Introduction
Pancreatic cancer (PC) is a malignant tumor with a
high mortality rate and has become the leading cause of
cancer-related death (1). At
present, the prognosis of patients with PC remains poor and the
5-year survival rate of PC patients is still low (2,3).
Therefore, it is urgent to understand the molecular mechanism of PC
carcinogenesis in order to unearth reliable diagnostic and
therapeutic targets for PC.
Long non-coding RNAs (lncRNAs) are
non-protein-coding transcripts longer than 200 nucleotides and
serve important roles in tumorigenesis (4,5). In
addition, mounting evidence has shown that lncRNAs are functional
in every stage of tumor progression (6), and are involved in tumor growth,
invasion, metastasis of multiple cancers including PC (7–9).
Previous studies have shown that the lncRNAs ATB, MALAT1, HOTAIR1,
H19 and HOTTIP are associated with the development and progression
of PC (10–14). lncRNA X inactive-specific transcript
(XIST) (18 kb) is important for inactivation of X chromosome in the
development of female mammals and is a prototype of gene-silencing
lncRNAs (15). Studies have
indicated that lncRNA XIST is involved in the progression of
glioblastoma, non-small cell lung and gastric cancer, and
hepatocellular carcinoma (16–20).
However, the role of lncRNA XIST in PC is not fully understood.
MicroRNAs (miRNAs) are small (~20 nucleotides in
length) non-coding RNA molecules that regulate gene expression by
inhibiting translation or degrading mRNA transcripts (21). Research has shown that miRNAs
participate in the occurrence and development of various diseases
(22). There is a growing body of
research showing that miRNAs are involved in the regulation of
various biological processes including proliferation,
differentiation, apoptosis, migration and invasion (23–26).
Therefore, miRNAs are new and effective biomarkers for human cancer
diagnostics. MicroRNA-34a-5p (miR-34a-5p) plays critical roles in
the progression of various diseases. However, the biological
functions and molecular mechanisms of miR-34a-5p in PC are not
entirely clear.
Recently, in molecular biology, competing endogenous
RNAs (abbreviated ceRNAs) regulate other RNA transcripts by
competing for shared microRNAs. This has been proposed and defined
as the crosstalk of RNA transcripts with miRNA response elements
(MREs) (27,28). An increasing number of studies have
found that the interaction between lncRNAs and miRNAs occurs in
many different types of cancers (29–32),
adding more puzzles for miRNA and lncRNA regulatory networks. For
example, lncRNA-UCA1 was found to serve as a ceRNA, promoting the
cell proliferation of esophageal cancer (33); lncRNA H19 functions as a ceRNA to
promote epithelial-to-mesenchymal transition of colorectal cancer
(30); lncRNA HOTAIR serves as a
ceRNA to regulate HER2 expression by targeting miR-331-3p in
gastric cancer (34); lncRNA FER1L4
inhibits cancer cell proliferation by functioning as a ceRNA
(35).
In the present study, we investigated the expression
and function of XIST in PC. We found that XIST expression was
markedly upregulated in PC tissues and cells. Overexpression of
XIST promoted PC cell growth, migration, invasion and metastasis;
knockdown of XIST suppressed PC cell growth, migration, invasion
and metastasis, implying a possible role of lncRNA XIST as an
oncogene in PC. Moreover, we demonstrated that XIST was involved in
the proliferation, migration, invasion and progression of PC by
targeting miR-34a-5p.
Materials and methods
Ethics statement and specimens
Paired PC (n=139) and corresponding non-tumor
control tissues were collected from the Japan Union Hospital of
Jilin University from 2013 to 2015. The present study was approved
by the Ethics Committee of Japan Union Hospital of Jilin
University, and written informed consent was obtained from all
patients. All tissue samples were immediately conserved at −80°C
after washing with sterile phosphate-buffered saline (PBS).
Cell culture
Six PC cell lines (PANC-1, ASPC-1, MIA PaCa-2, HPAC,
CFAPC-1 and BxPC-3), and human pancreatic ductal epithelial (HPDE),
and 293T cells were purchased from the Institute of Biochemistry
and Cell Biology of the Chinese Academy of Sciences (Shanghai,
China). Cells were grown in Dulbecco's modified Eagles medium
(DMEM) supplemented with 10% fetal bovine serum (FBS) (both from
Invitrogen Life Technologies, Carlsbad, CA, USA) and 100 U/ml
penicillin and 100 µg/ml streptomycin in an atmosphere of 5%
CO2 at 37°C.
Lentiviral vector construction
Human XIST full-length DNAs were amplified by PCR
from mRNA of BxPC-3 cells. Then, the cDNAs were inserted into a
pcDNA3.1 vector. Enhanced green fluorescent protein (EGFP) served
as a control. Specific shRNA against XIST and luciferase shRNA
(Luc-shRNA; control) were synthesized and validated effective by
RiboBio Co. (Guangzhou, China). The VSV-G pseudotyped lentiviruses
were produced by co-transfecting 293T cells with lentivirus
expression plasmid and packaging plasmids (pMD2.G, pMDL-G/P-RRE and
pRSV-REV). Cells (5×104) were then transduced with the
lentiviruses in the presence of 8 µg/ml Polybrene (Sigma-Aldrich,
St. Louis, MO, USA).
Cell transfection
For plasmid transfection, BxPC-3 cells
(2×105 cells/well) were seeded in 6-well plates, and
transfected with XIST or the control using Lipofectamine™ 3000
(Invitrogen, Carlsbad, CA, USA) respectively according to the
manufacturer's protocols. Likewise, PANC-1 cells were transfected
with shXIST or control, respectively.
For miRNA transfection, miR-34a-5p mimics, inhibitor
and negative control (miR-34a-5p NC) were purchased from GenePharma
Co., Ltd. (Shanghai, China). Cells were transfected with miR-34a-5p
mimics, miR-34a-5p inhibitors and miR-34a-5p NC respectively using
Lipofectamine™ 3000, respectively, according to the manufacturer's
protocols.
RNA isolation and quantitative
real-time PCR (qRT-PCR)
Total RNA from PC tissues or cells was isolated
using TRIzol reagent (Invitrogen) after treatments. Then, reverse
transcription reaction was performed with Revert Aid First Strand
cDNA Synthesis kit (Thermo Fisher Scientific Inc., Rockford, IL,
USA) and random primers to provide cDNA products. The thermocycling
conditions were 25°C for 5 min; 42°C for 60 min; 70°C for 10 min
for the reverse transcription. qRT-PCR assays were performed using
SYBR Premix Ex Taq (Takara, Tokyo, Japan), primers and cDNA
templates on the Applied Biosystems 7500 Real-Time PCR system
[Applied Biosystems Inc. (ABI) Carlsbad, CA, USA). Each individual
sample was performed in triplicate and the expression levels were
quantified using the comparative cycle threshold (CT) method.
Results were normalized to GAPDH expression and RNA enrichments
were calculated using the equation 2−ΔΔCt (36). Specific primers for XIST and
miR-34a-5p were designed and synthesized by RiboBio Co. The primer
sequences used in the present study are shown in Table I.
 | Table I.Primer sequences for qRT-PCR
analysis. |
Table I.
Primer sequences for qRT-PCR
analysis.
| Gene name | Primer
sequences |
|---|
| miR-34a-5p | Forward
5′-GGGGTGGCAGTGTCTTAGC-3′ |
|
| Reverse
5′-CAGTGCGTGTCGTGGAGT-3′ |
| U6 | Forward
5′-CTCGCTTCGGCAGCACA-3′ |
|
| Reverse
5′-AACGCTTCACGAATTTGCGT-3′ |
| XIST | Forward
5′-AGCTCCTCGGACAGCTGTAA-3′ |
|
| Reverse
5′-CTCCAGATAGCTGGCAACC-3′ |
| GAPDH | Forward
5′-TGTTCGTCATGGGTGTGAAC-3′ |
|
| Reverse
5′-ATGGCATGGACTGTGGTCAT-3′ |
Luciferase reporter assay
293T cells (1×105 cells/well) were placed
in a 24-well plate, and co-transfected 200 ng of either
pGL3-XIST-wt or pGL3-XIST-Mut vector and 80 ng of either miR-34a-5p
or miR-NC. After 48 h for transfection, cells were harvested and
luciferase activities were measured with the Dual-Luciferase
Reporter Assay System (Promega, Wisconsin, WI, USA). All
transfection experiments were conducted in triplicate.
Cell viability
Cell viability was assessed by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT)
assay. The treated cells (4,000 cells/well) were seeded in a
96-well plate and cultured in complete medium for 1, 2, 3, 4 and 5
days. MTT (0.5 mg/ml) was added to each well and incubation was
carried out at 37°C for 4 h. Afterward, the supernatant was
carefully aspirated and 100 µl of dimethyl sulfoxide (DMSO) was
added. The absorbance was measured at 490 nm using a microplate
reader. All experiments were repeated ≥3 times.
Colony formation assay
For the colony formation assay, the treated BxPC-3
and PANC-1 cells were seeded in 6-well plates at a density of 500
cells/well after transfection with different vectors. After 14
days, the cells were fixed with a 4% paraformaldehyde solution and
stained with crystal violet. The total number of colonies in each
plate from three independent transfections was counted under an
inverted microscope to evaluate the colony formation ability.
Wound-healing assay
BxPC-3 or PANC-1 cells were transfected with
different vectors and seeded in 6-well plates. Small linear wounds
were created by removing a line of cells with a disinfected
Eppendorf tip. After removing cell debris by washing with FBS-free
medium, the wound areas were photographed under a microscope. Three
different positions of distance between the two edges of the wound
were calculated and analyzed by image analysis software (National
Institute of Health, Bethesda, MD, USA).
Migration and invasion assays
Cell migration and invasion assays were performed
using the Transwell assay according to the manufacturer's
instructions. BxPC-3 or PANC-1 cells (5×104 cells/well)
were transfected with different vectors and seeded in the upper
compartment of the Transwell and incubated in serum-free media, and
the lower compartment was filled with complete medium supplemented
with 10% FBS. After 48 h of incubation at 37°C, non-invading cells
were removed. The migratory and invasive cells on the bottom
surface of the filters were fixed using 4% paraformaldehyde, and
stained with 0.1% crystal violet solution. Four randomly selected
fields of the fixed cells were counted for each group. The
experiments were performed in triplicate.
Flow cytometric analysis
Treated cells were washed with 1X PBS, trypsinized
and fixed with 70% ethanol for 30 min on ice. RNA was degraded with
20 mg/ml RNase (Sigma-Aldrich) for 1 h at 37°C. DNA was then
labeled with 20 mg/ml propidium iodide (PI; Sigma-Aldrich). The
cell cycle images were obtained and analyzed using FACSCalibur (BD
Biosciences, Franklin Lakes, NJ, USA) and FlowJo software (Tree
Star, Inc., Ashland, OR, USA).
Tumor formation assay in nude
mice
The BxPC-3 cells were transfected with control
plasmid or XIST plasmid. The PANC-1 cells were infected with
control or shXIST plasmid, respectively. Four-week-old male nude
mice were purchased from the National Laboratory Animal Center
(Shanghai, China) and divided into four groups for subcutaneous
injection using BxPC-3 or PANC-1 cells. Animals were sacrificed 40
days after injection and tumors were collected for measurement of
the volume every 10 days. The tumor volume (V) was calculated by
the formula: V (mm3) = length × width2/2. All
experiments were performed strictly in accordance with a protocol
approved by the Administrative Panel on Laboratory Animal Care of
Jilin University.
Statistical analysis
The data were analyzed by the Student's t-test and
one-way analysis of variance (ANOVA) using SPSS 15.0 software
(SPSS, Inc., Chicago, IL, USA). Pearson's correlation coefficient
was used to calculate the correlation between miR-34a-5p and XIST.
The Kaplan-Meier method test was utilized for survival analysis.
Each experiment was repeated at least three times. All results were
summarized and are presented as means ± SD. P<0.05 was
considered statistically significant.
Results
Upregulation of lncRNA-XIST predicts a
poor prognosis in PC patients
In the present study, we determined lncRNA-XIST
expression in 139 PC patients and analyzed the relationship between
XIST expression and clinicopathological characteristics [age, sex,
lymphatic metastasis, distal metastasis and TNM stage] of the PC
patients. The results showed that lncRNA-XIST was significantly
upregulated in tumor tissues compared with that noted in the
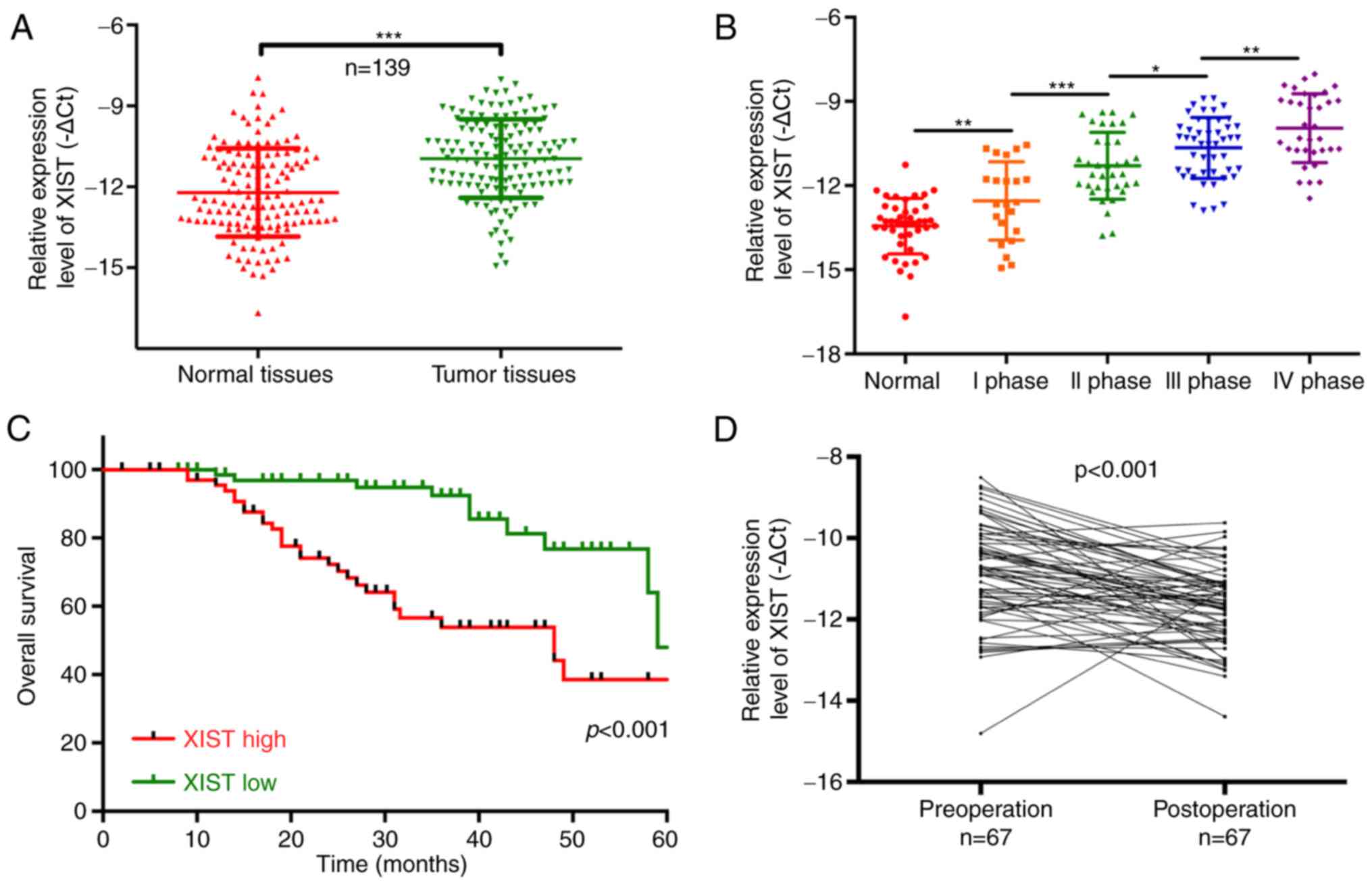
matched adjacent normal tissues (P<0.001; Fig. 1A). We also found that lncRNA-XIST
expression was significantly related to the tumor-node-metastasis
(TNM) stage, and was higher in the TNM I, II, III and IV stages
than that noted in the normal group (P<0.05, P<0.01,
P<0.001; Fig. 1B). Therefore,
our results indicated that lncRNA-XIST expression is correlated
with the malignant degree of PC. The Chi-square analysis indicated
that the expression level of lncRNA-XIST was positively correlated
with TNM stage (P<0.05) and distal metastasis (P<0.01),
suggesting that lncRNA-XIST may be a potential biomarker for PC
(Table II). In addition, PC
patients with a high expression of XIST had a shorter overall
survival than patients with low XIST expression (P<0.001;
Fig. 1C). The results also
indicated that lncRNA-XIST expression was significantly
downregulated in 67 post-operation patients compared with
pre-operation patients (P<0.001; Fig. 1D). All the above results
demonstrated that a high expression level of XIST is associated
with poor prognosis.
| Table II.Relationship between lncRNA-XIST
expression level (∆Ct) and clinicopathological characteristics of
the PC patients. |
Table II.
Relationship between lncRNA-XIST
expression level (∆Ct) and clinicopathological characteristics of
the PC patients.
|
|
| lncRNA-XIST |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | n (%) | Mean ± SD | P-value |
|---|
| Total no. of
patients | 139 |
|
|
| Age (years) |
|
| 0.101 |
|
>60 | 59 (42.4) | 11.85±1.73 |
|
|
≤60 | 80 (57.6) | 11.19±2.69 |
|
| Sex |
|
| 0.232 |
|
Male | 89 (64.0) | 11.59±1.75 |
|
|
Female | 50 (36.0) | 12.02±2.45 |
|
| Lymphatic
metastasis |
|
| 0.235 |
| N0 | 94 (67.6) | 12.05±1.49 |
|
|
N1-N2 | 45 (32.4) | 11.71±1.73 |
|
| Distal
metastasis |
|
|
0.003b |
| M0 | 105 (75.5) | 11.98±1.06 |
|
| M1 | 34 (24.5) | 11.14±2.16 |
|
| TNM stage |
|
|
0.012a |
| 0, I,
II | 91 (65.5) | 11.72±1.58 |
|
| III,
IV | 48 (34.5) | 10.96±1.83 |
|
XIST promotes PC tumor cell growth in
vitro
To further explore the oncogenic roles of XIST on PC
in vitro, the expression level of lncRNA-XIST was detected
by qRT-PCR in human pancreatic ductal epithelial (HPDE) cells and
PC cell lines (PANC-1, ASPC-1, MIA PaCa-2, HPAC, CFAPC-1, and
BxPC-3). The results revealed that lncRNA-XIST expression was
increased in the PC cell lines compared with that noted in the HPDE
cells (P<0.05, P<0.01, P<0.001; Fig. 2A). According to the lncRNA-XIST
expression level in PC cells, we established PC cell lines (BxPC-3
and PANC-1) with XIST stable overexpression or knockdown (shXIST),
respectively. The results indicated that lncRNA-XIST may play a
critical role in the progression of PC. As shown in Fig. 2B, the expression level of XIST was
increased in the BxPC-3 cells transfected with XIST-overexpressing
plasmid compared with the control. XIST expression was decreased in
PANC-1 cells transfected with shXIST compared with the control.
Overexpression of XIST significantly promoted BxPC-3 cell
viability, and knockdown of XIST significantly inhibited PANC-1
cell viability (P<0.01, P<0.001; Fig. 2C and D). Then, we evaluated the
effect of XIST on cell cycle distribution of the PC cells. In
agreement with the above results, we found that overexpression of
XIST observably decreased the percentage of cells in the G0/G1
stage and increased the percentage of cells in the S and G2/M
stages; knockdown of XIST significantly induced cell cycle arrest
in the G0/G1 stage and decreased the percentage of cells in the S
and G2/M stages (P<0.05, P<0.01; Fig. 2E).
 | Figure 2.XIST promotes PC cell growth in
vitro. (A) Expression level of lncRNA-XIST was detected by
qRT-PCR assay in human pancreatic ductal epithelial (HPDE) cells
and PC cell lines (PANC-1, ASPC-1, MIA PaCa-2, HPAC, CFAPC-1 and
BxPC-3). Each assay was performed for at least three biological
replicates (*P<0.05, **P<0.01, ***P<0.001). (B) XIST
expression was examined by qRT-PCR assay in BxPC-3 cells
transfected with XIST plasmid and control, and PANC-1 cells
transfected with shXIST and control. Relative expression was
normalized to GAPDH expression (**P<0.01). (C) MTT assay was
performed to measure the viability of BxPC-3 and PANC-1 cells
treated as in B (**P<0.01). (D) Colony formation assay was used
to detect the proliferation ability of the transfected cells. (E)
Cell Cycle distribution was analyzed by flow cytometry in BxPC-3
and PANC-1 cells after transfection (**P<0.01, ***P<0.001).
PC, pancreatic cancer; lncRNA, long non-coding RNA; XIST, X
inactive-specific transcript. |
XIST accelerates the migration and
invasion abilities of PC
We further studied whether XIST functions as an
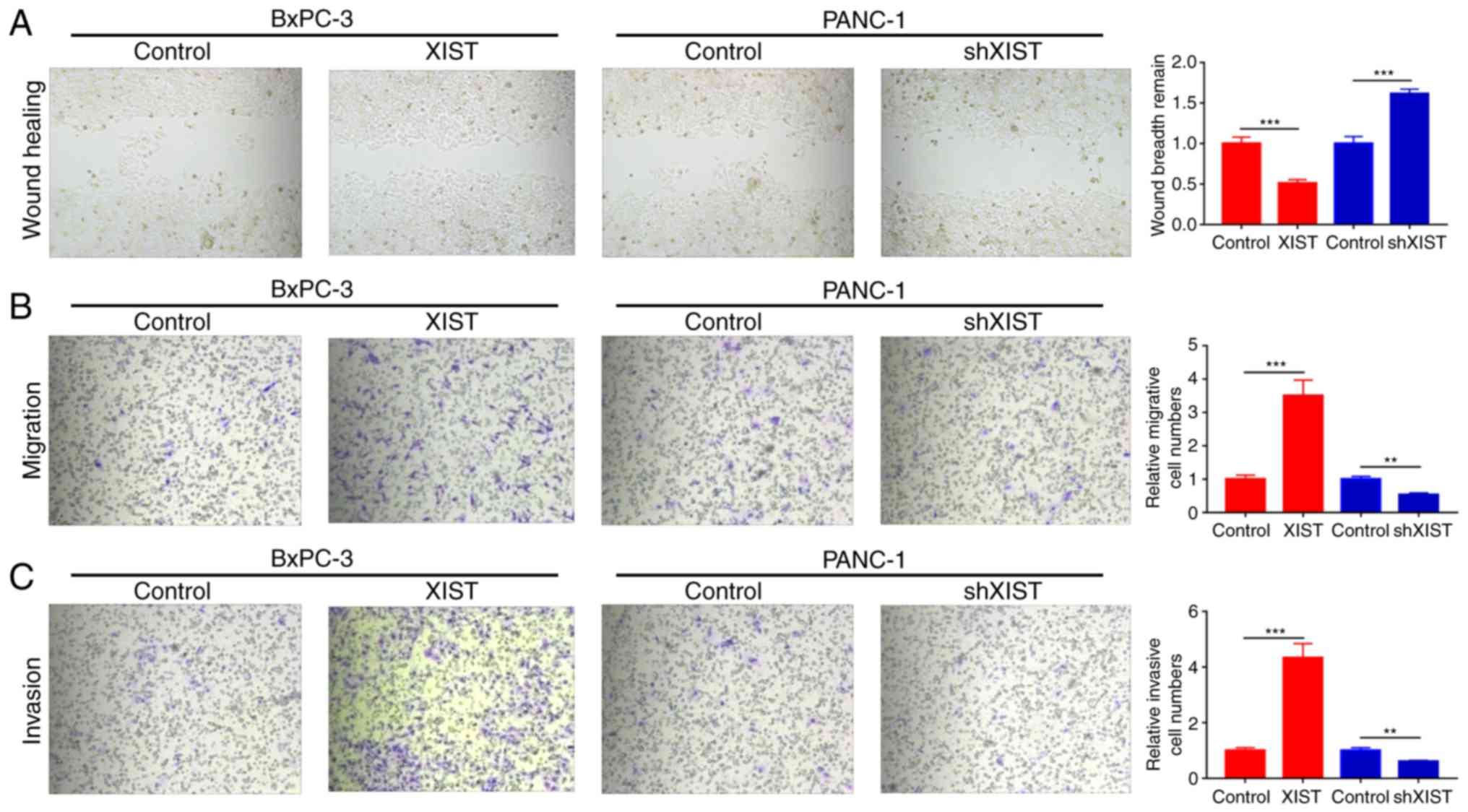
oncogene. Wound healing assays found that overexpression of XIST
significantly promoted the wound healing ability and knockdown of
XIST significantly decreased this ability (P<0.001; Fig. 3A). Moreover, we used Transwell assay
to explore the effects of XIST on the migration and invasion of
BxPC-3 and PANC-1 cells. The results indicated that XIST obviously
promoted the migration and invasion abilities of the BxPC-3 cells;
knockdown of XIST markedly inhibited the migration and invasion
abilities of the PANC-1 cells (P<0.01, P<0.001; Fig. 3B and C).
XIST promotes tumor formation in
vivo
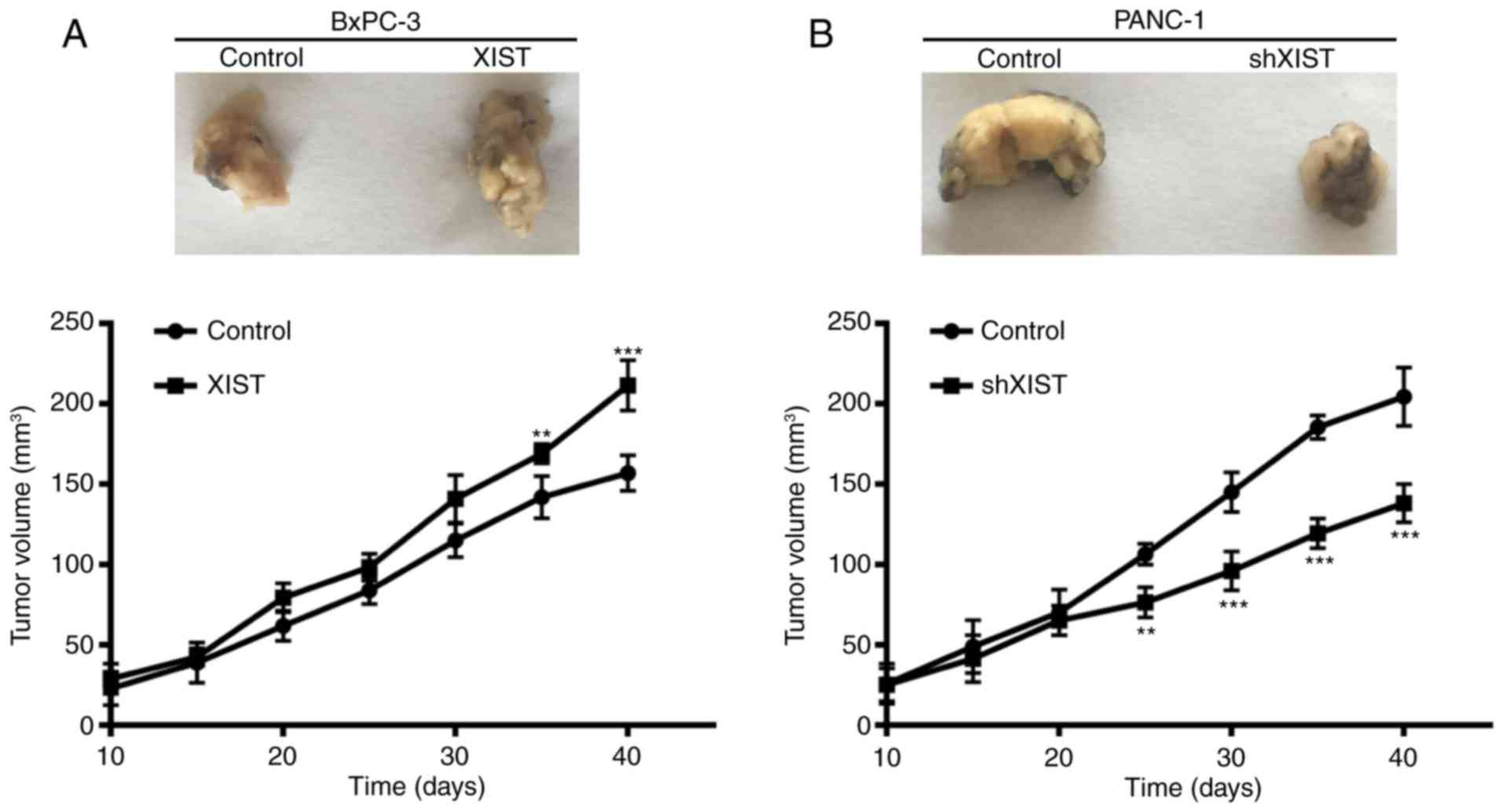
To further assess the growth effect of XIST on PC,
we examined the tumorigenicity in nude mice. The result showed that
overexpression of XIST in BxPC-3 cells accelerated tumor growth in
the nude mouse model, and knockdown of XIST in PANC-1 cells
inhibited tumor growth in the nude mouse model (P<0.01,
P<0.001; Fig. 4). At 10, 15, 20,
25, 30, 35 and 40 days after injection, the tumors were removed.
The tumor volume was accordance with above observation. From the
above results, we suggest that XIST is an oncogene and promotes
tumor formation.
Downregulation of miR-34a-5p predicts
a poor prognosis in PC patients
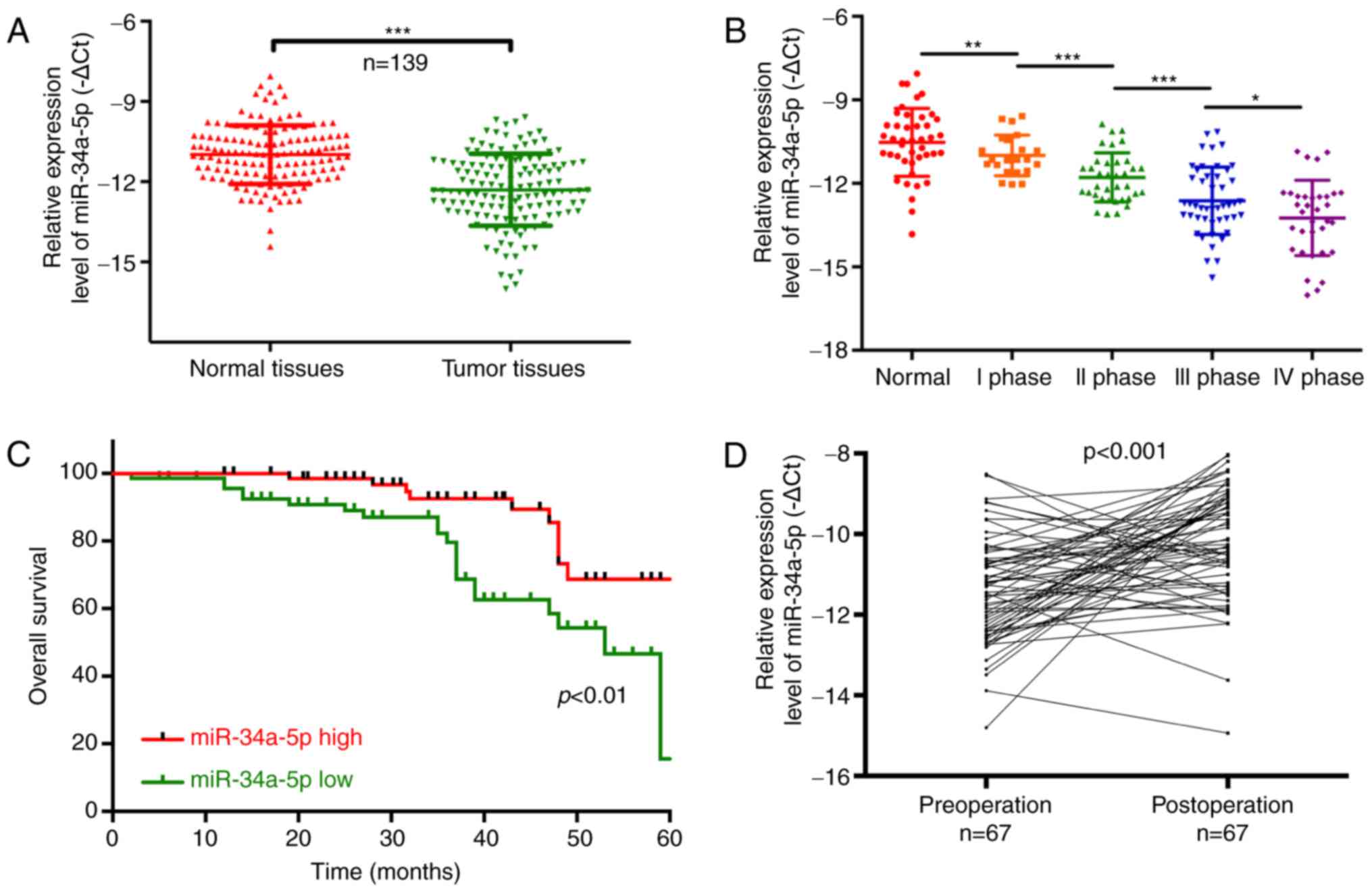
Similarly, we found that miR-34a-5p was
significantly decreased in tumor tissues compared with that noted
in the matched adjacent normal tissues (P<0.001; Fig. 5A). The results also showed that
miR-34a-5p expression was significantly related to TNM stage, and
was lower in the TNM I, II, III and IV stage than that in the
normal group (P<0.05, P<0.01, P<0.001, Fig. 5B). The Chi-square analysis also
indicated that miR-34a-5p expression was related to lymphatic
metastasis (P=0.006) and TNM stage (P=0.009), suggesting that
miR-34a-5p may be a potential biomarker for PC (Table III). Furthermore, high-expression
of miR-34a-5p in patients with PC had a longer overall survival
than patients with low miR-34a-5p expression (P<0.01; Fig. 5C). miR-34a-5p expression was
significantly increased in 67 post-operation patients compared with
that noted in pre-operation patients (P<0.001; Fig. 5D).
| Table III.Correlation between miR-34a-5p
expression level (∆Ct) and clinicopathological characteristics of
the PC patients. |
Table III.
Correlation between miR-34a-5p
expression level (∆Ct) and clinicopathological characteristics of
the PC patients.
|
|
| miR-34a-5p |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | n (%) | Mean ± SD | P-value |
|---|
| Total no. of
patients | 139 |
|
|
| Age (years) |
|
| 0.785 |
|
>60 | 59 (42.4) | 11.48±1.75 |
|
|
≤60 | 80 (57.6) | 11.39±2.03 |
|
| Sex |
|
| 0.202 |
|
Male | 89 (64.0) | 11.45±1.79 |
|
|
Female | 50 (36.0) | 11.04±1.84 |
|
| Lymphatic
metastasis |
|
|
0.006a |
| N0 | 94 (67.6) | 11.06±1.64 |
|
|
N1-N2 | 45 (32.4) | 11.91±2.52 |
|
| Distal
metastasis |
|
| 0.089 |
| M0 | 105 (75.5) | 11.24±1.93 |
|
| M1 | 34 (24.5) | 11.94±2.46 |
|
| TNM stage |
|
|
0.009a |
| 0 &
I & II | 91 (65.5) | 11.58±1.74 |
|
| III
& IV | 48 (34.5) | 12.35±1.39 |
|
miR-34a-5p abrogates the facilitation
of malignant behavior mediated by XIST
Furthermore, Pearson's correlation coefficient
indicated that miR-34a-5p was significantly negatively correlated
with XIST (R2=0.2175, P<0.001; Fig. 6A). We hypothesized that miR-34a-5p
may play roles in organisms by targeting XIST. 293T cells were
co-transfected with luciferase construct (pGL3-Control, pGL3-XIST
WT or pGL3-XIST-Mut) and negative control (miR-control) or
miR-34a-5p mimics, respectively. Luciferase reporter assay was
performed to detect the regulatory relationship between miR-34a-5p
and XIST. The results showed that miR-34a-5p markedly decreased the
relative fluorescence value in Luc-wt reporter constructs,
suggesting that miR-34a-5p was a target gene of XIST (P<0.001;
Fig. 6B). In addition, we found
that upregulation of XIST expression significantly decreased
miR-34a-5p expression in BxPC-3 cells, and miR-34a-5p mimics
significantly increased miR-34a-5p expression inhibited by XIST.
Meanwhile, knockdown of XIST significantly increased miR-34a-5p
expression in PANC-1 cells, and miR-34a-5p inhibitors significantly
decreased miR-34a-5p expression induced by XIST (P<0.01,
P<0.001; Fig. 6C). Furthermore,
we used Transwell chamber inserts to explore the effects of
miR-34a-5p and XIST on the migration and invasion of BxPC-3 and
PANC-1 cells. The results indicated that overexpression of XIST
promoted cell proliferation, while miR-34a-5p mimics inhibited cell
proliferation induced by XIST; knockdown of XIST decreased cell
proliferation, and miR-34a-5p inhibitors blocked this decrease
mediated by XIST (Fig. 6D).
Meanwhile, we also found that overexpression of XIST accelerated
cell migration and invasion, while miR-34a-5p mimics inhibited this
acceleration induced by XIST; knockdown of XIST decreased cell
migration and invasion, and miR-34a-5p inhibitors blocked this
decrease mediated by XIST (P<0.001; Fig. 6E).
 | Figure 6.miR-34a-5p abrogates the facilitation
of malignant behavior mediated by XIST. (A) The correlation between
miR-34a-5p and XIST was determined by Pearson's correlation
coefficient in clinical samples (R2=0.2175, P<0.001).
(B) Luciferase reporter assay was performed in 293T cells
co-transfected with luciferase construct (pGL3-Control, pGL3-XIST
WT or pGL3-XIST-Mut) and negative control (miR-control) or
miR-34a-5p mimics, respectively (***P<0.001). (C) BxPC-3 cells
were transfected with control, XIST, XIST plus miR-NC or XIST plus
miR-34a-5p mimics; PANC-1 cells were transfected with control,
shXIST, shXIST plus miR-NC or shXIST plus miR-34a-5p inhibitors,
respectively. qRT-PCR assay was used to detect miR-34a-5p
expression. GAPDH was used as control (**P<0.01, ***P<0.001).
(D) MTT assay was performed to determine the viability of BxPC-3
and PANC-1 cells treated as in C. (E) The migration and invasion
abilities were detected by Transwell assays in BxPC-3 and PANC-1
cells treated as in C (***P<0.001). XIST, X inactive-specific
transcript; |
Discussion
Pancreatic cancer (PC) has been the main cause of
cancer-related death worldwide for several decades (37,38).
Mortality of PC is projected to surpass breast and colorectal
cancer by 2030 in the US (39,40).
The prognosis for patients with PC is poor with a reduced 5-year
overall survival, and the median survival of patients with
untreated PC is only 6 months with an extremely low percentage of
long-term-surviving patients (41–43).
However, at present, effective therapeutic strategies for patients
with PC have been difficult to identify.
Emerging studies have identified numerous genes
which are involved in the pathogenesis of human PC, and lncRNAs are
important (12,14,44–46).
Numerous studies have indicated that lncRNAs participate in the
biological processes of various cancer cells, including cell
proliferation, development, apoptosis and metastases (47–49).
For example, IRAIN was found to promote proliferation and suppress
the apoptosis of PC cells (48);
MALAT-1 accelerates cell growth, migration and invasion in PC
(47). Nevertheless, the mechanisms
of long non-coding RNA XIST in PC are not clear.
MicroRNAs (miRNAs) have been demonstrated to be
involved in the pathogenesis of many diseases, including cancers
(50), infections (51,52)
and diabetes (53). In addition,
the differences in miRNA expression have regulatory functions in
post-transcriptional modification or degradation of their target
genes by binding to complementary regions in the 3′-untranslated
region (UTR) of their target mRNA transcripts (54,55).
Previous studies have demonstrated that miRNAs play a crucial role
in pancreatic development and function. For example, miR-34a-5p
inhibits colorectal cancer metastasis and is related to patient
recurrence (56); miR-34a-5p
enhances the multi-drug resistance of osteosarcoma (57); miR-34a-5p promotes chemoresistance
of osteosarcoma (58).
In the present study, we found that XIST was
significantly upregulated in human PC tissues and PC cell lines,
compared with that noted in the adjacent normal tissues and HBE
normal lung epithelial cell line. Our result also indicated that
XIST expression was markedly higher at later stages of tumor
development and in pre-operation patients with PC. In addition,
XIST significantly increased PC cell viability, G1-G0 phase arrest,
cell proliferation, migration and invasion and inhibited cell
apoptosis in vitro, while XIST knockdown had opposite
effects. The in vivo studies demonstrated that XIST
downregulation suppressed tumor growth. All of these data indicate
that XIST plays an important role in the development of PC.
However, the underlying mechanism by which XIST mediates gene
expression and participates in tumorigenesis remains to be
clarified. Recently, the ceRNA hypothesis proposed that lncRNAs
communicate with other protein-coding RNA transcripts via shared
common miRNA binding sites (32).
According to the ceRNA hypothesis, miRNA complementary base pairing
with XIST was predicted by starBase and TargetScan, and miR-34a-5p
was identified. Quantitative real-time PCR showed that the
expression of miR-34a-5p in BxPC-3 cells was decreased upon
overexpresion of XIST, however increased by transfected with
miR-34a-5p mimic. In PANC-1 cells, the expression of miR-34a-5p was
increased upon knockdown of XIST; however the expression of
miR-34a-5p was suppressed by miR-34a-5p inhibitor. Therefore, we
suggested that XIST is a direct target of miR-34a-5p and there was
interactive suppression between them. Furthermore, we used
Transwell chamber inserts to explore the effects of miR-34a-5p and
XIST on the migration and invasion of BxPC-3 and PANC-1 cells. The
results showed that the transfection of the miR-34a-5p mimic
abrogated the XIST-promoted BxPC-3 cell migration and invasion.
However, transfection of the miR-34a-5p inhibitor hampered
shXIST-decreased PANC-1 cell migration and invasion.
lncRNA XIST functions a tumor-promoting gene in PC.
It promotes cell proliferation and invasion in PC by directly
targeting and suppressing tumor-suppressor miR-34a-5p.
Consequently, XIST could be a potential target for the prevention
of the metastasis of PC.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ryan DP, Hong TS and Bardeesy N:
Pancreatic adenocarcinoma. N Engl J Med. 371:1039–1049. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Egawa S, Toma H, Ohigashi H, Okusaka T,
Nakao A, Hatori T, Maguchi H, Yanagisawa A and Tanaka M: Japan
pancreatic cancer registry; 30th year anniversary: Japan pancreas
society. Pancreas. 41:985–992. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Luo J, Xiao L, Wu C, Zheng Y and Zhao N:
The incidence and survival rate of population-based pancreatic
cancer patients: Shanghai Cancer Registry 2004–2009. PLoS One.
8:e760522013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boon RA, Jae N, Holdt L and Dimmeler S:
Long non-coding RNAs: From clinical genetics to therapeutic
targets? J Am Coll Cardiol. 67:1214–1226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qiu MT, Hu JW, Yin R and Xu L: Long
non-coding RNA: An emerging paradigm of cancer research. Tumour
Biol. 34:613–620. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Spizzo R, Almeida MI, Colombatti A and
Calin GA: Long non-coding RNAs and cancer: A new frontier of
translational research? Oncogene. 31:4577–4587. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang X, Zhi X, Gao Y, Ta N, Jiang H and
Zheng J: LncRNAs in pancreatic cancer. Oncotarget. 7:57379–57390.
2016.PubMed/NCBI
|
|
8
|
Peng W, Gao W and Feng J: Long non-coding
RNA HULC is a novel biomarker of poor prognosis in patients with
pancreatic cancer. Med Oncol. 31:3462014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhan HX, Wang Y, Li C, Xu JW, Zhou B, Zhu
JK, Han HF, Wang L, Wang YS and Hu SY: LincRNA-ROR promotes
invasion, metastasis and tumor growth in pancreatic cancer through
activating ZEB1 pathway. Cancer Lett. 374:261–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng Y, Jutooru I, Chadalapaka G, Corton
JC and Safe S: The long non-coding RNA HOTTIP enhances pancreatic
cancer cell proliferation, survival and migration. Oncotarget.
6:10840–10852. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kim K, Jutooru I, Chadalapaka G, Johnson
G, Frank J, Burghardt R, Kim S and Safe S: HOTAIR is a negative
prognostic factor and exhibits pro-oncogenic activity in pancreatic
cancer. Oncogene. 32:1616–1625. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu JH, Chen G, Dang YW, Li CJ and Luo DZ:
Expression and prognostic significance of lncRNA MALAT1 in
pancreatic cancer tissues. Asian Pac J Cancer Prev. 15:2971–2977.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma C, Nong K, Zhu H, Wang W, Huang X, Yuan
Z and Ai K: H19 promotes pancreatic cancer metastasis by
derepressing let-7's suppression on its target HMGA2-mediated EMT.
Tumour Biol. 35:9163–9169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qu S, Yang X, Song W, Sun W, Li X, Wang J,
Zhong Y, Shang R, Ruan B, Zhang Z, et al: Downregulation of
lncRNA-ATB correlates with clinical progression and unfavorable
prognosis in pancreatic cancer. Tumour Biol. 37:3933–3938. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Smola MJ, Christy TW, Inoue K, Nicholson
CO, Friedersdorf M, Keene JD, Lee DM, Calabrese JM and Weeks KM:
SHAPE reveals transcript-wide interactions, complex structural
domains, and protein interactions across the Xist lncRNA in living
cells. Proc Natl Acad Sci USA. 113:pp. 10322–10327. 2016;
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tantai J, Hu D, Yang Y and Geng J:
Combined identification of long non-coding RNA XIST and HIF1A-AS1
in serum as an effective screening for non-small cell lung cancer.
Int J Clin Exp Pathol. 8:7887–7895. 2015.PubMed/NCBI
|
|
17
|
Yao Y, Ma J, Xue Y, Wang P, Li Z, Liu J,
Chen L, Xi Z, Teng H, Wang Z, et al: Knockdown of long non-coding
RNA XIST exerts tumor-suppressive functions in human glioblastoma
stem cells by up-regulating miR-152. Cancer Lett. 359:75–86. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen DL, Ju HQ, Lu YX, Chen LZ, Zeng ZL,
Zhang DS, Luo HY, Wang F, Qiu MZ, Wang DS, et al: Long non-coding
RNA XIST regulates gastric cancer progression by acting as a
molecular sponge of miR-101 to modulate EZH2 expression. J Exp Clin
Cancer Res. 35:1422016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fang J, Sun CC and Gong C: Long non-coding
RNA XIST acts as an oncogene in non-small cell lung cancer by
epigenetically repressing KLF2 expression. Biochem Biophys Res
Commun. 478:811–817. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhuang LK, Yang YT, Ma X, Han B, Wang ZS,
Zhao QY, Wu LQ and Qu ZQ: MicroRNA-92b promotes hepatocellular
carcinoma progression by targeting Smad7 and is mediated by long
non-coding RNA XIST. Cell Death Dis. 7:e22032016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hayes EL and Lewis-Wambi JS: Mechanisms of
endocrine resistance in breast cancer: An overview of the proposed
roles of non-coding RNA. Breast Cancer Res. 17:402015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Ann Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Muluhngwi P and Klinge CM: Roles for
miRNAs in endocrine resistance in breast cancer. Endocr Relat
Cancer. 22:R279–R300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Filipska M, Skrzypski M, Bigda JJ and
Jassem J: Biological role of prognostic microRNAs (miRNAs) in
squamous lung cancer cell lines. J Thorac Oncol. 10:S391. 2015.
|
|
25
|
Kara M, Yumrutas O, Ozcan O, Celik OI,
Bozgeyik E, Bozgeyik I and Tasdemir S: Differential expressions of
cancer-associated genes and their regulatory miRNAs in colorectal
carcinoma. Gene. 567:81–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang P, Zuo Z, Wu A, Shang W, Bi R, Jin
Q, Wu J and Jiang L: miR-600 inhibits cell proliferation, migration
and invasion by targeting p53 in mutant p53-expressing human
colorectal cancer cell lines. Oncol Lett. 13:1789–1796. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ergun S and Oztuzcu S: Oncocers:
ceRNA-mediated cross-talk by sponging miRNAs in oncogenic pathways.
Tumour Biol. 36:3129–3136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Su X, Xing J, Wang Z, Chen L, Cui M and
Jiang B: microRNAs and ceRNAs: RNA networks in pathogenesis of
cancer. Chin J Cancer Res. 25:235–239. 2013.PubMed/NCBI
|
|
29
|
Guo LL, Song CH, Wang P, Dai LP, Zhang JY
and Wang KJ: Competing endogenous RNA networks and gastric cancer.
World J Gastroenterol. 21:11680–11687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang WC, Fu WM, Wong CW, Wang Y, Wang WM,
Hu GX, Zhang L, Xiao LJ, Wan DC, Zhang JF and Waye MM: The lncRNA
H19 promotes epithelial to mesenchymal transition by functioning as
miRNA sponges in colorectal cancer. Oncotarget. 6:22513–22525.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Qi X, Zhang DH, Wu N, Xiao JH, Wang X and
Ma W: ceRNA in cancer: Possible functions and clinical
implications. J Med Genet. 52:710–718. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long non-coding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiao C, Song Z, Chen J, Zhong J, Cai W,
Tian S, Chen S, Yi Y and Xiao Y: LncRNA-UCA1 enhances cell
proliferation through functioning as a ceRNA of Sox4 in esophageal
cancer. Oncol Rep. 36:2960–2966. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xia T, Chen S, Jiang Z, Shao Y, Jiang X,
Li P, Xiao B and Guo J: Long non-coding RNA FER1L4 suppresses
cancer cell growth by acting as a competing endogenous RNA and
regulating PTEN expression. Sci Rep. 5:134452015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Alzheimer's Association, . 2013
Alzheimer's disease facts and figures. Alzheimers Dement.
9:208–245. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Malvezzi M, Bertuccio P, Levi F, La
Vecchia C and Negri E: European cancer mortality predictions for
the year 2013. Ann Oncol. 24:792–800. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
No authors listed: Cancer statistics JAMA.
310:9822013.
|
|
41
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hidalgo M: Pancreatic cancer. N Engl J
Med. 362:1605–1617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li X, Deng SJ, Zhu S, Jin Y, Cui SP, Chen
JY, Xiang C, Li QY, He C, Zhao SF, et al: Hypoxia-induced
lncRNA-NUTF2P3-001 contributes to tumorigenesis of pancreatic
cancer by derepressing the miR-3923/KRAS pathway. Oncotarget.
7:6000–6014. 2016.PubMed/NCBI
|
|
45
|
Müller S, Raulefs S, Bruns P, Afonso-Grunz
F, Plötner A, Thermann R, Jäger C, Schlitter AM, Kong B, Regel I,
et al: Next-generation sequencing reveals novel differentially
regulated mRNAs, lncRNAs, miRNAs, sdRNAs and a piRNA in pancreatic
cancer. Mol Cancer. 14:942015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ye S, Yang L, Zhao X, Song W, Wang W and
Zheng S: Bioinformatics method to predict two regulation mechanism:
TF-miRNA-mRNA and lncRNA-miRNA-mRNA in pancreatic cancer. Cell
Biochem Biophys. 70:1849–1858. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiao F, Hu H, Yuan C and Wang L, Jiang W,
Jin Z, Guo Z and Wang L: Elevated expression level of long
non-coding RNA MALAT-1 facilitates cell growth, migration and
invasion in pancreatic cancer. Oncol Rep. 32:2485–2492. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lian Y, Wang J, Feng J, Ding J, Ma Z, Li
J, Peng P, De W and Wang K: Long non-coding RNA IRAIN suppresses
apoptosis and promotes proliferation by binding to LSD1 and EZH2 in
pancreatic cancer. Tumour Biol. 37:14929–14937. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zheng S, Chen H, Wang Y, Gao W, Fu Z, Zhou
Q, Jiang Y, Lin Q, Tan L, Ye H, et al: Long non-coding RNA
LOC389641 promotes progression of pancreatic ductal adenocarcinoma
and increases cell invasion by regulating E-cadherin in a
TNFRSF10A-related manner. Cancer Lett. 371:354–365. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sullivan CS and Ganem D: MicroRNAs and
viral infection. Mol Cell. 20:3–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Staedel C and Darfeuille F: MicroRNAs and
bacterial infection. Cell Microbiol. 15:1496–1507. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kato M, Castro NE and Natarajan R:
MicroRNAs: Potential mediators and biomarkers of diabetic
complications. Free Radic Biol Med. 64:85–94. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Filipowicz W: RNAi: The nuts and bolts of
the RISC machine. Cell. 122:17–20. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gao J, Li N, Dong Y, Li S, Xu L, Li X, Li
Y, Li Z, Ng SS, Sung JJ, et al: miR-34a-5p suppresses colorectal
cancer metastasis and predicts recurrence in patients with stage
II/III colorectal cancer. Oncogene. 34:4142–4152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pu Y, Zhao F, Wang H, Cai W, Gao J, Li Y
and Cai S: MiR-34a-5p promotes the multi-drug resistance of
osteosarcoma by targeting the CD117 gene. Oncotarget.
7:28420–28434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Pu Y, Zhao F, Li Y, Cui M, Wang H, Meng X
and Cai S: The miR-34a-5p promotes the multi-chemoresistance of
osteosarcoma via repression of the AGTR1 gene. BMC Cancer.
17:452017. View Article : Google Scholar : PubMed/NCBI
|