Introduction
The aetiologies and pathogenic mechanisms of
schizophrenia are complicated and likely involve various
neurotransmitter systems. Its clinical manifestations are also
complicated and varied. Currently, its exact pathogenic mechanism
has not been elucidated (1). As the
major atypical antipsychotic drug, risperidone primarily exerts
antipsychotic effects through the blocking of the serotonin
5-HT2A and dopamine D2 (DA2) receptors
(2). Although many clinical and
pre-clinical studies (3–5) have confirmed that the improvements of the
positive and negative symptoms of patients with schizophrenia via
risperidone are associated with the blocking of the 5-HT
2Aand DA2 receptors, the improvement of the
pathogenic mechanism of patients' cognitive symptoms has not been
elucidated (6).
As an important neurotropic factor in the brain, the
brain-derived neurotrophic factor (BDNF) is involved in the
maturation and survival of neurons as well as the growth and
development of synapses. Furthermore, it plays important roles in
synaptic plasticity as well as formation and consolidation of
learning and memory (7). Both clinical
and pre-clinical studies suggested that BDNF and tyrosine receptor
kinase B (TrkB) is extensively involved in the development and
treatment of mental illnesses, including schizophrenia (8). For example, one study (9) showed that risperidone may improve
cognitive impairment in patients through the upregulation of the
functions of the BNDF-TrkB signalling system.
Dizocilpine maleate (MK-801), a non-competitive
N-methyl-D-aspartate (NMDA) receptor, has been applied
extensively in recent animal models of schizophrenia with glutamate
dysfunction (10). Acute treatment of
different doses of MK-801 can induce schizophrenia-like behavioural
changes in rodents, including increases in spontaneous activities,
stereotyped behaviours, sensory gating function damage, and a
variety of abnormal cognitive behaviours (10–12).
Antipsychotics can reverse the above cognitive behavioural
alterations to different degrees; however, the underlying
mechanisms require additional investigation. A previous study
showed that the continuous administration of MK-801 (0.1 mg/kg) for
4 days during the neonatal stage significantly downregulated the
levels of BDNF, TrkB, and p-TrkB in the hippocampus of adult rats
(13). Kim et al also showed
that the continuous intraperitoneal administration of MK-801 (0.6
mg/kg) significantly downregulated BDNF expression in the
hippocampi of mice (14). However, to
the best of our knowledge, few studies have examined the effect of
acute MK-801 injection on the BDNF-TrkB signalling system.
The aims of the current study are two-fold. Firstly,
we investigated whether and how acute MK-801 treatment affected
animals' cognitive performance in the hippocampus-dependent spatial
object recognition task as well as the BDNF-TrkB signalling system
in the hippocampus. Secondly, we examined whether risperidone could
reverse MK-801-induced behavioural and neurobiological
alterations.
Materials and methods
Animals
A total of 52 adult male Sprague-Dawley rats, with a
body weight of 300–350 g at the time of inclusion, were purchased
from the animal laboratory of Jining Medical University (Shandong,
China). The animals were randomly grouped using a random number
table. The animals were housed in the same cage, 4 animals/cage,
and had access to food and water ad libitum. The animals
adapted to the feeding room for ≥7 days prior to the behavioural
experiments. The room temperature was maintained at 23±2°C,
humidity was maintained at 35–55%, and the circadian rhythm was 12
h (from 8:00 a.m. to 8:00 p.m.). Behavioural tests were completed
from 9:00 a.m. to 1:00 p.m. The Ethics Committee of Jining Medical
University approved the present study.
Drugs and pharmacological
procedures
MK-801 and risperidone were purchased from Sigma
(St. Louis, MO, USA). MK-801 was dissolved in normal saline, and
risperidone was dissolved in 0.5% glacial acetic acid saline. Based
on previous literature (15), 0.1
mg/kg MK-801 and risperidone were used. We chose a rather low dose
of MK-801, because it has been established that low-dose MK-801 may
induce cognitive deficits without affecting basic motor abilities,
thus ruling out the potential influence of abnormal peripheral
processes in cognitive performance. Drugs were administered through
intraperitoneal injection, and the drug volumes were both 2 ml/kg.
The control group received an equal volume of normal saline or 0.5
ml/kg glacial acetic acid saline.
Two animal cohorts were used in the
present study
The first cohort included a total of 20 rats (10
animals per group), which were randomly assigned to the control
group (vehicle) or the MK-801 (0.1 mg/kg) group, and received an
intraperitoneal injection of MK-801 or vehicle 30 min before
behavioural testing. After a one-week drug washout period, the
animals received intraperitoneal injections of MK-801 or vehicle
again and 30 min later they were sacrificed, with their brains
removed for western blot analysis. The second cohort comprised 32
animals, which were randomly divided into four groups (8 animals
per group): the control group (vehicle + vehicle), the MK-801 group
(MK-801 + vehicle), the risperidone group (vehicle + risperidone),
and the intervention group (MK-801 + risperidone). The interval
between the drug administration times was 30 min. After drug
treatment, the animals were subjected to behavioural testing or
sacrifice.
Spatial object recognition test
The task was performed as previously reported
(16). Briefly, this task consisted of
the habituation, familiarisation, and test phases. i) Habituation
phase: Before the formal behavioural test, each animal was allowed
to habituate to the test box for 10 min for 3 days to eliminate the
effects of new environmental stimulation on their activity and
exploration behaviours. ii) Familiarisation phase: Two solid
aluminium cubes with a side length of 6 cm were placed in the two
corners (A and B) of the test box. These objects were 9 cm from the
two side walls, and the distance between these objects was 20 cm.
The animals were placed inside, tracked and recorded for 10 min via
video. iii) Recognition phase: After 60 min, the animals were
placed inside the test box again to test for 10 min. During this
phase, the object in corner A or B was moved to corner C. After
each test, the odours were removed by wiping with 75% alcohol, and
the box was dried for the next experimental group. The exploration
time of the animals on each object was evaluated. Following the
scoring criteria reported in previous studies (17), only when the animal's nose or forepaws
were in contact with the object or directed to the object within 1
cm, was this scored as ‘exploring’ the object. Standing, sitting or
leaning on the object was not scored as exploration. The preference
index (PI) of the animals for the object in the new location was
calculated as PI = Tn/(Tn + To) × 100%. In this formula, Tn
represented the exploration time of the object in the new location,
and To was the exploration time of the object in the old location.
Animals with total exploration time of two objects <10 sec were
not included in the data analysis.
Western blot analysis
Thirty minutes after the last drug injection, the
animals were anesthetised using 20% chloral hydrate. After cervical
dislocation, the hippocampal tissues were isolated on ice, quickly
frozen using dry ice, and stored in a freezer at −70°C.
The tissues were removed prior to the experiments
and thawed on ice, placed in a lysis buffer containing a variety of
protease inhibitors, and mechanically lysed on ice. The total
protein concentration was determined using a BCA protein analysis
reagent kit (Pierce Biotechnology, Inc., Rockford, IL, USA). The
amount of protein for loading was calculated. Proteins were
denatured at 95°C for 10 min. Samples containing 30 µg of protein
were subjected to 10% SDS-PAGE. After the electrophoresis was
complete, the proteins on the gel were electro-transferred onto a
PVDF membrane for 90 min. After the electro-transfer was complete,
the membrane was washed with 10 ml of TBST for 2 min and blocked in
a 10 ml TBST solution containing 5% non-fat milk powder for 60 min.
The membrane was incubated with primary antibodies [BDNF: rabbit
anti-BDNF, 1:1,000, Ab108319; Abcam (Cambridge, UK); TrkB: rabbit
anti-TrkB, 1:1,000, sc-8316; Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA); p-TrkB: rabbit anti-phospho-TrkB, 1:2,000,
ABN1381; Millipore Corp. (Billerica, MA, USA): β-actin: mouse
anti-β-actin, 1:20,000, CS-3700; Cell Signaling Technology, Inc.
(Danvers, MA, USA)] at 4°C on a shaker overnight. After washing 3
times with TBST, the membrane was incubated with HRP-labelled goat
anti-rabbit or anti-mouse secondary antibody (Santa Cruz
Biotechnology, Inc.) at 1:5,000 or 1:20,000, respectively. After
incubation at room temperature for 2 h, the membrane was washed
with TBST 3 times. The proteins were exposed in a dark room using
ECL. The exposure time was adjusted according to the fluorescence
intensity, and the images were developed and fixed. The optic
density values of the bands were analysed using Quantity One
software. The optic density values of all the target bands were
normalised based on the optic density values of the corresponding
β-actin bands. All of the data were compared with those in the
control group and presented as percentages (the control group was
set at 100%).
Statistical analysis
The experimental results are presented as means ±
SD. Statistical analyses were performed using SPSS 17.0 (SPSS,
Inc., Chicago, IL, USA). The comparison between the two groups was
performed using an independent samples t-test. The comparison
between the time exploring objects in the novel and familiar
locations was carried out using paired samples t-tests. The one
sample t-test was used to examine whether the PI index in each
group was significantly higher than the chance level, i.e., 50%.
The risperidone intervention experiment analysis was performed
using two-factor analysis of variance (ANOVA). MK-801 (vehicle vs.
MK-801) and risperidone (vehicle vs. risperidone) were used as
between group factors. If the interaction was significant, then the
simple effect analysis was performed. Homogeneity of variance was
examined using Levene's test. If the variance was homogenous, then
the least significant difference (LSD) method was performed for
subsequent tests; if the variance was heterogeneous, then the
Games-Howell method was performed for subsequent tests. P<0.05
was considered to indicate a statistically significant
difference.
Results
Acute MK-801 injection decreases the
spatial object recognition ability of rats
The differences in the activity during the
habituation phase and the total time to explore objects in the
familiarisation phase between MK-801-treated and vehicle-treated
animals were not significant (P>0.05), indicating that acute
injection of MK-801 (0.1 mg/kg) did not significantly affect the
movement ability or the animals' natural exploration of objects.
The variance of all the data were homogeneity (Table I).
 | Table I.Acute MK-801 injection altered the
spatial object recognition ability of rats (means ± SD). |
Table I.
Acute MK-801 injection altered the
spatial object recognition ability of rats (means ± SD).
| Groups | No. of rats | Activity distance
during the habituation phase (m) | Exploration time
during the familiarisation phase (s) | Exploration time for
the objects in new locations (s) | Exploration time for
the objects in old locations (s) | PI of the objects in
new locations (%) |
|---|
| Control | 10 | 42.26±2.54 | 65.05±7.78 | 44.27±6.31 |
20.65±3.97b | 69.20±3.35 |
| MK-801 (0.1
mg/kg) | 10 | 44.07±2.64 | 78.83±8.52 | 43.84±4.70 | 35.12±3.75 |
54.89±3.13a |
During the test phase, a paired samples t-test
showed that the time used to explore objects at new locations by
the animals in the control group was significantly longer than that
used to explore objects at old locations
(t9=4.333, P<0.001). However, the times used
to explore the objects at the new and old locations by the animals
in the MK-801 group did not significantly differ
(t9=1.904, P=0.089). One sample t-test by
comparing PI to 50% further demonstrated that animals in the
control group showed clear preference to the object in the novel
location (t9=5.732, P<0.001), whereas animals
receiving MK-801 treatment failed to discriminate the novel from
the familiar object location (t9=1.566, P=0.152).
The PI for the objects at the new locations between these two
groups was also compared using an independent samples t-test. The
results showed that the PI of the control animals was significantly
higher than that of the MK-801 group (t18=3.122,
P<0.001). The above results indicated that rats in the MK-801
group did not accurately recognize the new or old locations of the
objects and that their spatial object recognition ability was
significantly lower than that of the control group.
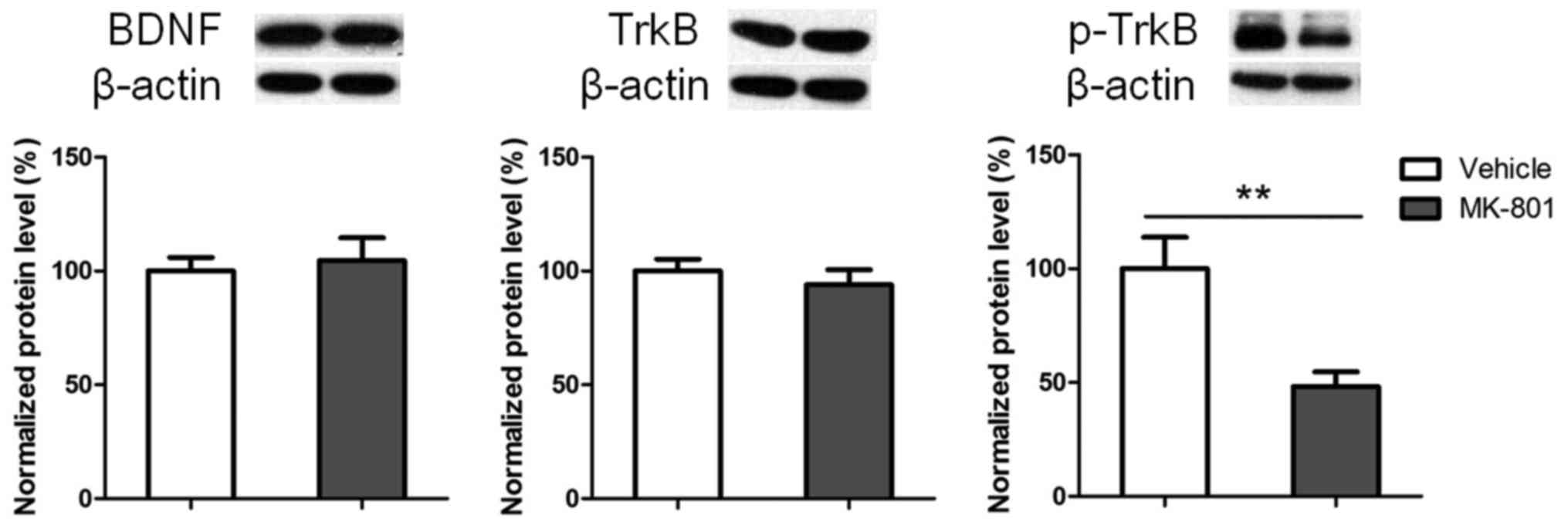
Acute MK-801 injection downregulates
the function of the hippocampal BDNF-TrkB signalling system
Although the acute intraperitoneal injection of
MK-801 did not significantly affect the concentration of BNDF or
the expression of the TrkB receptor in the rat hippocampus
(P>0.05), MK-801 significantly downregulated the expression
level of p-TrkB in the hippocampus (t18=3.385,
P<0.001). p-TrkB is the active form of the TrkB receptor; its
levels can represent the strength of the function of the BDNF-TrkB
signalling system. Thus, these results showed that acute MK-801
injection downregulated the function of the hippocampal BDNF-TrkB
signalling system (Fig. 1).
Risperidone reverses the role that
MK-801 plays in the change in the spatial object recognition
ability of rats
As shown in Table II,
there were no significant differences among the four groups
regarding the distance travelled during the habituation phase and
the total time used to explore the objects during the
familiarisation phase (Ps>0.05), indicating that both MK-801
(0.1 mg/kg) and risperidone (0.1 mg/kg) did not significantly
affect the movement ability or the object preferences of the
animals.
| Table II.Risperidone reversed the role that
MK-801 plays regarding the spatial object recognition ability
changes in rats (means ± SD). |
Table II.
Risperidone reversed the role that
MK-801 plays regarding the spatial object recognition ability
changes in rats (means ± SD).
| Group | Number of rats | Activity distance
during the habituation phase (m) | Exploration time
during the familiarisation phase (s) | Exploration time for
the objects in new locations (s) | Exploration time for
objects in old locations (s) | PI of the objects in
new locations (%) |
|---|
| Control | 8 | 37.54±1.84 | 62.20±6.79 | 42.78±5.23 |
18.43±2.24d |
68.53±4.28c |
| MK-801 | 8 | 37.21±3.13 | 71.33±6.63 | 34.99±4.58 | 33.83±4.47 |
50.78±3.99a |
| Risperidone | 8 | 33.97±3.58 | 54.65±4.15 | 40.23±4.85 |
19.68±2.05d |
66.19±3.46c |
| Intervention | 8 | 36.17±2.38 | 64.21±7.95 | 41.24±6.69 |
21.23±2.52d |
64.95±3.31b |
The results of the two-factor ANOVA regarding the PI
among the groups of animals showed that the main effect of MK-801
(F(1,28)=6.303, P=0.018) and the interaction
between MK-801 and risperidone (F(1,28)=4.762,
P=0.038) were significant, whereas the main effect of risperidone
was not significant (F(1,28)=2.448, P=0.129). An
additional simple main effect analysis showed that the PI of the
animals in the MK-801 group for objects at new locations
significantly decreased compared with that of the control group
(P<0.001). The PI value of the intervention group did not
significantly differ from that of the control group (P>0.05) but
was significantly higher than that of the MK-801 group
(P<0.001). One sample t-test by comparing PI to 50% further
demonstrated that animals received MK-801 treatment only spent
comparable time exploring novel and familiar object positions
(P>0.05); nevertheless, the other three groups showed clear
preference to the object in the novel location (P<0.001). This
was confirmed by a paired samples t-test showing that except for
the MK-801 group (P>0.05), all of the animals in the other
groups spent significantly more time exploring the object in the
novel location (Ps<0.001). The above results indicate that
risperidone reversed the role that MK-801 plays in the spatial
object recognition ability changes of rats.
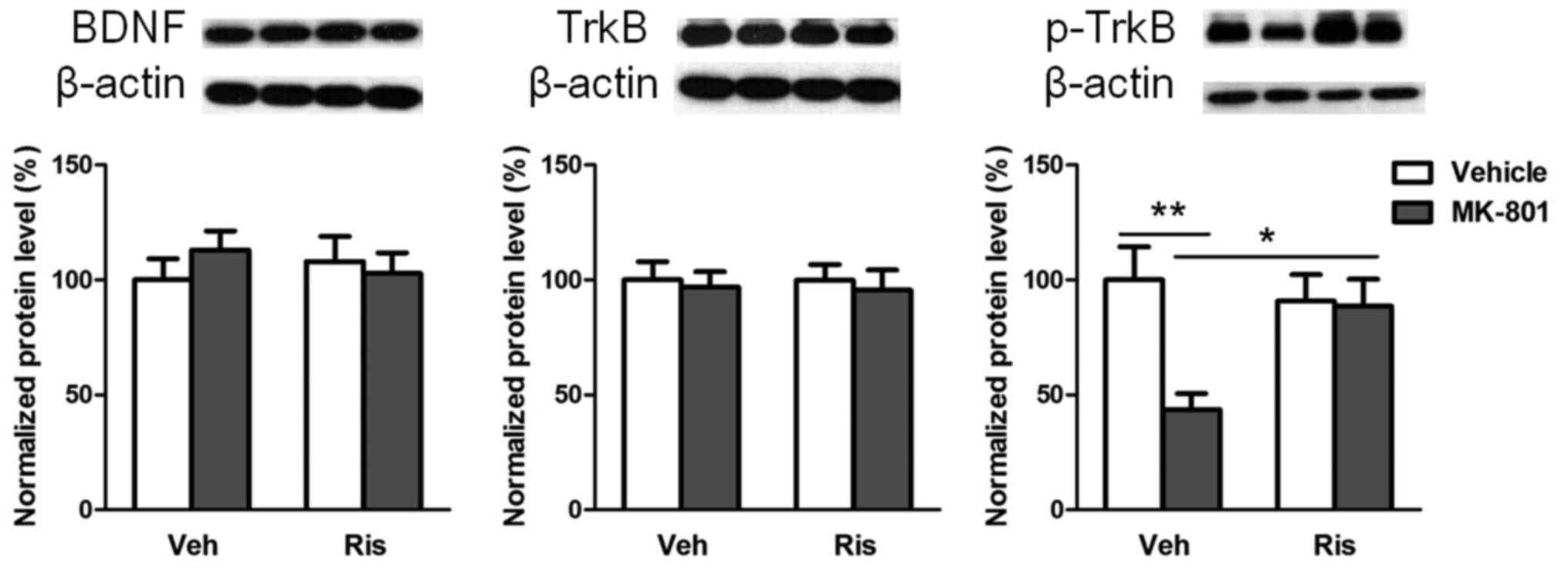
Intervention of risperidone on the
functional changes to the BDNF-TrkB signalling system in the rat
hippocampus caused by MK-801
A two-factor ANOVA showed that the main effects of
MK-801 and risperidone as well as their interaction were not
significant with regard to the expression levels of BDNF and TrkB
(Ps>0.05). However, the main effect of MK-801
(F(1,28)=6.552, P=0.016) as well as the
interaction between MK-801 and risperidone
(F(1,28=5.550, P=0.026) were significant with
regard to the p-TrkB expression level, whereas the main effect of
risperidone was not significant (F(1,28=2.423,
P=0.131). An additional simple main effect analysis showed that the
hippocampal p-TrkB level of the MK-801 group was significantly
decreased compared with that of the control group (P<0.01). The
p-TrkB level of the intervention group did not significantly differ
from that of the control group (P>0.05), but it was
significantly higher than that of the MK-801 group (P=0.010). The
above results indicate that risperidone reversed the role that
MK-801 plays in the downregulation of p-TrkB in the rat hippocampus
and repaired the function of the BDNF-TrkB signalling system
(Fig. 2).
Discussion
The present study used acute intraperitoneal
injection of the NMDA receptor antagonist MK-801 to establish an
animal model of schizophrenia. First, the results showed that an
injection at 0.1 mg/kg deteriorated the spatial object recognition
ability of adult male rats and modelled the reduction of spatial
learning and memory abilities in patients with schizophrenia. These
results have excellent face validity. Additionally, the acute
intraperitoneal injection of MK-801 downregulated the function of
the BNDF-TrkB signalling system in rat hippocampus. This finding is
consistent with the reduction of BDNF levels in the cerebrospinal
fluid of patients with schizophrenia and the reduction of the BDNF
protein and mRNA levels in cognition-related brain areas such as
the prefrontal cortex and hippocampus. These results support the
neurotrophic factor hypothesis of schizophrenia and showed good
construct validity. Finally, the atypical antipsychotic drug
risperidone (0.1 mg/kg) significantly reversed the aforementioned
behavioural and molecular changes caused by MK-801 and showed good
predictive validity. These results provide a basic theoretical
basis for the improvement of cognitive impairment in patients with
schizophrenia via the atypical antipsychotic drug, risperidone.
It has been previously confirmed that the acute and
chronic administration of NMDA receptor antagonists damages the
learning and memory ability of rodents (10,12,18–20).
Consistent with results of the present study, Nilsson et al
(19) and Rogóż and Kamińska (20) showed that the intraperitoneal
administration of MK-801 (at 0.1 and 0.2 mg/kg) 30 min prior to the
test during the familiarisation phase deteriorated the recognition
abilities of rats regarding novel objects. However, to the best of
our knowledge, no report has examined the effect of the acute
intraperitoneal injection of MK-801 on the spatial object
recognition ability of rats. To the best of our knowledge, this
study was the first to show that the acute peritoneal injection of
MK-801 deteriorates the spatial object recognition ability of rats.
This finding adds to the behavioural phenotype of this model.
The function of risperidone regarding the
improvement of the cognitive symptoms of patients with
schizophrenia remains controversial (21). Takekita et al performed a
6-month randomised, controlled, and double-blinded trial and found
that risperidone significantly improved cognitive skills such as
attention and verbal fluency (6). A
pre-clinical study also showed that the continuous intraperitoneal
administration of risperidone (0.2 mg/kg) for 14 days reversed the
spatial learning, memory, and attention deficits caused by MK-801
in rats (5). Rogóż and Kamińska found
that a low dose of risperidone (0.01 mg/kg) did not reverse the
changes in object recognition ability caused by MK-801; however, a
higher dose of risperidone (0.1 mg/kg) successfully reversed the
cognitive deterioration caused by MK-801 (20). The present study showed that
risperidone (0.1 mg/kg) reversed the role that MK-801 played in the
deterioration of the spatial objective recognition ability of rats,
providing pre-clinical evidence for cognitive function improvement
in patients with schizophrenia via the atypical antipsychotic drug
risperidone.
How acute MK-801 treatment may affect hippocampal
BDNF-TrkB signalling system is currently unclear. Hill et al
(22) showed that the acute
intraperitoneal injection of MK-801 (0.05 mg/kg) significantly
downregulated the expression of BDNF protein in the hippocampus
after 24 or 48 h. The results of the present study showed that
after 30 min of acute MK-801 injection, the expression levels of
BDNF and TrkB receptor were not significantly changed in the
hippocampus. This finding may be associated with the different time
points of detection (i.e., immediate vs. long-term effects).
However, the current findings have shown that an acute MK-801
injection significantly downregulated the phosphorylation level of
TrkB after 30 min. As the active form of the TrkB receptor, the
expression level of p-TrkB likely reflects the activity of the
BNDF-TrkB signalling system. Therefore, the acute MK-801 injection
downregulated the function of the BNDF-TrkB signalling system and
partially simulated the pathogenic mechanism of patients with
schizophrenia.
The mechanism that underlies the improvement of
cognitive symptoms in patients with schizophrenia via risperidone
has not been elucidated. However, one study showed that risperidone
may improve cognitive impairment in patients through the
upregulation of the functions of the BNDF-TrkB signalling system
(9). The present study showed that
acute MK-801 injection upregulated the functions of the BNDF-TrkB
signalling system to improve the spatial learning and memory
abilities of rats. However, this study found that risperidone did
not affect the BNDF-TrkB signalling system. A study in China showed
that risperidone upregulated the BNDF-TrkB signalling pathway in
the left and right prefrontal cortex, left and right temporal
cortex, and hippocampus (23). The
differences between these results may be associated with
inconsistent drug doses (0.1 vs. 0.25 mg/kg) and the length of drug
administration (one-time acute administration vs. continuous drug
administration for 14 days).
The present study showed that the atypical
antipsychotic drug risperidone reversed the cognitive behavioural
damage and changes in the hippocampal BDNF-TrkB signalling system
caused by MK-801, suggesting that the insufficiency of the function
of the hippocampal BDNF-TrkB signalling system may be involved in
the development of schizophrenia. However, the mechanism through
which risperidone regulates the BDNF-TrkB signalling system to
improve the cognitive function of patients with schizophrenia as
well as how the BDNF-TrkB signalling system participates in the
development of schizophrenia require additional investigation.
Acknowledgements
The present study was sponsored by the Fund of
Wenzhou Health Bureau (no. 2013B28 to G.D.C) and the Fund of
Tianjin Health Bureau (no. 2014KR02 to C.J.Z).
References
|
1
|
Keshavan MS, Nasrallah HA and Tandon R:
Schizophrenia, ‘just the facts’ 6. Moving ahead with the
schizophrenia concept: From the elephant to the mouse. Schizophr
Res. 127:3–13. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Corena-McLeod M: Comparative pharmacology
of risperidone and paliperidone. Drugs R D. 15:163–174. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Robinson DG, Gallego JA, John M, Petrides
G, Hassoun Y, Zhang JP, Lopez L, Braga RJ, Sevy SM, Addington J, et
al: A randomized comparison of aripiprazole and risperidone for the
acute treatment of first-episode schizophrenia and related
disorders: 3-month outcomes. Schizophr Bull. 41:1227–1236. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Subotnik KL, Casaus LR, Ventura J, Luo JS,
Hellemann GS, Gretchen-Doorly D, Marder S and Nuechterlein KH:
Long-acting injectable risperidone for relapse prevention and
control of breakthrough symptoms after a recent first episode of
schizophrenia. A randomized clinical trial. JAMA Psychiatry.
72:822–829. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kumar V, Ahmad MA, Najmi AK and Akhtar M:
Effect of sarcosine (a glycine transport 1 inhibitor) and
risperidone (an atypical antipsychotic drug) on MK-801 induced
learning and memory deficits in rats. Drug Res (Stuttg). 66:11–17.
2016.PubMed/NCBI
|
|
6
|
Takekita Y, Koshikawa Y, Fabbri C, Sakai
S, Sunada N, Onohara A, Nishida K, Yoshimura M, Kato M, Serretti A
and Kinoshita T: Cognitive function and risperidone long-acting
injection vs. paliperidone palmitate in schizophrenia: a 6-month,
open-label, randomized, pilot trial. BMC Psychiatry. 16:1722016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Waterhouse EG and Xu B: New insights into
the role of brain-derived neurotrophic factor in synaptic
plasticity. Mol Cell Neurosci. 42:81–89. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pandya CD, Kutiyanawalla A and Pillai A:
BDNF-TrkB signaling and neuroprotection in schizophrenia. Asian J
Psychiatr. 6:22–28. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang TL: Effects of antipsychotics on the
BDNF in schizophrenia. Curr Med Chem. 20:345–350. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li JT, Su Y, Yang Y, Feng Z, Huang R and
Si T: Effects of acute treatment of dizocilpine maleate on
locomotor activity, prepulse inhibition and object recognition
memory in rats. Chin J Psychiatry. 45:97–102. 2012.(In
Chinese).
|
|
11
|
Su Y, Si T, Zhou D, Guo C and Shu L:
Establishment of a schizophrenia mouse model of deficient
sensorimotor gating by MK-801. Chin J Nerv Ment Dis. 32:558–560.
2006.(In Chinese).
|
|
12
|
Li J, Su Y, Yang Y, Feng Y, Huang RH and
Si TM: The effects of low-dose MK-801 treatment on reference
memory, spatial working memory and reversal learning in rats. Chin
J Nerv Ment Dis. 37:261–265. 2011.(In Chinese).
|
|
13
|
Yang YJ, Li YK, Wang W, Wan JG, Yu B, Wang
MZ and Hu B: Small-molecule TrkB agonist 7,8-dihydroxyflavone
reverses cognitive and synaptic plasticity deficits in a rat model
of schizophrenia. Pharmacol Biochem Behav. 122:30–36. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim TW, Kang HS, Park JK, Lee SJ, Baek SB
and Kim CJ: Voluntary wheel running ameliorates symptoms of
MK-801-induced schizophrenia in mice. Mol Med Rep. 10:2924–2930.
2014.PubMed/NCBI
|
|
15
|
Su YA, Si TM, Zhou DF, Guo CM, Wang XD,
Yang Y, Shu L and Liang JH: Risperidone attenuates MK-801-induced
hyperlocomotion in mice via the blockade of serotonin 5-HT 2A/2C
receptors. Eur J Pharmacol. 564:123–130. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barker GR, Bird F, Alexander V and
Warburton EC: Recognition memory for objects, place, and temporal
order: a disconnection analysis of the role of the medial
prefrontal cortex and perirhinal cortex. J Neurosci. 27:2948–2957.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bevins RA and Besheer J: Object
recognition in rats and mice: a one-trial non-matching-to-sample
learning task to study ‘recognition memory’. Nat Protoc.
1:1306–1311. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lipina TV, Palomo V, Gil C, Martinez A and
Roder JC: Dual inhibitor of PDE7 and GSK-3-VP1.15 acts as
antipsychotic and cognitive enhancer in C57BL/6J mice.
Neuropharmacology. 64:205–214. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nilsson M, Hansson S, Carlsson A and
Carlsson ML: Differential effects of the N-methyl-d-aspartate
receptor antagonist MK-801 on different stages of object
recognition memory in mice. Neuroscience. 149:123–130. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rogóż Z and Kamińska K: The effect of
combined treatment with escitalopram and risperidone on the
MK-801-induced changes in the object recognition test in mice.
Pharmacol Rep. 68:116–120. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Houthoofd SA, Morrens M and Sabbe BG:
Cognitive and psychomotor effects of risperidone in schizophrenia
and schizoaffective disorder. Clin Ther. 30:1565–1589. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hill XL, Richeri A and Scorza C: Measure
of anxiety-related behaviors and hippocampal BDNF levels associated
to the amnesic effect induced by MK-801 evaluated in the modified
elevated plus-maze in rats. Physiol Behav. 147:359–363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu B, Wang X, Wei JX, Ni PY, Liang LH,
Zhao LS, Ma XH, Li T and Wang YC: Effect of risperidone on the
BDNF-TrkB signalling pathway in rat brain. Sichuan Da Xue Xue Bao
Yi Xue Ban. 46:528–532, 610. 2015.(In Chinese). PubMed/NCBI
|