Introduction
Global warming is a challenge to human development,
which causes irreversible changes to the ecosystem on the earth.
Climate warming not only causes a negative effect to the human
survival environment, but also brings serious damage to the health
of human beings. In recent years, the effect of a continuous high
temperature environment on health and well-being is receiving
increasing attention (1). A high
temperature environment as a source of stress triggers a series of
stress responses of the body. It causes the imbalance of internal
metabolism, damage to the tissue and organ, and fatigue (2). The imbalance of the body's heat may cause
the heat-stress reaction (3), and the
body's core temperature continues to rise due to the disorder of
the heat balance. Body temperature is vital for the body to
maintain the physiological function, as cell biological and
enzymatic reactions are affected by the temperature. Cell damage
will occur due to the denatured intraocular protein and enzyme
induced by the raised core temperature. By measuring the body's
core temperature of rats, the influence of heat exposure on the
body's core temperature can be explored.
The hypothalamus-pituitary-adrenal (HPA) axis is the
primary neuroendocrine system involved in the stress response
(4). The HPA axis secretes
adrenocorticotropic hormone (ACTH) and cortisol (Cor). The change
of ACTH and Cor adapt to the adverse environment, which is the
evaluated index for the degree of stress and plays an extremely
important role in the body (5). The
weight of the organs change under the state of stress (6), including the pituitary, adrenal and
hypothalamus. In the present study, by testing the organ
coefficient of the pituitary and adrenal glands, and by testing the
levels of ACTH and Cor in serum, whether the high temperature
environment (32°C) caused the heat-stress response in rats was
explored. Different types of stress can affect the immune function,
such as psychological, cold, thermal and fear stress. Interleukin-2
(IL-2) is mainly produced by the active T cells and has a variety
of functions in the immune system. IL-2 also affects the HPA axis,
and its level can be used as an important index of cellular immune
function (7). IL-12 enhances the
activity of T cells and natural killer (NK) cells. These indicators
were measured to explore the association between heat stress and
immunological function.
In general, under the condition of heat stress, the
damage to animal tissues and organs occurs; the damage to the
immune organs is more serious. The apoptotic cells activate the
immune response (8) and they also
start an immune tolerance (9). Of
note, numerous studies have shown that strong stress is harmful to
the organs, tissues, cells and immune system, but occasionally mild
stress is beneficial for the body in the heat acclimatization
(10–12). Therefore, the present study aimed to
explore whether the heat exposure is harmful to the body by
detecting the expression of apoptosis genes in order to observe
whether the cells are damaged under the moderate thermal
environment and to discuss the influence of heat stress on the body
by evaluating the changes of neuroendocrine and immune
function.
Materials and methods
Animals and heat exposure
protocol
Male Sprague-Dawley rats, weighing 180–200 g, were
purchased from the Laboratory Animal Center of Ningxia Medical
University (Ningxia, China). Rats were permitted to eat food ad
libitum. The cycle of light and dark was 12:12 h (6:00
p.m.-6:00 a.m. as the light cycle and 6:00 a.m.-6:00 p.m. as the
dark cycle every day). The rats were maintained in separate cages,
with only 1/cage. The cage size was 44×27×19 cm. Room temperature
was 24.0±0.1°C and relative humidity was 54±5%. In the course of
the experiment, cages were cleaned, and food and water were
replaced at a random time every 2 or 3 days. Rats were anesthetized
with intraperitoneal injection of phenobarbital sodium (50 mg/kg),
and subsequently temperature sensors were placed and kept for 2
weeks for recovery. Sixteen rats were randomly divided into the
control (CN) and heat exposure (HE) groups (n=8/group). Rats in the
CN group were fed at room temperature throughout the study. Rats in
the HE group received a fixed 8 h (9:00 a.m.-5:00 p.m.) heat
exposure process a day, and the exposure was finished inside the
artificial climate chamber with a temperature of 32°C (relative
humidity of 60±5%). Following each exposure the rats were kept at
room temperature. The heat exposure lasted for 7 days. The
experimental procedures of the present study were approved by the
Animal Ethics Committee of Ningxia Medical University and Use
Committee, in accordance with the guidelines of the Council of the
Physiological Society of China.
Body weight and viscera
coefficient
The body temperature of the rats was collected by a
wireless temperature sensor (TA10TA-F40; Data Sciences
International, St. Paul, MN, USA) and the processing was
respectively analyzed in accordance with the day and night.
Following the heat exposure, the body weight was analyzed. Each rat
was anesthetized by intraperitoneal injection of phenobarbital
sodium (50 mg/kg). The pituitary and adrenal gland were removed and
weighed (XS105 Dual Range; Mettler-Toledo Inc., Columbus, OH, USA)
quickly, and subsequently the organ weight was calculated by the
following formula: Organ coefficient = organ weight/animal body
weight.
Detection of the serum ACTH, Cor, IL-2
and IL-12
Blood was collected from the inferior vena cava,
centrifuged at 3,500 × g for 15 min and the serum was segregated
for detection. Cor and ACTH levels were detected by
radioimmunoassay. The serum IL-2 and IL-12 were detected using
commercially available sandwich ELISA kits, which were the Rat IL-2
ELISA kit and Rat IL-12 ELISA kit (Chenglin Biotechnology, Beijing,
China). All the detections were tested in accordance with the
manufacturer's instructions. Absorbance was read at 450 nm (Bio-Rad
680; Bio-Rad Laboratories Co., Ltd., Hercules, CA, USA). The
quantity of IL-2 in the serum was estimated from a calibration
curve, which ranged between 80 and 1,500 ng/l. The quantity of
IL-12 in the serum was estimated from a calibration curve, which
ranged between 2 and 40 pg/ml.
Detection of caspase-3, Bcl2 and Bax
in spleen
The spleens were quickly removed and placed into
liquid nitrogen. Frozen samples were reserved at −80°C until
further analysis. The spleen organs were homogenized with
glass-Teflon®. Total RNA was prepared from the splenic organ (100
mg) with TRIzol® reagent (Invitrogen Life Technologies, Carlsbad,
CA, USA) according to the manufacturer's instructions.
Complementary DNA (cDNA) was synthesized with a First Strand cDNA
Synthesis kit (Thermo Fisher Scientific Inc., Waltham, MA, USA).
Reverse transcription-polymerase chain reaction (PCR) was carried
out using a Maxima SYBR-Green PCR kit (Thermo Fisher Scientific
Inc.) with indicated primers. After an initial 10 min at 95°C, the
PCR program was finished as follows: 95°C for 15 sec, 60 or 57°C
for 30 sec and extension at 72°C for 30 sec, for 40 cycles. At the
end of the reaction, melting curve analysis was performed to ensure
the specificity of the reaction. β-actin was used as an internal
control and primers used for the PCR are shown in Table I.
 | Table I.Sequence of oligonucleotide
primers. |
Table I.
Sequence of oligonucleotide
primers.
| Gene | Sequence (5′→3′) | Length, bp | Temperature,°C | GenBank |
|---|
| Caspase-3 | F:
AGCTGGACTGCGGTATTGAG | 104 | 60 | NM_012922 |
|
| R:
GGGTGCGGTAGAGTAAGCAT |
|
|
|
| Bcl2 | F:
AGCCTGAGAGCAACCGAAC | 159 | 60 | NM_016993 |
|
| R:
AGCGACGAGAGAAGTCATCC |
|
|
|
| Bax | F:
TTGCTACAGGGTTTCATCCAG | 145 | 57 | NM_017059 |
|
| R:
TGTTGTTGTCCAGTTCATCG |
|
|
|
| β-actin | F:
CACCCGCGAGTACAACCTTC | 207 | 60 | NM_031144 |
|
| R:
CCCATACCCACCATCACACC |
|
|
|
Statistical analysis
All the data were analyzed by SPSS, Inc., (version
21.0; IBM Corp., Armonk, NY, USA), and the results are represented
as mean ± standard deviation. The statistical difference was
evaluated using the t-test. P<0.05 and P<0.01 were considered
to indicate a statistically significant difference.
Result
Body core temperature of rats
The dynamic change of the body core temperature of
Sprague-Dawley rats showed that the body's core temperature in the
two groups was lower in the daylight and higher at night, but there
was no statistical significance between the two groups (Fig. 1).
Body weight and organ coefficient
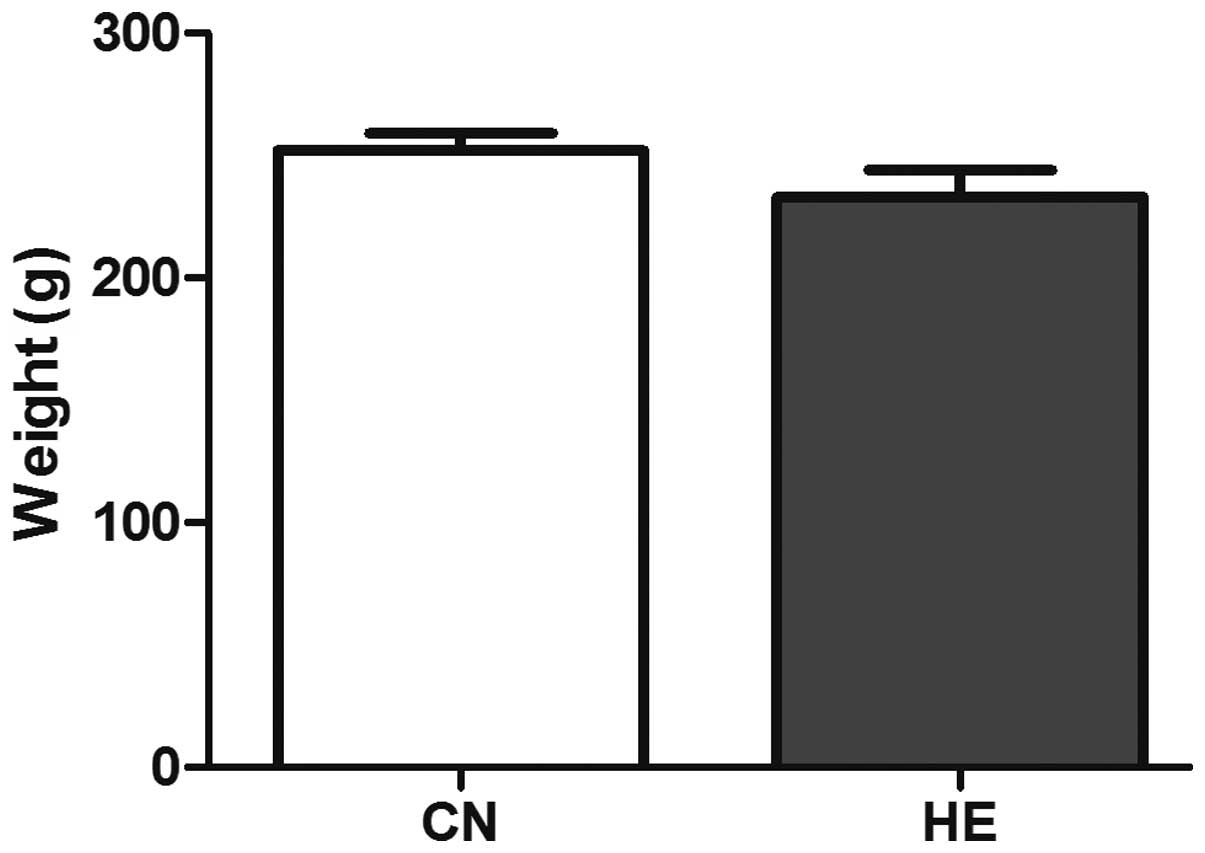
The body weight was reduced in the HE group, but no
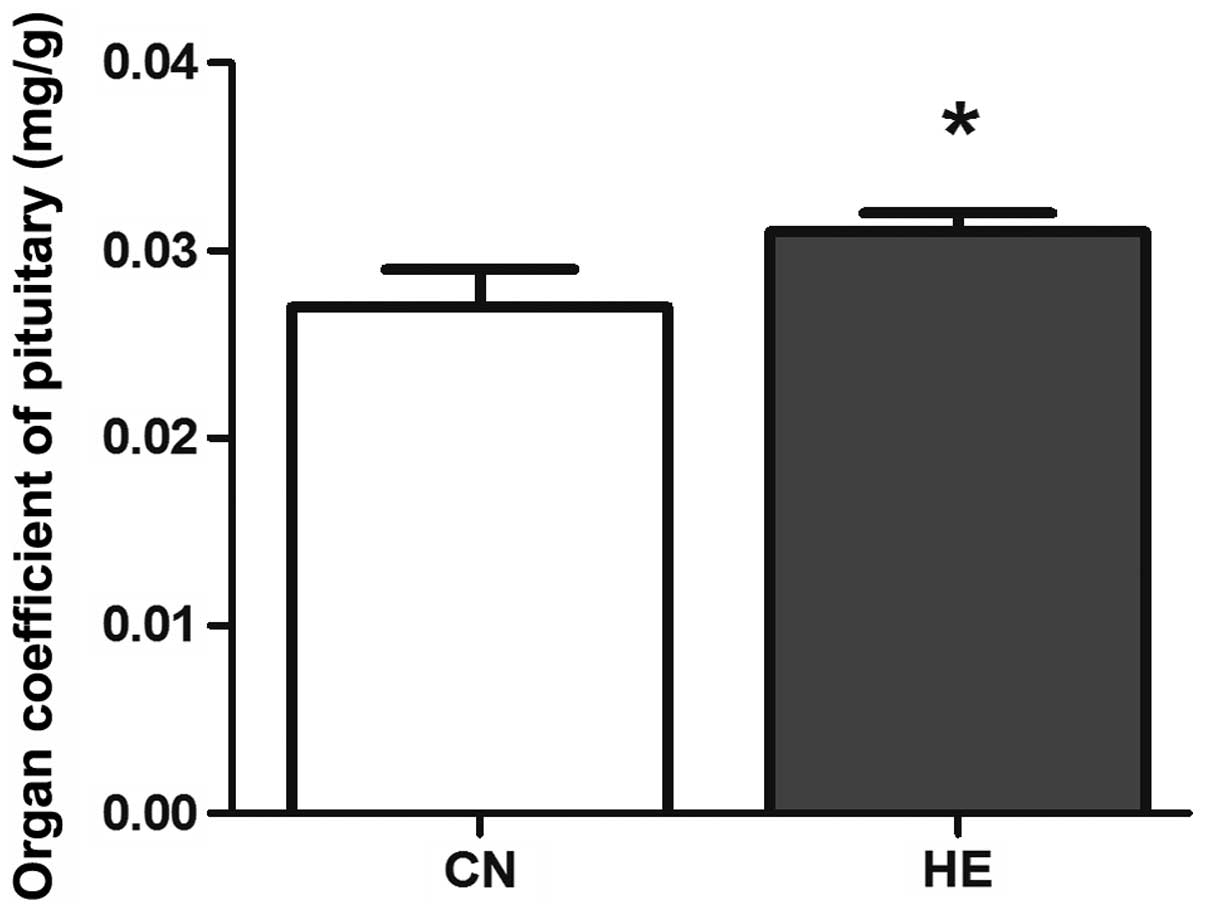
statistical significance was observed (P>0.05) (Fig. 2). The organ coefficient of the
pituitary in the HE group was increased significantly compared to
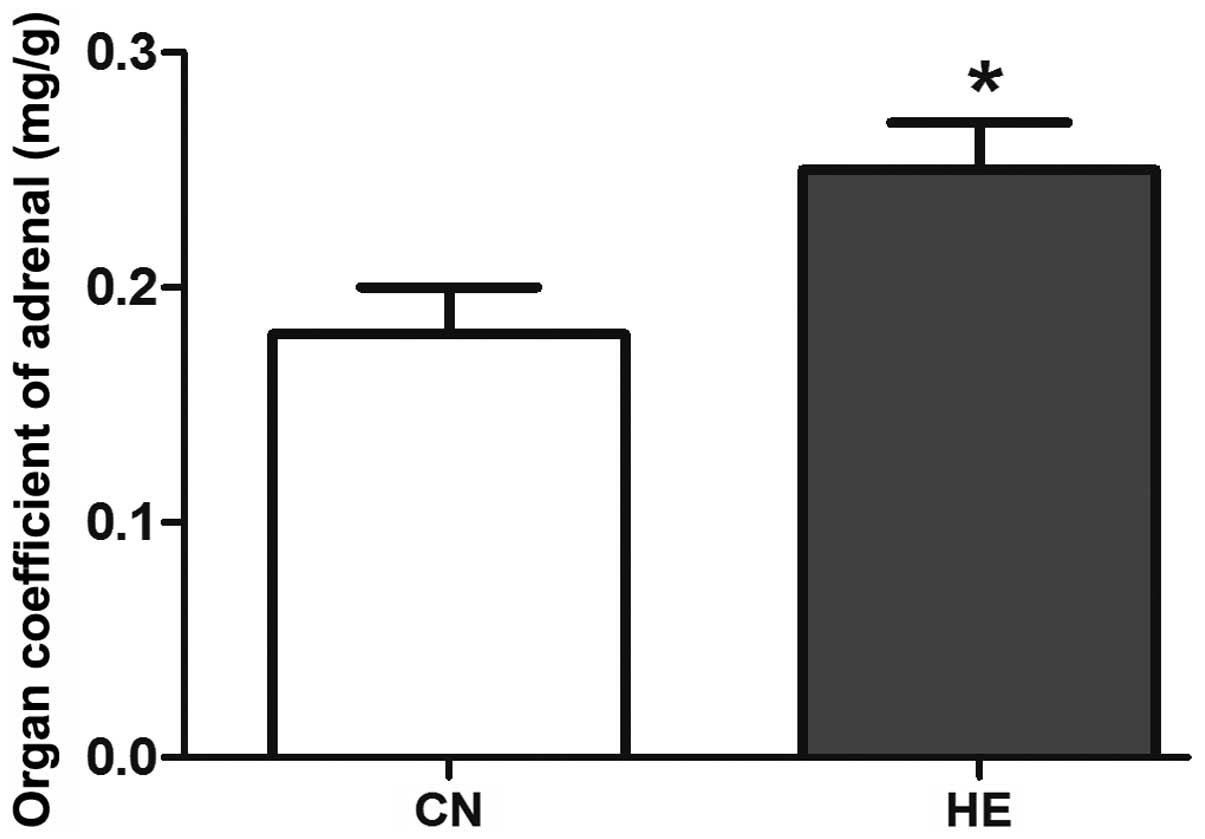
the CN group (P<0.05) (Fig. 3), and
the organ coefficient of the adrenal glands was evidently higher
than that of the CN group (P<0.05) (Fig. 4).
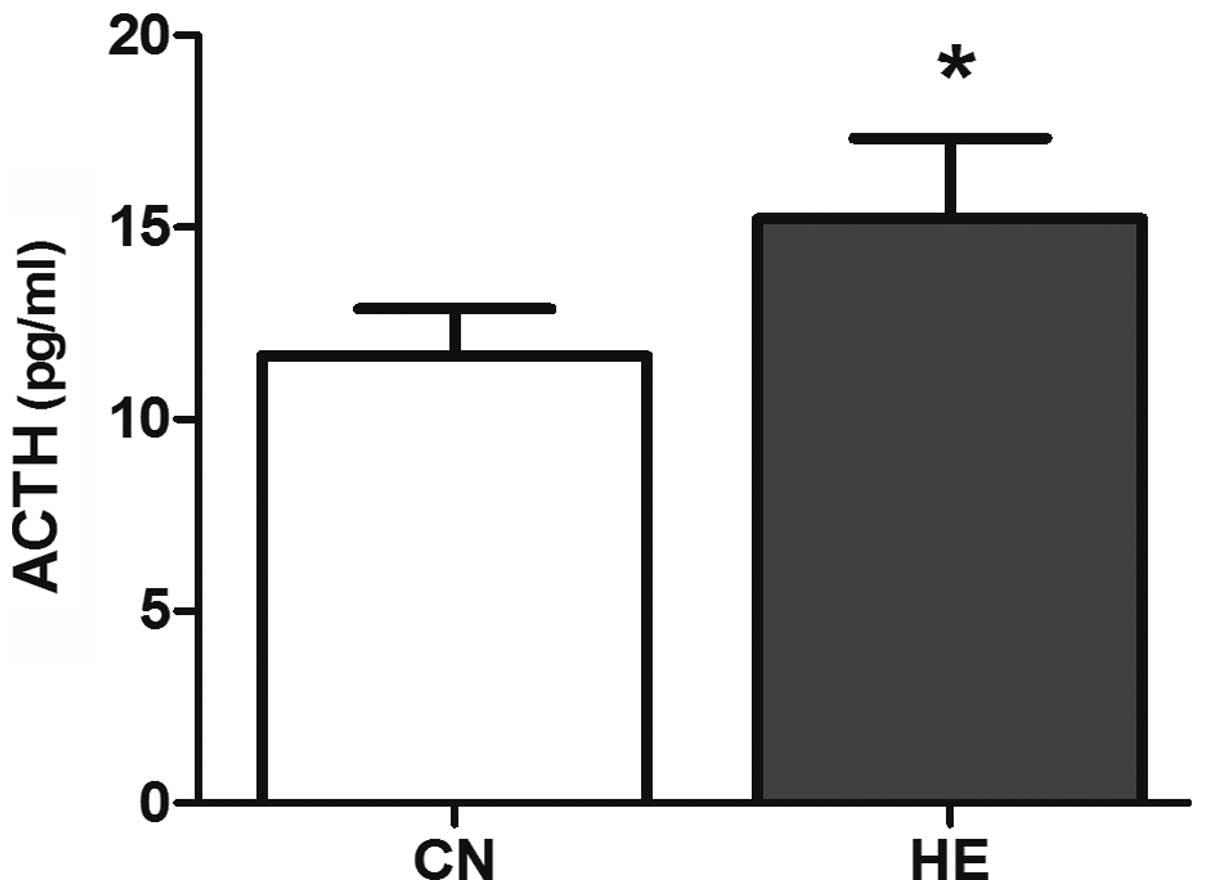
Serum ACTH and Cor concentrations
Compared with the CN group, the serum ACTH level was
higher (P<0.05) (Fig. 5) and the
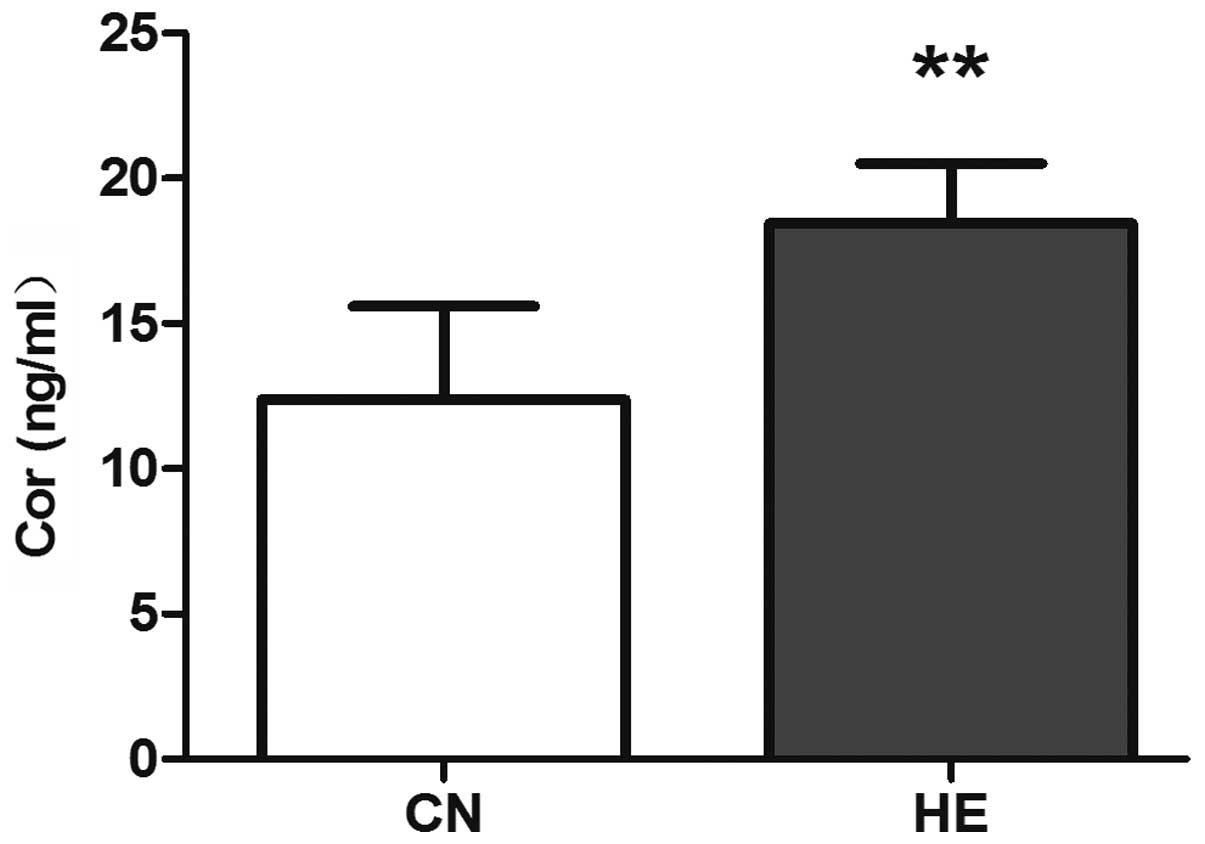
Cor level increased significantly (P<0.01) (Fig. 6) in the HE group.
Serum IL-2 and IL-12
concentrations
The serum IL-2 in the HE group is higher than that
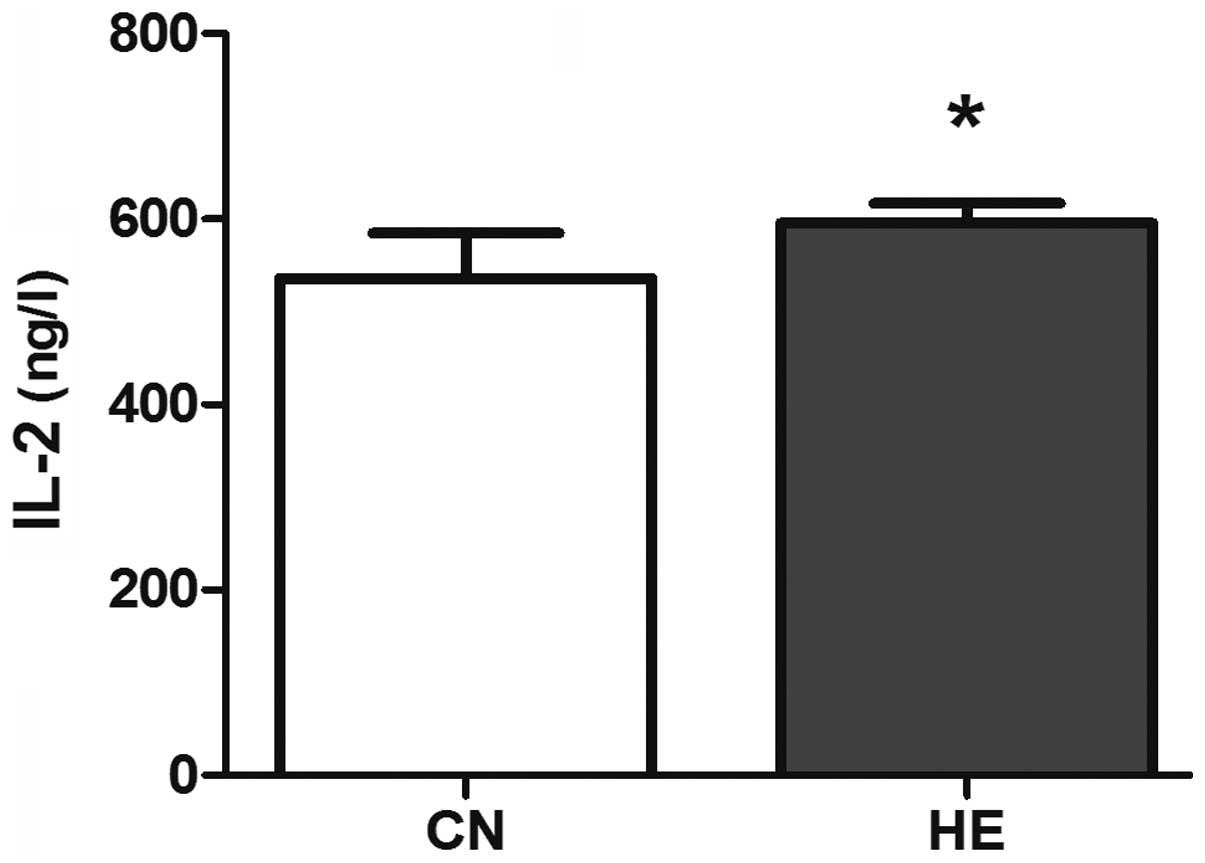
of the CN group (P<0.05) (Fig. 7),
and the level of IL-12 is significantly higher than that of the CN
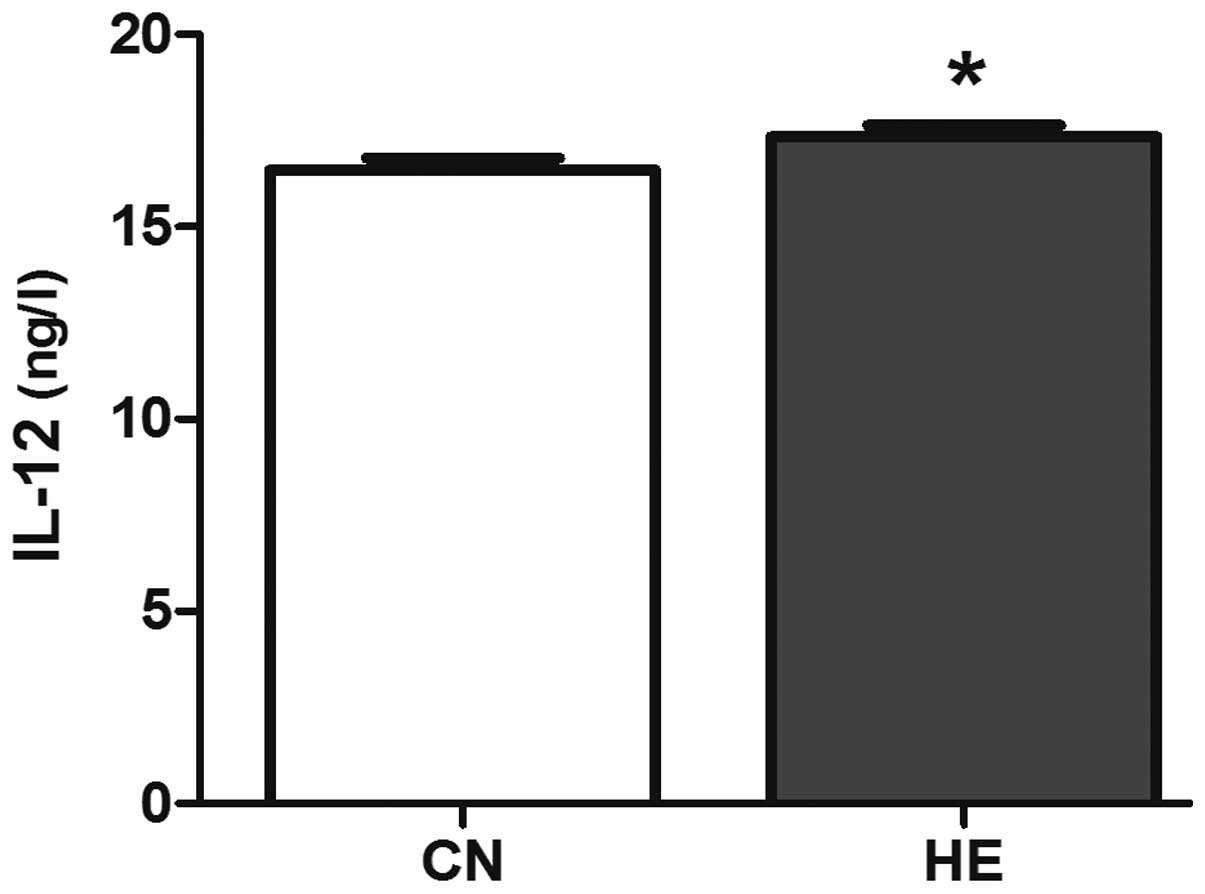
group (P<0.01) (Fig. 8).
Apoptosis gene expression
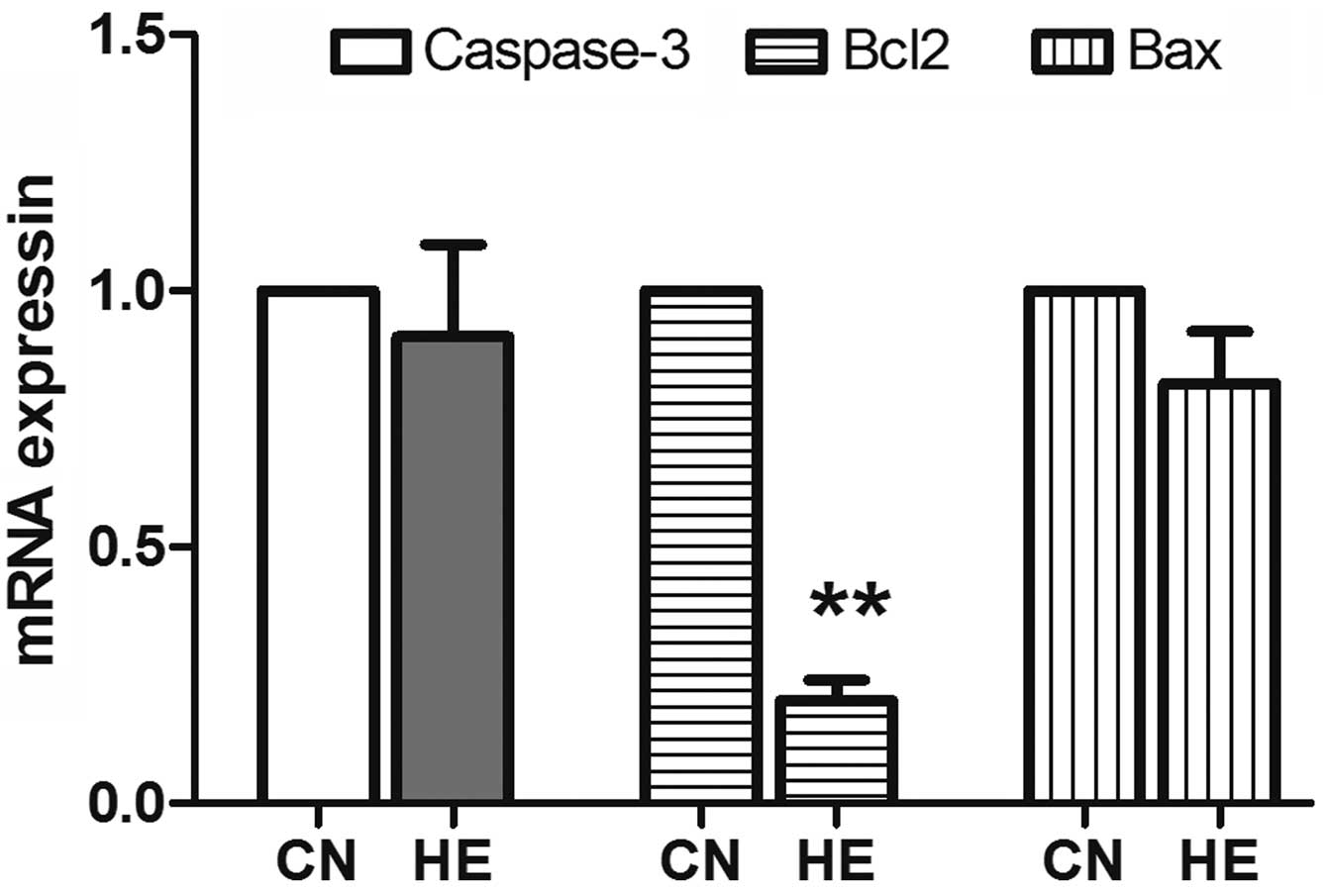
The expression of Bcl2 significantly reduced
(P<0.01) in the HE group, and there was no significant
difference in the reduction of caspase-3 and Bax between the
two groups (P>0.05) (Fig. 9).
Discussion
The present study demonstrated that moderate heat
stress can cause the body's stress response, enhance immune
function and defend against adverse stimulation from the
outside.
As the heat stress occurs, there will be a multiple
axis reaction in the endocrine system (13), such as the HPA, the
hypothalamus-pituitary-thyroid axis or the
hypothalamus-pituitary-gonadal axis. Among them, the main feature
of the stress reaction is the activation of the HPA axis leading to
the increase of ACTH and Cor (14).
ACTH is one of the important pituitary hormones, mainly produced in
the pituitary. The main physiological function of ACTH is promoting
the growth and development of adrenal cortex and stimulating the
synthesis and secretion of glucocorticoid. Cor is the terminal
product of the HPA axis, one of the adrenal cortical synthetic
glucocorticoid (15). Therefore, the
secretion of Cor depends on the integrity of the HPA axis.
Following the activation of the HPA axis, the anterior pituitary
secretes ACTH and the adrenal cortex secretes Cor in a few minutes
to affect the behavior and neuroendocrine activities associated
with the stress. Therefore, ACTH and Cor are often considered as
the most classic and important indicators in studying stress
(16,17). In the present study, the serum ACTH and
Cor levels increased in the HE group, which indicated that the HPA
axis of rats was activated in the process of exposure. The viscera
coefficient is the ratio of viscera weight and body weight. Under
normal circumstances, the viscera coefficient is relatively
constant. The index of the organ coefficient is commonly used in
experiments, as this method is simple and sensitive. The increased
activity of organs leads to the relative hyperplasia to adapt the
heat stress response. Under the environment of the high
temperature, the HPA axis was activated in the stress response in
the present study.
ACTH is released from the pituitary gland and immune
cells. In various immune cells, such as the rat thymus cells,
spleen cells, T cells and B cells, immune ACTH and precursor are
identified indicating that the HPA axis is closely associated with
the cellular immune function. ACTH regulates the production of IL-2
and regulates the immune system by ACTH receptors expressed on
immune cells (18–20). IL-2 is mainly produced by T cells or
the T cell line, and it plays an important role in the immune
adjustment. The level of IL-2 reflects the activity of T cells.
IL-2 as a heat-trapping factor can be applied to the hypothalamus
to cause a high body temperature (21). IL-12 is a cytokine with a broad range
of biological activities, and is mainly produced by the activated
inflammatory cells. IL-12 promotes T cell proliferation, enhances
the activity of NK cells and T cells, and induces tumor necrosis
factor and interferon. In the present study, the serum IL-2 and
IL-12 in the HE group is higher than that of the CN group
suggesting that the body's immune function will increase, due to
the activation of the HPA axis, to adapt the adverse
environment.
The constant body temperature is extremely important
in maintaining the normal function of the human body, as
biochemical cell and enzymatic reactions are affected by the
temperature (22). When the
temperature of the cell is reduced, the metabolic activity and
function will be limited; and by contrast, increased body
temperature enhances the cellular biochemistry. However, when the
body temperature is >42°C, it will cause the denaturation of
intracellular enzymes and other proteins, which leads to cell
damage (23). According to a recent
study, tissue and organ damage occurs during the heat stress
period, particularly the immune organ (24). In general, cell death can be divided
into two patterns, necrosis and apoptosis. Apoptosis plays an
important regulatory role in the body, as it not only ensures the
normal development of the body and maintains a stable internal
environment, but also affects tumorgenesis and development
(25). Caspase-3 is believed to be the
important apoptosis practitioner, as its activation is a sign of
the irreversible apoptosis stage (26). Bcl2 and Bax are important members of
the Bcl2 protein family. Increased Bax promotes cell apoptosis, and
increased Bcl2 inhibits cell apoptosis (27). The present study demonstrated that
under the condition of short-term heat exposure, stimulation is
insufficient to cause the apoptotic program to start and rats can
maintain a constant body temperature. These results indicate that
the cell survival and biochemical reactions were not affected by
the heat exposure process.
The present study showed a significant increase in
the serum concentration of ACTH, Cor, IL-2 and IL-12 of rats
exposed to short-term heat stress. In addition, the organ
coefficients associated with stress were increased and the
apoptosis process did not commence. In conclusion, moderate heat
stress can causes certain beneficial changes and increase the
immune function of the body.
Acknowledgements
The present study was supported by the Ningxia
Natural Science Foundation Key Project (grant no. NZ13055).
References
|
1
|
Shi YJ, Yu JR, Cen XN, Zhu Q and Ren HY:
Influence of HSP70 on combined method of hyperthermia and
immunologic effector cells to treat cancer. Beijing Da Xue Xue Bao.
37:175–178. 2005.(In Chinese). PubMed/NCBI
|
|
2
|
Maglara AA, Vasilaki A, Jackson MJ and
McArdle A: Damage to developing mouse skeletal muscle myotubes in
culture: Protective effect of heat shock proteins. J Physiol.
548:837–846. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim Y, Kim J, Kim M, Baek W and Kim I:
Effect of heat shock on the vascular contractility in isolated rat
aorta. J Pharmacol Toxicol Methods. 42:171–174. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Atkinson HC, Wood SA, Kershaw YM, Bate E
and Lightman SL: Diurnal variation in the responsiveness of the
hypothalamic-pituitary-adrenal axis of the male rat to noise
stress. J Neuroendocrinol. 18:526–533. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dong Q and Liu X: Effect of fear stress in
rats on the contents of ACTH, CORT, IL 2, IL 8. J Mol Diagn Ther.
5:173–176. 2013.
|
|
6
|
Watanabe T, Fujioka T, Hashimoto M and
Nakamura S: Stress and brain angiotensin II receptors. Crit Rev
Neurobiol. 12:305–317. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Padgett DA and Glaser R: How stress
influences the immune response. Trends Immunol. 24:444–448. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fadok VA, Bratton DL and Henson PM:
Phagocyte receptors for apoptotic cells: Recognition, uptake and
consequences. J Clin Invest. 108:957–962. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Urban BC, Willcox N and Roberts DJ: A role
for CD36 in the regulation of dendritic cell function. Proc Natl
Acad Sci USA. 98:8750–8755. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Garrett AT, Creasy R, Rehrer NJ, Patterson
MJ and Cotter JD: Effectiveness of short-term heat acclimation for
highly trained athletes. Eur J Appl Physiol. 112:1827–1837. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brazaitis M and Skurvydas A: Heat
acclimation does not reduce the impact of hyperthermia on central
fatigue. Eur J Appl Physiol. 109:771–778. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fujii N, Honda Y, Ogawa T, Tsuji B, Kondo
N, Koga S and Nishiyasu T: Short-term exercise-heat acclimation
enhances skin vasodilation but not hyperthermic hyperpnea in humans
exercising in a hot environment. Eur J Appl Physiol. 112:295–307.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Cui Y, Yang Y, et al: Study on
neuroendocrine mechanism of occurence of cerebral infarction in
hypertension rats induced by soaring temperature. Prog Mod Biomed.
11:1428–1431. 2011.
|
|
14
|
Hannon MJ and O'Halloran DJ: Isolated
acquired ACTH deficiency and primary hypothyroidism: A short series
and review. Pituitary. 14:358–361. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Beleen C, Martinez Fuentes AJ and Gracia
Navarro F: Role of SST, COR and ghrelin and its receptors at the
endocrine pancreas. Front Endocrinol. 3:1142012. View Article : Google Scholar
|
|
16
|
Servatius RJ, Beck KD, Moldow RL, Salameh
G, Tumminello TP and Short KR: A stress-induced anxious state in
male rats: Corticotropin-releasing hormone induces persistent
changes in associative learning and startle reactivity. Biol
Psychiatry. 57:865–872. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Serra M, Pisu MG, Floris I and Biggio G:
Social isolation-induced changes in the
hypothalamic-pituitary-adrenal axis in the rat. Stress. 8:259–264.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Clarke BL, Gebhardt BM and Blalock JE:
Mitogen-stimulated lymphocytes release biologically active
corticotropin. Endocrinology. 132:983–988. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kravchenco IV and Furalev VA: Secretion of
immunoreactive corticotropin releasing factor and
adrenocorticotropic hormone by T- and B-lymphocytes in response to
cellular stress factors. Biochem Biophys Res Commun. 204:828–834.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wermerskirchen AS, LaTocha DH and Clarke
BL: Adrenocorticotropic hormone controls Concanavalin A activation
of rat lymphocytes by modulating IL-2 production. Life Sci.
67:2177–2187. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Horowitz M: From molecular and cellular to
integrative heat defense during exposure to chronic heat. Comp
Biochem Physiol A Mol Integr Physiol. 131:475–483. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Johnson JS, Boddicker RL, Sanz-Fernandez
MV, Ross JW, Selsby JT, Lucy MC, Safranski TJ, Rhoads RP and
Baumgard LH: Effects of mammalian in utero heat stress on
adolescent body temperature. Int J Hyperthermia. 29:696–702. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Khajavi M, Rahimi S, Hassan ZM, Kamali MA
and Mousavi T: Effect of feed restriction early in life on humoral
and cellular immunity of two commercial broiler strains under heat
stress conditions. Br Poult Sci. 44:490–497. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kanikowska D, Sato M, Sugenoya J, Iwase S,
Shimizu Y, Nishimura N and Inukai Y: No effects of acclimation to
heat on immune and hormonal responses to passive heating in healthy
volunteers. Int J Biometeorol. 56:107–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Johnstone RW, Ruefli AA and Lowe SW:
Apoptosis: A link between cancer genetics and chemotherapy. Cell.
108:153–164. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fesik SW and Shi Y: Structural biology.
Controlling the caspases. Science. 294:1477–1478. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu Y, Zheng Q, Wu H, et al: The effects
on expression ratio of Bax/Bcl-2 and the expression of activated
caspace 3 in different types of tumor cells. Tumor. 33:138–145.
2013.
|