Introduction
Non-alcoholic fatty liver disease (NAFLD), which is
characterized by the presence of hepatic steatosis in the absence
of excessive alcohol consumption, is recognized as the typical
hepatic manifestation of metabolic syndrome (1). It has become the most common cause of
chronic liver disease worldwide, and its prevalence has risen
rapidly with the rise of obesity, type 2 diabetes mellitus and
metabolic syndrome epidemics (2,3). NAFLD
represents a wide spectrum of pathological hepatic changes ranging
from simple steatosis to steatohepatitis to different degrees of
fibrosis severity (4). The
pathogenesis of NAFLD, which is not fully understood, involves
several factors, including insulin resistance, alterations of lipid
metabolism, mitochondrial dysfunction and oxidative stress
(5). Since the mechanistic
understanding of and treatment options for NAFLD remain limited,
extensive research efforts have been made to develop novel
therapeutics for the disease (6).
Oxidative stress serves a pivotal role in the
development of NAFLD (7,8). Nuclear factor erythroid 2-related
factor 2 (Nrf2), a cytoprotective transcription factor, serves an
important role in cellular defence against oxidative stress by
interacting with the antioxidant response element (ARE) sequences
of antioxidant and cytoprotective genes (9,10). Under
normal conditions, Nrf2 is localized to the cytoplasm through its
interaction with the actin-bound protein, Kelch-like
epichlorohydrin-associated protein 1 (Keap1). When oxidative stress
occurs, Nrf2 dissociates from Keap1 and translocates to the
nucleus, where it binds to ARE sequences, leading to the expression
of antioxidant enzymes (9,10). During the process of oxidative
stress, activation of the Nrf2/ARE signalling pathway can protect
the liver against oxidative stress by inducing the expression of
antioxidant enzymes, including haeme oxygenase 1 (HO-1) and NAD(P)H
dehydrogenase [quinone] 1 (NQO1) (11,12). For
these reasons, the Nrf2/ARE signalling pathway has been regarded as
an important therapeutic target for the prevention and treatment of
NAFLD (13).
Berberine (BBR) is a type of isoquinoline alkaloid
that was originally isolated from the Chinese medicinal herb
Rhizoma coptidis, which has been extensively used to treat
diabetes mellitus for more than 1,500 years as part of traditional
Chinese medicine (14). BBR has been
reported to have pharmacological effects in lowering blood glucose
(15), reducing blood lipids
(16), improving insulin sensitivity
(17) and reducing inflammation
(18); these effects may contribute
to its efficacy in treating NAFLD. Furthermore, accumulating
evidence from animal models and clinical trials suggests that BBR
may be a potential drug for NAFLD (19,20), but
the underlying molecular mechanisms of the effects of BBR in the
prevention and treatment of NAFLD remain far from fully elucidated.
In the present study, the authors used a NAFLD rat model induced by
a high-fat diet to further investigate whether BBR has a beneficial
effect on NAFLD through the regulation of the Nrf2/ARE signalling
pathway and the amelioration of oxidative stress in the liver.
Materials and methods
Animals
A total of 24 specific pathogen-free male
Sprague-Dawley rats aged 6–7 weeks (220±20 g) were purchased from
the Laboratory Animal Research Center of Guangzhou University of
Traditional Chinese Medicine (Approval No. SYXK (Yue) 2013–0117;
Guangzhou, China). The rats were housed, approximately five per
cage, under conditions of controlled temperature (22–26°C) and
humidity (50–60%), with a 12-h light/dark cycle, and free access to
water and the specified diet.
Experimental design
After 1 week of adaptive feeding, the rats were
randomly divided into three groups of 8 rats each: The normal
control (NC) group, the high-fat diet (HFD) group and the BBR
group. The rat model was established according to a method
previously reported by our group; however, the duration was
adjusted from 12 to 8 weeks (21).
The rats in the NC group had free access to a standard diet (11%
kcal as fat, 20% kcal as protein and 69% kcal as carbohydrate),
while those in the HFD group and BBR group were fed a high-fat diet
(30% kcal as fat, 19% kcal as protein and 51% kcal as
carbohydrate). The two supplements were purchased from Guangdong
Medical Laboratory Animal Center (Guangzhou, China). The rats in
the BBR group were given BBR (Mysun Pharmaceutical Co., Ltd.,
Chifeng, China) at a dose of 100 mg/kg body weight by gastrogavage
each day (22,23), and those in NC and HFD group were
given 10 ml/kg body weight distilled water. The interventions
lasted for 8 weeks. At the end of week 8, all rats were
anesthetized by intraperitoneal injection of 2% pentobarbital (40
mg/kg body weight) and blood samples were then collected from the
abdominal aorta. The blood samples were centrifuged at 1,500 × g
for 10 min at 4°C and the clear supernatants were collected. The
livers were immediately removed and weighed. All rats were treated
in accordance with the Guiding Principles for Animal Experiments
approved by the Animal Experimental Ethics Committee of Jinan
University (Guangzhou, China).
Biochemical analysis
Serum total cholesterol (TC), triglyceride (TG),
high-density lipoprotein cholesterol (HDL-C) and low-density
lipoprotein cholesterol (LDL-C) were measured using an automatic
biochemical analyser (7600-020; Hitachi, Ltd., Tokyo, Japan). The
liver tissues were placed into isopropanol for 15 min at 4°C.
Homogenates were then acquired using a homogenizer (Qiagen GmbH,
Hilden, Germany) and centrifuged at 3,000 × g for 10 min at 4°C.
The clear supernatants were then collected. Liver TC and TG levels
were determined using the automatic biochemical analyser, and
expressed as a value relative to initial sample wet weight.
Histopathological evaluation
Fresh liver tissues were fixed in 10% formalin for
24 h at 4°C or frozen in liquid nitrogen for subsequent
experiments. Formalin-fixed liver tissues were dehydrated in ethyl
alcohol, dealcoholized in xylene, embedded in paraffin, sliced to a
thickness of 5 µm and then stained with haematoxylin and eosin. At
room temperature, liver sections were stained with haematoxylin for
5 min and eosin for 2 min. Frozen liver tissues were embedded in
optimum cutting temperature compound (Sakura Finetek USA Inc.,
Torrance, CA, USA), sliced at a thickness of 8 µm at −18°C and
stained with Oil Red O (cat. no. D027; Nanjing Jiancheng Technology
Co., Ltd., Nanjing, China) for 10 min at room temperature. All
liver sections were observed under a light microscope
(magnification, ×200). The percentage of Oil Red O-stained area was
measured using Image-Pro Plus 6.0 (Media Cybernetics, Rockville,
MD, USA). For transmission electron microscopy (TEM), liver tissues
were fixed with 2.5% buffered glutaraldehyde solution overnight at
4°C and post-fixed with 1% osmium tetroxide solution for 1 h at
4°C, then embedded in Epon812 epoxy resin. Samples were sliced at a
thickness of 80 nm and stained with uranyl acetate for 15 min and
lead citrate for 5 min at room temperature. Images were acquired
using a transmission electron microscope (magnification,
×3,900).
Measurement of oxidative stress
markers in the liver
Liver tissues were homogenized in PBS at 4°C. The
homogenates were then centrifuged at 3,000 × g for 10 min at 4°C
and the supernatants were collected for subsequent examinations.
The protein concentration of supernatants was measured using a
bicinchoninic acid assay (BCA) protein assay kit (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China). The levels of superoxide
dismutase (SOD; cat. no. A001-3), glutathione (GSH; cat. no.
A006-2) and malondialdehyde (MDA; cat. no. A003-1) were measured
using corresponding commercial kits (Nanjing Jiancheng Technology
Co., Ltd.) following the manufacturer's protocol.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from liver tissues using
RNAiso Plus and cDNA was synthesized with PrimeScript RT reagent
kit (both Takara Biotechnology Co., Ltd., Dalian, China) according
to the manufacturer's instructions. Reaction were performed as
follows: 37°C for 15 min, 85°C for 5 sec. qPCR was then performed
using SYBR Premix Ex Taq (Takara Biotechnology Co., Ltd.) using the
CFX96 Touch real-time PCR detection system (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) following the manufacturers' protocol.
Reaction conditions were as follows: 95°C for 30 sec, followed by
40 cycles of 95°C for 5 sec, 60°C for 30 sec and 72°C for 30 sec.
The primers were synthesized by Generay Biotech Co., Ltd.
(Shanghai, China). GAPDH was used as an internal control. The
primer sequences are listed in Table
I. Relative mRNA levels were calculated using the
2−ΔΔCq method (24).
 | Table I.Primer sequences for reverse
transcription-quantification polymerase chain reaction. |
Table I.
Primer sequences for reverse
transcription-quantification polymerase chain reaction.
| Gene | Primer sequence
(5′-3′) | Product (bp) |
|---|
| GAPDH | Forward:
CAACGGGAAACCCATCACCA | 133 |
|
| Reverse:
ACGCCAGTAGACTCCACGACAT |
|
| Nrf2 | Forward:
CTCCTTAGACTCAAATCCCACCTT | 163 |
|
| Reverse:
GGACAGATCACAGCCCTCAAT |
|
| Keap1 | Forward:
AACTCGGCAGAATGTTACTACCC | 190 |
|
| Reverse:
CTACGAAAGTCCAGGTCTCTGTCTC |
|
| NQO1 | Forward:
TCAAGAGGAGCAGAAAAAGAACAAG | 162 |
|
| Reverse:
CTGAAAGCAAGCCAGGCAAAC |
|
| HO-1 | Forward:
ATGAGGAACTTTCAGAAGGGTC | 130 |
|
| Reverse:
GTGGGGCATAGACTGGGTT |
|
Western blot analysis
Western blotting was used to determine the protein
expression levels of Nrf2, Keap1, HO-1, NQO1 and GAPDH. GAPDH was
used as an internal control. Total protein was extracted from the
homogenate with radioimmunoprecipitation assay buffer (Beyotime
Institute of Biotechnology, Shanghai, China) according to the
manufacturer's protocol. Protein concentrations were measured using
bicinchoninic acid Protein Assay kit (Nanjing KeyGen Biotech Co.,
Ltd.). Equal amounts of protein (30 µg/lane) were separated using
10% SDS-PAGE gels and transferred to polyvinylidene difluoride
membranes. Membranes were blocked with 5% skim milk in TBS with
0.1% Tween-20 for 1 h at room temperature, then incubated with
anti-Nrf2 (cat. no. ab137550; 1:500; Abcam, Cambridge, UK),
anti-Keap1 (cat. no. 7705; Cell Signaling Technology, Inc.,
Danvers, MA, USA), anti-HO-1 (cat. no. AJ1338b; Abgent, Inc., San
Diego, CA, USA), anti-NQO1 (cat. no. ab28947; Abcam; all 1:1,000)
or anti-GAPDH (cat. no. KC-5G5; 1:10,000; KangChen BioTech, Co.,
Ltd., Shanghai, China) antibodies overnight at 4°C, followed by an
incubation with horseradish peroxidase-conjugated goat anti-rabbit
antibodies (cat. no. 4050-05; 1:10,000, Southern Biotech,
Birmingham, AL, USA) for 1 h at room temperature. The protein bands
were visualized with enhanced chemiluminescence Plus
electrochemical luminescence reagent (cat. no. P0018; Beyotime
Institute of Biotechnology). Blots were scanned using the ChemiDoc
Imaging system (Bio-Rad Laboratories, Inc.) and then the
densitometric analysis of band intensities was performed with Image
Lab 2.3 (Bio-Rad Laboratories, Inc.).
Statistical analysis
The data are expressed as mean ± standard deviation
and statistically analysed with SPSS 20.0 (IBM Corp., Armonk, NY,
USA). Experiments were repeated ≥3 times. Differences between
groups were analysed using one-way analysis of variance followed by
Bonferroni post-hoc test. P<0.05 indicated that the difference
between groups was statistically significant.
Results
BBR decreases body weight and
decreases liver weight
As shown in Fig. 1,
at the end of the experimental period, the body weight at week 6,
liver weight and the liver/body weight ratio in the HFD group were
significantly higher compared with the corresponding NC group (all
P<0.01). Additionally the body weight at week 4 and 8 in the HFD
group were significantly higher compared with the corresponding NC
group (both P<0.05). Compared with the HFD group, the body
weight at weeks 6 and 8, and liver weight in the BBR group were
significantly decreased (all P<0.01). The liver/body weight
ratio in the BBR group was also decreased compared with the HFD
group, although not in a significant manner. In addition, rats in
the HFD and BBR groups demonstrated decreased food intake, but the
differences were not significant.
BBR ameliorates HFD-induced hepatic
histopathological changes
As shown in Fig. 2A,
histological observations revealed microvesicular steatosis in the
livers of the model rats, as demonstrated by excessive small lipid
droplets inside the cytoplasm and the swelling of hepatocytes
(25). However, there was little
evidence of inflammation. This confirmed the successful
establishment of the NAFLD model by feeding the rats a high-fat
diet for 8 weeks. Rat livers of the NC group exhibited a normal
histological structure with no indication of steatosis and
inflammation. Compared with the HFD group, the histological
structure of rat livers was improved in the BBR group. In addition,
the percentage of the Oil Red O-stained area was significantly
higher in the HFD group when compared with the NC group (P<0.01;
Fig. 2B). Transmission electron
microscopy demonstrated a decreased number of mitochondria in the
HFD group compared with the NC group. In contrast to the HFD group,
the intensity of hepatic steatosis was significantly ameliorated in
the BBR group, as demonstrated by the decreased percentage of the
Oil Red O-stained area (P<0.01; Fig.
2B).
 | Figure 2.BBR ameliorates HFD-induced hepatic
histopathological changes and decreases liver fats. (A)
Representative H&E staining (scale bar, 100 µm), Oil Red O
staining (scale bar, 100 µm) and TEM (scale bar, 10 µm) of the
liver tissue. (B) The percentage of Oil Red O-stained area was
measured. Liver (C) TC and (D) TG levels were assayed. The values
are expressed as mean ± standard deviation. **P<0.01 vs. the NC
group; #P<0.05 and ##P<0.01 vs. the HFD
group. NC, normal control; HFD, high-fat diet; BBR, berberine; TC,
total cholesterol; TG, triglyceride; TEM, transmission electron
microscopy; H&E, haematoxylin and eosin. |
BBR decreases lipid metabolic
parameters
To confirm the role of BBR in lipid metabolism,
serum and liver lipid profiles were assayed. The histopathological
changes revealed that the liver levels of TC and TG were
significantly increased in the HFD group compared with the NC group
(both P<0.01; Fig. 2C and D). By
contrast, the BBR group exhibited significant decreases in the
liver levels of TC (P<0.05) and TG (P<0.01) compared with the
HFD group. As shown in Fig. 3,
compared with the corresponding NC group, serum TC, TG (both
P<0.05) and LDL-C (P<0.01) levels were significantly
increased in the HFD group. The serum HDL-C level was significantly
decreased in the HFD group compared with the NC group (P<0.01).
These results suggested the presence of a lipid metabolism disorder
in the rats with NAFLD. When BBR was administered, the levels of
TC, TG and LDL-C were significantly suppressed while the HDL-C
level was significantly increased compared with the HFD group (all
P<0.05).
BBR increases SOD and GSH, and
decreases MDA
To evaluate the possible role of BBR in hepatic
oxidative stress, SOD, GSH and MDA levels in the liver were
assayed. As shown in Fig. 4, rats in
the HFD group exhibited significant decreases in SOD (P<0.01)
and GSH (P<0.05) levels compared with the NC group. By contrast,
MDA levels in the HFD group were significantly higher compared with
those in the NC group (P<0.01). In the BBR group, SOD
(P<0.01) and GSH (P<0.05) levels were significantly
increased, whereas MDA levels (P<0.05) were significantly
reduced compared with those in the HFD group.
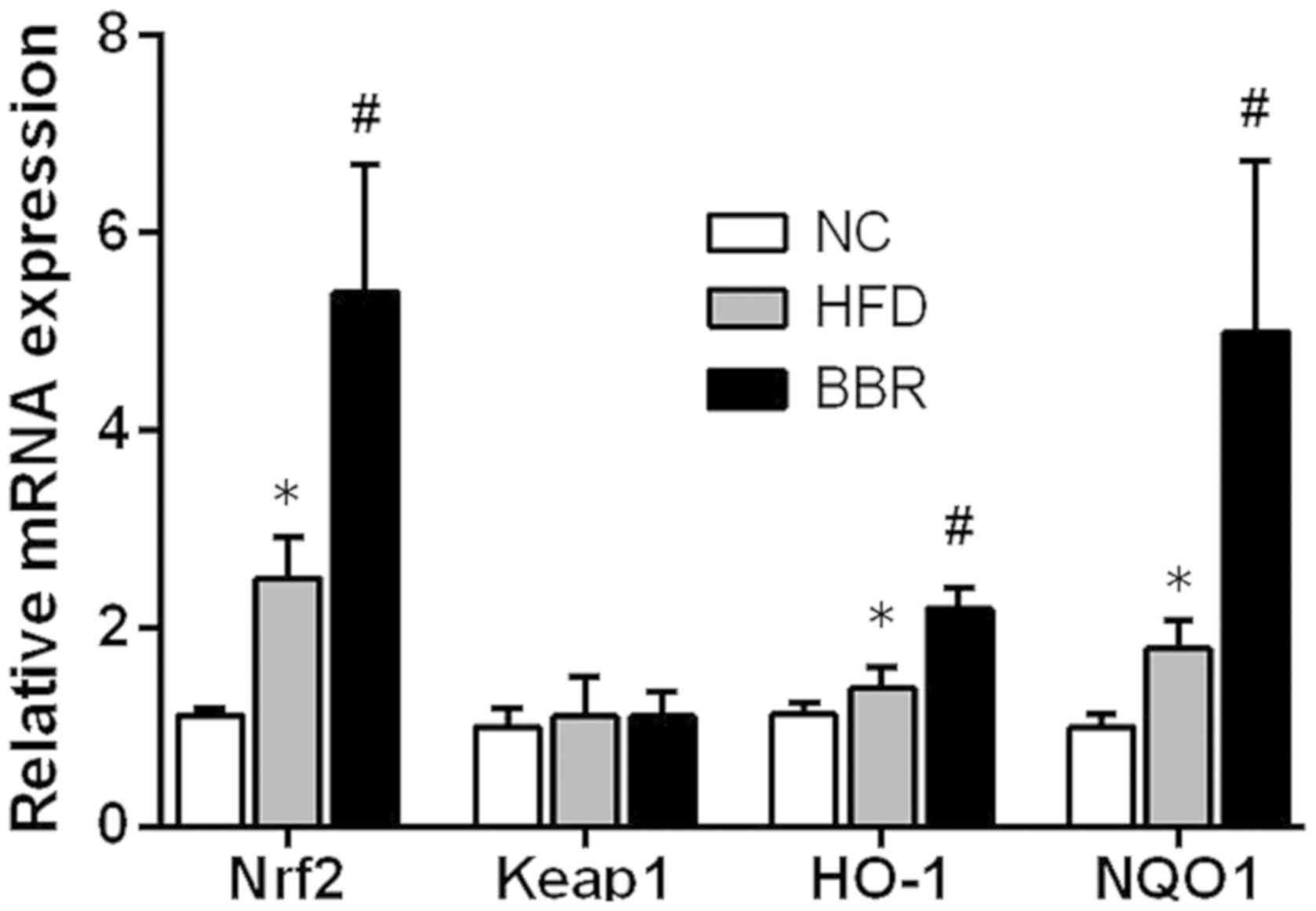
BBR increases the expression of genes
in the Nrf2/ARE signalling pathway in the liver
To further evaluate the role of the Nrf2/ARE
signalling pathway in the development of NAFLD, some key mRNAs and
proteins involved in the Nrf2/ARE signalling pathway were measured.
As shown in Fig. 5, compared with
the NC group, the mRNA levels of Nrf2, HO-1 and NQO1 in the HFD
group were significantly increased (all P<0.05). Western blot
analysis also demonstrated that the expression of HO-1 protein was
significantly increased in the HFD group compared with the NC group
(P<0.01), while the expression levels of Nrf2 and NQO1 protein
were markedly increased compared with the NC group (Fig. 6). Additionally, BBR administration
further increased the mRNA levels of Nrf2, HO-1 and NQO1; they were
significantly increased compared with the HFD group (all P<0.05;
Fig. 5). Western blot analysis
revealed that the protein expression levels of Nrf2, NQO1 (both
P<0.01) and HO-1 (P<0.05) were significantly increased in the
BBR group compared with the HFD group (Fig. 6). No significant changes in Keap1
mRNA and protein levels were observed among the groups (Figs. 5 and 6).
 | Figure 6.BBR increases the protein expression
of genes in the Nrf2/ARE signalling pathway in the liver. (A)
Western blot analysis and (B) relative protein expression of Nrf2,
Keap1, HO-1 and NQO1. The values are expressed as mean ± standard
deviation. **P<0.01 vs. the NC group; #P<0.05 and
##P<0.01 vs. the HFD group. NC, normal control; HFD,
high-fat diet; BBR, berberine; Nrf2, nuclear factor erythroid
2-related factor 2; Keap-1, kelch-like epichlorohydrin-associated
protein 1; HO-1, haeme oxygenase-1; NQO1, NAD(P)H dehydrogenase
[quinone] 1; ARE, antioxidant response element. |
Discussion
In the current study, the authors established a
NAFLD rat model by feeding rats a high-fat diet for 8 weeks. The
authors demonstrated that H&E- and Oil Red O-stained liver
sections from the model group displayed typical hepatic steatosis,
as demonstrated by excessive lipid droplets in the cytoplasm and
the swelling of hepatocytes (21).
Additionally, serum and liver lipid levels were significantly
higher in the model group compared with the normal group. Body
weight and liver weight were also significantly increased in the
model group. Taken together, these results indicate that rats fed a
high-fat diet for 8 weeks can develop the typical signs of hepatic
steatosis and lipid metabolism disorder, suggesting that the
authors of the current study successfully established a rat model
of NAFLD.
Although the pathogenesis of NAFLD remains unclear,
the ‘two-hit theory’ is a widely accepted interpretation of the
pathogenic mechanism of NAFLD. The ‘first hit’ refers to the
accumulation of excessive hepatic fat due to insulin resistance,
while oxidative stress has been recognized as the ‘second hit’ in
the pathogenesis of NAFLD (26,27).
During the ‘second hit’, oxidative stress caused by excess fat
accumulation leads to mitochondrial dysfunction and further liver
cell injury, which contributes to the development of NAFLD
(8). As important indicators of
oxidative stress, antioxidant enzymes, such as SOD, and reactive
species, such as MDA, are often used to assess the level of
oxidative stress in NAFLD (28). In
the current study, the authors observed that rats fed a HFD
exhibited a higher MDA level along with lower SOD and GSH levels.
The results suggested that oxidative stress could be induced in rat
livers after 8 weeks of high-fat feeding.
Over the past decade, a growing body of evidence
suggests that activation of the Nrf2/ARE signalling pathway can
protect hepatic cells from oxidative stress and prevent the
progression of NAFLD (29,30). HO-1 and NQO1 are important
antioxidant enzymes regulated by the Nrf2/ARE signalling pathway
(9). The results of the current
study indicated that the expression of proteins in the Nrf2/ARE
signalling pathway was slightly elevated in the livers of the rats
high-fat diet-induced NAFLD. This finding indicates that Nrf2
activation could be induced by 8 weeks of high-fat feeding, which
is consistent with a previous study (30). Under normal conditions,
Nrf2-dependent transcription is repressed by the regulator Keap1
(10). During exposure to oxidative
stress, Nrf2 dissociates from Keap1 and is therefore not repressed
by Keap1; Nrf2 then translocates into the nucleus where it combines
with ARE sequences, activating the downstream transcription of
antioxidant enzymes (11,13). The oxidative stress induced by
high-fat feeding may activate the Nrf2/ARE signalling pathway,
which is important to cellular antioxidant defence (13). A number of studies have demonstrated
that, in Nrf2-deficient mice, the inability to adapt to hepatic
oxidative stress may hasten the development of NAFLD (31,32).
Nevertheless, adaptive Nrf2 activation induced solely by high-fat
feeding may not be sufficient to avoid the progression of NAFLD in
the absence of an additional Nrf2 activator. Therefore,
pharmacological activation of Nrf2 expression has the potential to
attenuate the onset of fatty liver disease (12,31).
Collectively, these results indicated that Nrf2 serves an important
role in inhibiting the progression of NAFLD, and Nrf2 activators
have the potential to help prevent and treat this disease.
BBR is a natural compound extracted from the dry
roots of Rhizoma coptidis that has been revealed to have
beneficial effects on metabolic disorders, such as diabetes
mellitus (33,34). Several studies have reported that BBR
can reduce body weight, liver weight and serum lipid levels in
animal models (35,36). Similarly, the data from the current
study demonstrated that BBR exhibits similar beneficial effects in
rats with NAFLD induced by a high-fat diet. In the present study,
body weight, liver weight and liver lipids in the BBR group were
significantly decreased compared with the HFD group. The results of
the histopathological evaluation also demonstrated that BBR
markedly alleviated hepatic steatosis. These results are in
agreement with a previous study our group (21). In brief, these data indicate that BBR
successfully slowed down the development of NAFLD in rats through
biochemical and histological improvements.
BBR has been reported to attenuate oxidative stress
through activation of antioxidative pathways (18,37).
Thus, the authors of the current study hypothesized that the
Nrf2/ARE signalling pathway may be involved in the mechanisms of
the protective effect of BBR in high-fat diet-induced NAFLD. The
authors investigated whether BBR can regulate a number of key
proteins of the Nrf2/ARE signalling pathway in the liver. The
results revealed that the liver levels of the Nrf2, HO-1 and NQO1
proteins in the BBR group were significantly higher when compared
with those in the HFD group. These data suggest that BBR
administration in rats fed with a high-fat diet could up-regulate
Nrf2 expression as well as the expression of some genes and
proteins involved in the Nrf2/ARE signalling pathway. As predicted,
the results demonstrated that liver MDA content, the product of
lipid peroxidation during oxidative stress (8), was decreased by BBR co-administration.
Additionally, BBR increased the liver levels of SOD and GSH, which
could help to relieve oxidative stress. This effect may have been
attributable to the up-regulation of the Nrf2/ARE signalling
pathway. In addition, studies have demonstrated that the Nrf2/ARE
signalling pathway may be involved in regulation of hepatic lipid
metabolism (11,31). A number of pharmacological Nrf2
activators have been reported to reduce liver lipid accumulation
and lipogenic gene expression in vivo (38). In the present study, biochemical and
histological examination partly confirmed that Nrf2 overexpression
following BBR administration reduced lipid content and attenuated
hepatic steatosis. Taken together, the findings of the current
suggest that BBR can induce the activation of the hepatic Nrf2/ARE
signalling pathway, and that this effect may contribute to the
amelioration of oxidative stress and its deleterious effects. The
Nrf2/ARE signalling pathway may be an important target for BBR in
the prevention and treatment of NAFLD. However, the precise
mechanisms by which BBR affects the Nrf2/ARE signalling pathway,
oxidative stress and lipid metabolism in the development of NAFLD
requires further clarification.
In conclusion, the current study demonstrated that
BBR may up-regulate the hepatic Nrf2/ARE signalling pathway in rats
fed a high-fat diet, and this effect may be associated with the
amelioration of oxidative stress. Based on these findings, the
authors of the current study concluded that the activation of the
Nrf2/ARE signalling pathway may be one of the important mechanisms
by which BBR exerts its protective effect against NAFLD.
Acknowledgements
Not applicable.
Funding
The present work was supported by Natural Science
Foundation of China (grant nos. 81774165 and 81273617) and
Traditional Chinese Medicine Bureau of Guangdong Province (grant
no. 20152112).
Availability of data and material
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
QY and YZ designed and organized the study. YD, KT,
RC and JZ performed the animal experiments. HN and SL performed the
data analysis. YD and KT wrote the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal
Experimental Ethics Committee of Jinan University (Guangzhou,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BBR
|
berberine
|
|
GSH
|
glutathione
|
|
HDL-C
|
high-density lipoprotein
cholesterol
|
|
HO-1
|
haeme oxygenase-1
|
|
Keap-1
|
kelch-like epichlorohydrin-associated
protein 1
|
|
LDL-C
|
low-density lipoprotein
cholesterol
|
|
MDA
|
malondialdehyde
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
NQO1
|
NAD(P)H dehydrogenase [quinone] 1
|
|
SOD
|
superoxide dismutase
|
|
TC
|
total cholesterol
|
|
TG
|
triglyceride
|
References
|
1
|
Chalasani N, Younossi Z, Lavine JE, Diehl
AM, Brunt EM, Cusi K, Charlton M and Sanyal AJ: The diagnosis and
management of non-alcoholic fatty liver disease: Practice Guideline
by the American Association for the Study of Liver Diseases,
American College of Gastroenterology, and the American
Gastroenterological Association. Hepatology. 55:2005–2023. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Than NN and Newsome PN: A concise review
of non-alcoholic fatty liver disease. Atherosclerosis. 239:192–202.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Farrell GC and Larter CZ: Nonalcoholic
fatty liver disease: From steatosis to cirrhosis. Hepatology. 43 (2
Suppl 1):S99–S112. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Musso G, Cassader M and Gambino R:
Non-alcoholic steatohepatitis: Emerging molecular targets and
therapeutic strategies. Nat Rev Drug Discov. 15:249–274. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Videla LA, Rodrigo R, Araya J and
Poniachik J: Insulin resistance and oxidative stress
interdependency in non-alcoholic fatty liver disease. Trends Mol
Med. 12:555–558. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedman SL, Neuschwander-Tetri BA,
Rinella M and Sanyal AJ: Mechanisms of NAFLD development and
therapeutic strategies. Nat Med. 24:908–922. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sakaida I and Okita K: The role of
oxidative stress in NASH and fatty liver model. Hepatol Res.
33:128–131. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rolo AP, Teodoro JS and Palmeira CM: Role
of oxidative stress in the pathogenesis of nonalcoholic
steatohepatitis. Free Radical Bio Med. 52:59–69. 2012. View Article : Google Scholar
|
|
9
|
Kaspar JW, Niture SK and Jaiswal AK:
Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radical Bio
Med. 47:1304–1309. 2009. View Article : Google Scholar
|
|
10
|
Vomhof-DeKrey EE and Picklo MJ Sr: The
Nrf2-antioxidant response element pathway: A target for regulating
energy metabolism. J Nutr Biochem. 23:1201–1206. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chambel SS, Santos-Goncalves A and Duarte
TL: The dual role of Nrf2 in nonalcoholic fatty liver disease:
Regulation of antioxidant defenses and hepatic lipid metabolism.
Biomed Res Int 2015. 5971342015.
|
|
12
|
Tang W, Jiang YF, Ponnusamy M and Diallo
M: Role of Nrf2 in chronic liver disease. World J Gastroentero.
20:13079–13087. 2014. View Article : Google Scholar
|
|
13
|
Bataille AM and Manautou JE: Nrf2: A
potential target for new therapeutics in liver disease. Clin
Pharmacol Ther. 92:340–348. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yin J, Zhang H and Ye J: Traditional
chinese medicine in treatment of metabolic syndrome. Endocr Metab
Immune Disord Drug Targets. 8:99–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang C, Li J, Lv X, Zhang M, Song Y, Chen
L and Liu Y: Ameliorative effect of berberine on endothelial
dysfunction in diabetic rats induced by high-fat diet and
streptozotocin. Eur J Pharmacol. 620:131–137. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kong W, Wei J, Abidi P, Lin M, Inaba S, Li
C, Wang Y, Wang Z, Si S, Pan H, et al: Berberine is a novel
cholesterol-lowering drug working through a unique mechanism
distinct from statins. Nat Med. 10:1344–1351. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Campbell T, Perry B, Beaurepaire C
and Qin L: Hypoglycemic and insulin-sensitizing effects of
berberine in high-fat diet- and streptozotocin-induced diabetic
rats. Metabolism. 60:298–305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mo C, Wang L, Zhang J, Numazawa S, Tang H,
Tang X, Han X, Li J, Yang M, Wang Z, et al: The crosstalk between
Nrf2 and AMPK signal pathways is important for the
anti-inflammatory effect of berberine in LPS-stimulated macrophages
and endotoxin-shocked mice. Antioxid Redox Sign. 20:574–588. 2014.
View Article : Google Scholar
|
|
19
|
Liu Y, Zhang L, Song H and Ji G: Update on
berberine in nonalcoholic Fatty liver disease. Evid Based
Complement Alternat Med 2013. 3081342013.
|
|
20
|
Yang J, Ma XJ, Li L, Wang L, Chen YG, Liu
J, Luo Y, Zhuang ZJ, Yang WJ, Zang SF and Shi JP: Berberine
ameliorates non-alcoholic steatohepatitis in ApoE(−/-) mice. Exp
Ther Med. 14:4134–4140. 2017.PubMed/NCBI
|
|
21
|
Yang QH, Hu SP, Zhang YP, Xie WN, Li N, Ji
GY, Qiao NL, Lin XF, Chen TY and Liu HT: Effect of berberine on
expressions of uncoupling protein-2 mRNA and protein in hepatic
tissue of non-alcoholic fatty liver disease in rats. Chin J Integr
Med. 17:205–211. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu D, Wen W, Qi CL, Zhao RX, Lü JH, Zhong
CY and Chen YY: Ameliorative effect of berberine on renal damage in
rats with diabetes induced by high-fat diet and streptozotocin.
Phytomedicine. 19:712–718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gomes AP, Duarte FV, Nunes P, Hubbard BP,
Teodoro JS, Varela AT, Jones JG, Sinclair DA, Palmeira CM and Rolo
AP: Berberine protects against high fat diet-induced dysfunction in
muscle mitochondria by inducing SIRT1-dependent mitochondrial
biogenesis. Biochim Biophys Acta 1822. 185–195. 2012.
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jensen VS, Hvid H, Damgaard J, Nygaard H,
Ingvorsen C, Wulff EM, Lykkesfeldt J and Fledelius C: Dietary fat
stimulates development of NAFLD more potently than dietary fructose
in Sprague-Dawley rats. Diabetol Metab Syndr. 10:42018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Malaguarnera M, Di Rosa M, Nicoletti F and
Malaguarnera L: Molecular mechanisms involved in NAFLD progression.
J Mol Med (Berl). 87:679–695. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Day CP and James OF: Steatohepatitis: A
tale of two ‘hits’? Gastroenterology. 114:842–845. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu R, Wang Y, Zhang L and Guo Q:
Oxidative stress and liver disease. Hepatol Res. 42:741–749. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tanaka Y, Aleksunes LM, Yeager RL, Gyamfi
MA, Esterly N, Guo GL and Klaassen CD: NF-E2-related factor 2
inhibits lipid accumulation and oxidative stress in mice fed a
high-fat diet. J Pharmacol Exp Ther. 325:655–664. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xu W, Shao L, Zhou C, Wang H and Guo J:
Upregulation of Nrf2 Expression in Non-Alcoholic Fatty Liver and
Steatohepatitis. Hepatogastroenterology. 58:2077–2080. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Okada K, Warabi E, Sugimoto H, Horie M,
Gotoh N, Tokushige K, Hashimoto E, Utsunomiya H, Takahashi H, Ishii
T, et al: Deletion of Nrf2 leads to rapid progression of
steatohepatitis in mice fed atherogenic plus high-fat diet. J
Gastroenterol. 48:620–632. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Meakin PJ, Chowdhry S, Sharma RS, Ashford
FB, Walsh SV, McCrimmon RJ, Dinkova-Kostova AT, Dillon JF, Hayes JD
and Ashford ML: Susceptibility of Nrf2-Null mice to steatohepatitis
and cirrhosis upon consumption of a high-fat diet is associated
with oxidative stress, perturbation of the unfolded protein
response, and disturbance in the expression of metabolic enzymes
but not with insulin resistance. Mol Cell Biol. 34:3305–3320. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Z, Geng Y, Jiang J and Kong W:
Antioxidant and anti-inflammatory activities of berberine in the
treatment of diabetes mellitus. Evid-Based Compl Alt 2014.
2892642014.
|
|
34
|
Yin J, Xing H and Ye J: Efficacy of
berberine in patients with type 2 diabetes mellitus. Metabolism.
57:712–717. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chang X, Yan H, Fei J, Jiang M, Zhu H, Lu
D and Gao X: Berberine reduces methylation of the MTTP promoter and
alleviates fatty liver induced by a high-fat diet in rats. J Lipid
Res. 51:2504–2515. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou J, Zhou S, Tang J, Zhang K, Guang L,
Huang Y, Xu Y, Ying Y, Zhang L and Li D: Protective effect of
berberine on beta cells in streptozotocin- and
high-carbohydrate/high-fat diet-induced diabetic rats. Eur J
Pharmacol. 606:262–268. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hsu YY, Chen CS, Wu SN, Jong YJ and Lo YC:
Berberine activates Nrf2 nuclear translocation and protects against
oxidative damage via a phosphatidylinositol 3-kinase/Akt-dependent
mechanism in NSC34 motor neuron-like cells. Eur J Pharm Sci.
46:415–425. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yates MS, Tran QT, Dolan PM, Osburn WO,
Shin S, McCulloch CC, Silkworth JB, Taguchi K, Yamamoto M, Williams
CR, et al: Genetic versus chemoprotective activation of Nrf2
signaling: Overlapping yet distinct gene expression profiles
between Keap1 knockout and triterpenoid-treated mice.
Carcinogenesis. 30:1024–1031. 2009. View Article : Google Scholar : PubMed/NCBI
|