Introduction
Extracellular vesicles, which consist of lipid
bilayer membranes, are secreted from various cells into mammalian
body fluid, such as blood, saliva and urine (1). Such vesicles are generally classified
into two groups: Exosomes and microvesicles, according to their
size, physical properties and biogenesis (2). Exosomes are small particles with lipid
bilayer membranes, ranging from 40 to 100 nm in diameter. They are
associated with characteristic markers such as CD9, CD63, CD81 and
HSP70 and are derived from the endocytic recycling pathway.
Microvesicles are extracellular particles of 100 to 1,000 nm in
diameter that are produced directly from the plasma membrane as a
consequence of outward budding. Thus, the markers associated with
microvesicles are considered to depend predominantly on the
receptors that are present on the surface of the cells from which
they are derived.
Extracellular vesicles contain proteins, microRNAs,
mRNAs and DNA from their parent cells (3,4), and it
appears that indicated these vesicles are important mediators of
the transfer of information among cells (5). These features have suggested the
potential utility of extracellular vesicles as diagnostic
biomarkers of disease, as already proposed for ovarian cancer, lung
cancer, pancreatic cancer and renal deficiency (6–10). Several
methods have been developed for isolation of extracellular vesicles
from biological samples, for example, ultracentrifugation, gel
dialysis, immunoprecipitation, filtration and density-gradient
centrifugation (11–15), but there is as yet no consensus as to
optimal methods with respect to convenience and purity.
Aptamers are artificial DNA or RNA oligonucleotides
that bind to targets with high affinity and specificity, and were
initially investigated by Ellington and Szostak (16) and Tuerk and Gold (17) and their colleagues in the early 1990s.
Aptamers can be selected using molecular-evolution technology, via
in vitro selection or the systematic evolution of ligands by
exponential enrichment (SELEX). Aptamers have been isolated that
are specific for a large variety of targets, such as small
molecules, nucleotides, peptides and proteins (18). In addition, live cells have been used
as targets for selection of aptamers specific for receptors on cell
surfaces (19–23).
Given that extracellular vesicles are more stable
than cells and contain lipids, sugars and proteins within and/or on
their membranes, as do cells themselves, the authors designed a
procedure for isolation of RNA aptamers that bind to targets on
extracellular vesicles, with the goal of developing new diagnostic
systems. The current study reports the isolation of two aptamers
specific for extracellular vesicles and the description of the
target-specific affinity and structural features of these
aptamers.
Materials and methods
Cell lines and preparation of
extracellular vesicles
293T cells were a generous gift from Professor A.
Fukamizu (Tsukuba University, Tsukuba, Japan). HeLa S3 cells were
obtained from the RIKEN BioResource Center (Tsukuba, Japan). Both
cell lines were maintained in Dulbecco's Modified Eagle's medium
(DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
supplemented with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.). For experiments, cells were transferred to DMEM
without serum and cultured for 2 or 3 days. The resultant
conditioned medium was collected and passed through a 0.22 µm
filter (EMD Millipore, Billerica, MA, USA) to remove cell debris.
Then each sample was loaded into a 10 kDa ultrafiltration cartridge
(Amicon Ultra; EMD Millipore), subjected to these exchanges of
buffer with 40 ml PBS and concentrated to a final volume of 1 ml.
The concentrated sample was placed in a 100 kDa ultrafiltration
cartridge (VIVACON 500; Sartorius AG, Göttingen, Germany) and
subjected to these exchanges of 500 µl PBS. The protein
concentration of each preparation of vesicles was determined in
terms of absorbance at 260 nm using a NanoDrop spectrophotometer
(Thermo Fisher Scientific, Inc., Wilmington DE, USA).
Selection of aptamers
Selection in vitro from two different library
pools was performed as described previously (24–26). The
libraries used are as follows: An N55 library that contained a
region of 55 randomized nucleotides was used [5′-GGG AGG fUGG AAfC
fUGA AGG AGA-(N55)-ACfU fUCG fCAA fUfCG fCfUfC fUAfC GfCA-3′]; an
N30 library that contained a region of 30 randomized nucleotides
[5′-GGfU AGA fUAC GAfU GGA-(N30)-fCAfU G AfC GfCG fCAG fCfCA-3′],
where f represents a 2′-fluoro modification. Selections were
performed in binding buffer [20 mM Tris-HCl (pH 7.5), 150 mM NaCl,
5 mM KCl, 0.5 mM MgCl2 and 1.5 mM CaCl2].
Each pool of RNA was denatured at 98°C for 2 min
and, following cooling, was incubated with extracellular vesicles
in the presence of transfer (t)RNA (100 µg/ml) at room temperature.
The mixture was passed through a 0.45 µm HAWP nitrocellulose filter
(EMD Millipore) and the filter was washed three times with binding
buffer. The RNA that had bound to extracellular vesicles and was
retained on the filter was recovered by incubation with 7 M urea/10
mM EDTA at 98°C for 5 min, extraction with phenol/chloroform and
precipitation in ethanol.
Reverse transcription was performed with a
PrimeScript II first-strand cDNA synthesis kit (Takara
Biotechnology Co., Ltd., Dalian, China). Specifically, preparations
of recovered RNA were supplemented with 1.25 µmol dNTPs and 25 pmol
each reverse primer [reverse primer N55, 5′-TGC GTA GAG CGA TTG CGA
AGT-3′; reverse primer N30, 5′-TGG CTG CGC GTC ATG-3′] then
incubated at 65°C for 5 min. The RNase inhibitor (Takara
Biotechnology Co., Ltd.) and 100 U reverse transcriptase (Takara
Biotechnology Co., Ltd.) were added to the solution, which was
incubated at 30°C for 10 min, 37°C for 10 min, 42°C for 40 min,
52°C for 30 min and 98°C for 5 min.
The cDNA was amplified by polymerase chain reaction
(PCR) (98°C for 10 sec, 55°C for 30 sec and 72°C for 60 sec), with
10 mM Tris-HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 0.2 mM
dNTPs, 0.5 U Ex Taq (Takara Biotechnology Co., Ltd.) and 0.125 µM
each forward primer (N55 forward primer, 5′-TGTA ATA CGA CTC ACT
ATA GGG AGG TGG AAC TGA AGG AGA-3′; N30 forward primer, 5′-TGT AAT
ACG ACT CAC TAT AGG TAG ATA CGA TGG A-3′) and reverse primer (N55
reverse primer, 5′-TGC GTA GAG CGA TTG CGA AGT-3′; N30 reverse
primer, 5′-TGG CTG CGC GTC ATG-3′), and products were precipitated
with ethanol. RNAs, whose pyrimidine residues were 2-fluoro
modified, were transcribed from template DNAs with a
DuraScribe® kit (Epicentre; Illumina, Inc., San Diego,
CA, USA) at 37°C for >6 h or overnight. Transcribed RNAs were
treated with DNase I and purified on micro Bio-Spin columns (P-30;
Bio-Rad Laboratories, Inc. Hercules, CA, USA) or by denaturing on
8% SDS-PAGE gels that contained 7 M urea in Tris/borate/EDTA buffer
(Wako Pure Chemical Industries, Ltd., Osaka, Japan). Then, they
were subjected to the next round of selection.
Sequencing
Sequencing was performed with a MiSeq sequencing
system (Illumina, Inc.) next-generation sequencer and the reagent
kit v2 (Illumina, Inc.) exactly according to the manufacturer's
protocol. The sequences obtained were ranked in order of decreasing
frequency.
Predictions of secondary structure and
search for consensus sequences
Secondary structures were predicted using the
CentroidFold prediction program (National Institute of Advanced
Industrial Science and Technology, Tokyo, Japan) for RNA secondary
folding (27). Consensus sequence were
identified using the motif-discovering algorithm and the MEME Suite
4.10.0 online server (http://meme.nbcr.net/meme/tools/meme), applying the
criterion of a minimum motif length of 8 nucleotides (28).
Minimized oligonucleotides
MO-1 [5′-GGC Cafc gafc gfcg gag gfug fugg ggg afufc
gfuG GCC-3′] and MO-2 [5′-ggfu aga fuafc gafu gga aga ggg aaa ggg
agg gfufu fcfua fcfc-3′] were purchased from Hokkaido System
Science Co., Ltd. (Sapporo, Japan). Uppercase letters indicate
deoxyribonucleic acids and lowercase letters indicate ribonucleic
acids. The abbreviations fu and fc refer to 2-fluoro modified
pyrimidines.
Filter-binding assay
For each assay, a 5′-biotinylated aptamer was
incubated with extracellular vesicles in binding buffer that
contained 100 µg/ml tRNA at room temperature for 2 h. The solution
was passed through a nitrocellulose filter (EMD Millipore) and
washed with 5 ml binding buffer. The filter was exposed to UV
irradiation (254 nm) and was blocked in binding buffer that
contained 3% BSA and 0.1% Tween-20 (Wako Pure Chemical Industries,
Ltd.) for 60 min. Following washing, the filter was incubated with
streptavidin-horseradish peroxidase conjugate (Thermo Fisher
Scientific, Inc.; cat. no. N100; dilution, 1:1,000) for 60 min at
room temperature and washed. Then, the Amersham ECL Prime Western
Blotting Detection reagent (GE Healthcare Life Sciences, Chalfont,
UK) was used to visualize the chemiluminescence of the
5′-biotinylated aptamer on the filter.
Surface plasmon resonance (SPR)
assay
SPR assays were performed with Biacore X™ (GE
Healthcare Life Sciences). Biotinylated oligonucleotides were
immobilized on SA sensor chips (GE Healthcare Life Sciences). Then,
extracellular vesicles were injected at the indicated
concentrations (150, 75, 50, 43 and 38 µg/ml), and running buffer
[binding buffer supplemented with 0.005% Tween-20 (Wako Pure
Chemical Industries, Ltd)] was loaded onto flow cells 1 (blank) and
2 (aptamer) of the sensor chip. Data were analyzed with BiacoreTM
evaluation software (GE Healthcare Life Sciences; version, 2.0).
following subtracting the background (flow cell 1) from values for
flow cell 2.
Prediction of G-quadruplex structure
and measurements of circular dichroism (CD) and melting temperature
(Tm)
The presence of a G-quadruplex structure in
individual aptamers was predicted by the QGRS Mapper (http://bioinformatics.ramapo.edu/QGRS/index.php)
(29).
Circular dichroism (CD) spectra were recorded with a
spectropolarimeter (J-820; Jasco International Co., Ltd., Tokyo,
Japan) as described previously (30).
A cell with a 1 mm path length was used for analyses of titrations
with KCl (at 0 mM, 0.1, 0.3, 1, 5, 10, 50 and 100 mM) in 20 mM
Tris-HCl buffer (pH 7.5). Spectra were recorded four times, from
220 to 320 nm at 25°C. Then, CD intensities were expressed in terms
of [θ] per residue.
Melting temperatures of aptamers were determined at
270 nm in buffer that included 100 mM KCl as the temperature was
raised from 25 to 95°C.
Fractionation of extracellular
vesicles and dot-blotting assay
Preparations of extracellular vesicles were
fractionated on a column of sepharose 2B (GE Healthcare Life
Sciences). Protein concentrations were determined with a DC protein
assay kit (Bio-Rad Laboratories, Inc.) according to the
manufacturer's instructions. Then, 2 µl aliquots of fractionated
and concentrated extracellular vesicles were spotted on a
nitrocellulose membrane, which was dried for 30 min at room
temperature.
For detection of binding of aptamers, the membrane
was blocked with binding buffer that contained 1% BSA and 100 µg/ml
tRNA for 60 min at room temperature. The membrane was then
incubated with 100 µM 5′-biotinylated aptamer in binding buffer
supplemented with 0.1% Tween-20, 1% BSA and 100 µg/ml tRNA for 60
min at room temperature and washed three times with binding buffer
supplemented with 0.1% Tween-20. The membrane was exposed to UV
irradiation (254 nm) and then to streptavidin-horseradish
peroxidase conjugate (Thermo Fisher Scientific, Inc.; cat. no.
N100; dilution, 1:4,000) for 60 min at room temperature. It was
washed three times and, finally, the Amersham ECL Prime Western
Blotting Detection reagent (GE Healthcare Life Sciences; cat. no.
RPN2232) was used to visualize the chemiluminescence of the aptamer
on the membrane.
For detection of antibodies, the membrane was
blocked in PBS supplemented with 0.1% Tween-20 (PBS-T) and
containing 3% BSA for 60 min at room temperature. The membrane was
incubated with human CD63-specific mouse monoclonal antibody (Santa
Cruz Biotechnology, Inc., Dallas, TX, USA; cat. no. Mx-49.129.5;
dilution, 1:1,000) for 60 min at room temperature and washed three
times with PBS-T. The membrane was exposed to second antibody-HRP
conjugate (GE Healthcare Life Sciences; cat. no. NA931; dilution,
1:2,000) for 60 min at room temperature and washed three times.
Finally, the Amersham ECL Prime Western Blotting Detection reagent
(GE Healthcare Life Sciences; RPN2232) was used to visualize
chemiluminescence.
Results
Isolation of RNA aptamers with
affinity for extracellular vesicles
To isolate RNA aptamers with an affinity for
extracellular vesicles, the authors used a selection method in
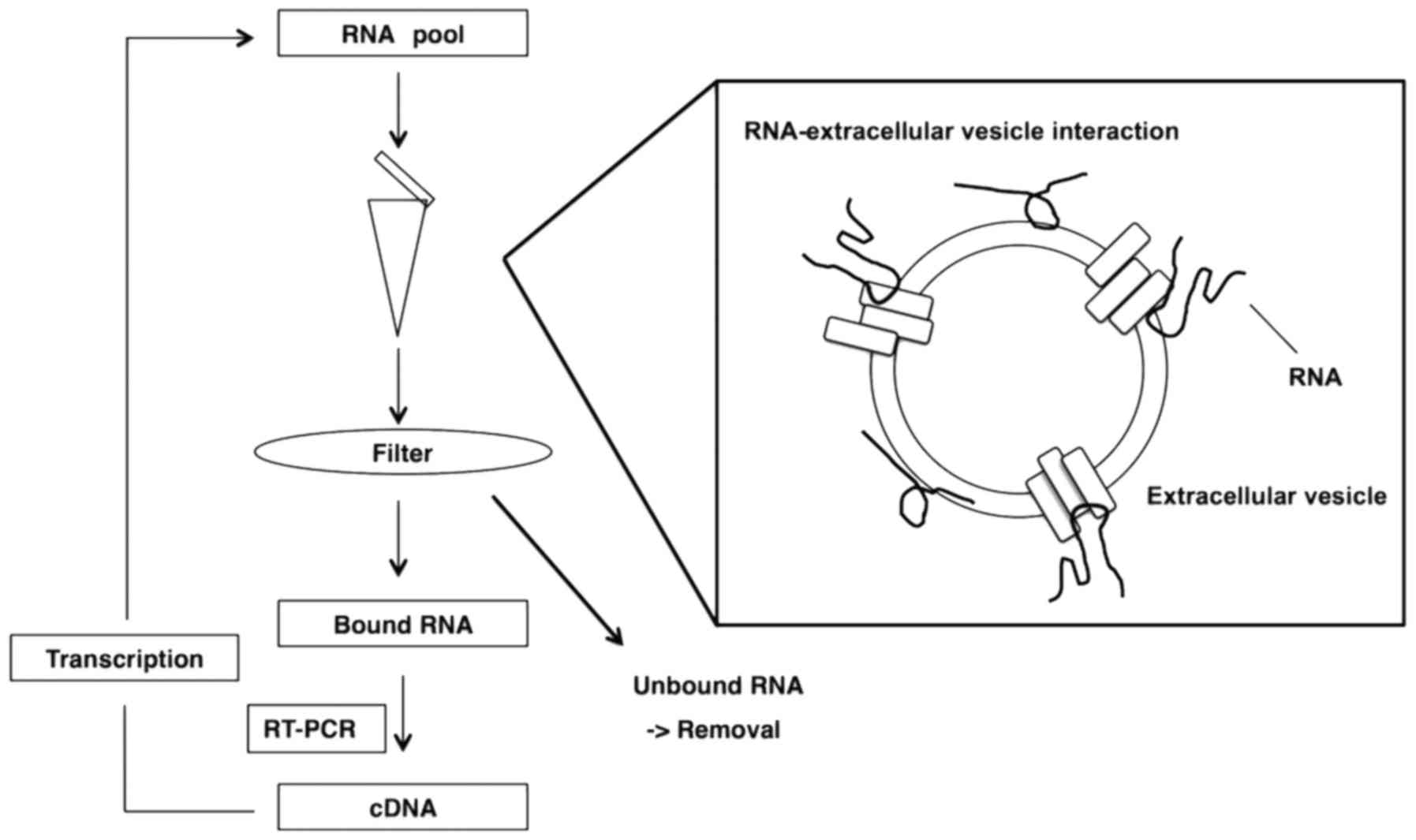
vitro that was based on a previously reported method (Fig. 1) (24–26). A 97-nt
RNA library pool (N55 pool) and a 60-nt RNA library pool (K pool)
was prepared, with respective central randomized 55 and 30-nt
sequences, by in vitro transcription with 2′-fluoro-modified
pyrimidines from synthesized DNA libraries, as templates. Selection
was performed as follows. A transcribed RNA library pool was
incubated with extracellular vesicles in binding buffer for 30 min.
The RNA-extracellular vesicle complexes were added to a
nitrocellulose filter and washed. Bound RNAs were eluted, purified
and subjected to reverse-transcription and amplification. An
enriched RNA pool was transcribed from the amplified products of
PCR and used for the next round of selection.
After 10 rounds of selection, the enriched pools
were sequenced from the eighth, ninth and tenth rounds using a
next-generation sequencer. The 50 most abundant unique sequences
were ranked according to frequency and the results are presented in
Tables I and II.
 | Table I.The sequences obtained from S
pool. |
Table I.
The sequences obtained from S
pool.
| Clone | Number | Sequence
(5′-3′) |
|---|
| S1 | 935 |
ACGCTAGCTGTGGAAAGACGCTAAATCGGGAGGTGGGTTGGGGTGCTAGCGATCA |
| S2 | 375 |
CTAGGGTGGGGAAATATGGAGGGCGCTCAGGGCTACACTGACTACGTGATTAGAG |
| S3 | 365 |
GTCCGCTTCCCGCCACCCGGTGGAAGGAGCCGCGAGGTCACGGAGGTGAGGGGGT |
| S4 | 362 |
TGCTCATTCGTCATAGGTGGAGGGTAGTAGGGGGCGAACAGTTCCACGCTAGCGT |
| S5 | 357 |
GGTGGGTTATAGAGGTTGGGTCCCCGACGTTGCTTCCCTATATAGAGTGGCGAGG |
| S6 | 344 |
TCTCTCGCAGCGGTCTGCATGGTAGCGGGTCAACCACGGAGGAGTGGGGGTGTTG |
| S7 | 310 |
CACGGAGGTGTGGGGGTTCTCAAAACGTTCTATGCTTGGCCAATCCCGAGCCGTC |
| S8 | 223 |
CAGCATTGCGCACGACGCGGAGGTGTGGGGGATCGTGTGGAGAAATGGCTGGCTG |
| S9 | 180 |
ATGGGATGGGAATAGTTCACCACAATTTTTGGGCTCCTTAGCTACACGCGATGGA |
| S10 | 169 |
AGCAACGAAAATTACTGCCGCGGAGGAGAGGGGGAGCAGTATTCGTTGGGTCATA |
| S11 | 137 |
TAGACCCACGCGCATGGCACGGAGGAGAGGGGGACACGCGCAAGGTCTTCACATG |
| S12 | 134 |
AAATTCGCGTTGTAGGTGCACGGTGGTGTGGGGGTTATCCGGCAACGCGTTTGGT |
| S13 | 124 |
GGTGCACCATCTCCGGTCTCGCGCGGTGGAGTGGGGGTTGAGACCGGGGTTGGTT |
| S14 | 107 |
CTTACTCGCAGACACGGAGGTGTGGGGGTTCTGTGGGAGCTTGCACGAGACCTTC |
| S15 | 98 |
TGAGTTAGTGGTGCTGCGCGCGGAGGAGTGGGGGTTGCAGCGTTACGTACGAGTC |
| S16 | 80 |
TGCCGTGTCCGGAATTGCACGGAGGTGAGGGGGATATATTCTGGATAGCGCGCGG |
| S17 | 78 |
TTTGGGGCGTTTCATCAAGATTCGGGTTGGGGACTCTAATAGTAGCCGGTTGTGG |
| S18 | 78 |
ATTTTCGAATCCGCGTCCCGGCGGATAGCACGCGAGGTTACGGCGGAGTGGGGAA |
| S19 | 77 |
CACGGAGGTGTGGGGGTTCTCAAAACGTTCTATGCTTGGCCAATCCCGAGCGGTC |
| S20 | 77 |
ACTGTTGAACGCTCACGTGTACGGAGGTGTGGGGAATACGTGGGGGGTAACATCT |
| S21 | 75 |
CACGGCGGAGTGGGGAGACTCTTGAAATGTTCCGGGTTGCGAACGTCGGTGCTGG |
| S22 | 71 |
CACCGGGGGCAACATGGCGCACGGAGGAGAGGGGGTCGTCTAGTTAGTCCCGGTG |
| S23 | 63 |
TGGCGCGGGTTCGCTGACACGGCGGTGTGGGGGTTCAGGAAACAATGCGCACGAA |
| S24 | 63 |
GGTATTGGGCGGGGGTTCCCAAAAGTAGCGCGACGTCATACGAAGCCGTACGTCG |
| S25 | 62 |
GGCGGAGCACGTATTGACACGGTGGAGAGGGGGTTCATACGGCTCAGGCACCTAA |
| S26 | 56 |
TCGGGACGGCCTGAGGCGCACGGTGGTGTGGGGGTTGCCTACTGCCGTTTGGTCA |
| S27 | 54 |
TCTCTCGCAGCGTCTGCATGGTAGCGGGTCAACCACGGAGGAGTGGGGGTGTTG |
| S28 | 43 |
GAGCGCAATCTAACACGGAGGTGTGGGGGTTTAGCGTGCGCAAACTCAGATGCGT |
| S29 | 38 |
CTCAGTTCATCGTCCCTAGACGCACGGCGGAATTGGGAGTACGGAGGTGGGGGGA |
| S30 | 38 |
ACGAAGTTTAGGATCGCTCCTTCCACGGCGGTGAGGGGGTGAAGCTGCGACCCAG |
| S31 | 38 |
AAGCCGCATCCGGCGGGCGACACGGAGGAGAGGGGGATCGCCACAGCCTGCGGTG |
| S32 | 36 |
TTGGAGGGGGTGTGCCTATTTGGGGTTCAGCAGGGGCACATACGCGGTTGACGAG |
| S33 | 35 |
CGTCAATCACGGCGGTGAGGGGGCATTGATGTCACGCAAAGTAGGCCTAATACCC |
| S34 | 29 |
CACGGAGGAGTGGGGGTTCTCTGGGAAGTTTCGTCTTGTTGGTACGTAACCCGGC |
| S35 | 28 |
AGGCCCGTACGGAGGTGTGGGGATTGGGTCTTTTGACTTGCGAGGCCGTTGTGCG |
| S36 | 27 |
ACGCTAGCTGTGGAAAGACGCTAAATCGGGAGGTGGGTTGGGGTGCTAGCGATCG |
| S37 | 26 |
TGAGGGATGGCCACGGAGGAGTGGGGGTGCCATCGCAACGTTGACACGGGTTGCA |
| S38 | 26 |
GTGTGCCGAGCGATAGGGACACGGTGGAGTGGGGGGACCTATGCCCGGCAAGACA |
| S39 | 26 |
GGGGTCATTAGGAGGGGCCTTTAGAAAAATAGTAGCCGCTGCGGGTCCTTTCGGG |
| S40 | 24 |
ACGCTAGCTGTGGAAAGACGCTAAATCGGGAGGTGGGTTGGGGTGCTAGTGATCA |
| S41 | 21 |
GATTTGGGGGCCACGGAGGTGAGGGGGCGCTCCTATTCTCGTGTTTTGCGTGCGA |
| S42 | 21 |
CGGGCGCCGGGTTAGATCACGGAGGTGTGGGGGTATCTACCCCGTGAGGCGCCAC |
| S43 | 20 |
GTGTTGCACCCGCCTGGGGCACGGAGGTGTGGGGGATCCCAGGGTGAGCGACTAA |
| S44 | 20 |
CTAGGGTGGGGAAATATGGAGGGCGCTCAGGGCTACACTGACTATGTGATTAGAG |
| S45 | 20 |
ATTTTTTTAATGCATCATTTTTACACTCCTTTTGGACCAACCCAACGGGCGCTGC |
| S46 | 19 |
CACTCGGGTTGGAGGCGCACGGCGGTGTGGGGGTCGCTTCAAGTGACGGGTGTCA |
| S47 | 18 |
TGCACAGGTGACACGCCGATCACGGAGGTGTGGGGAGATCGGCAATGGCACGGTG |
| S48 | 18 |
GTCGAAGTGTAACTTATTTGTGTGATTTTTTTGTTTTATGCTTACACGCGGCTCA |
| S49 | 18 |
GGTGGGTTATAGAGGTTGGGTCCCCGACGTTGCTTCCCTATATAGAGTGGCGCGG |
| S50 | 18 |
ATTTTGTTTTGTTGTTTTTGTGTACTCCTTGAGATGCTGGCTACTGTCCGAGCCG |
| Table II.The sequences obtained from K
pool. |
Table II.
The sequences obtained from K
pool.
| Clone | Number | Sequence
(5′-3′) |
|---|
| K1 | 918 |
ATAGCGGGAGGGAGGGTTCTACCTGGTGGG |
| K2 | 692 |
GGGTGGAGGGAAGGAGTGGGGTTCTACCGG |
| K3 | 645 |
AGAGGGAAAGGGAGGGTTCTACCGGGTGCA |
| K4 | 552 |
ACGTGGGAGGGATTGGGGTATCTCCGGTTG |
| K5 | 475 |
AGAGGGTGGGAAAGGGTTCTACCACAGTGC |
| K6 | 471 |
AAGTGGGAGGGGAGGGTTCTACCGGGCCGC |
| K7 | 436 |
GTAGGGAGGTATGTATCTCCTGGTTGGGGG |
| K8 | 403 |
GAGGGAAGGGATATGGGGTATCTAGGGCCG |
| K9 | 367 |
AAGAGGGAGGGTAGGGTTCTACCAGCTGGG |
| K10 | 361 |
GTACGGAGGTGAGGGGAACTCCACGGTCGG |
| K11 | 356 |
GCCCGGCACGGTGGAGAGGGGGTCCGGGGC |
| K12 | 355 |
AGTGGGACTGGGATGGGGTGTATCGCCCGG |
| K13 | 349 |
ATGAGGGAGGGTTGGGGTATCTCCCCGGTG |
| K14 | 338 |
TGAGGGAATAGGGAAAGGGGTATCGTTGGG |
| K15 | 287 |
ACGAGGGAGGGAGGGGTATCACCGGGCCGG |
| K16 | 276 |
GAGGGATGAGGGTGGGCTCTACCTGGCCGG |
| K17 | 271 |
GCCTACGCACGGTGGAGAGGGGGTTGTGGG |
| K18 | 236 |
AGTGGGAAGGGTTTGGGTGGTCTACCGTGG |
| K19 | 230 |
GAGGGAGGGAGGGCTCTACCTTTGTGGCCC |
| K20 | 225 |
CACGGTGGAGTGGGGAGTTCATTGGGCGGG |
| K21 | 222 |
AGTGGGATGGGTAGGGTTCTACATATGCTG |
| K22 | 207 |
GCGGGATTGGGTTTTGGGGTATCTGGGCGG |
| K23 | 205 |
CGGTCAATGCCCACGGTGGAGAGGGGGTGG |
| K24 | 188 |
CCACGGAGGTGAGGGGGTGTCCACGGTGGC |
| K25 | 185 |
TAGAGGGAGGGAGGGATCTACCAGGTGGGG |
| K26 | 174 |
TAGTGGGAGGGAATGGGATTTCTACCGGGG |
| K27 | 170 |
CACTGGAGGAGGGTGGGGAGTTCATCCGGG |
| K28 | 158 |
TTGAGGGAAAGGGTGGGGCATCTACCGTGG |
| K29 | 151 |
CCGTAACACGGAGGAGAGGGGGGAACGGTG |
| K30 | 146 |
CGCGGAGGTGTGGGGGATCCGTCGTGGTGG |
| K31 | 149 |
GAGGGAGGGACTGGGGTATCTTCAGGCGGC |
| K32 | 143 |
ATGAGGGTAGGGAAAGGGGTATCTCGGCGG |
| K33 | 136 |
GGGCACGGTGGAGTGGGGGTCCTTCCTGGG |
| K34 | 132 |
GAGGGTCAGGGATTTGGGGTATCTGGGTGG |
| K35 | 127 |
TTGGAGTGGTGGGTGGGGATCGTGAGGCGG |
| K36 | 124 |
ACACGGTGGTGTGGGGGTTTCCAGGGCGGG |
| K37 | 121 |
CTGTCGGTGCCCACGGTGGAGAGGGGGTGG |
| K38 | 117 |
TGAGGGAGGGACAGGGGTATCTTGGTGCGG |
| K39 | 116 |
CCCTGTCACGGTGGTGTGGGGGTATAGGGC |
| K40 | 114 |
AGAGGGAGGGCAAGGGTTCTACCAGGTCG |
| K41 | 113 |
CCAGCACGCGGTGGAGAGGGGGATGTTGGC |
| K42 | 111 |
TGGCCGATCACGGAGGAGAGGGGGTATCGG |
| K43 | 106 |
GAGGGTTAAGGGACGGGGTATCTGAGGCGG |
| K44 | 97 |
GTGGGATAGGGTTTACGGGGTATCGGTGGG |
| K45 | 94 |
CACGGAGGAGAGGGGGTTCCATCGTTGTCG |
| K46 | 92 |
TTGAGGTGGGATAGGGTAGGGGTCGTGTGC |
| K47 | 92 |
GTGGGAGGGTGGGCTCTGCCAGAACCGGC |
| K48 | 92 |
AGTGGGTTGGGTAATGGGGTATCTACGGGG |
| K49 | 88 |
CACGGTGGAGAGGGGGATCCTACTACTCGG |
| K50 | 88 |
ACAGTGGGAGGGAGGGTTATCACCGGGCCG |
The MEME suite (28)
was used to search for sequence similarities among all 50 sequences
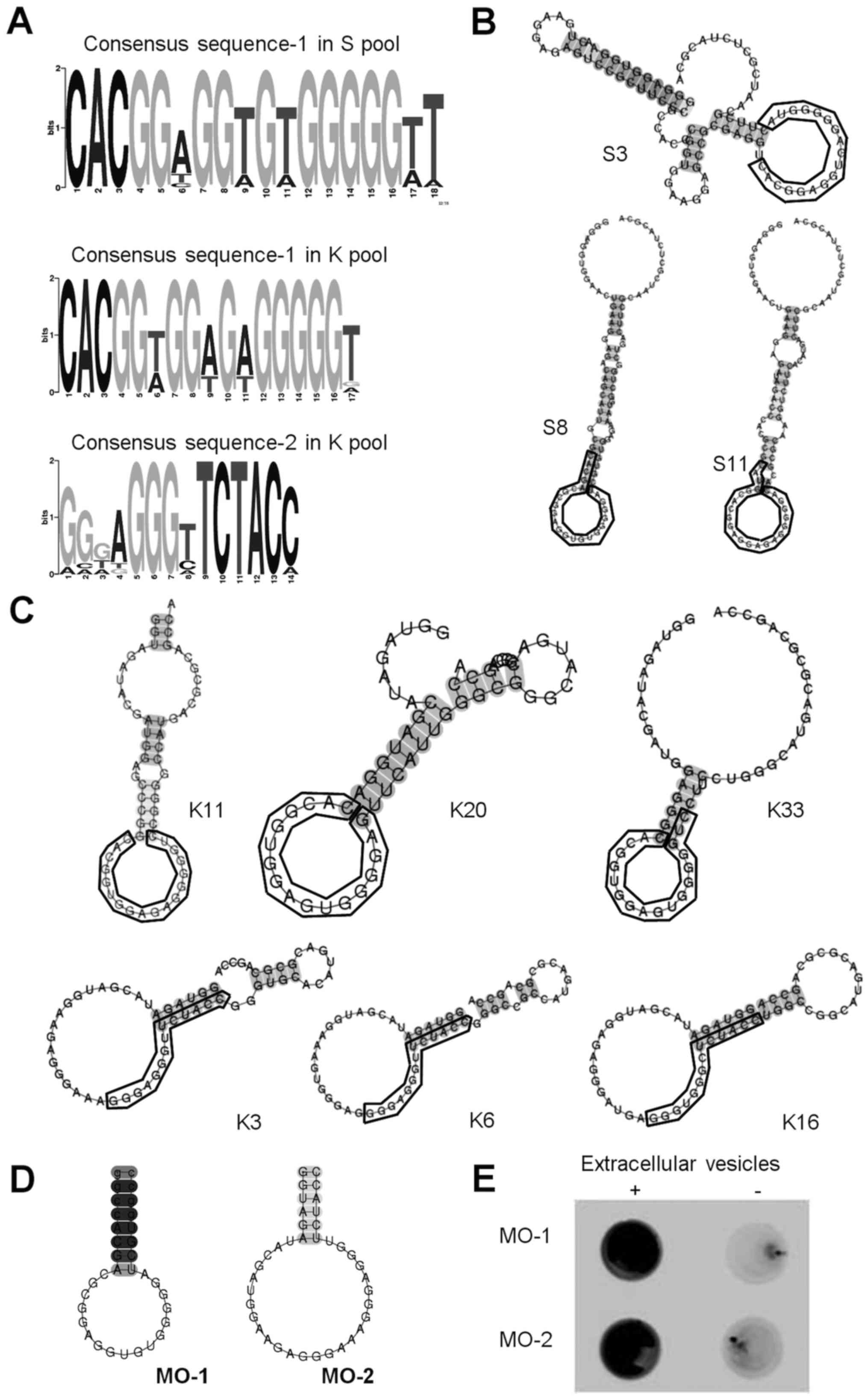
from the respective tenth rounds of enrichment (Fig. 2A). The analysis revealed two common
sequences, the consensus sequence-1 ‘cac gg(t/a) gg(t/a) g(t/a) g
ggg gt’ in the S pool (31 out of 50 clones), the K pool (5 out of
50 clones) and the consensus sequence-2 ‘ggg agg gtt cta cc’ in the
K pool (17 out of 50 clones), respectively. Consensus sequence-1
was identified from both library pools, even through the design of
the two libraries (lengths of randomized region and the fixed
primer regions) were different. The independent isolation of
consensus sequence-1 from the two very different libraries
suggested that the sequence may have strong affinity for
extracellular vesicles.
Next, the secondary RNA structures of the complete
sequences of 53 clones were analyzed (36 clones that included
consensus sequence-1 and 17 clones that included consensus
sequence-2) with primer regions, using CentroidFold software, to
identify common secondary structures (Fig.
2B) (27). All of the sequences
were predicted to have a non-conserved stem with a loop. Some
examples of this structure are presented in Fig. 2C.
Next, the authors attempted to shorten a full-length
sequence to a minimized sequence with the step-loop structure
(Fig. 2D). The sequences of clone S8
and clone K3 were selected as representatives of aptamers with each
respective consensus sequence because both were in the upper ranks
of respective rankings and were likely to be relatively stable as a
result of their longer stems. The authors synthesized minimized
oligonucleotide-1 (MO-1), including consensus sequence-1 referenced
to clone S8, and minimized oligonucleotide-2 (MO-2), including
consensus sequence-2 referenced to clone K3, respectively.
To examine the binding affinities of MO-1 and MO-2
for extracellular vesicles, filter-binding assays were performed
with the respective 5′-biotinylated minimized aptamers. Both
aptamers bound to the extracellular vesicles (Fig. 2E) and not to BSA or mouse IgG (data not
shown), indicating that the minimized sequences had retained
binding affinity and specificity with respect to extracellular
vesicles.
Analysis of interactions of aptamers
with extracellular vesicles
To examine the interaction of the each minimized
aptamer with extracellular vesicles, affinities were measured by
SPR. The 5′-biotinylated minimized oligonucleotides were
immobilized on streptavidin-coated sensor chips. Extracellular
vesicles at various concentrations were then passed over the
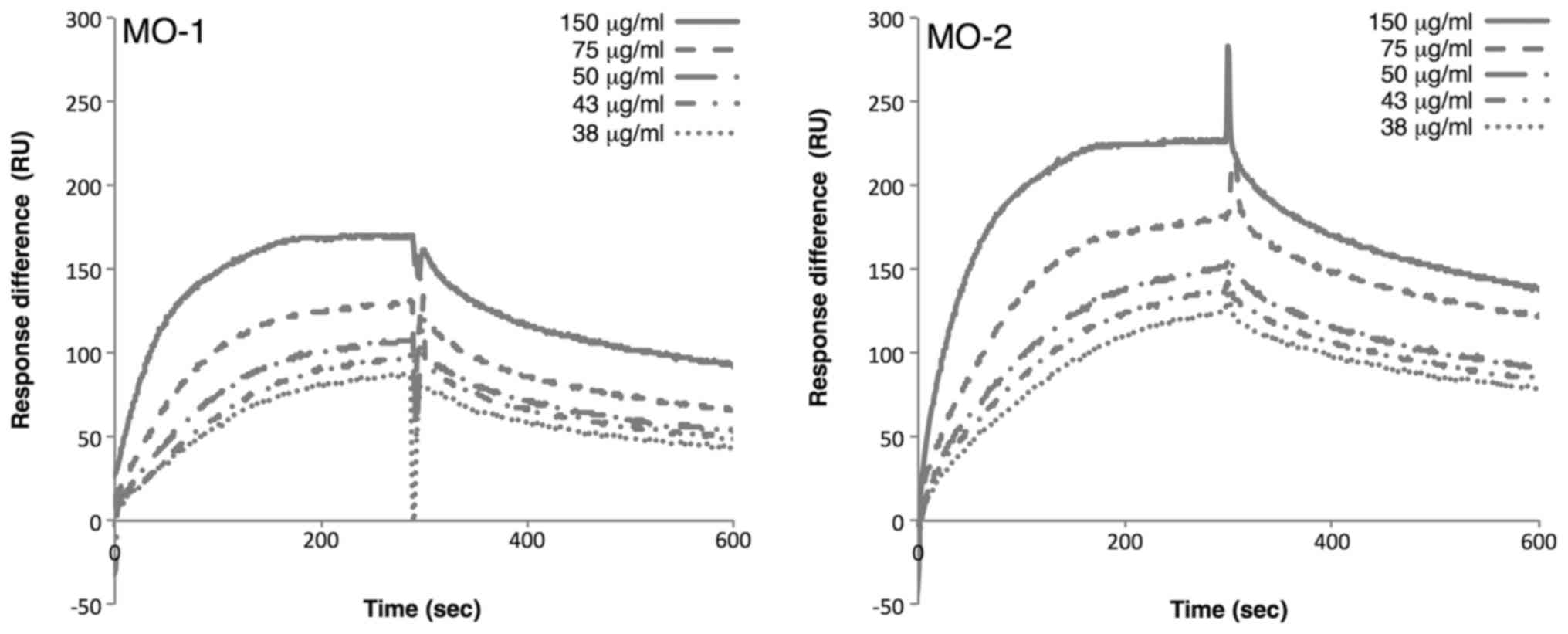
immobilized oligonucleotides. SPR analysis revealed that MO-1 bound
to extracellular vesicles with a KD of 1.17×10–15 mg/ml, while MO-2
bound to extracellular vesicles with a KD of 6.08×10–16 mg/ml
(Fig. 3). These rather similar values
suggested that two aptamers may bind to the same target on
extracellular vesicles.
Since both aptamers bound to extracellular vesicles
derived from 293T cells, the authors examined whether the aptamers
could bind to extracellular vesicles derived from another cell
line. Extracellular vesicles were prepared from HeLa S3 cells and
the SPR analysis was repeated. The results demonstrated that the
MO-1 aptamer bound to HeLa S3 extracellular vesicles with a
KD of 6.83×10−15 mg/ml, and the MO-2 aptamer
bound to the HeLa S3 extracellular vesicles with a KD of
2.82×10−16 mg/ml. These results were similar to those
obtained with 293T-derived extracellular vesicles. Thus, these
observations suggested that the two aptamers may recognize a common
target on extracellular vesicles from a variety of sources.
Spectroscopic analysis of the
structures of minimized aptamers
There are many examples of guanine-rich sequences
folding into G-quadruplex structures in the presence of potassium
ions (31,32). In previous studies of the authors, it
was demonstrated that an RNA aptamer directed against a bovine
prion forms a G-quadruplex structure (25,26,33–35). Since
guanine-rich sequences were present in the MO-1 and MO-2 aptamers,
the authors examined whether the guanine-rich sequences may
resemble the G-quadruplex consensus motif, using the QGRS Mapper,
namely, G-quadruplex analysis software (29). There was a high probability of the
presence of a G-quadruplex structure in both the MO-1 aptamer
(G-score=19) and the MO-2 aptamer (G-score=20).
To confirm these predictions experimentally,
structural and thermodynamic analyses were performed of each
aptamer by circular dichroism (CD) spectroscopy and thermal melting
point (Tm) analysis (Fig. 4A). Both
aptamers yielded a negative band at 240 nm and a positive band at
260 nm in the CD spectrum. The minimum band at 240 nm and the
maximum band at 260 nm were induced by potassium ions in a
concentration-dependent manner (K+ from 0 mM to 100 mM;
Fig. 4A). The Tm values of
the aptamers were relatively high (approximately 70°C; Fig. 4B), indicating that the aptamers were
tightly folded, as expected from a G-quadruplex structure. All of
the results supported the hypothesis that both aptamers were folded
into a G-quadruplex.
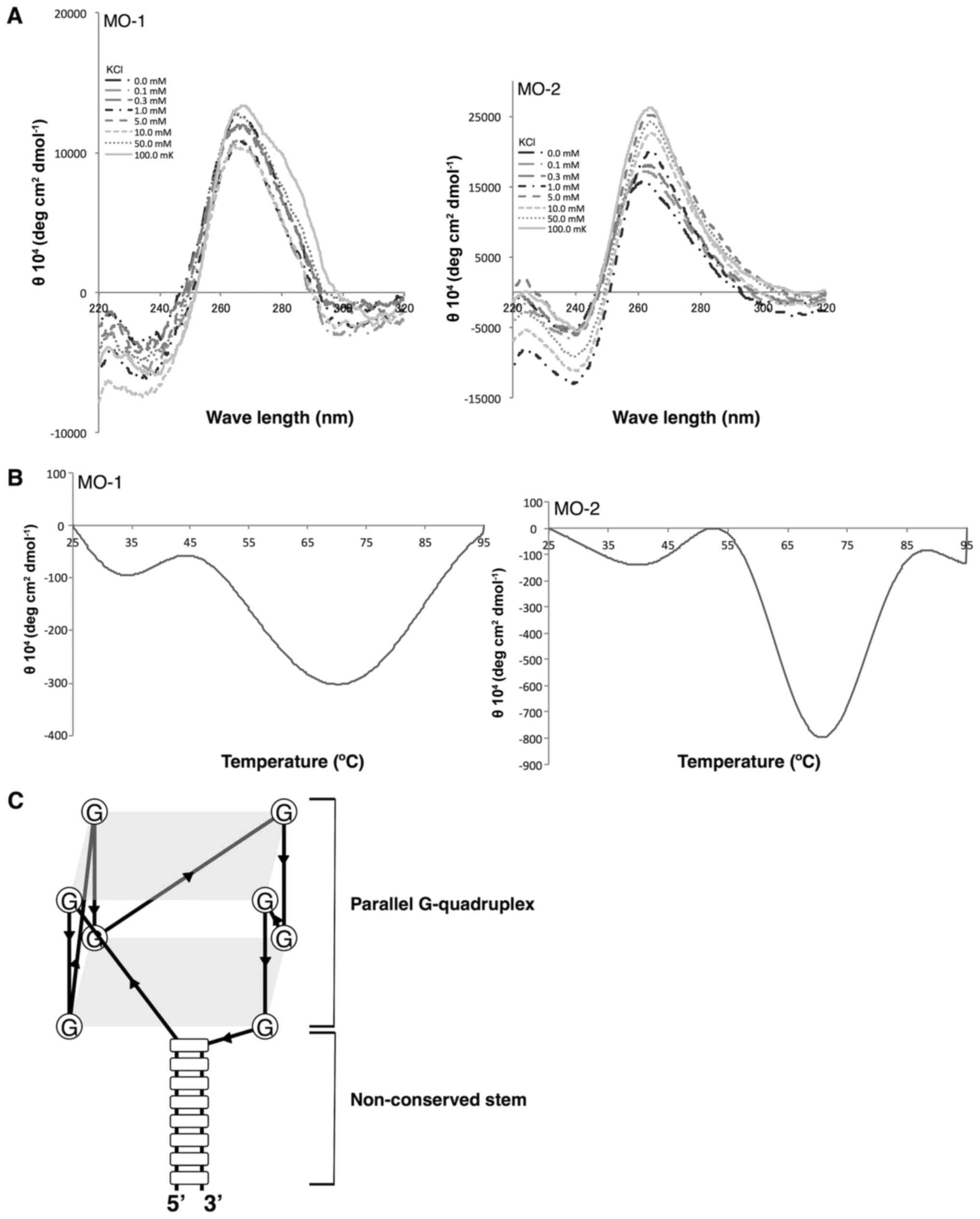 | Figure 4.Analysis of quadruplex structures.
(A) CD spectra of aptamers at various concentrations of KCl (0,
0.1, 0.3, 1, 5, 10, 50 and 100 mM). The aptamers yielded a negative
peak at 240 nm and a positive peak at 260 nm, an indication of the
formation of a parallel quadruplex structure. (B) CD
melting-temperature curves of aptamers at 270 nm in 100 mM KCl. (C)
Putative G-quadruplex structure of the aptamers. The loop region of
the hairpin structure folds into the G-quadruplex structure. CD,
circular dichroism; MO, minimized oligonucleotides; KCl, potassium
chloride. |
G-quadruplex structures have been classified into
two characteristic spectral forms (36) on the basis of their CD spectra: The
parallel G-quadruplex with a maximum at ~260 nm and a minimum at
~240 nm, and the anti-parallel G-quadruplex with a maximum at ~290
nm and a minimum at ~260 nm. Both the MO-1 and the MO-2 aptamers
yielded a maximum at ~260 nm and a minimum at ~240 nm, strongly
suggesting the presence of a parallel G-quadruplex in each one
(25,26,33–35), presented schematically in Fig. 4C.
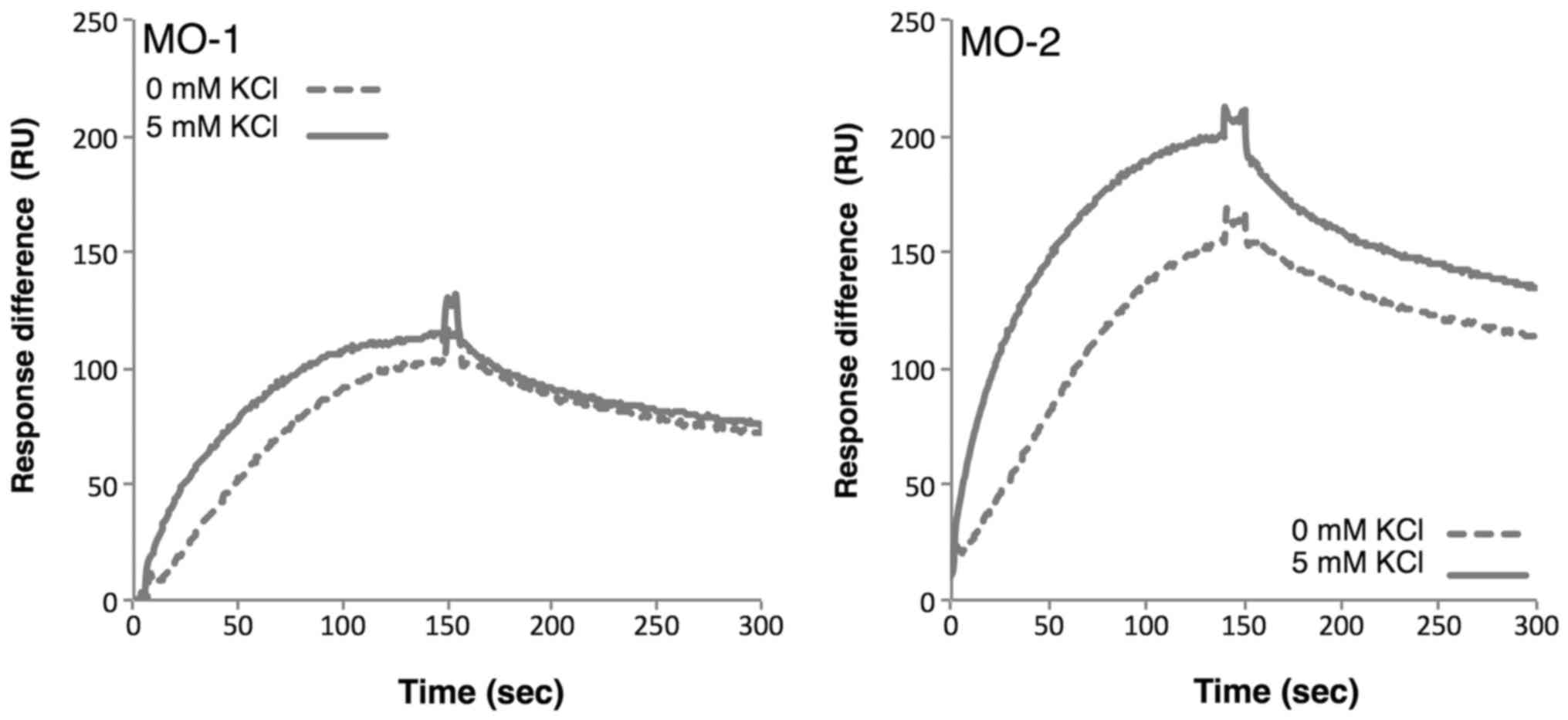
Next, the binding of aptamers to extracellular
vesicles in the presence and absence of potassium ions was
examined. SPR analysis indicated that the binding affinities of
both aptamers increased in the presence of potassium ions (Fig. 5), reflecting the observation that the
G-quadruplex structure is stabilized by potassium ions. The strong
affinities of the aptamers for extracellular vesicles may be
sustained by the tight structure of the G-quadruplex.
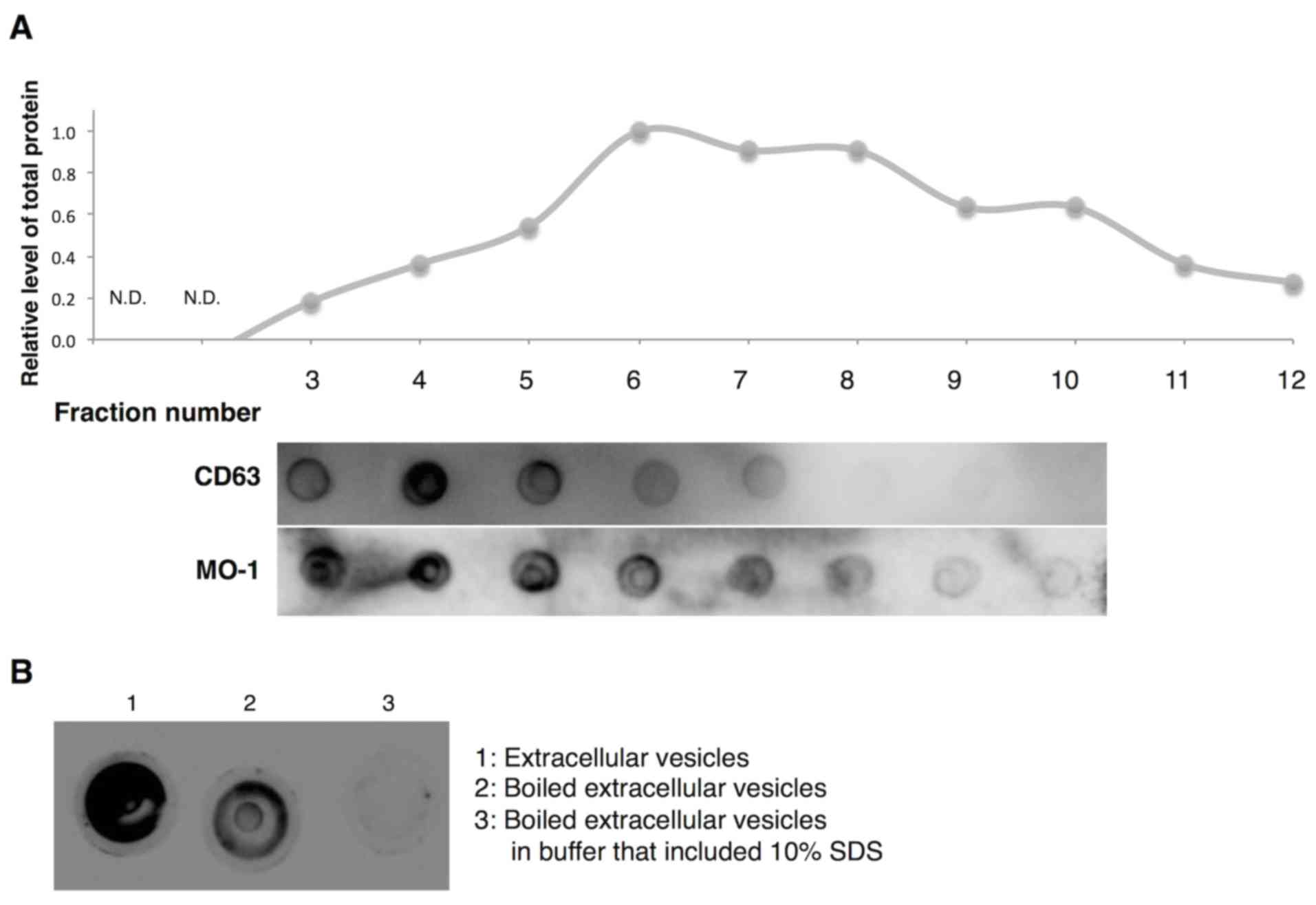
Properties of extracellular vesicles
recognized by aptamer MO-1
Extracellular vesicles, including exosomes and
microvesicles, exist as heterogeneous populations of vesicles of
different sizes, densities and constituent molecules because of
difference in origin and mode of generation (37). Therefore, whether the MO-1 aptamer
recognized all fractions of extracellular vesicles or only some
members of populations of vesicles was investigated. The authors
fractioned extracellular vesicles in terms of size on a sepharose
2B column and monitored the distribution of amounts of total
protein (upper graph in Fig. 6A) and
CD63 protein, a marker of exosomes (middle panel in Fig. 6A). The CD63-specific antibody bound
strongly around fraction 4. By contrast, aptamer MO-1 exhibited
strong affinity for fractions 3 through 6 (lower panel in Fig. 6A). Thus, the aptamer appeared to
recognize the population of vesicles that included a majority of
exosomes.
Also examined was the binding of the MO-1 aptamer to
denatured extracellular vesicles (Fig.
6B). The binding affinity of the aptamer (MO-1) for
heat-denatured and for SDS-denatured extracellular vesicles was
very much reduced. Thus, it seems likely that the aptamer may
recognize a specific protein structure on the extracellular
vesicles.
Discussion
This study is the first, to the best of the authors'
knowledge, to identify RNA aptamers (33-nucleotide MO-1 and
38-nucleotide MO-2) that bind to extracellular vesicles. The
sequences were identified with a next-generation sequencer and each
consisted of a non-conserved stem with a conserved loop (Fig. 2D). SPR analysis revealed that the
aptamers MO-1 and MO-2 had similar strong affinity for
extracellular vesicles (Fig. 3).
Structural analysis by CD spectroscopy demonstrated that both
aptamers formed a G-quadruplex in the loop region (Fig. 4).
The two aptamers MO-1 and MO-2, with consensus
sequence-1, were obtained from two independent libraries, in which
the lengths of the randomized regions and the sequences of fixed
primer regions were completely different (see Materials and
methods). This result may reflect the fact that the aptamers were
effectively and efficiently selected from a huge variety of
randomized sequences. Alternatively, it is possible that the
presence of consensus sequence-1 in both aptamers was due to its
strong affinity and shorter length.
All the candidate aptamers with consensus sequence-1
and consensus sequence-2, respectively, had non-conserved stem
regions. In other words, the regions surrounding the consensus
sequences were different but were able to form stable double-strand
structures. This method allowed the authors to concentrate related
sequences from a massive diversity of libraries sequences
effectively, comprehensively and individually.
Spectroscopic analysis suggested that the MO-1 and
MO-2 aptamers formed a G-quadruplex structure in their loop
regions, and the stable conformation of the G-quadruplex in the
presence of potassium ions may contribute to the binding affinity
of the aptamers for extracellular vesicles (Fig. 5). The stem-G-quadruplex structure of
these aptamers may be one of essential structures for recognition
of target molecules as a binding scaffold with strong affinity and
compact size.
To confirm the interaction between MO-1 and
extracellular vesicles, the authors designed ‘pull-down’
experiments using the 5′-biotinylated MO-1 aptamer. In spite of
several attempts, these experiments failed, presumably because of
difficulties in capturing vesicles of high molecular weight (data
not shown). The binding of MO-1 to size-fractionated extracellular
vesicles suggested that a majority of its targets may be exosomes
but the targets may also be other entities of similar molecular
weight. The SPR analyses demonstrated that extracellular vesicles
derived from HeLa S3 cells also bound to the MO-1 aptamer,
suggesting that the same molecule(s) derived from both 293T and
HeLa S3 cells may be its target (Fig.
3). Moreover, binding affinity was severely depressed under
denaturing conditions (Fig. 6B). Thus,
it is very likely that the targets of the MO-1 and MO-2 aptamers
are proteins on the surface of extracellular vesicles, for example,
CD9, CD13, CD26, CD63, CD81, heat shock protein 70, integrins,
intercellular adhesion molecule-1, major histocompatibility complex
and tumor necrosis factor (38–41).
Previously, a patent was filed for a sequence very
similar to that of MO-1 (42). The
sequence was identified as an aptamer that bound to
platelet-derived growth factor (PDGF). In the patent application,
the claim was made that the sequence was able to bind to PDGF
receptors, to vascular endothelial growth factor and its receptors.
Therefore, the target of MO-1 may be such a ligand or receptor on
extracellular vesicles.
For development of diagnostic technologies that
exploit extracellular vesicles, aptamers have several advantages
over antibodies. The most significant advantage is that
extracellular vesicles can be purified intact, under mild
conditions, from body fluids (11).
Captured vesicles can be eluted from an aptamer-conjugated support
by chelating agents, such as EDTA, which can unfold aptamers by
sequestering magnesium ions. In addition, aptamers can target small
molecules, lipids and sugars (18).
Aptamers can be easily improved by inclusion of modified nucleic
acids or artificial nucleic acids that enhance stability and/or the
ability to bind to nucleic acids. Furthermore, a method using
counter-SELEX (43) may allow
isolation of aptamers against disease-specific extracellular
vesicles via ‘subtractive’ use of disease-derived and normal
vesicles. In such counter-SELEX, the aptamers identified in the
present study, which recognize whole extracellular vesicles, may be
useful in the future as blocking reagents.
It is hoped that the current work will lead to the
development of extracellular vesicle-based diagnostic systems in
which aptamers are used to target disease-specific molecules on
extracellular vesicles.
Acknowledgements
The present study was supported by a Grant-in-Aid
for Challenging Exploratory Research from the Japan Society for the
Promotion of Science (grant no. 26640097). The authors would like
to thank Dr. M. Ikemoto for his helpful comments and Dr. T.
Yamasaki for her skilled technical assistance.
References
|
1
|
Simpson RJ, Jensen SS and Lim JW:
Proteomic profiling of exosomes: Current perspectives. Proteomics.
8:4083–4099. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee Y, El Andaloussi S and Wood MJ:
Exosomes and microvesicles: Extracellular vesicles for genetic
information transfer and gene therapy. Hum Mol Genet. 21:R125–R134.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Valadi H, Ekström K, Bossios A, Sjostrand
M, Lee JJ and Lötvall JO: Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells.
Nat Cell Biol. 9:654–659. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Skog J, Wurdinger T, van Rijn S, Meijer
DH, Gainche L, Sena-Esteves M, Curry WT Jr, Carter BS, Krichevsky
AM and Breakefield XO: Glioblastoma microvesicles transport RNA and
proteins that promote tumour growth and provide diagnostic
biomarkers. Nat Cell Biol. 10:1470–1476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Simons M and Raposo G: Exosomes-vesicular
carriers for intercellular communication. Curr Opin Cell Biol.
21:575–581. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taylor DD and Gercel-Taylor C: MicroRNA
signatures of tumor-derived exosomes as diagnostic biomarkers of
ovarian cancer. Gynecol Oncol. 110:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhou H, Pisitkun T, Aponte A, Yuen PS,
Hoffert JD, Yasuda H, Hu X, Chawla L, Shen RF, Knepper MA and Star
RA: Exosomal Fetuin-A identified by proteomics: A novel urinary
biomarker for detecting acute kidney injury. Kidney Int.
70:1847–1857. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yamashita T, Kamada H, Kanasaki S, Maeda
Y, Nagano K, Abe Y, Inoue M, Yoshioka Y, Tsutsumi Y, Katayama S, et
al: Epidermal growth factor receptor localized to exosome membranes
as a possible biomarker for lung cancer diagnosis. Pharmazie.
68:969–973. 2013.PubMed/NCBI
|
|
9
|
Zoller M: Pancreatic cancer diagnosis by
free and exosomal miRNA. World J Gastrointest Pathophysiol.
4:74–90. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Melo SA, Luecke LB, Kahlert C, Fernandez
AF, Gammon ST, Kaye J, LeBleu VS, Mittendorf EA, Weitz J, Rahbari
N, et al: Glypican-1 identifies cancer exosomes and detects early
pancreatic cancer. Nature. 523:177–182. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thery C, Amigorena S, Raposo G and Clayton
A: Isolation and characterization of exosomes from cell culture
supernatants and biological fluids. Curr Protoc Cell Biol. Chapter
3: Unit 3.22. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rekker K, Saare M, Roost AM, Kubo AL,
Zarovni N, Chiesi A, Salumets A and Peters M: Comparison of serum
exosome isolation methods for microRNA profiling. Clin Biochem.
47:135–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
van der Pol E, Hoekstra AG, Sturk A, Otto
C, van Leeuwen TG and Nieuwland R: Optical and non-optical methods
for detection and characterization of microparticles and exosomes.
J Thromb Haemost. 8:2596–2607. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Grant R, Ansa-Addo E, Stratton D,
Antwi-Baffour S, Jorfi S, Kholia S, Krige L, Lange S and Inal J: A
filtration-based protocol to isolate human plasma membrane-derived
vesicles and exosomes from blood plasma. J Immunol Methods.
371:143–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tauro BJ, Greening DW, Mathias RA, Ji H,
Mathivanan S, Scott AM and Simpson RJ: Comparison of
ultracentrifugation, density gradient separation, and
immunoaffinity capture methods for isolating human colon cancer
cell line LIM1863-derived exosomes. Methods. 56:293–304. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ellington AD and Szostak JW: In vitro
selection of RNA molecules that bind specific ligands. Nature.
346:818–822. 1990. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tuerk C and Gold L: Systematic evolution
of ligands by exponential enrichment: RNA ligands to bacteriophage
T4 DNA polymerase. Science. 249:505–510. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Stoltenburg R, Reinemann C and Strehlitz
B: SELEX-a (r)evolutionary method to generate high-affinity nucleic
acid ligands. Biomol Eng. 24:381–403. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ohuchi SP, Ohtsu T and Nakamura Y:
Selection of RNA aptamers against recombinant transforming growth
factor-beta type III receptor displayed on cell surface. Biochimie.
88:897–904. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Daniels DA, Chen H, Hicke BJ, Swiderek KM
and Gold L: A tenascin-C aptamer identified by tumor cell SELEX:
Systematic evolution of ligands by exponential enrichment. Proc
Natl Acad Sci USA. 100:pp. 15416–15421. 2003; View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wilner SE, Wengerter B, Maier K, de
Lourdes Borba Magalhães M, Del Amo DS, Pai S, Opazo F, Rizzoli SO,
Yan A and Levy M: An RNA alternative to human transferrin: A new
tool for targeting human cells. Mol Ther Nucleic Acids. 1:e212012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li N, Nguyen HH, Byrom M and Ellington AD:
Inhibition of cell proliferation by an anti-EGFR aptamer. PLoS One.
6:e202992011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shigdar S, Lin J, Yu Y, Pastuovic M, Wei M
and Duan W: RNA aptamer against a cancer stem cell marker
epithelial cell adhesion molecule. Cancer Sci. 102:991–998. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sekiya S, Noda K, Nishikawa F, Yokoyama T,
Kumar PK and Nishikawa S: Characterization and application of a
novel RNA aptamer against the mouse prion protein. J Biochem.
139:383–390. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Murakami K, Nishikawa F, Noda K, Yokoyama
T and Nishikawa S: Anti-bovine prion protein RNA aptamer containing
tandem GGA repeat interacts both with recombinant bovine prion
protein and its beta isoform with high affinity. Prion. 2:73–80.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nishikawa F, Murakami K, Matsugami A,
Katahira M and Nishikawa S: Structural studies of an RNA aptamer
containing GGA repeats under ionic conditions using microchip
electrophoresis, circular dichroism and 1D-NMR. Oligonucleotides.
19:179–190. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hamada M, Sato K, Kiryu H, Mituyama T and
Asai K: Predictions of RNA secondary structure by combining
homologous sequence information. Bioinformatics. 25:i330–i338.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bailey TL, Boden M, Buske FA, Frith M,
Grant CE, Clementi L, Ren J, Li WW and Noble WS: MEME SUITE: Tools
for motif discovery and searching. Nucleic Acids Res. 37(Web Server
issue): W202–W208. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kikin O, D'Antonio L and Bagga PS: QGRS
Mapper: A web-based server for predicting G-quadruplexes in
nucleotide sequences. Nucleic Acids Res. 34:W676–W682. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matsugami A, Ouhashi K, Kanagawa M, Liu H,
Kanagawa S, Uesugi S and Katahira M: An intramolecular quadruplex
of (GGA)(4) triplet repeat DNA with a G:G:G:G tetrad and a
G(:A):G(:A):G(:A):G heptad and its dimeric interaction. J Mol Biol.
313:255–269. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Catasti P, Chen X, Moyzis RK, Bradbury EM
and Gupta G: Structure-function correlations of the insulin-linked
polymorphic region. J Mol Biol. 264:534–545. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xia T, SantaLucia J Jr, Burkard ME,
Kierzek R, Schroeder SJ, Jiao X, Cox C and Turner DH: Thermodynamic
parameters for an expanded nearest-neighbor model for formation of
RNA duplexes with Watson-Crick base pairs. Biochemistry.
37:14719–14735. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mashima T, Matsugami A, Nishikawa F,
Nishikawa S and Katahira M: Unique quadruplex structure and
interaction of an RNA aptamer against bovine prion protein. Nucleic
Acids Res. 37:6249–6258. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mashima T, Nishikawa F, Kamatari YO,
Fujiwara H, Saimura M, Nagata T, Kodaki T, Nishikawa S, Kuwata K
and Katahira M: Anti-prion activity of an RNA aptamer and its
structural basis. Nucleic Acids Res. 41:1355–1362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Matsugami A, Mashima T, Nishikawa F,
Murakami K, Nishikawa S, Noda K, Yokoyama T and Katahira M:
Structural analysis of r(GGA)4 found in RNA aptamer for bovine
prion protein. Nucleic Acids Symp Ser (Oxf). 79–180.
2008.PubMed/NCBI
|
|
36
|
Zhang AY, Bugaut A and Balasubramanian S:
A sequence-independent analysis of the loop length dependence of
intramolecular RNA G-quadruplex stability and topology.
Biochemistry. 50:7251–7258. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kowal J, Arras G, Colombo M, Jouve M,
Morath JP, Primdal-Bengtson B, Dingli F, Loew D, Tkach M and Théry
C: Proteomic comparison defines novel markers to characterize
heterogeneous populations of extracellular vesicle subtypes. Proc
Natl Acad Sci USA. 113:pp. E968–E977. 2016; View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Christianson HC, Svensson KJ, van
Kuppevelt TH, Li JP and Belting M: Cancer cell exosomes depend on
cell-surface heparan sulfate proteoglycans for their
internalization and functional activity. Proc Natl Acad Sci USA.
110:pp. 17380–17385. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thery C, Ostrowski M and Segura E:
Membrane vesicles as conveyors of immune responses. Nat Rev
Immunol. 9:581–593. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Mark M Tesic, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Clayton A, Court J, Navabi H, Adams M,
Mason MD, Hobot JA, Newman GR and Jasani B: Analysis of antigen
presenting cell derived exosomes, based on immuno-magnetic
isolation and flow cytometry. J Immunol Methods. 247:163–174. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Epstein D, Grate D, Stanton M, et al:
Stabilized aptamers to platelet derived growth factor and their use
as oncology therapeutics US Patent No US20040253679 A1. April
21–2004, issued December 16, 2004.
|
|
43
|
Jenison RD, Gill SC, Pardi A and Polisky
B: High-resolution molecular discrimination by RNA. Science.
263:1425–1429. 1994. View Article : Google Scholar : PubMed/NCBI
|