Introduction
As one of the seventh most common types of cancer in
women and the most lethal gynecologic cancer, ovarian cancer
affects more than 295,000 females worldwide annually (1). Currently, epithelial ovarian cancer
(EOC) is the major type of ovarian cancer (more than 90% of ovarian
cancer cases are EOCs), and EOC has a higher prevalent rate
compared to non-EOC (2). Based on
statistical reports from 2008 to 2014 in the USA, the overall
5-year survival rate of EOC remains low at 29% (3). This is largely due to the fact that EOC
has few early symptoms and rapidly progress to an advanced stage
(4). More than 70% of patients with
EOC are diagnosed at an advanced stage of the disease (5). Additionally, molecular-targeted
therapies (e.g. anti-PD-1/PD-L1 antibody and CAR-T) have not been
demonstrated to be successful in EOC (6,7).
Innovative treatment with poly(ADP-ribose) polymerase-1 (PARP)
inhibitors (PARPis) has demonstrated outstanding activity in EOC
and can change clinical practice, but at present PARPis are mainly
used in the treatment of BRCA-mutant patients (8). Currently, the mainstay for treatment of
advanced EOC is tumor debulking surgery followed by a combination
of platinum-based combination chemotherapy (usually for six
cycles), with a platinum (carboplatin or cisplatin/CDDP) and a
taxane (paclitaxel/PTX or docetaxel/DTX). Compared to other
chemotherapeutic agents, PTX exhibits high tolerability, decreases
hematological toxicities and neutropenia, thereby leading to an
increase in the duration of the progression-free interval and
overall survival (9). PTX is less
myelosuppressive than DTX (10).
PTX-CDDP chemotherapy has been shown to improve the complete and
partial clinical response by 38.7 and 42.7%, respectively. Thus,
PTX-CDDP chemotherapy is now used as the first-line treatment for
patients with advanced EOC (11). In
spite of initial responsiveness, an estimated 85% patients, showing
increased chemoresistance to the first-line chemotherapy agents,
relapse within two years and ultimately die of recurrent disease
(12). Second-line treatments can
improve survival and quality of life but are not curative (13). Therefore, chemoresistance remains the
main cause of treatment failure associated with advanced EOC
(14), and there is an urgent need to
elucidate the molecular mechanisms underlying chemoresistance to
develop novel therapeutic targets.
MicroRNAs (miRNAs), a cluster of 21–25 nt non-coding
regulatory RNAs, regulate gene expression at the
posttranscriptional level, inducing the degradation of target mRNAs
or translational repression. Over 70% of all human genes are
estimated to be regulated by miRNAs, indicating that gene
regulation by miRNAs is likely involved in nearly every genetic
pathway (15). In this regard,
identification of the function of key miRNAs and a fuller
understanding of their molecular bases should deepen insight into
diverse biological processes and render novel clues to develop
molecular targets for therapeutic intervention (16).
Compelling biochemical data suggest that
dysregulation of miRNA expression is closely associated with
response to chemotherapeutic agents, and alteration in the
expression levels of miRNAs can cause substantial changes during
oncogenesis (17). Hsa-miR-105 is
such a striking example. The complex roles of miR-105 during cancer
initiation and progression are gradually emerging. It can act as a
tumor suppressor by inhibiting tumor growth and metastasis or as an
oncogene by promoting tumor initiation and invasion, depending on
the particular tumor context and base-pairing genes, in a variety
of cancer types including colorectal cancer, hepatocellular
carcinoma, breast cancer, lung cancer and glioma (18).
Interestingly, hsa-miR-105 can be secreted by solid
tumors into circulation in which it further interacts with
Agonature2 (Ago2) to increase stability. This makes hsa-miR-105 a
valuable diagnostic/prognostic biomarker candidate because of its
stability and availability and ease of measurement (19). In the present study, we sought to
determine the expression profile of hsa-miR-105 in PTX-resistant
EOC tissues and cell lines, and to uncover its role and
corresponding molecular basis in the pathogenesis of paclitaxel
resistance in EOC. Overall, our systematic analysis should pave the
way for a better understanding of this unique miRNA in EOC.
Materials and methods
Data mining
A microRNA profiling microarray expression dataset
consisting of three pairs of parental EOC cells and their
PTX-resistant sublines (GSE148251) (20) was downloaded from Gene Expression
Omnibus (GEO platform). The miRNA expression levels were analyzed
using GEO2R bioconductor project and provided as log2 ratios
(21). We explored three mRNA
target-predicting algorithms including mirSVR, Target scan and
miRDB to screen the potential downstream targets of hsa-miR-105
(22).
Cell treatment
The human EOC cell lines (SKOV3, TOV21G and OVCAR8)
and 293T cells, obtained from the American Type Culture Collection
(ATCC), were maintained in RPMI-1640 medium supplemented with 10%
(v/v) fetal bovine serum (FBS), penicillin (100 U/ml), streptomycin
(100 U/ml) and amphotericin B (25 µg/ml) at 37°C in a 5%
CO2 atmosphere. The PTX-resistant cell lines (SKOV3/PTX
and TOV21G/PTX) were generated by exposing parental cells to
gradually increased doses of PTX (Sigma-Aldrich; Merck KGaA), and
were finally maintained in complete medium containing 50 nM PTX
(23). All cell lines used were
recently authenticated using short tandem repeat analysis performed
by Suzhou Genetic Testing Biotechnology, and were tested for
mycoplasma contamination using Universal Mycoplasma Detection Kit
(ATCC) prior to the experiments.
Oligonucleotide transfection and
lentiviral transduction
Hsa-miR-105 mimics and inhibitors for in
vitro transfection and hsa-miR-105 micrON™ agomir for in
vivo transfection, along with their negative controls, were
purchased from Ribobio. Zinc And Ring Finger 2 (ZNRF2) lentiviral
shRNA particle was obtained from GeneCopoeia. To manipulate the
hsa-miR-105 expression, cells were transfected with 100 nM mimics
or inhibitors with the aid of Lipofectamine™ 3000 (Thermo Fisher
Scientific, Inc.) for 48 h. For lentiviral transduction, the viral
particles were prepared by co-transfection of expression plasmid
and packaging plasmids into 293T cells, followed by harvest of
virus-containing supernatants 48 h after transfection. SKOV3 and
TOV21G cells were then transduced with the lentivirus, followed by
selection of positive clones using 1 µg/ml of puromycin
(Sigma-Aldrich; Merck KGaA) for ~14 days.
Measurement of in vitro
cytotoxicity
Forty-eight hours after transfection with the
indicated oligonucleotides, EOC cells were seeded in 96-well plates
at the density of 5×103 cells/well. Cells were then
exposed to different doses of PTX as indicated or DMSO for 24 h
after adhering to the plate. Upon completion of PTX treatment,
fresh medium was added and cell viability was determined using Cell
Counting Kit-8 (MedChemExpress), as per the manufacturers'
instructions. Each concentration was conducted in triplicate and
repeated at least twice. The half maximal inhibitory concentration
(IC50) of cell viability was calculated as the level
that caused 50% reduction in cell viability vs. the parental cells
(24).
Clonogenic assay
About 500 cells were seeded into the 6-well plates.
After a 48-h culture, PTX (20 nM) was added into the cultures.
After a 3-day treatment with PTX, cells were transferred to fresh
medium and cultured for another 12 days until colonies were large
enough to be clearly discerned. Cells were then fixed using 4%
paraformaldehyde (Sigma-Aldrich; Merck KGaA) for 20 min, followed
by staining with crystal violet stain solution (0.5%) (Sangon
Biotech). Counting colonies was finally achieved by using Oracle
Java SE software (25), with the
average number of colonies being plotted (mean ± SD, n=3).
Cell invasion assay
The invasiveness of EOC cells in PTX-containing
medium was determined by Transwell chamber (26). Briefly, 1×105 cells were
seeded in the upper chamber of a Transwell with Matrigel-coated
membrane (Corning, Inc.). After a 24-h culture, cells were then
treated with 20 nM of PTX for another 24 h, followed by removal of
cells from the upper chamber. The cells that had migrated to the
lower surface were fixed and stained with
4′,6-diamidino-2-phenylindole (DAPI; Thermo Fisher Scientific,
Inc.) for 5 min and then counted under a Leica M205 FCA
fluorescence microscope (Leica Microsystems, Inc.). Ten randomly
selected fields were chosen for cell number quantification.
In vivo inhibition of hsa-miR-105 and
tumor xenograft assay
All animal research was conducted in accordance with
Guide for the Care and Use of Laboratory Animals from NIH, and was
approved by the Animal Research Ethics Board of First Affiliated
Hospital of Xi'an Jiaotong University (approval no. XMU-2012-1682).
Animals were maintained under a constant 12-h light:12-h darkness
cycle (lights on at 08:00 h) and controlled conditions of humidity
(between 70 and 80%) and temperature (22±1°C). They were allowed
free access to experimental diets and water, and were allowed to
acclimatize for at least 1 week before the experiment. For
establishment of the tumor xenograft model, 2×106
SKOV3/PTX or TOV21G/PTX cells were subcutaneously injected into
flanks of female BALB/cA-nu mice (n=125, 6-weeks-old, weighing
19–23 g, obtained from the Animal Facility of The First Affiliated
Hospital of Xi'an Medical University). On day 13 post cell
inoculation, mice received intraperitoneal injections of 5 mg/kg of
PTX (dissolved in PBS)/kg body weight every three days. On the
following day of the first PTX injection, 10 nmol of hsa-miR-105
micrONTM agomir or negative control (dissolved in 0.1 ml
PBS) was locally injected into the tumor mass once every 3 days for
two weeks. Mice receiving PBS injection served as the negative
controls. The animals were checked daily, and tumor volume (V) was
recorded and calculated using the formula: V=(length ×
width2)/2. Humane endpoints used in this study included
tumor ulceration, infection, or necrosis, loss of ability to
ambulate (inability to access food or water) and labored
respiration. Four weeks after cell inoculation, mice were
sacrificed by cervical dislocation following sodium pentobarbital
anesthesia (200 mg/kg, intraperitoneally). Some xenograft tissues
were snap frozen in liquid nitrogen, followed by being stored at
−80°C. The other xenografts were immediately fixed in 10% formalin
and processed for histological examination.
Clinical specimens
Upon receipt of the written informed consent, a
total of 115 female patients with EOC and known clinical follow-up
were recruited from the Department of Obstetrics and Gynecology,
First Affiliated Hospital of Xi'an Medical University during August
2012 and June 2019. All patients with EOC underwent initial
cytoreductive surgery followed by 6–8 cycles of standard
paclitaxel-based chemotherapy (27).
Patients with progressive disease during primary chemotherapy
according to medical image analysis or with recurrent disease
within 6 months of completing primary chemotherapy were termed
PTX-resistant (28). Some tumor
samples were snap frozen in liquid nitrogen, followed by being
stored at −80°C until use, whereas other tumor samples were fixed
in 10% formalin and processed for histological examination. The
protocols involved in the human study strictly conformed to the
2008 Revised Declaration of Helsinki, and were approved by the
Ethics Committee of the First Affiliated Hospital of Xi'an Medical
University (approval no. XMU-2012-1683).
Real-time RT-qPCR
Real-time RT-qPCR was carried out as described
elsewhere (29,30). Total RNA was isolated and purified
using PureLink™ miRNA Isolation Kit (Thermo Fisher Scientific,
Inc.). RT-qPCR analysis of miRNA expression was performed using
Applied Biosystems TaqMan microRNA assay system (Applied
Biosystems), with the aid of microRNA-specific primers
(5′-AAAAGCUGGGUUGAGAGGGCGA-3′) and TaqMan™ MicroRNA Reverse
Transcription Kit (Thermo Fisher Scientific, Inc.). To quantify the
expression levels of other target genes, 500 ng of total RNA was
used to synthesize the first strand cDNA using iScript™ cDNA
Synthesis Kit (Bio-Rad Laboratories, Inc.). Subsequent qPCR was
carried out on the Applied Biosystems® 7500 Fast
Real-Time PCR System. Relative expression levels of target miRNA
and mRNAs were determined using the standard ΔΔCq method
(31), with human U6 snRNA or
18S RNA serving as the internal controls (32). Primers used in the current study are
listed in Table I.
 | Table I.Primer sequences used for real-time
PCR analysis. |
Table I.
Primer sequences used for real-time
PCR analysis.
| Gene | Primer
sequence |
|---|
| 18S | F:
CTCGCCGCGCTCTACCTACCTA |
|
| R:
ATGAGCCATTCGCAGTTTCACTGTA |
| TJP1 | F:
AGCCATTCCCGAAGGAGTTG |
|
| R:
ATCACAGTGTGGTAAGCGCA |
| SEPT14 | F:
CTGTGCCTGACCCACAATGA |
|
| R:
TGCTGCTATGCGTCAGACAT |
| TM9SF2 | F:
GCCTGAAGAGGCCAGTCAAT |
|
| R:
AATTCATTCTCGGGGAGGGC |
| ANKRD44 | F:
CGGTGTTGTGACTGAAACCC |
|
| R:
AGGTTCTCGGGTCAGATCCA |
| SNX4 | F:
AGTTTTCAGTGAATGGAGTGCC |
|
| R:
ATGTTTCCTGCACACAGCCC |
| ZNRF2 | F:
AGCACGACTGCCTTGTCTAT |
|
| R:
TTCTGGCTGAGTATGCCTGC |
|
C1orf168 | F:
GGTTCGAGCACCTCAACTTC |
|
| R:
CCCCTGATAGTGTGACCAGC |
| EIF4A2 | F:
TCCAATGAGACAGGGCACAC |
|
| R:
ATGGCTGCCTCAGTCCTTTC |
| LMO7 | F:
AGAGAATCCACAAGGAATTGA |
|
| R:
TAGGGACTGTCATTTGGCCC |
| MLF1 | F:
GGCCCACTTAGAGCGAAACT |
|
| R:
ATGAGAACCCCAGCCTTTGG |
| ZFPM2 | F:
ACCCGGAAGTGAGATGCAAG |
|
| R:
TCTCTTGGTCTGGTATTCCATCA |
| YTHDF3 | F:
CACCATTTCTTGGAACCCCAC |
|
| R:
TGCCCTTAAAGCTACACATACA |
| RAPGEF2 | F:
GATGCGAAGGGATTGCCCC |
|
| R:
CGTACAGGAGGTCAGTGGTG |
| F13A1 | F:
CAACAGCTTGGAGGAGGGAG |
|
| R:
CCCTTTTTGCAGCAACCCAG |
| SLC5A1 | F:
CCAGCTTTTGTCACAGGTGC |
|
| R:
AAACCCTGCAGGACAGATGG |
| HNRNPK | F:
CAAAGGCCCACGGAGTTCTA |
|
| R:
TATTCGCACTCCCCCTACCT |
| LDLRAD3 | F:
GGCGTCCCCCTACCCTGTCAT |
|
| R:
TGCTGCACACGGCTCCTCAC |
Western blot analysis
Western blot analysis was conducted according to
published protocols (33). Total
protein was prepared using ReadyPrep™ Protein Extraction Kit
(Bio-Rad Laboratories, Inc.). Protein samples (~30 µg) were
separated on 10% SDS-PAGE and transferred to PDVF membranes
(Millipore Corp.). After being blocked by 5% nonfat dry milk in
TBST, the membranes were incubated with primary antibodies
(Table II) at 4°C overnight.
Positive signals were developed using an EZ-ECL Chemiluminescence
Detection kit (Biological Industries) on a GOBlot™ Western Blot
Processor (Cytoskeleton) after incubation with proper secondary
antibodies.
| Table II.Sources of antibodies and the working
dilutions that were used for western blotting or immunostaining in
the present study. |
Table II.
Sources of antibodies and the working
dilutions that were used for western blotting or immunostaining in
the present study.
| Antibody | Vendor | Catalog no. | Dilutions |
|---|
| Rabbit
anti-Ki67 | Abcam | ab15580 | 1:500 (IHC) |
| Rabbit
anti-ZNRF2 | Thermo Fisher
Scientific, Inc. | PA5-84890 | 1:1,000 (IB) 1:150
(IHC) |
| Rabbit
anti-Flag | Sigma-Aldrich/Merck
KGaA | F7425 | 1:1,000 (IB) |
| Rabbit
anti-ACTIN | Abcam | ab8227 | 1:2,500 (IB) |
|
VECTASTAIN® Elite®
ABC HRP Kit | Vector
Laboratories, Inc. | PK-7200 | 1:500 (IHC) |
| Goat anti-rabbit
IgG-HRP secondary antibody | Abcam | ab97200 | 1:10,000 (IB) |
Morphological examination
Localization of hsa-miR-105 in EOC cells was
revealed by RNA fluorescence in situ hybridization (FISH)
using a locked nucleic acid (LNA) probe for human mature
hsa-miR-105 (5′-UCAAAUGCUCAGACUCCUGUGGU-3′) and scramble miRNA
control probe (5′-GTGTAACACGTCTATACGCCCA-3′; Exiqon), both of which
were labeled with digoxigenin at the 5′end with the aid of the DIG
RNA Labeling Kit (Roche). EOC cells were fixed in 4%
paraformaldehyde, followed by hybridization at 37°C overnight in a
dark moist chamber. After thorough rinse, cells were incubated with
TSA fluorescent signal reaction solution (PerkinElmer) at room
temperature for 30 min and were then sealed using Fluoroshield™
with DAPI histology mounting medium (Sigma-Aldrich; Merck KGaA).
Positive staining was observed under a Leica M205 FCA fluorescence
microscope.
The distribution of hsa-miR-105 in EOC tissues was
revealed using chromogenic in situ hybridization (CISH)
(29). Briefly, after a routine
deparaffinization and rehydration, 4% paraformaldehyde-fixed
paraffin-embedded EOC tissue sections were treated with 10 µg/ml of
Proteinase K (Sigma-Aldrich; Merck KGaA) to expose RNA. Sections
were then incubated sequentially with hybridization buffer without
probes at 21°C for 2 h and with hybridization buffer containing 40
nM of the above-mentioned DIG-labeled LNA-modified probes
overnight. After incubation with the blocking solution (PBST
containing 2% goat serum and 2 mg/ml bovine albumin) for 1 h,
sections were treated with anti-digoxigenin-AP antibody
(Sigma-Aldrich; Merck KGaA) at 4°C overnight, followed by signal
development using the BCIP/NBT Substrate Kit (Vector Laboratories,
Inc.) according to the manufacturer's instruction.
Localization of the zinc and ring finger (ZNRF2)
protein in EOC tissues was detected using immunohistochemistry as
described elsewhere (34). In brief,
4% paraformaldehyde-fixed paraffin-embedded EOC tissue sections
were routinely deparaffinized and rehydrated, followed by antigen
retrieval by incubating slides in 0.01% citrate acid in a heated
water bath (95°C) for 30 min. After elimination of endogenous
peroxides activity by treatment for 10 min with 0.3% hydrogen
peroxide in methanol, the sections were incubated with the primary
antibody (Table II) at 4°C
overnight. Positive signals were finally developed with the aid of
the VECTASTAIN® Elite® ABC system (Vector
Laboratories, Inc.) according to the protocol recommended by the
manufacturer.
Luciferase reporter assay
The 3′UTR segment of human ZNRF2 was amplified from
ovarian cell genomic DNA and cloned into psiCHECK-2 luciferase
reporter vector using NotI/XhoI sites. Site-directed
mutagenesis of the hsa-miR-105 binding site on the ZNRF2 3′UTR was
achieved using QuikChange Site-Directed Mutagenesis Kit (Agilent
Technologies, Inc.). Co-transfection of reporter plasmids and
hsa-miR-105 mimics into 293T cells was carried out using
Lipofectamine™ 3000. Forty-eight hours after transfection, relative
luciferase activity was measured by the Dual-Luciferase Reporter
Assay System (Promega Corp.).
Ago2-mediated
RNA-immunoprecipitation
FLAG-Ago2 was a gift from Edward Chan (Addgene
plasmid no. 21538; Addgene) (35).
The FLAG-Ago2 construct and hsa-miR-105 mimics were con-transfected
into 293T cells for 48 h using Lipofectamine™ 3000, followed by
cell harvest using RIPA buffer supplemented with RNase A, DNase and
protease-free from Thermo Fisher Scientific, Inc. The
FLAG® Tag antibody (Thermo Fisher Scientific, Inc.) was
preincubated with Protein G-agarose beads (Sigma-Aldrich; Merck
KGaA) at 4°C overnight. Subsequently, the resultant cell lysates
were incubated with preconjugated antibody at 4°C overnight.
Pellets from immunoprecipitation were then dissolved in TRIsure™
(BioLine) and subjected to PCR analysis.
Statistical analysis
All experiments were conducted in triplicate and
repeated at least twice. Quantitative values are expressed as the
mean ± SD. Differences among/between experimental groups were
analyzed by one-way analysis of variance (ANOVA) followed by
Tukey's test or Student's t-test, wherever appropriate. The
association between hsa-miR-105 expression and overall survival was
assessed using the log-rank Kaplan-Meier analysis. The receiver
operating characteristic (ROC) curve was used to examine the
predictive power. P<0.05 was considered to be statistically
significant.
Results
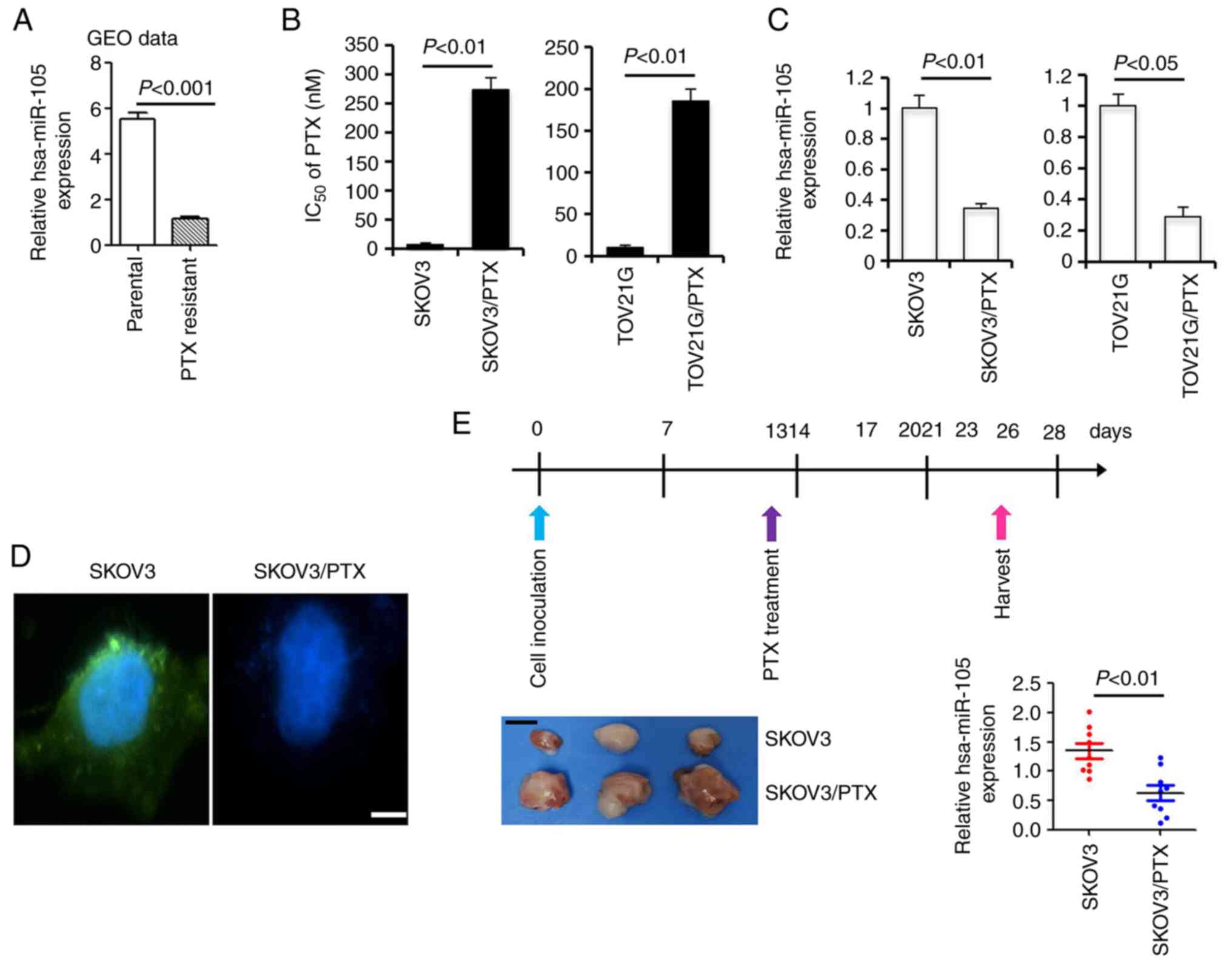
Downregulation of hsa-miR-105
expression in paclitaxel-resistant EOC cells
As an initial effort to understand the potential
involvement of hsa-miR-105 in paclitaxel resistance, we examined
the expression of hsa-miR-105 in a microRNA profiling microarray
expression dataset consisting of three pairs of parental EOC cells
and their PTX-resistant sublines (GSE148251) (20). Student's t-test demonstrated that the
expression of hsa-miR-105 was significantly downregulated in
PTX-resistant sublines compared to their parental cell lines
(percentage decrease >78.8%, P<0.0001, Fig. 1A). To validate these findings, we
generated the PTX-resistant SKOV3/PTX and TOV21G/PTX cell lines
using a previously validated protocol. SKOV3/PTX and TOV21G/PTX
cells were −31.4 and −18.1 times more resistant to PTX than their
parental SKOV3 and TOV21G cells, respectively (Fig. 1B). Compared to the expression level of
hsa-miR-105 in the parental cells which was set at 100%, the
relative percentage of hsa-miR-105 expression in SKOV3/PTX and
TOV21G/PTX cells was 34.2 and 28.6%, respectively (Fig. 1C). Moreover, FISH analysis revealed a
highly enriched expression of hsa-miR-105 in the parental SKOV3
cells compared to the PTX-resistant SKOV3/PTX subline (Fig. 1D). To further provide the in
vivo evidence for the association between deregulated
expression of hsa-miR-105 and paclitaxel resistance, we established
a xenograft model to mimic the clinical treatment with PTX. At two
weeks after PTX treatment, xenografts derived from SKOV3/PTX cells
were clearly larger than those derived from parental SKOV3 cells,
indicative of a successful resistance of SKOV3/PTX cells to PTX.
Subsequent qPCR analysis showed that the expression of hsa-miR-105
in the SKOV3/PTX xenografts was significantly reduced by ~53.6%
compared to the SKOV3 ×enografts (P<0.01, Fig. 1E). These data together suggest that
hsa-miR-105 deficiency correlates with paclitaxel resistance in EOC
cells.
Inhibition of hsa-miR-105 confers
paclitaxel resistance in EOC cells
To explore the oncogenic roles of the hsa-miR-105
deficiency, we silenced the expression of hsa-miR-105 in SKOV3 and
TOV21G cells using specific inhibitors (anti-miR-105) (Fig. 2A). Following PTX treatment, knockdown
of hsa-miR-105 expression conferred a stronger pro-survival effect
in a dose-dependent manner, as revealed by viability assays
(Fig. 2B). Consistently, higher
efficiency of colony formation upon PTX co-incubation was observed
in the hsa-miR-105 inhibitor (anti-miR-105)-transfected SKOV3 and
TOV21G cells (Fig. 2C). Additionally,
ablation of endogenous expression of hsa-miR-105 significantly
increased cell invasiveness by ~3.6- and ~3.2-fold in
PTX-challenged SKOV3 and TOV21G cells, respectively (Fig. 2D). We also used another serous EOC
cell line (OVCAR8) to elucidate the potential involvement of
hsa-miR-105 in the development of resistance to PTX (Fig. S1). These findings suggest that
hsa-miR-105 deficiency may confer resistance to paclitaxel in EOC
cells.
Restoration of hsa-miR-105 expression
in situ leads to dramatic inhibition of EOC growth
In order to investigate the antitumor effects of
hsa-miR-105 at the in vivo level, we employed the
micrONTM agomir system which has been shown to be
promising for therapeutic gene delivery in vivo (36). We evaluated the therapeutic effects of
micrONTM agomir-mediated hsa-miR-105 restoration on
tumor growth in a xenograft model derived from either SKOV3/PTX or
TOV21G/PTX cells (Fig. 3A).
Intratumoral injection of hsa-miR-105 micrONTM agomir
resulted in a significant increase in hsa-miR-105 expression in
xenografts at 28 days after cell inoculation (Fig. 3B), as well as an inhibition of tumor
growth, especially during the late phase of PTX treatment (Fig. 3C). In accordance, the
hsa-miR-105-overexpressing and PTX-treated tumors were smaller at
28 days after cell inoculation (Fig.
3D), and expressed lower levels of Ki67 (a well-known
proliferation marker for human tumor cells, Fig. 3E), compared to the Ctrl group. These
results indicate that in vivo inhibition of hsa-miR-105
expression by using the micrONTM agomir may have the
potential to be used as an alternative therapeutic option for
EOC.
Identification of ZNRF2 as a direct
target of hsa-miR-105 in EOC
To further dissect the molecular basis underlying
the effects of hsa-miR-105 on PTX responsiveness, we explored three
mRNAtarget-predicting algorithms including mirSVR, Target scan and
miRDB (22). A total of 17 genes were
identified to be potential downstream targets of hsa-miR-105, as
these genes were observed to be overlapped in all three databases
(Fig. 4A). We then evaluated the
transcriptional expression levels of these 17 genes in four pair of
PTX-responsive and PTX-resistant EOC tissues using RT-qPCR.
ZNRF2 was the only candidate gene whose expression was found
to be increased by more than 3-fold in the PTX-resistant tissues
compared with the PTX-responsive EOC tissues (Fig. 4B). Transfection with hsa-miR-105
mimics caused a significant decrease in the expression of ZNRF2 in
SKOV3/PTX and TOV21G/PTX cells, at both mRNA (Fig. 4C) and protein levels (Fig. 4D). Consistently, immunostaining and
ISH analyses revealed that the expression of hsa-miR-105 was
negatively correlated to the expression of ZNRF2 protein in EOC
specimens from our clinical cohort (Fig.
4E). To provide the direct evidence for the direct regulation
of ZNRF2 expression by hsa-miR-105, we performed luciferase
reporter assay. Co-transfection of reporter plasmids and
hsa-miR-105 mimics into 293T cells resulted in a ~66.4% reduction
in the transcriptional activity of ZNRF2 3′UTR, while this
inhibitory effect was totally abolished when the binding site of
hsa-miR-105 on the 3′UTR of ZNRF2 was mutated (Fig. 4F). To better confirm the interaction
between hsa-miR-105 and ZNRF2, we conducted Ago2-mediated RNA-IP
assay. In the cells that were co-overexpressed with hsa-miR-105 and
FLAG-Ago2, ZNRF2 mRNA was found to bind hsa-miR-105
(Fig. 4G). These results collectively
suggest that hsa-miR-105 may regulate cellular responsiveness to
PTX by directly targeting ZNRF2 in EOC cells.
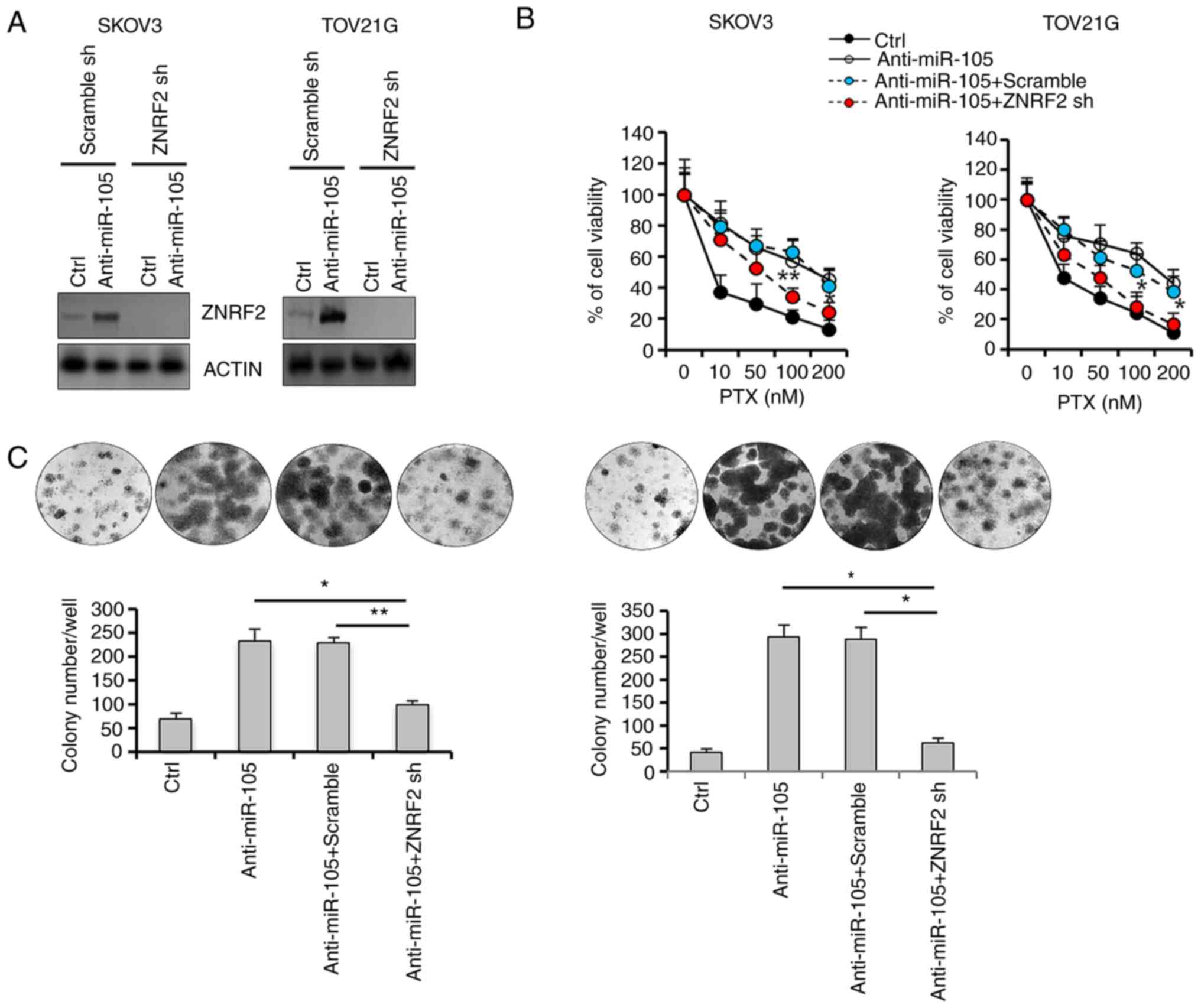
Validation of the functional role of
ZNRF2 in hsa-miR-105-mediated PTX resistance
Since ZNRF2 has been shown to be involved in
chemoresistance (37), we next
investigated the biological effects of the manipulation of ZNRF2
expression in hsa-miR-105-mediated chemoresistance by generating
SKOV3 and TOV21G cells that were stably transfected with ZNRF2
shRNA. Transfection with hsa-miR-105 inhibitors (anti-miR-105) in
control cells resulted in a significant increase in the expression
level of ZNRF2, whereas this induction was totally abolished in the
SKOV3 and TOV21G cells that were stably deprived of endogenous
ZNRF2 (Fig. 5A). Hsa-miR-105
inhibition-induced cell survival upon PTX challenge was attenuated
in the SKOV3 and TOV21G cells that were stably transfected with
ZNRF2 shRNA (Fig. 5B). Furthermore,
ZNRF2 depletion enhanced the ability of PTX to decrease colony
formation efficiency (Fig. 5C), in
the presence of hsa-miR-105 inhibition in both SKOV3 and TOV21G
cells. Thus, we identified ZNRF2 as a main downstream effector
mediating the action of hsa-miR-105 in the promotion of resistance
to PTX.
Tumor and circulating hsa-miR-105
serves as a biomarker to predict PTX survival in EOC patients
In order to broaden the translational research
enterprise of the current study, we measured the expression levels
of hsa-miR-105 in tumor samples from a total of 115 EOC patients
that had received standard paclitaxel-based chemotherapy. The
association between hsa-miR-105 expression and clinicopathological
parameters of the patients with EOC is summarized in Table SI. Patients with high levels of
hsa-miR-105 were more likely to have clinical response to PTX
chemotherapy (P<0.0042). Tumor stage and grade did not
significantly differ between patients with high levels of
hsa-miR-105 and patients with low levels of hsa-miR-105 (Table SI). RT-qPCR revealed that hsa-miR-105
was significantly downregulated in the PTX-resistant EOC specimens
compared with that in the PTX-responsive EOC specimens
(P<0.0001, Fig. 6A). A ROC curve
was then generated to assess the predictive power of hsa-miR-105.
The corresponding AUC was 0.8649 (95% CI, 0.8024–0.9275) (Fig. 6B), suggesting that hsa-miR-105 may
serve as a valuable diagnostic biomarker for EOC chemotherapy.
Importantly, the patients with lower levels of hsa-miR-105
exhibited poorer survival following PTX-based chemotherapy
(P=0.0014; Fig. 6C). Previous studies
have identified hsa-miR-105 as a circulating miRNA (19). Given that circulating miRNAs are
potent and convenient biomarkers for the evaluation of tumor
development and progression (38), we
sought to determine the clinical implication of hsa-miR-105
expression in the peripheral circulation using our 115-patient
cohort. Expression levels of hsa-miR-105 were significantly
decreased in the plasma of patients with PTX-resistant EOC,
compared with those from patients with PTX-responsive EOC
(P<0.0001; Fig. 6D). The
contingency plot indicated that high levels of plasma hsa-miR-105
were associated with improved responsiveness to PTX (Fig. 6E). Subsequent ROC curve analysis
revealed that the corresponding AUC was 0.8797 (95% CI,
0.8169–0.9424) (Fig. 6F), verifying
the predictive power of the circulating hsa-miR-105. These clinical
data support our pre-clinical findings and show that downregulation
of hsa-miR-105 occurs in both tumor and peripheral circulation in
the EOC patients with acquired PTX resistance.
Discussion
Hsa-miR-105 is located on human chromosome Xq28
(miRBase). Accumulated data suggest that hsa-miR-105 can play
differential roles in various cancer types. For instance,
hsa-miR-105 promotes the invasion and metastasis of colorectal
cancer cells by directly targeting the RAP2C signaling (39). Likewise, hsa-miR-105 functions as an
essential oncomiR in both non-small lung cancer (40) and breast cancer (41). By contrast, we demonstrated here that
the expression of hsa-miR-105 was significantly downregulated in
paclitaxel-resistant epithelial ovarian cancer (EOC) tissues and
cell lines, and inhibition of the expression of hsa-miR-105
conferred paclitaxel resistance in EOC cells. Our findings are in
accordance with previous reports that hsa-miR-105 exerts its
anti-proliferative effects on gastric cancer (42) and inhibits cell aggressiveness in
prostate cancer cells (43), thus
indicating a tumor-suppressor role of this unique miRNA. Based on
the available data, we reason that hsa-miR-105 may function as a
double-faced gene expression regulator in cancer contexts, and
whether it acts as an oncomiR or a tumor-suppressor miRNA is
largely dependent on cancer type and cell context.
It is significant to know what causes the
downregulation of hsa-miR-105 particularly in PTX-resistant EOC.
Two possibilities are herein proposed. First, compelling data have
suggested that epigenetic regulation functions as an essential
mechanism modulating the expression of miRNAs (44,45). In
this regard, Zhou et al reported that hsa-miR-105 expression
is decreased in gastric cancer cells, concomitantly with the DNA
hypermethylation in the upstream region of the hsa-miR-105 promoter
(42). Meanwhile, hypomethylation of
CpGs in the promoter of hsa-miR-105 was found to activate the
expression of the downstream CT-GABRA3 signaling pathway in
melanoma cells (46). In the present
study, treatment with a DNA methylation inhibitor RG108 partially
restored the expression levels of hsa-miR-105 in PTX-resistant
SKOV3/PTX or TOV21G/PTX cells (data not shown). Thus, deregulation
of DNA hypermethylation in the hsa-miR-105 promoter may at least in
part explain the reduced expression of hsa-miR-105 in PTX-resistant
EOC cells. On the other hand, we cannot exclude the possibility
that hsa-miR-105 expression is also subjected to a delicate control
at the genetic level. In favor of this hypothesis, MYC and the core
component of the NF-κB transcription complex (namely p65) are both
proved to be crucial upstream regulators of hsa-miR-105 by directly
targeting its chromatin (47,48). Interestingly, by using bioinformatics,
we observed that the hsa-miR-105 promoter contains a potential
binding site of the MTA1 transcription factor. Given that there is
an interactive feedback loop between MTA1 and the net transcription
of the components of NF-κB pathway (49), it will be of future interest to
investigate if a MTA1/hsa-miR-105 axis is also at play in drug
resistance in EOC.
By using luciferase reporter assay, site-directed
mutagenesis and Ago2-mediated RNA-immunoprecipitation, we further
identify zinc and ring finger 2 (ZNRF2) as a direct target of
hsa-miR-105 in EOC cells. ZNRF2 functions as a component of the
amino acid sensing machinery that calibrates the cell's amino acid
uptake, regulates proteasomal ubiquitin-dependent protein catabolic
process, and governs cell fate (50).
Recent advances in this field point out two characteristics
regarding the roles of ZNRF2 in tumor biology. First, ZNRF2 is
tightly associated with mammalian target of rapamycin (mTOR)
signaling. ZNRF2 promotes amino acid-stimulated mTORC1
translocation to lysosomes to activate mTOR signaling (51). Given that mTORC1 and its downstream
mRNA translation effectors protect cells against genotoxic DNA
damage (52), it is therefore a
logical observation that ZNRF2 is involved in the pathogenesis of
paclitaxel resistance in EOC. Moreover, ZNRF2 expression is tightly
regulated at the posttranscriptional level by miRNAs. For example,
miR-100 suppresses human osteosarcoma cell proliferation and
chemoresistance by directly targeting ZNRF2 (37). Similarly, miR-153-3p targets ZNRF2 to
modulate tumorigenesis of papillary thyroid cancer (53). In our study, stable depletion of ZNRF2
expression enhanced the ability of PTX to decrease colony formation
efficiency and to increase apoptosis in the presence of hsa-miR-105
inhibition in both SKOV3 and TOV21G cells. These findings lend
functional support to the notion that ZNRF2 may serve as a main
downstream target of hsa-miR-105 during the pathogenesis of
paclitaxel resistance. Additionally, our in vivo assays have
important therapeutic implications for EOC treatment, since
restoration of the hsa-miR-105 expression in situ by
intratumoral injection of micrONTM agomir significantly
potentiated sensitivity to PTX and inhibited tumor growth. Thus,
replenishment of hsa-miR-105 expression may contribute to
chemotherapeutic sensitization as an alternative approach for EOC
treatment.
The importance of the hsa-miR-105 pathway in the
pathogenesis of chemoresistance in EOC was further attested by our
in vitro studies, using combined paclitaxel and cisplatin
treatment in SKOV3 and OVCAR8 cells that were deprived of
hsa-miR-105 expression. Apparently, ablation of the endogenous
hsa-miR-105 potentiated collateral resistance to cisplatin and
paclitaxel in ovarian cancer cells (Fig.
S2). In patients with cisplatin-resistant EOC, single
chemotherapeutic agent (such as paclitaxel) has been shown to
produce an objective response rate of less than 30%. It is
currently unclear how this collateral resistance occurs (54). To this end, our results suggest that
acquired cisplatin resistance and acquired paclitaxel resistance
may share a common pathogenic mechanism (e.g. regulation by
miRNAs). Thus, further elucidating the molecular basis of
hsa-miR-105 should provide valuable insights into therapeutic
strategies for EOC patients.
Admittedly, a limitation of our study is that, due
to the complex nature of this area, the exact mechanism underlying
paclitaxel resistance cannot be elucidated by one single miRNA.
Moreover, while our expression analyses focused on hsa-miR-105, we
cannot rule out the potential involvement of additional miRNAs in
the cancerous control of ZNRF2 expression, as our in silico
analyses also highlighted other miRNAs with the ability to bind
directly to the ZNRF2 gene. In this context, the future
lines of research should be focused on investigating how
differential miRNAs in EOC concordantly build regulatory networks
that control different cellular responses to paclitaxel.
To summarize, in this study, we report that
hsa-miR-105 expression levels were significantly decreased in
PTX-resistant EOC tissues and cells. hsa-miR-105 regulates the
chemosensitivity to PTX by directly targeting the ZNRF2 signaling
pathway. Considering that hsa-miR-105 is a validated circulating
miRNA (19), and that patients with
lower levels of serum hsa-miR-105 tend to exhibit a poorer survival
following PTX-based chemotherapy (present study), testing patients
for hsa-miR-105 expression may provide more accurate prognostic
information and could influence the recommended course of PTX
treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The author(s) received no financial support for the
research, authorship, and/or publication of this article.
Availability of data and materials
Data and materials are available upon reasonable
request.
Authors' contributions
RA designed and supervised the experiments, and
wrote the main manuscript text. ML performed the experiments and
data analysis, and prepared the figures. SZ, YM, and YY provided
administrative, technical, and material support. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The protocols involved in the human study strictly
conformed to the 2008 Revised Declaration of Helsinki, and were
approved by the Ethics Committee of The First Affiliated Hospital
of Xi'an Jiaotong University (approval no. XMU-2012-1683). All
animal research was conducted in accordance with the Guide for the
Care and Use of Laboratory Animals from the NIH, and was approved
by the Animal Research Ethics Board of The First Affiliated
Hospital of Xi'an Jiaotong University (approval no.
XMU-2012-1682).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Pokhriyal R, Hariprasad R, Kumar L and
Hariprasad G: Chemotherapy resistance in advanced ovarian cancer
patients. Biomark Cancer. 11:1179299X198608152019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sawai T, Miyahara M, Nakajima H, Nakamori
S, Sakai M, Tamada H, Tanigawa T, Yamada N, Nakamura M and Ito M:
The impact of preprocedural hemoglobin A1c on the efficacy of
sirolimus-eluting and paclitaxel-eluting stents. Cardiovasc Interv
Ther. 27:72–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Suh DH, Kim M, Lee KH, Eom KY, Kjeldsen
MK, Mirza MR and Kim JW: Major clinical research advances in
gynecologic cancer in 2017. J Gynecol Oncol. 29:e312018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Winter WE III, Maxwell GL, Tian C, Carlson
JW, Ozols RF, Rose PG, Markman M, Armstrong DK, Muggia F and
McGuire WP; Gynecologic Oncology Group Study, : Prognostic factors
for stage III epithelial ovarian cancer: A gynecologic oncology
group study. J Clin Oncol. 25:3621–3627. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu G, Zhang Q, Li D, Zhang L, Gu Z, Liu
J, Liu G, Yang M, Gu J, Cui X, et al: PD-1 silencing improves
anti-tumor activities of human mesothelin-targeted CAR T cells. Hum
Immunol. 82:130–138. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Le Saux O, Dubois B, Stern MH, Terme M,
Tartour E, Classe JM, Chopin N, Trédan O, Caux C and Ray-Coquard I:
Current advances in immunotherapy in ovarian cancer. Bull Cancer.
107:465–473. 2020.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cook SA and Tinker AV: PARP inhibitors and
the evolving landscape of ovarian cancer management: A review.
BioDrugs. 33:255–273. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Naskou J, Beiter Y, van Rensburg R,
Honisch E, Rudelius M, Schlensog M, Gottstein J, Walter L, Braicu
EI, Sehouli J, et al: EZH2 loss drives resistance to carboplatin
and paclitaxel in serous ovarian cancers expressing ATM. Mol Cancer
Res. 18:278–286. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vasey PA, Paul J, Birt A, Junor EJ, Reed
NS, Symonds RP, Atkinson R, Graham J, Crawford SM, Coleman R, et
al: Docetaxel and cisplatin in combination as first-line
chemotherapy for advanced epithelial ovarian cancer. Scottish
gynaecological cancer trials group. J Clin Oncol. 17:2069–2080.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
du Bois A, Luck HJ, Meier W, Adams HP,
Möbus V, Costa S, Bauknecht T, Richter B, Warm M, Schröder W, et
al: A randomized clinical trial of cisplatin/paclitaxel versus
carboplatin/paclitaxel as first-line treatment of ovarian cancer. J
Natl Cancer Inst. 95:1320–1329. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Foley OW, Rauh-Hain JA and del Carmen MG:
Recurrent epithelial ovarian cancer: An update on treatment.
Oncology (Williston Park). 27:288–294, 298. 2013.PubMed/NCBI
|
|
13
|
Hennessy BT, Coleman RL and Markman M:
Ovarian cancer. Lancet. 374:1371–1382. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang SF, Wang XY, Fu ZQ, Peng QH, Zhang
JY, Ye F, Fu YF, Zhou CY, Lu WG, Cheng XD and Xie X: TXNDC17
promotes paclitaxel resistance via inducing autophagy in ovarian
cancer. Autophagy. 11:225–238. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huh JH, Kim TH, Kim K, Song JA, Jung YJ,
Jeong JY, Lee MJ, Kim YK, Lee DH and An HJ: Dysregulation of
miR-106a and miR-591 confers paclitaxel resistance to ovarian
cancer. Br J Cancer. 109:452–461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei L, Sun J, Zhang N, Zheng Y, Wang X, Lv
L, Liu J, Xu Y, Shen Y and Yang M: Noncoding RNAs in gastric
cancer: Implications for drug resistance. Mol Cancer. 19:622020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marengo B, Pulliero A, Izzotti A and
Domenicotti C: MiRNA regulation of glutathione homeostasis in
cancer initiation, progression and therapy resistance. Microrna.
9:187–197. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Zhang Z, Chen F, Hu T, Peng W, Gu Q
and Sun Y: The diverse oncogenic and tumor suppressor roles of
microRNA-105 in cancer. Front Oncol. 9:5182019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li HY, Liang JL, Kuo YL, Lee HH, Calkins
MJ, Chang HT, Lin FC, Chen YC, Hsu TI, Hsiao M, et al:
MiR-105/93-3p promotes chemoresistance and circulating
miR-105/93-3p acts as a diagnostic biomarker for triple negative
breast cancer. Breast Cancer Res. 19:1332017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kazmierczak D, Jopek K, Sterzynska K,
Ginter-Matuszewska B, Nowicki M, Rucinski M and Januchowski R: The
significance of MicroRNAs expression in regulation of extracellular
matrix and other drug resistant genes in drug resistant ovarian
cancer cell lines. Int J Mol Sci. 21:26192020. View Article : Google Scholar
|
|
21
|
Chai Y, Tan F, Ye S, Liu F and Fan Q:
Identification of core genes and prediction of miRNAs associated
with osteoporosis using a bioinformatics approach. Oncol Lett.
17:468–481. 2019.PubMed/NCBI
|
|
22
|
Pla A, Zhong X and Rayner S: MiRAW: A deep
learning-based approach to predict microRNA targets by analyzing
whole microRNA transcripts. PLoS Comput Biol. 14:e10061852018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Y, Hu D, Qiu J, Xie X, Ye F and Lu WG:
Overexpression of glycogen synthase kinase-3 in ovarian carcinoma
cells with acquired paclitaxel resistance. Int J Gynecol Cancer.
21:439–444. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun NK, Huang SL, Chang TC and Chao CC:
TLR4 and NFKB signaling is critical for taxol resistance in ovarian
carcinoma cells. J Cell Physiol. 233:2489–2501. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Niyazi M, Niyazi I and Belka C: Counting
colonies of clonogenic assays by using densitometric software.
Radiat Oncol. 2:42007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao G, Li Y and Wang T: Potentiation of
docetaxel sensitivity by miR-638 via regulation of STARD10 pathway
in human breast cancer cells. Biochem Biophys Res Commun.
487:255–261. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fu Z, Wang C, Chen Y, Zhang X, Wang X and
Xie X: Down-regulation of UTP23 promotes paclitaxel resistance and
predicts poorer prognosis in ovarian cancer. Pathol Res Pract.
215:1526252019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li X, Lu Y, Chen Y, Lu W and Xie X:
MicroRNA profile of paclitaxel-resistant serous ovarian carcinoma
based on formalin-fixed paraffin-embedded samples. BMC Cancer.
13:2162013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang LL, Ma J, Yang B, Zhao J, Yan BY,
Zhang YQ and Li W: Interference with lactate metabolism by
mmu-miR-320-3p via negatively regulating GLUT3 signaling in mouse
Sertoli cells. Cell Death Dis. 9:9642018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dong YS, Hou WG, Li Y, Liu DB, Hao GZ,
Zhang HF, Li JC, Zhao J, Zhang S, Liang GB and Li W: Unexpected
requirement for a binding partner of the syntaxin family in
phagocytosis by murine testicular Sertoli cells. Cell Death Differ.
23:787–800. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He K, Qu H, Wang H, Zhang S, Qian XH and
Li W: Regulated and functional expression of the corepressor MTA3
in rodent testis. Endocrinology. 157:4400–4410. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang C, Lai JH, Hu B, Zhang S, Zhao J and
Li W: A chromatin modifier regulates Sertoli cell response to
mono-(2-ethylhexyl) phthalate (MEHP) via tissue inhibitor of
metalloproteinase 2 (TIMP2) signaling. Biochim Biophys Acta.
1839:1170–1182. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang S, Li W, Zhu C, Wang X, Li Z, Zhang
J, Zhao J, Hu J, Li T and Zhang Y: Sertoli cell-specific expression
of metastasis-associated protein 2 (MTA2) is required for
transcriptional regulation of the follicle-stimulating hormone
receptor (FSHR) gene during spermatogenesis. J Biol Chem.
287:40471–40483. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lian SL, Li S, Abadal GX, Pauley BA,
Fritzler MJ and Chan EK: The C-terminal half of human Ago2 binds to
multiple GW-rich regions of GW182 and requires GW182 to mediate
silencing. RNA. 15:804–813. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hou J, Lin L, Zhou W, Wang Z, Ding G, Dong
Q, Qin L, Wu X, Zheng Y, Yang Y, et al: Identification of miRNomes
in human liver and hepatocellular carcinoma reveals miR-199a/b-3p
as therapeutic target for hepatocellular carcinoma. Cancer Cell.
19:232–243. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xiao Q, Yang Y, An Q and Qi Y:
MicroRNA-100 suppresses human osteosarcoma cell proliferation and
chemo-resistance via ZNRF2. Oncotarget. 8:34678–34686. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang L and Zhang L: Circulating exosomal
miRNA as diagnostic biomarkers of neurodegenerative diseases. Front
Mol Neurosci. 13:532020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shen Z, Zhou R, Liu C, Wang Y, Zhan W,
Shao Z, Liu J, Zhang F, Xu L, Zhou X, et al: MicroRNA-105 is
involved in TNF-α-related tumor microenvironment enhanced
colorectal cancer progression. Cell Death Dis. 8:32132017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jin X, Yu Y, Zou Q, Wang M, Cui Y, Xie J
and Wang Z: MicroRNA-105 promotes epithelial-mesenchymal transition
of nonsmall lung cancer cells through upregulating Mcl-1. J Cell
Biochem. 120:5880–5888. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhou W, Fong MY, Min Y, Somlo G, Liu L,
Palomares MR, Yu Y, Chow A, O'Connor ST, Chin AR, et al:
Cancer-secreted miR-105 destroys vascular endothelial barriers to
promote metastasis. Cancer Cell. 25:501–515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou GQ, Han F, Shi ZL, Yu L, Li XF, Yu C,
Shen CL, Wan DW, Zhu XG, Li R and He SB: DNMT3A-mediated
down-regulation of microRNA-105 promotes gastric cancer cell
proliferation. Eur Rev Med Pharmacol Sci. 21:3377–3383.
2017.PubMed/NCBI
|
|
43
|
Honeywell DR, Cabrita MA, Zhao H,
Dimitroulakos J and Addison CL: MiR-105 inhibits prostate tumour
growth by suppressing CDK6 levels. PLoS One. 8:e705152013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cheng FHC, Lin HY, Hwang TW, Chen YC,
Huang RL, Chang CB, Yang W, Lin RI, Lin CW, Chen GCW, et al: E2F6
functions as a competing endogenous RNA, and transcriptional
repressor, to promote ovarian cancer stemness. Cancer Sci.
110:1085–1095. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Braga EA, Loginov VI, Filippova EA,
Burdennyi AM, Pronina IV, Kazubskaya TP, Khodyrev DS, Utkin DO,
Kushlinskii DN, Adamyan LV and Kuslinskii NE: Diagnostic value of a
group of MicroRNA genes hypermethylated in ovarian carcinoma. Bull
Exp Biol Med. 166:253–256. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Loriot A, Van Tongelen A, Blanco J,
Klaessens S, Cannuyer J, van Baren N, Decottignies A and De Smet C:
A novel cancer-germline transcript carrying pro-metastatic miR-105
and TET-targeting miR-767 induced by DNA hypomethylation in tumors.
Epigenetics. 9:1163–1171. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yan W, Wu X, Zhou W, Fong MY, Cao M, Liu
J, Liu X, Chen CH, Fadare O, Pizzo DP, et al: Cancer-cell-secreted
exosomal miR-105 promotes tumour growth through the MYC-dependent
metabolic reprogramming of stromal cells. Nat Cell Biol.
20:597–609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ji Q, Xu X, Xu Y, Fan Z, Kang L, Li L,
Liang Y, Guo J, Hong T, Li Z, et al: MiR-105/Runx2 axis mediates
FGF2-induced ADAMTS expression in osteoarthritis cartilage. J Mol
Med (Berl). 94:681–694. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bui-Nguyen TM, Pakala SB, Sirigiri RD, Xia
W, Hung MC, Sarin SK, Kumar V, Slagle BL and Kumar R: NF-kappaB
signaling mediates the induction of MTA1 by hepatitis B virus
transactivator protein HBx. Oncogene. 29:1179–1189. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hoxhaj G, Caddye E, Najafov A, Houde VP,
Johnson C, Dissanayake K, Toth R, Campbell DG, Prescott AR and
MacKintosh C: The E3 ubiquitin ligase ZNRF2 is a substrate of
mTORC1 and regulates its activation by amino acids. Elife.
5:e122782016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yang M, Zhai Z, Zhang Y and Wang Y:
Clinical significance and oncogene function of long noncoding RNA
HAGLROS overexpression in ovarian cancer. Arch Gynecol Obstet.
300:703–710. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Musa F, Alard A, David-West G, Curtin JP,
Blank SV and Schneider RJ: Dual mTORC1/2 inhibition as a novel
strategy for the resensitization and treatment of
platinum-resistant ovarian cancer. Mol Cancer Ther. 15:1557–1567.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cui Z, Luo Z, Lin Z, Shi L, Hong Y and Yan
C: Long non-coding RNA TTN-AS1 facilitates tumorigenesis of
papillary thyroid cancer through modulating the miR-153-3p/ZNRF2
axis. J Gene Med. 21:e30832019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ribeiro JR, Schorl C, Yano N, Romano N,
Kim KK, Singh RK and Moore RG: HE4 promotes collateral resistance
to cisplatin and paclitaxel in ovarian cancer cells. J Ovarian Res.
9:282016. View Article : Google Scholar : PubMed/NCBI
|