +
paclitaxel | 14 | 5 | 53
(1–114) | 3 | 65
(3–114) |
| FEC + docetaxel +
trastuzumab | 5 | 2 | 25 (4–38) | 1 | 27
(5–38) |
| FEC + paclitaxel +
trastuzumab | 5 | 2 | 70
(22–96) | 1 | 76
(53–96) |
| FEC | 5 | 1 | 75
(13–106) | 0 | 79
(33–108) |
| CEF | 4 | 2 | 73
(15–104) | 1 | 70
(18–104) |
| FEC + paclitaxel +
vinorelbine | 1 | 1 | 3 | 1 | 14 |
| Docetaxel | 1 | 0 | 15 | 0 | 15 |
| Paclitaxel +
carboplatin | 1 | 0 | 45 | 0 | 45 |
| Paclitaxel +
cisplatin | 1 | 0 | 32 | 1 | 32 |
| Paclitaxel +
trastuzumab | 1 | 0 | 22 | 0 | 22 |
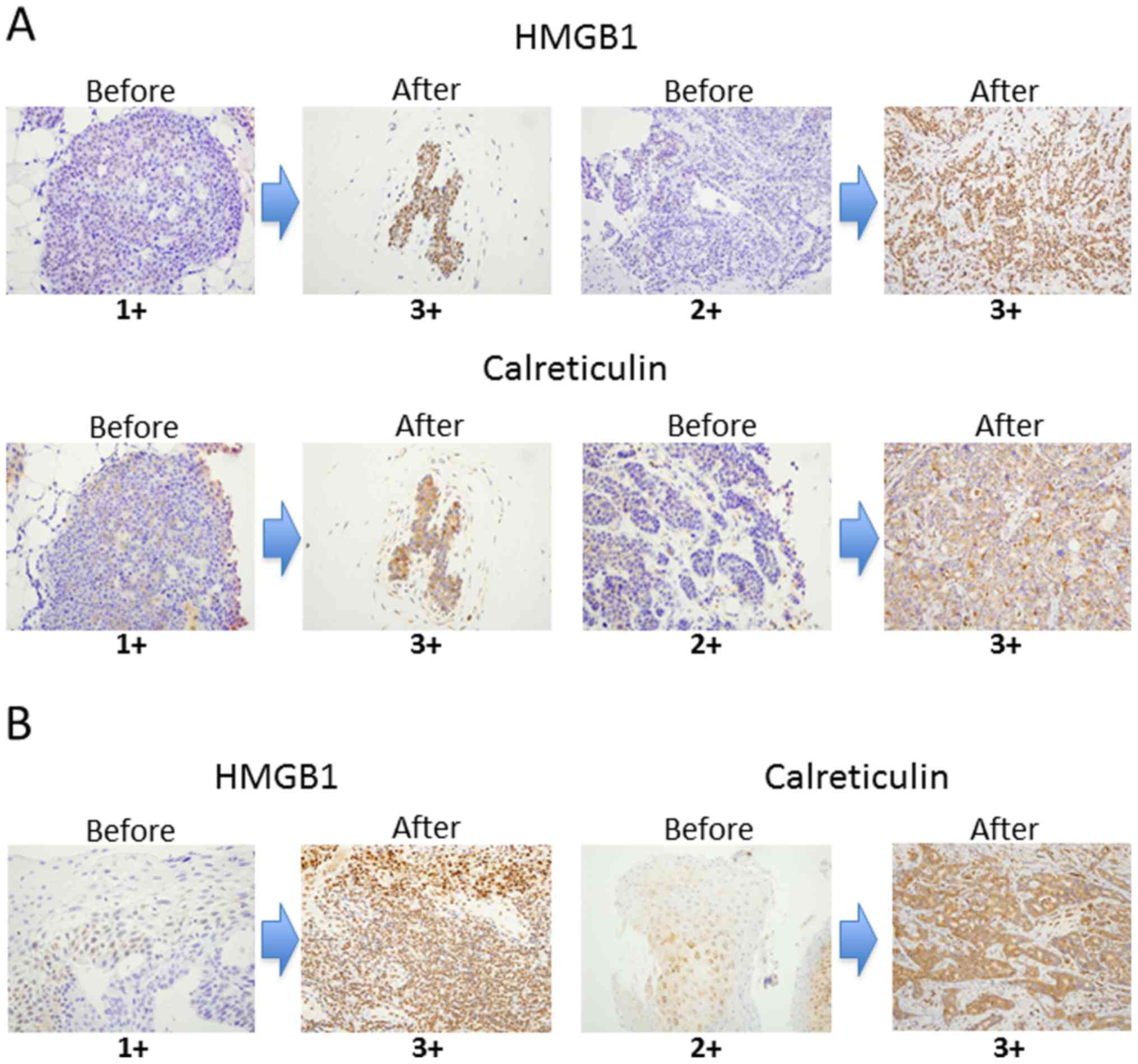
Both HMGB1 and calreticulin expression was
significantly upregulated after NAC compared to pre-treatment
samples in breast cancer and ESCC (Fig.
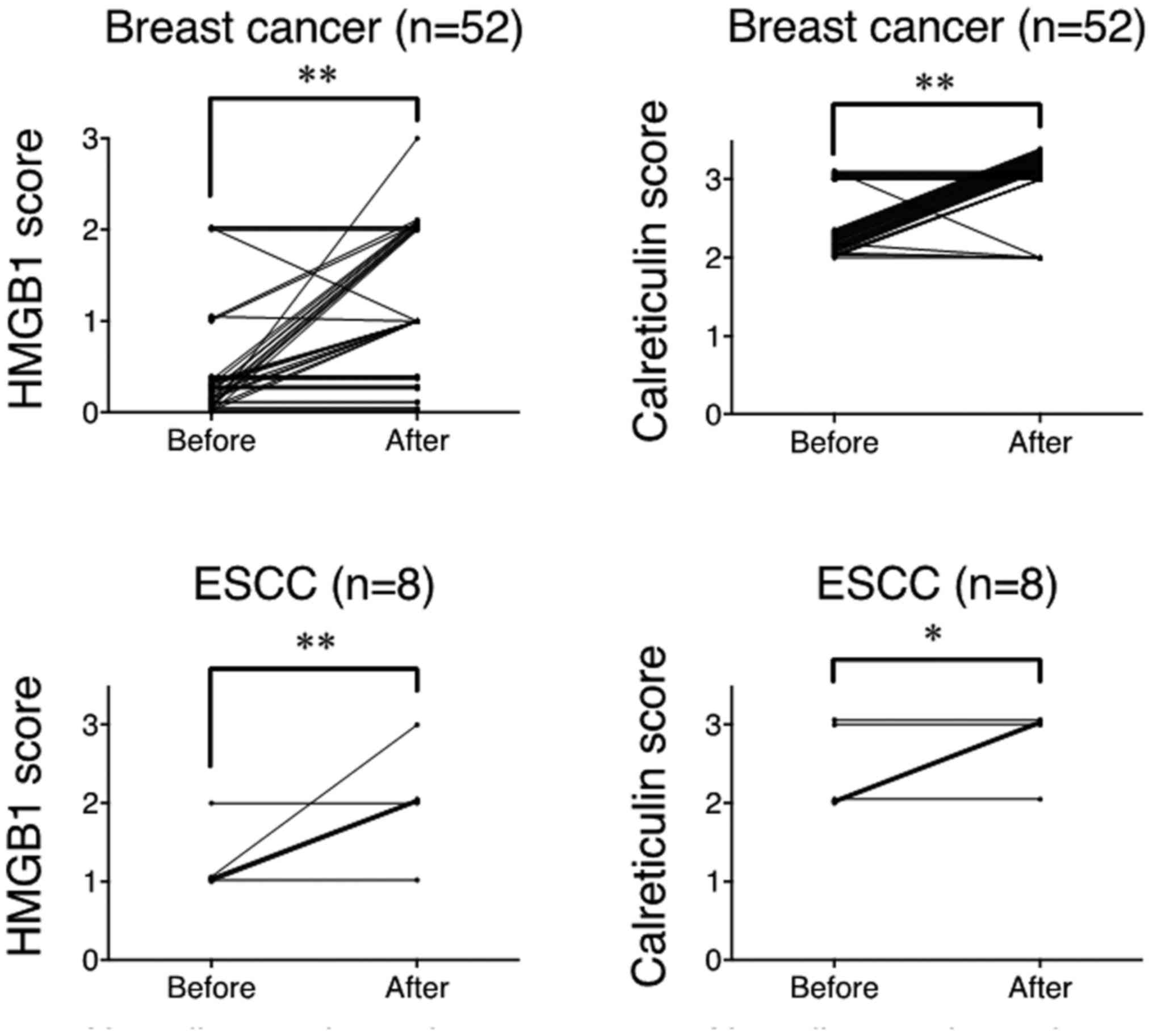
2). Summarized data from all samples showed that the degree of
HMGB1 and calreticulin expression was significantly upregulated
after NAC compared to the pre-treatment samples in breast cancer
and ESCC (Fig. 3). Thus, it is
strongly suggested that chemotherapy alone could upregulated HMGB1
and calreticulin expression in the tumor microenvironments in
breast cancer and ESCC.
Correlation of HMGB1 and calreticulin
expression with pathological responses after NAC and patient
survival
Since the number of patients with breast cancer was
enough to evaluate the clinical data, the evaluations of the
response rate and survival rate were performed in the breast cancer
specimens only. Patient and tumor characteristics in the breast
cancer cases are shown in Table
II. Tumors were classified according to the TNM Classification
of Malignant Tumors (UICC 7th edition) and histological criteria
made by the Japanese Breast Cancer Society for assessment of
therapeutic response was used to evaluate the pathological response
to NAC (21). As shown in Table III, there was no significant
correlation between HMGB1 score in the pre-treatment samples and
pathological response, or between HMGB1 score in the post-treatment
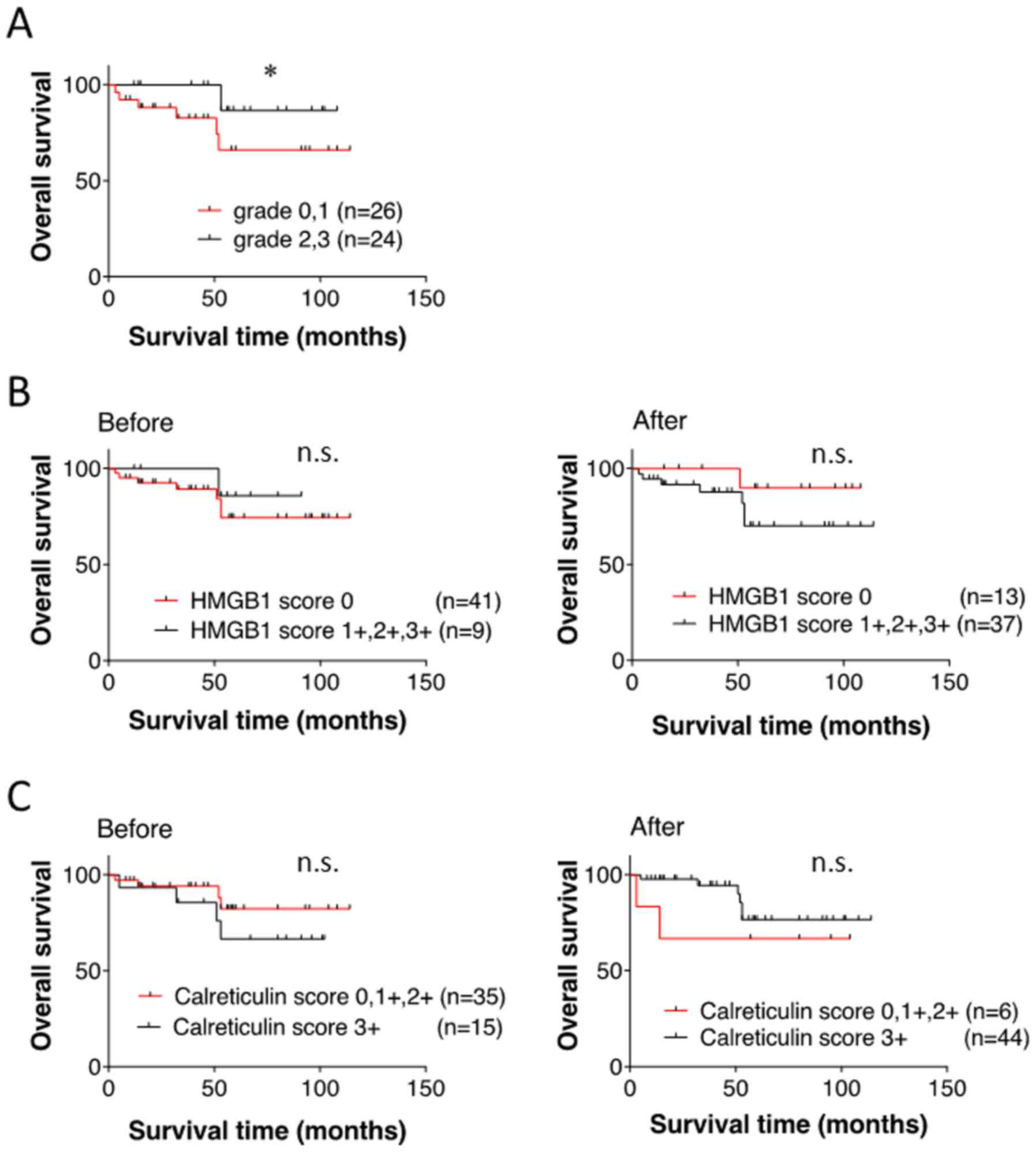
samples and pathological response. OS in the responder group to NAC
was significantly superior to that in the non-responder group
(Fig. 4A). However, there was no
significant difference in survival between HMGB1-high and HMGB1-low
scores in the pre-treatment samples or the post-treatment samples
(Fig. 4B). Similarly, calreticulin
expression in the pre- and post-treatment samples did not affect
pathological response and OS (Table
IV and Fig. 4C).
| Table II.Patient and tumor characteristics of
the breast cancer cases (n=52). |
Table II.
Patient and tumor characteristics of
the breast cancer cases (n=52).
|
Characteristics | n |
|---|
| Age (years) |
|
|
Mean | 53.4 |
|
Range | 26–75 |
| Sex |
|
|
Male | 1 |
|
Female | 51 |
| Tumora |
|
| T1 | 7 |
| T2 | 24 |
| T3 | 7 |
| T4 | 14 |
| Lymph node
metastasisa |
|
| N0 | 2 |
| N1 | 41 |
| N2 | 7 |
| N3 | 2 |
| Stagea |
|
| I | 0 |
| II | 27 |
|
III | 23 |
| IV | 2 |
| Histological
classification |
|
|
Invasive ductal carcinoma | 41 |
|
Non-invasive carcinoma | 1 |
|
Mucinous carcinoma | 2 |
|
Medullary carcinoma | 1 |
|
Metaplastic carcinoma | 2 |
|
Invasive micropapillary
carcinoma | 1 |
| Spindle
cell carcinoma | 1 |
|
Others | 1 |
|
Missing | 2 |
| Subtype |
|
|
Luminal | 21 |
|
HER2 | 5 |
|
Triple-negative | 17 |
| Luminal
+ HER2 | 8 |
| Table III.Correlation between HMGB1 score and
pathological response. |
Table III.
Correlation between HMGB1 score and
pathological response.
| Before NAC |
|---|
|
|---|
| HMGB1 score | Grade 0, 1 | Grade 2, 3 |
|---|
| 0 | 23 | 18 |
| 1+ | 2 | 4 |
| 2+ | 1 | 2 |
| 3+ | 0 | 0 |
| P-value | 0.4648 |
|
| After
NAC |
|
| HMGB1
score | Grade 0,
1 | Grade 2,
3 |
|
| 0 | 5 | 8 |
| 1+ | 5 | 6 |
| 2+ | 8 | 6 |
| 3+ | 8 | 4 |
| P-value | 0.5076 |
| Table IV.Correlation between calreticulin
score and pathological response. |
Table IV.
Correlation between calreticulin
score and pathological response.
| Before NAC |
|---|
|
|---|
| Calreticulin
score | Grade 0, 1 | Grade 2, 3 |
|---|
| 0 | 0 | 0 |
| 1+ | 0 | 0 |
| 2+ | 18 | 17 |
| 3+ | 8 | 7 |
| P-value | 0.9017 |
|
|
| After
NAC |
|
| Calreticulin
score | Grade 0,
1 | Grade 2,
3 |
|
| 0 | 0 | 0 |
| 1+ | 0 | 0 |
| 2+ | 3 | 3 |
| 3+ | 23 | 21 |
| P-value | 0.9167 |
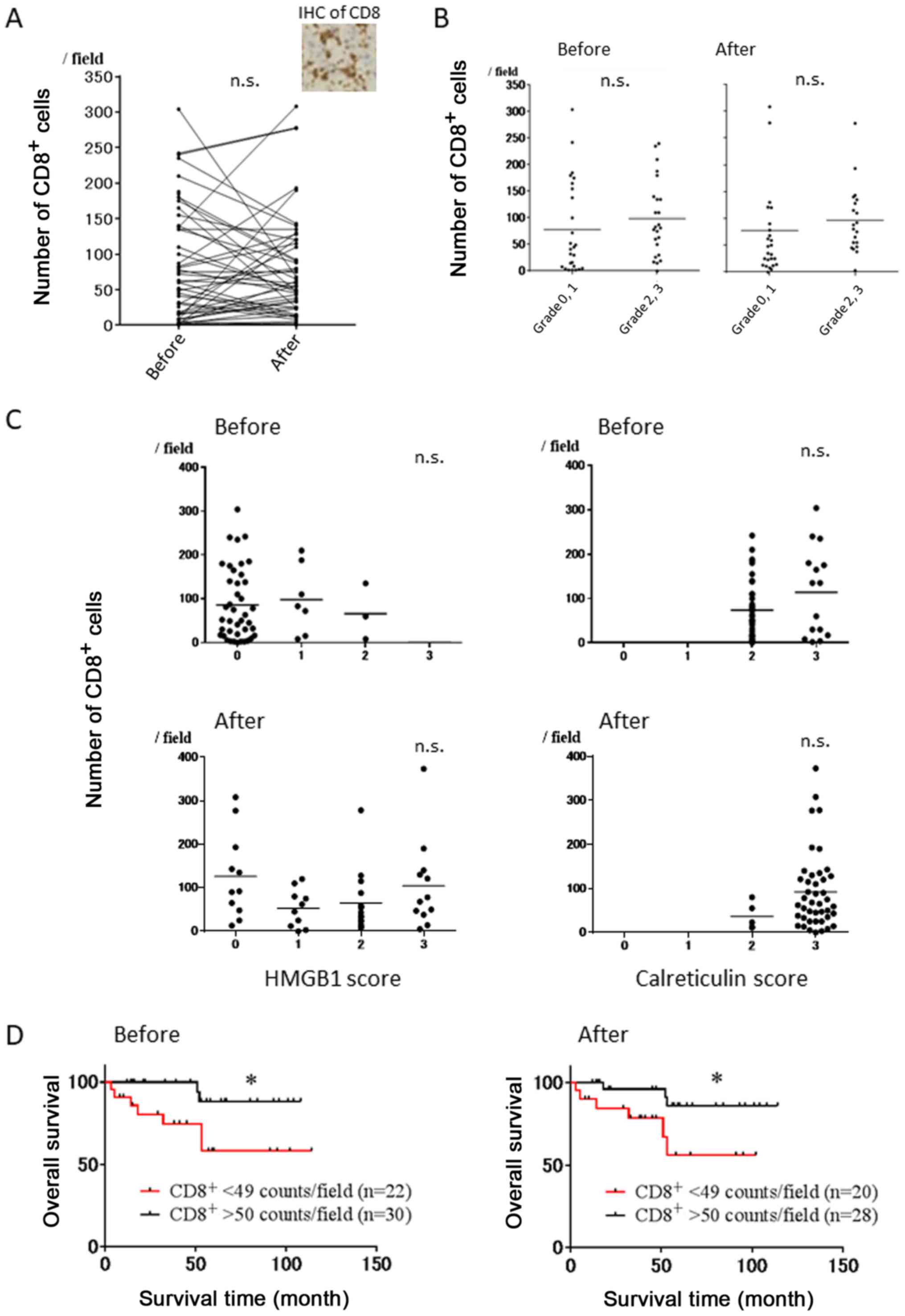
Correlation of infiltrating
CD8+ T cells before and after NAC with HMGB1 and
calreticulin score, and patient survival in breast cancer
patients
Representative immunostaining using CD8 mAb is shown
in Fig. 5A. There was no
significant correlation between the number of CD8+ T
cells before and after NAC (p=0.9228) (Fig. 5A). Furthermore, there were no
significant differences in the number of CD8+ T cells
before and after NAC between the cases with pathological grade 0
and 1 or those with grade 2 and 3 (Fig.
5B). We next evaluated the correlation between the number of
CD8+ T cells before/after NAC and HMGB1 score
before/after NAC, and no correlations were observed (Fig. 5C, left). Similarly, the number of
CD8+ T cells before/after NAC did not affect
calreticulin score before/after NAC (Fig. 5C, right). However, OS rates in the
high infiltration group of CD8+ T cells (>50
counts/field) before and after NAC were significantly superior to
those in the low infiltration group (<50 counts/field) (Fig. 5D).
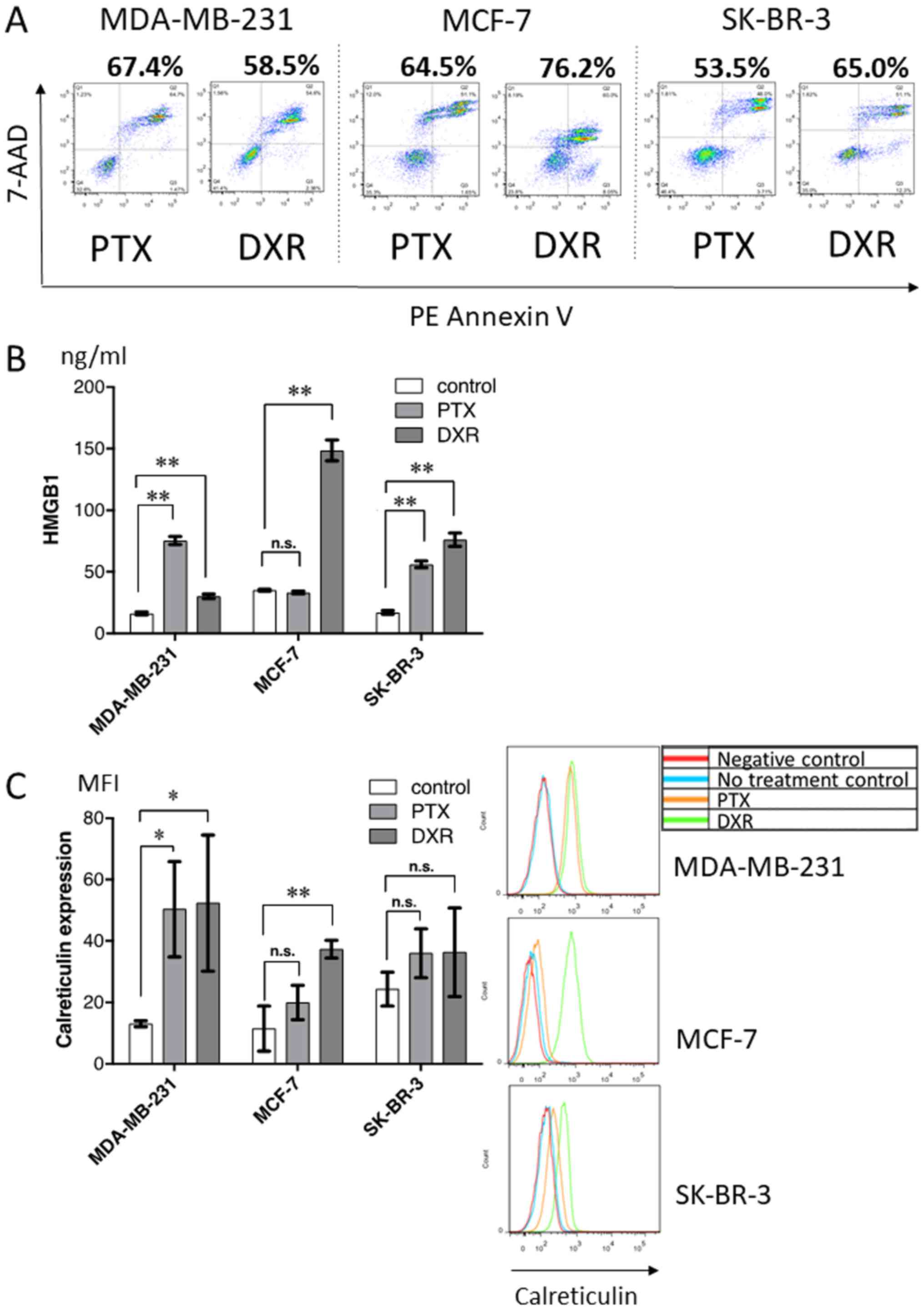
In vitro treatment of breast cancer
cell lines with chemotherapeutic drugs
To further evaluate HMGB1 and calreticulin
expression following treatment with chemotherapeutic drugs, three
breast tumor cell lines were treated with paclitaxel or doxorubicin
in vitro, and the production of HMGB1 and surface expression
of calreticulin, along with the proportion of dying cells (Fig. 6A), were analyzed. As shown in
Fig. 6B and C, chemotherapeutic
drugs alone induced variable levels of HMGB1 production (Fig. 6B) and surface calreticulin
expression (Fig. 6C) depending on
the drug and cell line, regardless of approximately the same
proportion of dying cells.
Discussion
The present study contains novel findings supporting
the concept that ICD can be induced by chemotherapy alone in
patients with breast cancer and ESCC. Firstly, both HMGB1 and
calreticulin expression were significantly upregulated after NAC.
Secondly, chemotherapeutic drugs induced the upregulation of HMGB1
and calreticulin in several tested breast cancer cell lines.
We and others have recently reported that danger
signals from dying cells following treatment with radiotherapy or
certain chemotherapeutic drugs, such as anthracyclines and
oxaliplatin, can induce Toll-like receptor (TLR)-dependent,
antigen-specific T-cell immunity (22,23).
Additional therapeutic modalities shown to induce ICD include
oncolytic virus therapy (24–26)
and photodynamic therapy (27,28).
Furthermore, among various danger signals released from dying cells
in a tumor-bearing mouse model, HMGB1, but not other known TLR4
ligands, could be a mandatory factor to induce tumor
antigen-specific T-cell immunity (22,23).
Moreover, it has been shown that early membrane exposure of
calreticulin induced by certain chemotherapeutic drugs, such as
anthracyclines and oxaliplatin (15,23,29–31),
could enhance the phagocytosis of dying tumor cells by DCs in
vitro (32–34). Both HMGB1 release and calreticulin
cell surface expression were found to be required for
antigen-specific T-cell response in a murine model. In the present
study, we showed for the first time in a human clinical study that
conventional chemotherapy alone significantly induced the
upregulation of HMGB1 and calreticulin in breast cancer and ESCC,
indicating that some degree of ICD was significantly induced in the
tumor microenvironment after chemotherapy. Contrary to
expectations, no correlation was observed between these expression
levels and the number of CD8+ T cells in the present
study, although OS in the high infiltration group of
CD8+ T cells was significantly superior to that in the
low infiltration group in breast cancer patients. In addition, the
present in vitro study indicated that there were substantial
variations in HMGB1 and calreticulin production following
chemotherapy in the breast cancer cell lines regardless of
approximately the same proportion of dying cells. Apetoh et
al reported that breast cancer patients with a TLR4
loss-of-function allele relapsed more quickly after chemotherapy
than those with a normal TLR4 allele (23). This finding indicates a clinically
relevant immune reaction triggered by TLR-dependent ICD induced by
chemotherapy, and the possibility of predicting clinical outcome
for CRT or chemotherapy by evaluating ICD status. However, in the
present study, there was no significant correlation between the
degree of mediators (HMGB1 and calreticulin) and pathological
response after NAC, or between the degree of the mediators and
patient survival. It is therefore likely that the induction of ICD
alone is not a suitable biomarker to predict the clinical response
to NAC, and a more complicated network is involved in ICD
induction. Regarding this problem, Bruchard et al, recently
reported that myeloid-derived suppressor cells (MDSCs) treated with
gemcitabine and 5-fluorouracil produced IL-1β, and MDSC-derived
IL-1β induced secretion of IL-17 by CD4-positive T cells, leading
to blunt the anticancer efficacy of the chemotherapy (35). Further investigation is required to
find a suitable biomarker for the response to NAC.
Taking our present and previous studies into
consideration (36), it is clear
that some degree of ICD was induced in cancer patients after CRT or
chemotherapy alone. Combination therapy of CRT or chemotherapy with
immune checkpoint inhibitors may therefore induce a synergistic
effect. It has been shown that the anti-CTLA4 antibody enhances the
antigen presentation of DCs activated by HMGB1 and calreticulin,
and that anti-programmed death 1/anti-programmed death ligand 1
antibodies enhance the cytotoxic ability of tumor antigen-specific
T-cells in the tumor microenvironment, resulting in the improvement
of patient survival (37,38).
In conclusion, the findings of the present study
indicate that chemotherapy alone can induce ICD in breast cancer
and ESCC patients. Hence, a combination therapy of chemotherapy
with immune checkpoint inhibitors may enhance the ICD, resulting in
the improvement of both the pathological response and survival of
breast cancer and ESCC patients.
Glossary
Abbreviations
Abbreviations:
|
CRT
|
chemo-radiotherapy
|
|
DCs
|
dendritic cells
|
|
ESCC
|
esophageal squamous cell carcinoma
|
|
HMGB1
|
high-mobility group box 1 protein
|
|
ICD
|
immunogenic tumor cell death
|
|
NAC
|
neoadjuvant chemotherapy
|
|
TLR
|
Toll-like receptor
|
References
|
1
|
Ishikura S, Nihei K, Ohtsu A, Boku N,
Hironaka S, Mera K, Muto M, Ogino T and Yoshida S: Long-term
toxicity after definitive chemoradiotherapy for squamous cell
carcinoma of the thoracic esophagus. J Clin Oncol. 21:2697–2702.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Manzoni G, Pedrazzani C, Laterza E,
Pasini F, Grandinetti A, Bernini M, Ruzzenente A, Zerman G,
Tomezzoli A and Cordiano C: Induction chemoradiotherapy for
squamous cell carcinoma of the thoracic esophagus: Impact of
increased dosage on long-term results. Ann Thorac Surg.
80:1176–1183. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Early Breast Cancer Trialists’
Collaborative Group (EBCTCG), . Effects of chemotherapy and
hormonal therapy for early breast cancer on recurrence and 15-year
survival: An overview of the randomised trials. Lancet.
365:1687–1717. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wolmark N, Wang J, Mamounas E, Bryant J
and Fisher B: Preoperative chemotherapy in patients with operable
breast cancer: Nine-year results from National Surgical Adjuvant
Breast and Bowel Project B-18. J Natl Cancer Inst Monogr.
30:96–102. 2001. View Article : Google Scholar
|
|
5
|
Bear HD, Anderson S, Brown A, Smith R,
Mamounas EP, Fisher B, Margolese R, Theoret H, Soran A, Wickerham
DL, et al National Surgical Adjuvant Breast and Bowel Project
Protocol B-27, : The effect on tumor response of adding sequential
preoperative docetaxel to preoperative doxorubicin and
cyclophosphamide: Preliminary results from National Surgical
Adjuvant Breast and Bowel Project Protocol B-27. J Clin Oncol.
21:4165–4174. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rastogi P, Anderson SJ, Bear HD, Geyer CE,
Kahlenberg MS, Robidoux A, Margolese RG, Hoehn JL, Vogel VG, Dakhil
SR, et al: Preoperative chemotherapy: Updates of National Surgical
Adjuvant Breast and Bowel Project Protocols B-18 and B-27. J Clin
Oncol. 26:778–785. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kono K and Mimura K: Immunogenic tumor
cell death induced by chemoradiotherapy in a clinical setting.
OncoImmunology. 2:e221972013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kono K, Mimura K and Kiessling R:
Immunogenic tumor cell death induced by chemoradiotherapy:
Molecular mechanisms and a clinical translation. Cell Death Dis.
4:e6882013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kroemer G, Galluzzi L, Kepp O and Zitvogel
L: Immunogenic cell death in cancer therapy. Annu Rev Immunol.
31:51–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Krysko DV, Garg AD, Kaczmarek A, Krysko O,
Agostinis P and Vandenabeele P: Immunogenic cell death and DAMPs in
cancer therapy. Nat Rev Cancer. 12:860–875. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Krysko O, Løve Aaes T, Bachert C,
Vandenabeele P and Krysko DV: Many faces of DAMPs in cancer
therapy. Cell Death Dis. 4:e6312013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ladoire S, Enot D, Andre F, Zitvogel L and
Kroemer G: Immunogenic cell death-related biomarkers: Impact on the
survival of breast cancer patients after adjuvant chemotherapy.
OncoImmunology. 5:e10827062015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Stoll G, Enot D, Mlecnik B, Galon J,
Zitvogel L and Kroemer G: Immune-related gene signatures predict
the outcome of neoadjuvant chemotherapy. OncoImmunology.
3:e278842014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gebremeskel S and Johnston B: Concepts and
mechanisms underlying chemotherapy induced immunogenic cell death:
Impact on clinical studies and considerations for combined
therapies. Oncotarget. 6:41600–41619. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Casares N, Pequignot MO, Tesniere A,
Ghiringhelli F, Roux S, Chaput N, Schmitt E, Hamai A, Hervas-Stubbs
S, Obeid M, et al: Caspase-dependent immunogenicity of
doxorubicin-induced tumor cell death. J Exp Med. 202:1691–1701.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wong DY, Ong WW and Ang WH: Induction of
immunogenic cell death by chemotherapeutic platinum complexes.
Angew Chem Int Ed Engl. 54:6483–6487. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Galluzzi L, Buqué A, Kepp O, Zitvogel L
and Kroemer G: Immunological Effects of conventional chemotherapy
and targeted anticancer agents. Cancer Cell. 28:690–714. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hodge JW, Garnett CT, Farsaci B, Palena C,
Tsang KY, Ferrone S and Gameiro SR: Chemotherapy-induced
immunogenic modulation of tumor cells enhances killing by cytotoxic
T lymphocytes and is distinct from immunogenic cell death. Int J
Cancer. 133:624–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kepp O, Senovilla L, Vitale I, Vacchelli
E, Adjemian S, Agostinis P, Apetoh L, Aranda F, Barnaba V, Bloy N,
et al: Consensus guidelines for the detection of immunogenic cell
death. OncoImmunology. 3:e9556912014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okano M, Kumamoto K, Saito M, Onozawa H,
Saito K, Abe N, Ohtake T and Takenoshita S: Upregulated Annexin A1
promotes cellular invasion in triple-negative breast cancer. Oncol
Rep. 33:1064–1070. 2015.PubMed/NCBI
|
|
21
|
Masafumi K, Sadako A, Futoshi A, Y K, H M
and S N: Histological criteria for assessment of therapeutic
response in breast cancer (2007 version). Breast Cancer. 15:5–7.
2007.
|
|
22
|
Apetoh L, Ghiringhelli F, Tesniere A,
Criollo A, Ortiz C, Lidereau R, Mariette C, Chaput N, Mira JP,
Delaloge S, et al: The interaction between HMGB1 and TLR4 dictates
the outcome of anticancer chemotherapy and radiotherapy. Immunol
Rev. 220:47–59. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Apetoh L, Ghiringhelli F, Tesniere A,
Obeid M, Ortiz C, Criollo A, Mignot G, Maiuri MC, Ullrich E,
Saulnier P, et al: Toll-like receptor 4-dependent contribution of
the immune system to anticancer chemotherapy and radiotherapy. Nat
Med. 13:1050–1059. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miyamoto S, Inoue H, Nakamura T, Yamada M,
Sakamoto C, Urata Y, Okazaki T, Marumoto T, Takahashi A, Takayama
K, et al: Coxsackievirus B3 is an oncolytic virus with
immunostimulatory properties that is active against lung
adenocarcinoma. Cancer Res. 72:2609–2621. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Diaconu I, Cerullo V, Hirvinen ML,
Escutenaire S, Ugolini M, Pesonen SK, Bramante S, Parviainen S,
Kanerva A, Loskog AS, et al: Immune response is an important aspect
of the antitumor effect produced by a CD40L-encoding oncolytic
adenovirus. Cancer Res. 72:2327–2338. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Takasu A, Masui A, Hamada M, Imai T, Iwai
S and Yura Y: Immunogenic cell death by oncolytic herpes simplex
virus type 1 in squamous cell carcinoma cells. Cancer Gene Ther.
23:107–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Garg AD, Krysko DV, Verfaillie T,
Kaczmarek A, Ferreira GB, Marysael T, Rubio N, Firczuk M, Mathieu
C, Roebroek AJ, et al: A novel pathway combining calreticulin
exposure and ATP secretion in immunogenic cancer cell death. EMBO
J. 31:1062–1079. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanaka M, Kataoka H, Yano S, Sawada T,
Akashi H, Inoue M, Suzuki S, Inagaki Y, Hayashi N, Nishie H, et al:
Immunogenic cell death due to a new photodynamic therapy (PDT) with
glycoconjugated chlorin (G-chlorin). Oncotarget. 7:47242–47251.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Michaud M, Martins I, Sukkurwala AQ,
Adjemian S, Ma Y, Pellegatti P, Shen S, Kepp O, Scoazec M, Mignot
G, et al: Autophagy-dependent anticancer immune responses induced
by chemotherapeutic agents in mice. Science. 334:1573–1577. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zappasodi R, Pupa SM, Ghedini GC,
Bongarzone I, Magni M, Cabras AD, Colombo MP, Carlo-Stella C,
Gianni AM and Di Nicola M: Improved clinical outcome in indolent
B-cell lymphoma patients vaccinated with autologous tumor cells
experiencing immunogenic death. Cancer Res. 70:9062–9072. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fucikova J, Kralikova P, Fialova A,
Brtnicky T, Rob L, Bartunkova J and Spísek R: Human tumor cells
killed by anthracyclines induce a tumor-specific immune response.
Cancer Res. 71:4821–4833. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tesniere A, Schlemmer F, Boige V, Kepp O,
Martins I, Ghiringhelli F, Aymeric L, Michaud M, Apetoh L, Barault
L, et al: Immunogenic death of colon cancer cells treated with
oxaliplatin. Oncogene. 29:482–491. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zitvogel L, Kepp O, Senovilla L, Menger L,
Chaput N and Kroemer G: Immunogenic tumor cell death for optimal
anticancer therapy: The calreticulin exposure pathway. Clin Cancer
Res. 16:3100–3104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Obeid M, Tesniere A, Ghiringhelli F, Fimia
GM, Apetoh L, Perfettini JL, Castedo M, Mignot G, Panaretakis T,
Casares N, et al: Calreticulin exposure dictates the immunogenicity
of cancer cell death. Nat Med. 13:54–61. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bruchard M, Mignot G, Derangère V, Chalmin
F, Chevriaux A, Végran F, Boireau W, Simon B, Ryffel B, Connat JL,
et al: Chemotherapy-triggered cathepsin B release in
myeloid-derived suppressor cells activates the Nlrp3 inflammasome
and promotes tumor growth. Nat Med. 19:57–64. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Suzuki Y, Mimura K, Yoshimoto Y, Watanabe
M, Ohkubo Y, Izawa S, Murata K, Fujii H, Nakano T and Kono K:
Immunogenic tumor cell death induced by chemoradiotherapy in
patients with esophageal squamous cell carcinoma. Cancer Res.
72:3967–3976. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tumeh PC, Harview CL, Yearley JH, Shintaku
IP, Taylor EJ, Robert L, Chmielowski B, Spasic M, Henry G, Ciobanu
V, et al: PD-1 blockade induces responses by inhibiting adaptive
immune resistance. Nature. 515:568–571. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Twyman-Saint Victor C, Rech AJ, Maity A,
Rengan R, Pauken KE, Stelekati E, Benci JL, Xu B, Dada H, Odorizzi
PM, et al: Radiation and dual checkpoint blockade activate
non-redundant immune mechanisms in cancer. Nature. 520:373–377.
2015. View Article : Google Scholar : PubMed/NCBI
|