Lighter skin tone has long been associated with
youth and beauty among a variety of Asian cultures. Investment in
skin-whitening agents, boosted by markets in Asian countries,
especially those in China, India and Japan, is increasing annually
(1). Skin color is influenced by a
number of intrinsic factors, including skin types and genetic
background, and extrinsic factors, including the degree of sunlight
exposure and environmental pollution (2-4).
Skin color is determined by the quantity of melanosomes and their
extent of dispersion in the skin (5). Under physiological conditions,
pigmentation can protect the skin against harmful UV injury.
However, excessive generation of melanin can result in extensive
aesthetic problems, including melasma, pigmentation of ephelides
and post-inflammatory hyperpigmentation (1,6).
Traditional pharmacological agents, including corticosteroids,
hydroquinone and aminomercuric chloride, lighten skin tone through
the inhibition of either melanocyte maturation or interference with
the process of melanogenesis. However, most if not all of the
aforementioned agents are closely associated with adverse effects
including prickling sensation, contact dermatitis, irritation, high
toxicity and sensitivity (7-10).
Therefore, recent research by cosmetic companies and research
institutions has been focusing on the development of novel
whitening agents that selectively suppress the activity of
tyrosinase (TYR) to reduce hyperpigmentation whilst avoiding
cytotoxicity to normal, healthy melanocytes. As a result, natural
skin whitening compounds are currently garnering significant
attention in the cosmetic and medical industry (11,12).
The present review summarizes the biosynthetic
process of melanogenesis and the associated core regulatory
signaling pathways. It also reviews natural skin-whitening agents
in terms of their compound classification and discusses their
efficacy based on their mechanism of action on melanogenesis. In
addition, an overview of the current research methodology applied
for the evaluation of compound bioactivity is provided. The aim of
the present review is to provide informative guidance for the
development of safe and effective depigmenting agents for use in
the cosmetic industry.
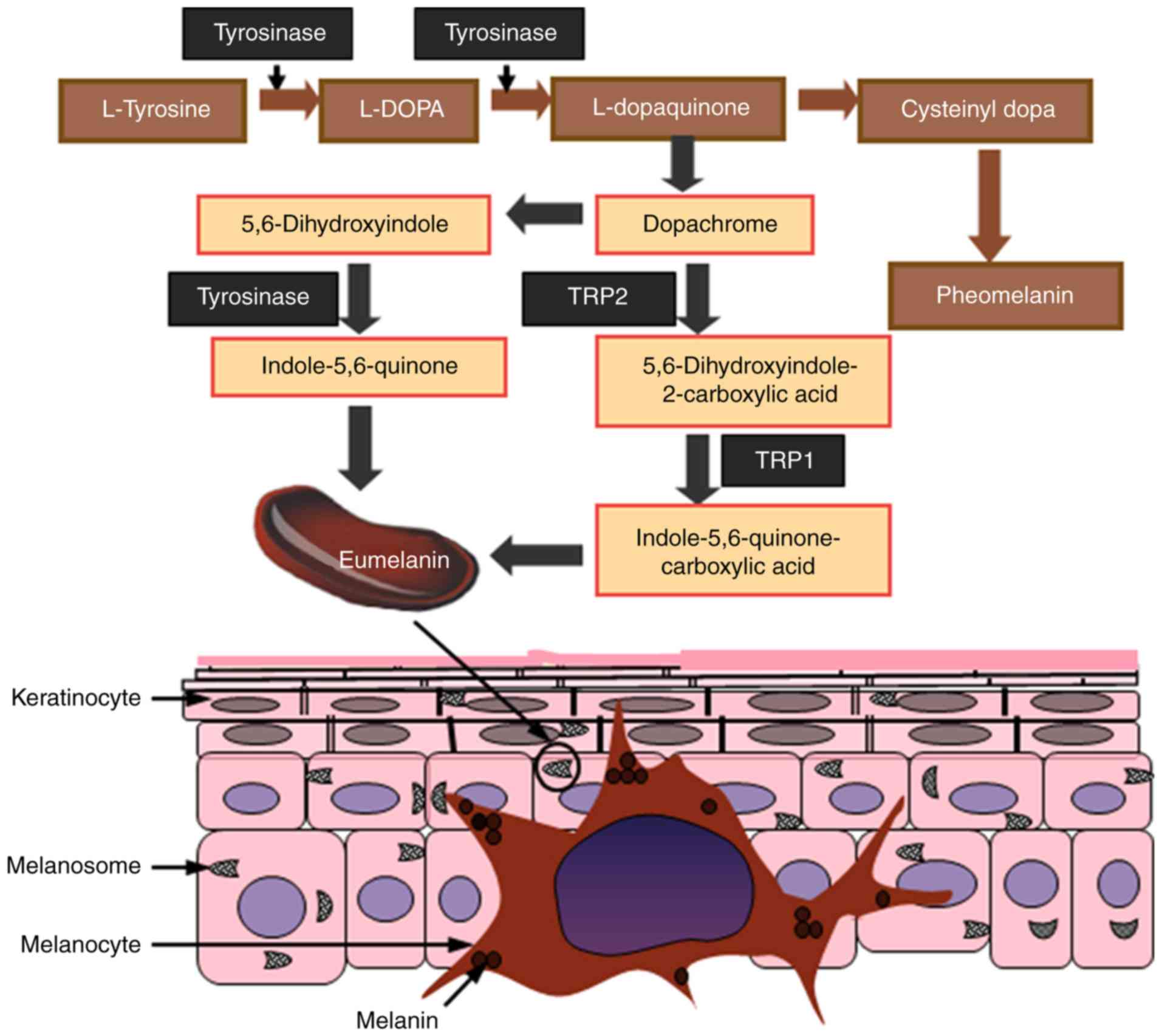
Melanin is mainly produced by melanocytes that are
localized in the epidermis, the outermost layer of the skin; it is
also this layer that determines skin color in humans (4). Melanin is primarily synthesized in
melanosomes, which function as specialized organelles in
melanocytes. Melanogenesis is a complex process that involves a
series of enzymatic and chemical reactions inside the melanosomes,
resulting in the production of two types of melanin: Eumelanin and
pheomelanin. Eumelanin is an insoluble polymer that is dark
brown-black in color, whereas pheomelanin is a soluble polymer
light red-yellow in color that also contain sulfur (13). Both eumelanin and pheomelanin are
formed by the conjugation of cysteine or glutathione (14-16).
To gain an understanding of the mechanism of whitening agents, a
summary of the signaling pathways associated with skin
melanogenesis is presented in Fig.
1. The pigmentation process starts with the oxidation of
L-tyrosine to L-dopaquinone (DQ) in the presence of the
rate-limiting enzyme TYR. Following DQ formation, the resulting
quinone undergoes intramolecular cyclization and oxidation, where
it serves as a substrate for the synthesis of eumelanin and
pheomelanin (17,18). During the process of melanogenesis,
hydroxylation of L-tyrosine to form L-3,4-dihydroxyphenylalanine
(L-DOPA) is the rate-limiting step of the whole process, which is
catalyzed by TYR.
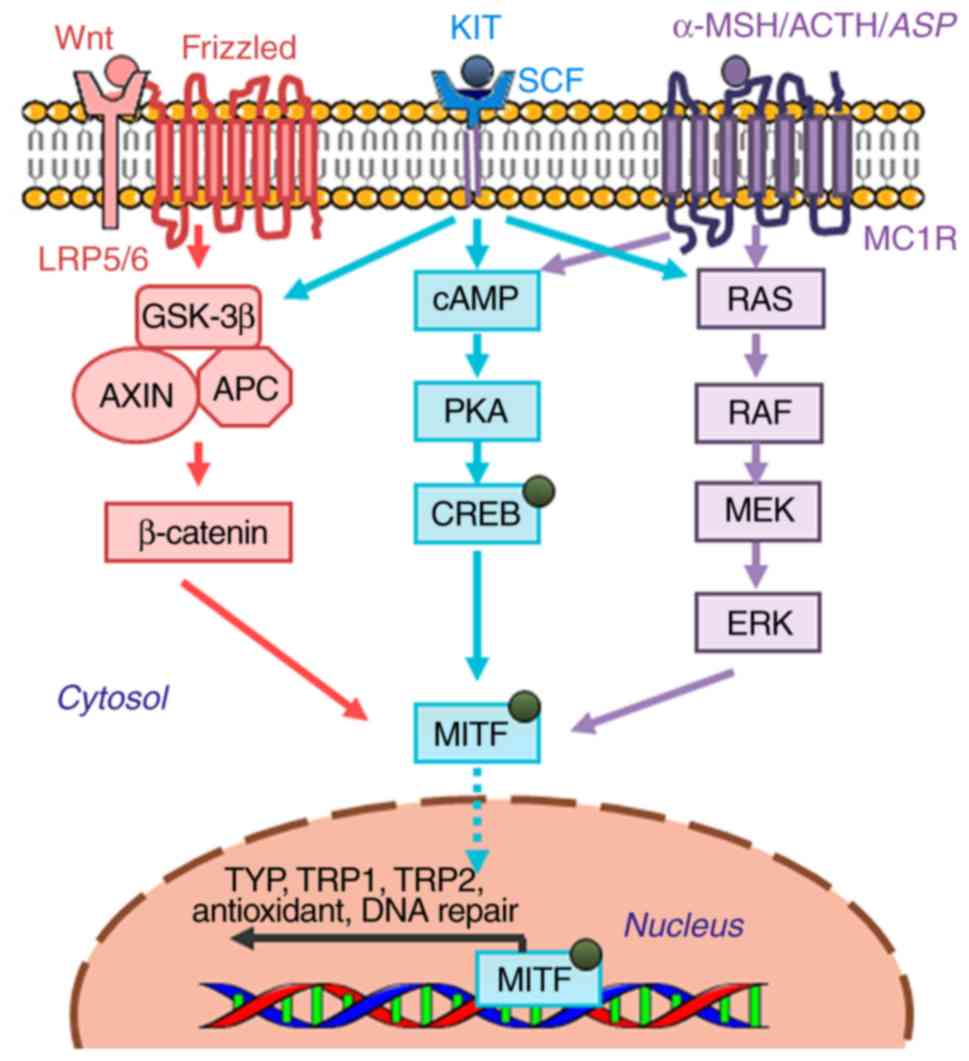
Melanogenesis is a complex process that is modulated
by a network of pivotal signaling cascades and transcription
factors, which is controlled at different levels. In particular,
modulation of TYR activity is the most commonly applied strategy
for the clinical intervention of pigmentation disorders. Since
naturally occurring inhibitors of melanogenesis usually garner more
attention compared with chemically synthesized compounds due to the
cosmetic demands of consumers, the present review focuses on
natural compounds that have been documented to exhibit
skin-whitening effects through the inhibition of TYR activity. The
three core signal pathways involved in the regulation of
melanogenesis are: i) melanocortin-1 receptor (MC1R) signaling; ii)
the Wnt/β-catenin signaling pathway; and iii) the tyrosine kinase
receptor KIT/stem cell factor (SCF) pathway, all of which converge
downstream to activate the master
regulator-microphthalmia-associated transcription factor (MITF)
(Fig. 2) (19). The following sections will describe
the genetic and molecular modulators that are involved in the
control of melanogenesis by these three key pathways.
α-MSH is a precursor polypeptide derived from
pro-opiomelanocortin that can modulate pigmentation through
paracrine action, whilst MC1R is a member of the G-protein-coupled
receptor family (20). α-MSH binding
to MC1R results in the activation of adenylyl cyclase, increasing
the intracellular levels of cAMP and subsequently upregulating TYR,
tyrosinase related protein-1 (TRP-1) and tyrosinase related
protein-2 (TRP-2) expression. The biological effects downstream of
cAMP elevation have been previously demonstrated to be
predominately mediated by cAMP-dependent protein kinase (PKA),
which phosphorylates cAMP-response element (CRE) binding protein
(CREB) (21). However, it has also
been suggested that neither TRP-1 nor TRP-2 have cAMP response
elements in their respective promoter regions. Evidence has
indicated that regulating the gene expression of TRP-1 and TRP-2 by
cAMP is directly associated with MITF, which binds to the M-box
sequence (AGTCATGTGCT) located in the tyrosinase distal elements
(TDEs) after its activation (22).
Since the promoter region of MITF contains the consensus CRE
sequence, the expression of MITF can also be increased by α-MSH
stimulation in a cAMP-dependent manner (23). This demonstrated that the α-MSH-MC1R
signaling pathway induces melanin production predominantly by
elevating intracellular cAMP levels, and the inhibition of which
can exert inhibitory effects on melanogenesis.
The Wnt signaling pathway has been previously
reported to serve an important role in melanogenesis (24,25). Wnt
ligands bind to Frizzled receptors on the cell surface, resulting
in the increased stability of cytoplasmic β-catenin, and its
subsequent translocation into the nucleus, where it activates the
transcription of MITF by interplay with lymphoid enhancer-binding
factor 1 (LEF1)/T-cell factor (LEF1/TCF) (26). Previous studies on melanocytes
suggest that β-catenin and LEF1 synergistically regulate the M
promoter activity of MITF via LEF1 binding sites, which upregulate
MITF expression in melanoma (27,28). By
regulating MITF transcription, the Wnt/β-catenin signaling pathway
can control the expression of TYR and other pigmentation
enzymes.
Recent studies have verified the important roles of
the SCF-KIT signaling pathway in melanocyte proliferation and
differentiation, and the process of melanogenesis (29,30). SCF
is a paracrine factor that is secreted by fibroblasts, whereas
c-KIT, its receptor, is expressed on melanocytes (31). When SCF binding to its
receptor-c-KIT, it stimulates tyrosine kinase activity, resulting
in receptor auto-phosphorylation to initiate signal transduction
(32,33). c-KIT phosphorylation directly
activates p38 mitogen-activated protein kinase (MAPK), a member of
the MAP kinase family, which in turn phosphorylates CREB and
subsequently activates MITF to promote TYR transcription (34). c-KIT can also activate ERK.
c-KIT-mediated ERK signaling pathway can induces CREB
phosphorylation to activate melanin synthesis on one hand, and on
the other hand, the activation of ERK signaling has been
demonstrated to phosphorylate MITF at the serine 73 residue, which
leads to the ubiquitination and degradation of MITF, this is the
feedback mechanism of the ERK pathway to regulate melanin
production (3,35). In addition to p38 MAPK and ERK, c-KIT
activation is associated with the phosphoinositide 3-kinase (PI3K)
signaling pathway, which not only regulates cell survival but also
causes pigmentation by activating the serine/threonine-specific
protein kinase AKT. Downstream, PI3K activation leads to the
phosphorylation of glycogen synthase kinase 3β (GSK-3β) to increase
MITF activity (36). Therefore,
inhibitors of the SCF-KIT signaling pathway can potentially exhibit
anti-melanogenesis activity.
MITF serves as the central hub of the regulatory
network of melanin synthesis that is comprised of numerous
transcription factors and signaling pathways that modulate the
survival, proliferation and differentiation of melanoblasts and
melanocytes (37). The MITF gene
contains multiple promoters, the M promoter is one of such
promoters, which is located adjacent to the common downstream exons
and is targeted by several transcriptional factors, including CREB,
paired box gene 3 (PAX3), LEF1/TCF, SRY-related HMG-box 10 (SOX10),
SOX9 and MITF itself (38). In
melanocytes, these transcription factors bind to the promoter of
MITF-M to regulate MITF expression whilst controlling the
transcription of several important genes. These genes are not only
related to the production of melanin, including TYR, TRP-1 and
TRP-2, but are also linked to the regulation of melanocyte
differentiation, proliferation and cell cycle progression.
Cyclin-dependent kinase 2 (CDK2), B-cell lymphoma-2 (BCL-2) and
Hypoxia-inducible factor 1-alpha (HIF-1α) are such genes that are
regulated by MITF. In addition, MAPK, ribosomal S6 kinase (RSK),
glycogen synthase kinase-3β (GSK-3β) and p38 can all phosphorylate
MITF and simultaneously modulate its transcriptional activity in
response to specific environmental cues (39-43).
Naturally occurring skin-whitening agents exert
their effects by regulating melanin production through a number of
mechanisms, including inhibiting the expression and activity of TYR
and suppressing the uptake and distribution of melanosomes. In the
cosmetics industry, since skin-whitening compounds from natural
sources are usually more appealing to consumers, a greater demand
exists for inhibitors of melanogenesis derived from herbal plants
that prevent hyperpigmentary disorders. Naturally occurring
bioactive compounds, including flavonoids, terpenoids,
polysaccharides and coumarin derivatives, all of which have been
previously demonstrated to exhibit antioxidant and
anti-inflammatory properties, are now becoming increasing
recognized to possess anti-melanogenesis functions (44,45).
Therefore, this section focuses on the natural active
skin-whitening agents that are currently known based on their
compound classification along with their mechanism of action on
melanogenesis.
MITF serves an indispensable role in melanogenesis
as it controls the transcription of TYR and other
pigmentation-associated enzymes (46-48).
Naturally occurring bioactive compounds have now been reported to
exert an anti-melanogenesis function by interfering with signaling
pathways to downregulate MITF expression. Among them, phenolic
compounds, including [6]-Shogaol (49), derived from Heracleum
moellendorffii Hance extracts (50), the ethyl acetate fraction of
Oroxylum indicum Vent. seeds (51) and
2-[4-(3-hydroxypropyl)-2-methoxyphenoxy]-1,3-propanediol (35) from Juglans mandshurica plants,
inhibit melanogenesis by mediating the degradation of MITF in a
manner that is associated with ERK signaling. By contrast, other
phenolic compounds (52-55)
exert anti-melanogenic properties by downregulating the cAMP/CREB
signaling pathway and/or activating related caspases to trigger
apoptosis of melanocyte cells (Table
I). Flavonoids, including isoorientin, catechin, coumaric acid
and kaempferol-7-O-D-glucuronide, derived from Gentiana
(56), Phyllostachys nigra
(57), Cryptotaenia japonica
(57) and dried pomegranate
concentrate powder (58) exhibit
skin-whitening effects by downregulating PKA/CREB-mediated MITF
expression. A list of other bioactive compounds, including
terpenoids, polysaccharides and lignanoids, and their respective
molecular mechanism of action on the melanogenesis pathway is
provided in Table I (49,59-72).
It can be observed that bioactive compounds are able to suppress
MITF or TYR activity by either binding to transcription factors
directly or by inhibiting melanogenic pathways upstream, including
that of cAMP/PKA, ERK, Wnt/β-catenin and MAPK. Therefore, these
aforementioned compounds represent promising skin-whitening agents,
but those targeting TYR gene expression are not recommended for
clinical use mainly for their non-specific effects through
intracellular signaling cascades (73).
TYR is a popular target for the development of
skin-whitening agents due to its position at the rate-limiting step
of the melanogenesis pathway. Additionally, TYR inhibitors have
highly specificity for targeting melanogenesis, reducing the risk
of side effects. Therefore, TYR inhibitors remain as the most
successful and commonly applied skin-whitening agents. The majority
of the naturally occurring compounds currently applied are
botanical inhibitors of TYR, where their mechanism of action mainly
entails two processes.
A number of studies have reported TYR inhibitors
from natural sources, most of which are originate from Asia.
Table II provides a summary of
studies that have previously applied such types of TYR inhibitors.
In a substantial number of these studies, mushroom TYR has been
used as the protein model, and the IC50 values of the
prospective TYR inhibitor were compared with those of other
established inhibitors, including kojic acid and arbutin. TYR is a
multi-functional type-3 copper-containing glycoprotein that is
located on the membrane of the melanosome (1,74).
Structurally, the active site of TYR consists two copper ions
surrounded by three histidine residues (75). Anthraquinones, flavonoids and
phenylpropanoids can serve as competitive inhibitors of TYR due
chemical structures similar to those of L-tyrosine or L-DOPA
(76,77).
Within the quinone family of compounds, the most
frequently applied skin-whitening agents are hydroquinone (HQ)
(78,79) and arbutin (80,81).
Although HQ can function as an alternative substrate for TYR, the
subsequent enzymatic reaction results in the production of reactive
oxygen species (ROS), which is thought to be responsible for its
skin-lightening properties, with possible associated side effects
including leukoderma and exogenous ochronosis (82,83).
Therefore, HQ has been banned in the EU, USA and a number of
African and Asian countries (5). By
contrast, arbutin is an effective agent for treating for
hyperpigmentation in the cosmetics industry, which is also commonly
applied as a positive control for melanogenesis studies.
Phenylpropanoids and olefinic unsaturated compounds,
which include ferulic acid, benzaldehyde (105), astaxanthin, curcumin and cinnamic
acid esters (106), have been
revealed to exert inhibitory effects on TYR. According to a study
by Park et al (107),
ferulic acid, one of the main phenolic components found in
Tetragonia tetragonioides, suppressed melanin synthesis by
reducing the expression of TYR and MITF in B16-F10 cells at
concentrations of between 5 and 20 µM. Additionally, Rao et
al (108), Niwano et al
(109) and Tu et al
(110) demonstrated that
astaxanthin and curcumin exhibit suppressive properties on melanin
synthesis and cellular TYR activity. Other typical agents with
reported inhibitory activities on TYR include kojic acid (111,112),
methyl gentisate (113,114), ganodermanondiol (71,115),
10-hydroxy-2-decenoic acid (116),
Stichopus japonicus extracts (69) and bis (4-hydroxybenzyl)sulphide
(117). Information on their
specific respective mechanisms of action are shown in Table II.
Substances that can regulate melanin synthesis by
affecting protein levels of the melanogenic enzymes without any
changes in mRNA levels likely regulate the activity of melanogenic
enzymes at post-translational levels. Post-translational
modification of components in this pathway primarily lead to the
inhibition of melanin synthesis. Currently, two main pathways are
known for the degradation of TYR, namely proteasomal and lysosomal
degradation (118,119). Unsaturated fatty acids, including
oleic acid (C18:1), linoleic acid (C18:2) and α-linolenic acid
(C18:3), have been demonstrated to accelerate the protein
degradation of TYR by activating one of these two pathways, leading
to anti-melanogenesis activity (120). These agents downregulate
intracellular TYR protein levels by promoting ubiquitin-dependent
degradation, inhibiting melanin synthesis and suppressing
hyperpigmentation. According to previous studies by Park et
al (121) and Lee et al
(122), terrein, a novel fungal
metabolite reduces TYR expression by downregulating MITF in a
manner that is dependent on ERK activation, with its inhibitory
effects on melanin synthesis prolonged by ubiquitin-mediated
proteasomal degradation. By contrast, lysosomes can also target TYR
for degradation. Geoditin A, an isomalabaricane triterpene compound
derived from the South China Sea Sponge Geodia japonica, has
been previously found to suppress melanogenesis by
post-translational regulation in the endoplasmic reticulum and the
degradation of TYR in the lysosome (123). Resveratrol, a promising
pigment-lightening flavonoid found in red wine, was recently found
to suppress TYR expression not via the inhibition of MITF
expression, but by directly inhibiting TYR activity by a
post-translational modification that reduces the levels of fully
mature TYR protein (119).
Retention of misfolded TYR proteins in the endoplasmic reticulum
results in the loss of pigmentation, which has also been proposed
to be one of the major post-translational mechanisms responsible
for the effects of resveratrol.
Following melanin synthesis, one key step of
melanogenesis in the skin is the translocation of mature
melanosomes into keratinocytes, which are then transported up to
the epidermidis where the melanin is dispersed. Therefore, agents
that can inhibit the transfer of melanosomes and/or accelerate
epidermal turnover can result in the whitening of the skin.
A number of studies have previously proposed
regulatory mechanisms of melanosome movement in dendrites and
interplay between keratinocytes and melanocytes during the transfer
process (124,125). In this regard, early skin-whitening
compounds, including niacinamide and soybean extracts, are reported
to interfere with this process. Niacinamide is has been shown to
reduce pigmentation by inhibiting melanosome transfer using a
skin-co-culture model (126),
whilst soymilk and soybean extracts have been previously suggested
to inhibit protease-activated receptor 2 activation in the skin,
which may enhance pigment transfer that results in skin whitening
(127,128). In addition, it was recently
reported that ginsenoside F1 exhibited skin lightening effects by
disrupting melanin transfer from the basal layer of melanocytes to
the upper layer of keratinocytes (129). Further microscopic research
revealed that melanosome transport between cells requires a number
of steps, including bidirectional long-range transfer to the apical
surface on microtubules, transfer to actin filaments, irreversible
short-range transfer by actin dynamics followed by binding to the
cell membrane (130). A number of
important molecules, including Rab27A, melanophilin (MLPH) /SLP
homolog lacking C2 domains-A, synaptotagmin-like protein (SLP)
2A/synaptotagmin 2 and myosin Va are involved in the regulation of
melanosome transport (130,131). Kudo et al (132) reported that O-methylated flavones
extracted from Scutellaria baicalensis Georgi, such as
wogonin, can inhibit the transport of intracellular melanosomes by
degrading melanophilin (MLPH), a carrier protein associated with
melanosome transport on actin filaments. Additionally, gagunin D, a
highly oxygenated diterpenoid from the marine sponge Phorbas
sp., was also found to exhibit anti-melanogenic properties by
downregulating the expression of proteins associated with
melanosome transfer, including Rab27A, MLPH and myosin Va (133). Therefore, these observations
suggest that downregulating the expression and activity of the
aforementioned proteins associated with melanosome transport may be
useful for reversing the process of skin hyperpigmentation.
A number of compounds have been documented to
possess the capacity to inhibit the dispersion of melanin granules
and accelerate skin turnover, which can result in a lighter skin
tone. Topical application of these compounds to the skin has been
demonstrated to effectively reduce the visibility of skin spots
without affecting their size or quantity, which can be used for
treating melasma. Examples of these compounds include α-hydroxy
acids, salicylic acid, linoleic acid and retinoic acids, which can
promote cellular renewal and facilitate the elimination of
melanized keratinocytes, leading to the loss of melanin
pigmentation (134,135). However, the application of those
acids is associated with side effects including erythema, scaling
and increased risk of sunburn (136-138). Therefore,
current research efforts are mainly focused on the discovery of
novel components of natural compounds with minimal off-target
effects. Liquiritin, a flavonoid glycoside of liquorice, has
previously been shown to significantly reduce hyperpigmentation in
20 women with a clinical diagnosis of melasma. The mechanism was
proposed to be associated with melanin dispersion mediated by the
pyran ring of the flavonoid chemical structure and the acceleration
of epidermal turnover (139). This
suggest flavonoids to be promising candidates for the development
of safe and effective interventions for hyperpigmentation.
Over the past few years, significant progress has
been made on the understanding of melanocyte biology and its
underlying mechanism, opening up new research avenues for the
discovery of novel melanogenesis inhibitors. In addition to the
direct suppression of TYR catalytic activity, other methods for the
disruption of melanogenesis include the post-transcriptional
control of TYR, regulation of melanosome transfer and the
suppression TYR transcription by suppressing upstream signaling
pathways. These mainly involve the inhibition of the master
regulator MITF, resulting from the reduction of intracellular cAMP
levels, increases in cytoplasmic β-catenin and/or the activation of
ERK signaling. Therefore, a large number of inhibitors acting
through these aforementioned alternative mechanisms have been
successfully identified (73).
Among these inhibitors, a mild, stable, safe and
effective compound is sought from natural extracts as a raw
material for the development of skin-whitening and skin care
products. For such prospective melanogenesis inhibitors, further
in vitro/in vivo studies and clinical trials are
required to evaluate efficacy and safety. To accelerate the process
of drug discovery, a variety of models and methodologies should be
applied to assess their potential hypopigmentation activity. From a
methodological perspective (140,141),
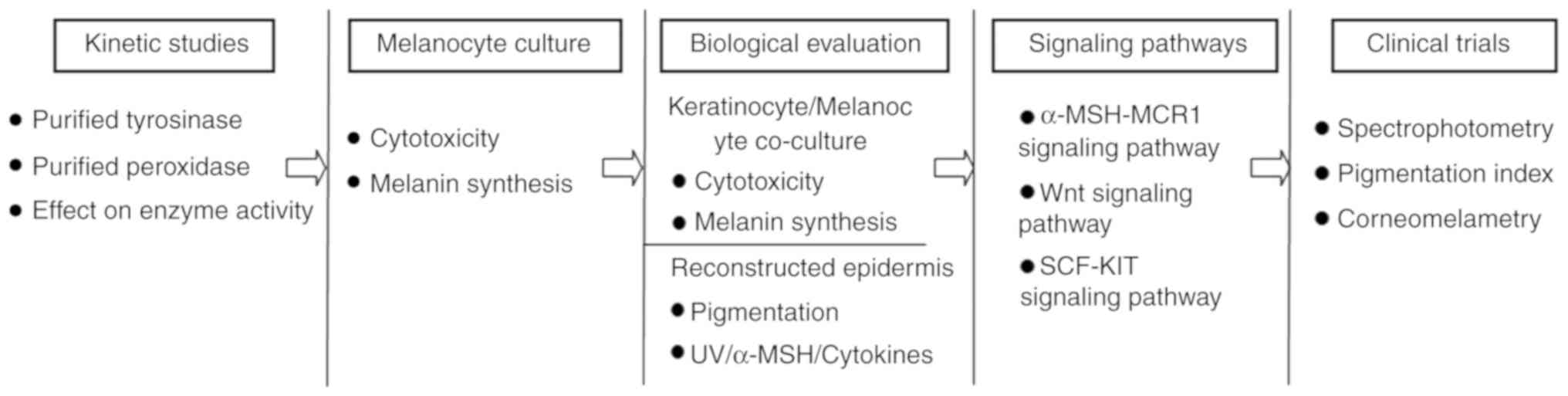
a multi-step process should be adopted for these investigations
(Fig. 3). Initial evaluation of
whitening properties in vitro should be conducted on
purified TYR and/or other melanogenic proteins, followed by the use
of melanocyte cultures to examine potential cytotoxic and melanin
synthetic effects. For further biological evaluation, co-culture
systems and reconstructed skin models should be adopted to screen
for the ability of the novel compounds to interfere with the
melanogenesis process, especially following stimuli including UV
irradiation, exposure to a-MSH or proinflammatory cytokines. In
addition, investigation into the regulatory mechanism involved in
melanogenesis should be performed. Finally, the in vivo
activity of the prospective agents should be evaluated using
non-invasive techniques such as UV light photography or
spectrophotometry to obtain comparable results (12,142).
It is expected that the aforementioned research methodology can
provide better opportunities for the development of novel
lightening agents that are effective and safe for use in the
clinical and cosmetic industries.
Although promising, the use of skin-lightening
compounds requires further research due to diverse modes of action
or off-target effects (77). Kojic
acid and arbutin remain the classic compounds that can be topically
used as skin-lightening agents in a clinical setting due to proven
efficacy. Additional natural skin-lightening compounds, including
mulberry, licorice and lemon extract, are regularly supplemented
into skin care products, to strengthen the effect of arbutin or
kojic acid (143,144). The ideal skin-lightening cosmetic
product should include a formulation that comprises compounds
acting on different pathways during the melanogenesis process. This
future combination should contain multiple targets and layers,
including the control of TYR expression on transcription and
protein levels, inhibition of enzymatic activity in the
melanogenesis pathway, suppression of melanocyte proliferation and
the transport of melanosomes on a cellular level. Although the
mechanisms of these inhibitors have been well characterized in
vitro, they have not been topically applied in cosmetics and
cosmeceuticals. Therefore, further evaluation of their
skin-whitening activity in vivo or in parallel human
clinical trials are required. In summary, from a clinical point of
view, additional mechanistic investigations of the novel natural
modulators on melanogenesis are urgently required.
Not applicable.
This project was supported by the National Natural
Science Foundation of China (grant no. 31671026), the Research
Project of Nanjing General Hospital (grant no. 2015056).
Not applicable.
GG and HS designed the theme of the review. WL, YC
and AT retrieved the relevant literature. WQ wrote and reviewed the
article. DZ helped to revise the manuscript and provided important
intellectual revision suggestions.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Pillaiyar T, Manickam M and Namasivayam V:
Skin whitening agents: Medicinal chemistry perspective of
tyrosinase inhibitors. J Enzyme Inhib Med Chem. 32:403–425.
2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Videira IF, Moura DF and Magina S:
Mechanisms regulating melanogenesis. An Bras Dermatol. 88:76–83.
2013.PubMed/NCBI View Article : Google Scholar
|
|
3
|
D'Mello SA, Finlay GJ, Baguley BC and
Askarian-Amiri ME: Signaling pathways in melanogenesis. Int J Mol
Sci. 17(pii: E1144)2016.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Gillbro JM and Olsson MJ: The
melanogenesis and mechanisms of skin-lightening agents--existing
and new approaches. Int J Cosmet Sci. 33:210–221. 2011.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Desmedt B, Courselle P, De Beer JO,
Rogiers V, Grosber M, Deconinck E and De Paepe K: Overview of skin
whitening agents with an insight into the illegal cosmetic market
in Europe. J Eur Acad Dermatol Venereol. 30:943–950.
2016.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Costin GE and Hearing VJ: Human skin
pigmentation: Melanocytes modulate skin color in response to
stress. Faseb J. 21:976–994. 2007.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Takizawa T, Imai T, Onose J, Ueda M,
Tamura T, Mitsumori K, Izumi K and Hirose M: Enhancement of
hepatocarcinogenesis by kojic acid in rat two-stage models after
initiation with N-bis (2-hydroxypropyl)nitrosamine or
N-diethylnitrosamine. Toxicol Sci. 81:43–49. 2004.PubMed/NCBI View Article : Google Scholar
|
|
8
|
García-Gavín J, González-Vilas D,
Fernández-Redondo V and Toribio J: Pigmented contact dermatitis due
to kojic acid. A paradoxical side effect of a skin lightener.
Contact Dermatitis. 62:63–64. 2010.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Chung KW, Jeong HO, Jang EJ, Choi YJ, Kim
DH, Kim SR, Lee KJ, Lee HJ, Chun P, Byun Y, et al: Characterization
of a small molecule inhibitor of melanogenesis that inhibits
tyrosinase activity and scavenges nitric oxide (NO). Biochim
Biophys Acta. 1830:4752–4761. 2013. View Article : Google Scholar
|
|
10
|
Hong YH, Jung EY, Noh DO and Suh HJ:
Physiological effects of formulation containing tannase-converted
green tea extract on skin care: Physical stability, collagenase,
elastase and tyrosinase activities. Integr Med Res. 3:25–33.
2014.PubMed/NCBI View Article : Google Scholar
|
|
11
|
Chiang HM, Chien YC, Wu CH, Kuo YH, Wu WC,
Pan YY, Su YH and Wen KC: Hydroalcoholic extract of Rhodiola rosea
L. (Crassulaceae) and its hydrolysate inhibit melanogenesis in
B16F0 cells by regulating the CREB/MITF/tyrosinase pathway. Food
Chem Toxicol. 65:129–139. 2014.PubMed/NCBI View Article : Google Scholar
|
|
12
|
Lajis AFB and Ariff AB: Discovery of new
depigmenting compounds and their efficacy to treat
hyperpigmentation: Evidence from in vitro study. J Cosmet Dermatol.
18:703–727. 2019.PubMed/NCBI View Article : Google Scholar
|
|
13
|
Ito S and Wakamatsu K: Quantitative
analysis of eumelanin and pheomelanin in humans, mice and other
animals: A comparative review. Pigment Cell Res. 16:523–531.
2003.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Slominski A, Tobin DJ, Shibahara S and
Wortsman J: Melanin pigmentation in mammalian skin and its hormonal
regulation. Physiol Rev. 84:1155–1228. 2004.PubMed/NCBI View Article : Google Scholar
|
|
15
|
Schiaffino MV: Signaling pathways in
melanosome biogenesis and pathology. Int J Biochem Cell Biol.
42:1094–1104. 2010.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Pillaiyar T, Manickam M and Jung SH:
Inhibitors of melanogenesis: A patent review (2009-2014). Expert
Opin Ther Pat. 25:775–788. 2015.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Hearing VJ and Jiménez M: Mammalian
tyrosinase-the critical regulatory control point in melanocyte
pigmentation. Int J Biochem. 19:1141–1147. 1987.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Halaban R, Patton RS, Cheng E, Svedine S,
Trombetta ES, Wahl ML, Ariyan S and Hebert DN: Abnormal
acidification of melanoma cells induces tyrosinase retention in the
early secretory pathway. J Biol Chem. 277:14821–14828.
2002.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Hou L, Panthier JJ and Arnheiter H:
Signaling and transcriptional regulation in the neural
crest-derived melanocyte lineage: Interactions between KIT and
MITF. Development. 127:5379–5389. 2000.PubMed/NCBI
|
|
20
|
Ryu S, Johnson A, Park Y, Kim B, Norris D,
Armstrong CA and Song PI: The alpha-melanocyte-stimulating hormone
suppresses TLR2-mediated functional responses through IRAK-M in
normal human keratinocytes. PLoS One. 10(e0136887)2015.PubMed/NCBI View Article : Google Scholar
|
|
21
|
Edelman AM, Blumenthal DK and Krebs EG:
Protein serine/threonine kinases. Annu Rev Biochem. 56:567–613.
1987.PubMed/NCBI View Article : Google Scholar
|
|
22
|
Yasumoto K, Yokoyama K, Shibata K, Tomita
Y and Shibahara S: Microphthalmia-associated transcription factor
as a regulator for melanocyte-specific transcription of the human
tyrosinase gene. Mol Cell Biol. 14:8058–8070. 1994.PubMed/NCBI View Article : Google Scholar
|
|
23
|
Bertolotto C, Abbe P, Hemesath TJ, Bille
K, Fisher DE, Ortonne JP and Ballotti R: Microphthalmia gene
product as a signal transducer in cAMP-induced differentiation of
melanocytes. J Cell Biol. 142:827–835. 1998.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Zhu PY, Yin WH, Wang MR, Dang YY and Ye
XY: Andrographolide suppresses melanin synthesis through
Akt/GSK3β/β-catenin signal pathway. J Dermatol Sci. 79:74–83.
2015.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Hwang I, Park JH, Park HS, Choi KA, Seol
KC, Oh SI, Kang S and Hong S: Neural stem cells inhibit melanin
production by activation of Wnt inhibitors. J Dermatol Sci.
72:274–283. 2013.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Steingrimsson E, Copeland NG and Jenkins
NA: Melanocytes and the microphthalmia transcription factor
network. Annu Rev Genet. 38:365–411. 2004.PubMed/NCBI View Article : Google Scholar
|
|
27
|
Takeda K, Yasumoto K, Takada R, Takada S,
Watanabe K, Udono T, Saito H, Takahashi K and Shibahara S:
Induction of melanocyte-specific microphthalmia-associated
transcription factor by Wnt-3a. J Biol Chem. 275:14013–14016.
2000.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Widlund HR, Horstmann MA, Price ER, Cui J,
Lessnick SL, Wu M, He X and Fisher DE: Beta-catenin-induced
melanoma growth requires the downstream target
Microphthalmia-associated transcription factor. J Cell Biol.
158:1079–1087. 2002.PubMed/NCBI View Article : Google Scholar
|
|
29
|
Martinez-Anton A, Gras D, Bourdin A,
Dubreuil P and Chanez P: KIT as a therapeutic target for
non-oncological diseases. Pharmacol Ther. 197:11–37.
2019.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Niwano T, Terazawa S, Nakajima H and
Imokawa G: The stem cell factor-stimulated melanogenesis in human
melanocytes can be abrogated by interrupting the phosphorylation of
MSK1: Evidence for involvement of the p38/MSK1/CREB/MITF axis. Arch
Dermatol Res. 310:187–196. 2018.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Li PH, Liu LH, Chang CC, Gao R, Leung CH,
Ma DL and David Wang HM: Silencing stem cell factor gene in
fibroblasts to regulate paracrine factor productions and enhance
c-Kit expression in melanocytes on melanogenesis. Int J Mol Sci.
19(pii: E1475)2018.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Flaherty KT, Hodi FS and Fisher DE: From
genes to drugs: Targeted strategies for melanoma. Nat Rev Cancer.
12:349–361. 2012.PubMed/NCBI View Article : Google Scholar
|
|
33
|
Bonaventure J, Domingues MJ and Larue L:
Cellular and molecular mechanisms controlling the migration of
melanocytes and melanoma cells. Pigment Cell Melanoma Res.
26:316–325. 2013.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Ahn JH, Jin SH and Kang HY: LPS induces
melanogenesis through p38 MAPK activation in human melanocytes.
Arch Dermatol Res. 300:325–329. 2008.PubMed/NCBI View Article : Google Scholar
|
|
35
|
Kim JY, Lee EJ, Ahn Y, Park S, Kim SH and
Oh SH: A chemical compound from fruit extract of Juglans
mandshurica inhibits melanogenesis through p-ERK-associated MITF
degradation. Phytomedicine. 57:57–64. 2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Hwang E, Lee TH, Lee WJ, Shim WS, Yeo EJ,
Kim S and Kim SY: A novel synthetic Piper amide derivative NED-180
inhibits hyperpigmentation by activating the PI3K and ERK pathways
and by regulating Ca2+ influx via TRPM1 channels. Pigment Cell
Melanoma Res. 29:81–91. 2016.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Vance KW and Goding CR: The transcription
network regulating melanocyte development and melanoma. Pigment
Cell Res. 17:318–325. 2004.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Seberg HE, Van Otterloo E and Cornell RA:
Beyond MITF: Multiple transcription factors directly regulate the
cellular phenotype in melanocytes and melanoma. Pigment Cell
Melanoma Res. 30:454–466. 2017.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Price ER, Horstmann MA, Wells AG,
Weilbaecher KN, Takemoto CM, Landis MW and Fisher DE:
alpha-Melanocyte-stimulating hormone signaling regulates expression
of microphthalmia, a gene deficient in Waardenburg syndrome. J Biol
Chem. 273:33042–33047. 1998.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Bondurand N, Pingault V, Goerich DE,
Lemort N, Sock E, Le Caignec C, Wegner M and Goossens M:
Interaction among SOX10, PAX3 and MITF, three genes altered in
Waardenburg syndrome. Hum Mol Genet. 9:1907–1917. 2000.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Jacquemin P, Lannoy VJ, O'Sullivan J, Read
A, Lemaigre FP and Rousseau GG: The transcription factor onecut-2
controls the microphthalmia-associated transcription factor gene.
Biochem Biophys Res Commun. 285:1200–1205. 2001.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Saito H, Yasumoto K, Takeda K, Takahashi
K, Fukuzaki A, Orikasa S and Shibahara S: Melanocyte-specific
microphthalmia-associated transcription factor isoform activates
its own gene promoter through physical interaction with
lymphoid-enhancing factor 1. J Biol Chem. 277:28787–28794.
2002.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Hsiao JJ and Fisher DE: The roles of
microphthalmia-associated transcription factor and pigmentation in
melanoma. Arch Biochem Biophys. 563:28–34. 2014.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Hasegawa T, Takano F, Takata T, Niiyama M
and Ohta T: Bioactive monoterpene glycosides conjugated with gallic
acid from the leaves of Eucalyptus globulus. Phytochemistry.
69:747–753. 2008.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Choi MH, Jo HG, Yang JH, Ki SH and Shin
HJ: Antioxidative and anti-melanogenic activities of bamboo stems
(Phyllostachys nigra variety henosis) via PKA/CREB-mediated MITF
downregulation in B16F10 melanoma cells. Int J Mol Sci. 19(pii:
E409)2018.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Yasumoto K, Yokoyama K, Takahashi K,
Tomita Y and Shibahara S: Functional analysis of
microphthalmia-associated transcription factor in pigment
cell-specific transcription of the human tyrosinase family genes. J
Biol Chem. 272:503–509. 1997.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Tachibana M: MITF: A stream flowing for
pigment cells. Pigment Cell Res. 13:230–240. 2000.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Fang D, Tsuji Y and Setaluri V: Selective
down-regulation of tyrosinase family gene TYRP1 by inhibition of
the activity of melanocyte transcription factor, MITF. Nucleic
Acids Res. 30:3096–3106. 2002.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Huang HC, Chang SJ, Wu CY, Ke HJ and Chang
TM: [6]-Shogaol inhibits α-MSH-induced melanogenesis through the
acceleration of ERK and PI3K/Akt-mediated MITF degradation. Biomed
Res Int. 2014(842569)2014.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Alam MB, Seo BJ, Zhao P and Lee SH:
Anti-melanogenic activities of heracleum moellendorffii via
ERK1/2-mediated MITF downregulation. Int J Mol Sci. 17(pii:
E1844)2016.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Zhao P, Alam MB, An H, Choi HJ, Cha YH,
Yoo CY, Kim HH and Lee SH: Antimelanogenic effect of an oroxylum
indicum seed extract by suppression of MITF expression through
activation of MAPK signaling protein. Int J Mol Sci. 19(pii:
E760)2018.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Wu PY, You YJ, Liu YJ, Hou CW, Wu CS, Wen
KC, Lin CY and Chiang HM: Sesamol inhibited melanogenesis by
regulating melanin-related signal transduction in B16F10 cells. Int
J Mol Sci. 19(pii: E1108)2018.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Truong XT, Park SH, Lee YG, Jeong HY, Moon
JH and Jeon TI: Protocatechuic acid from pear inhibits
melanogenesis in melanoma cells. Int J Mol Sci. 18(pii:
E1809)2017.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Sun L, Guo Y, Zhang Y and Zhuang Y:
Antioxidant and Anti-tyrosinase activities of phenolic extracts
from rape bee pollen and inhibitory melanogenesis by cAMP/MITF/TYR
pathway in B16 mouse melanoma cells. Front Pharmacol.
8(104)2017.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Chen YS, Lee SM, Lin CC and Liu CY:
Hispolon decreases melanin production and induces apoptosis in
melanoma cells through the downregulation of tyrosinase and
microphthalmia-associated transcription factor (MITF) expressions
and the activation of caspase-3, -8 and -9. Int J Mol Sci.
15:1201–1215. 2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Wu QY, Wong ZC, Wang C, Fung AH, Wong EO,
Chan GK, Dong TT, Chen Y and Tsim KW: Isoorientin derived from
Gentiana veitchiorum Hemsl. flowers inhibits melanogenesis by
down-regulating MITF-induced tyrosinase expression. Phytomedicine.
57:129–136. 2019. View Article : Google Scholar
|
|
57
|
Seong ZK, Lee SY, Poudel A, Oh SR and Lee
HK: Constituents of cryptotaenia japonica inhibit melanogenesis via
CREB- and MAPK-associated signaling pathways in murine B16 melanoma
cells. Molecules. 21(pii: E1296)2016.PubMed/NCBI View Article : Google Scholar
|
|
58
|
Kang SJ, Choi BR, Lee EK, Kim SH, Yi HY,
Park HR, Song CH, Lee YJ and Ku SK: Inhibitory effect of dried
pomegranate concentration powder on melanogenesis in B16F10
melanoma cells; involvement of p38 and PKA signaling pathways. Int
J Mol Sci. 16:24219–24242. 2015.PubMed/NCBI View Article : Google Scholar
|
|
59
|
Lee HJ, Lee WJ, Chang SE and Lee GY:
Hesperidin, A popular antioxidant inhibits melanogenesis via Erk1/2
Mediated MITF degradation. Int J Mol Sci. 16:18384–18395.
2015.PubMed/NCBI View Article : Google Scholar
|
|
60
|
Chae JK, Subedi L, Jeong M, Park YU, Kim
CY, Kim H and Kim SY: Gomisin N inhibits melanogenesis through
regulating the PI3K/Akt and MAPK/ERK signaling pathways in
melanocytes. Int J Mol Sci. 18(pii: E471)2017.PubMed/NCBI View Article : Google Scholar
|
|
61
|
Su TR, Lin JJ, Tsai CC, Huang TK, Yang ZY,
Wu MO, Zheng YQ, Su CC and Wu YJ: Inhibition of melanogenesis by
gallic acid: Possible involvement of the PI3K/Akt, MEK/ERK and
Wnt/β-catenin signaling pathways in B16F10 cells. Int J Mol Sci.
14:20443–20458. 2013.PubMed/NCBI View Article : Google Scholar
|
|
62
|
Lee DH, Ahn SS, Kim JB, Lim Y, Lee YH and
Shin SY: Downregulation of α-melanocyte-stimulating hormone-induced
activation of the Pax3-MITF-tyrosinase axis by sorghum ethanolic
extract in B16F10 melanoma cells. Int J Mol Sci. 19(pii:
E1640)2018.PubMed/NCBI View Article : Google Scholar
|
|
63
|
Tsao YT, Huang YF, Kuo CY, Lin YC, Chiang
WC, Wang WK, Hsu CW and Lee CH: Hinokitiol inhibits melanogenesis
via AKT/mTOR signaling in B16F10 mouse melanoma cells. Int J Mol
Sci. 17(248)2016.PubMed/NCBI View Article : Google Scholar
|
|
64
|
Ko GA, Shrestha S and Kim Cho S: Sageretia
thea fruit extracts rich in methyl linoleate and methyl linolenate
downregulate melanogenesis via the Akt/GSK3β signaling pathway.
Nutr Res Pract. 12:3–12. 2018.PubMed/NCBI View Article : Google Scholar
|
|
65
|
Lee SJ, Lee WJ, Chang SE and Lee GY:
Antimelanogenic effect of ginsenoside Rg3 through extracellular
signal-regulated kinase-mediated inhibition of
microphthalmia-associated transcription factor. J Ginseng Res.
39:238–242. 2015.PubMed/NCBI View Article : Google Scholar
|
|
66
|
Cho BR, Jun HJ, Thach TT, Wu C and Lee SJ:
Betaine reduces cellular melanin content via suppression of
microphthalmia-associated transcription factor in B16-F1 murine
melanocytes. Food Sci Biotechnol. 26:1391–1397. 2017.PubMed/NCBI View Article : Google Scholar
|
|
67
|
Alam MB, Bajpai VK, Lee J, Zhao P, Byeon
JH, Ra JS, Majumder R, Lee JS, Yoon JI, Rather IA, et al:
Inhibition of melanogenesis by jineol from Scolopendra subspinipes
mutilans via MAP-Kinase mediated MITF downregulation and the
proteasomal degradation of tyrosinase. Sci Rep. 7(45858)2017.
View Article : Google Scholar
|
|
68
|
Hu S, Huang J, Pei S, Ouyang Y, Ding Y,
Jiang L, Lu J, Kang L, Huang L, Xiang H, et al: Ganoderma lucidum
polysaccharide inhibits UVB-induced melanogenesis by antagonizing
cAMP/PKA and ROS/MAPK signaling pathways. J Cell Physiol.
234:7330–7340. 2019.PubMed/NCBI View Article : Google Scholar
|
|
69
|
Oh CT, Kwon TR, Jang YJ, Yoo KH, Kim BJ
and Kim H: Inhibitory effects of Stichopus japonicus extract on
melanogenesis of mouse cells via ERK phosphorylation. Mol Med Rep.
16:1079–1086. 2017.PubMed/NCBI View Article : Google Scholar
|
|
70
|
Huang HC, Wei CM, Siao JH, Tsai TC, Ko WP,
Chang KJ, Hii CH and Chang TM: Supercritical fluid extract of spent
coffee grounds attenuates melanogenesis through downregulation of
the PKA, PI3K/Akt and MAPK signaling pathways. Evid Based
Complement Alternat Med. 2016(5860296)2016. View Article : Google Scholar
|
|
71
|
Kim JW, Kim HI, Kim JH, Kwon OC, Son ES,
Lee CS and Park YJ: Effects of ganodermanondiol, a new
melanogenesis inhibitor from the medicinal mushroom ganoderma
lucidum. Int J Mol Sci. 17(pii: E1798)2016.PubMed/NCBI View Article : Google Scholar
|
|
72
|
Oh TI, Jung HJ, Lee YM, Lee S, Kim GH, Kan
SY, Kang H, Oh T, Ko HM, Kwak KC, et al: Zerumbone, a tropical
ginger sesquiterpene of zingiber officinale roscoe, attenuates
α-MSH-induced melanogenesis in B16F10 cells. Int J Mol Sci. 19(pii:
E3149)2018.PubMed/NCBI View Article : Google Scholar
|
|
73
|
Chang TS: Natural melanogenesis inhibitors
acting through the down-regulation of tyrosinase activity.
Materials (Basel). 5:1661–1685. 2012. View Article : Google Scholar
|
|
74
|
Sánchez-Ferrer A, Rodríguez-López JN,
García-Cánovas F and García-Carmona F: Tyrosinase: A comprehensive
review of its mechanism. Biochim Biophys Acta. 1247:1–11.
1995.PubMed/NCBI View Article : Google Scholar
|
|
75
|
Matoba Y, Kumagai T, Yamamoto A, Yoshitsu
H and Sugiyama M: Crystallographic evidence that the dinuclear
copper center of tyrosinase is flexible during catalysis. J Biol
Chem. 281:8981–8990. 2006.PubMed/NCBI View Article : Google Scholar
|
|
76
|
Menter JM, Etemadi AA, Chapman W, Hollins
TD and Willis I: In vivo depigmentation by hydroxybenzene
derivatives. Melanoma Res. 3:443–449. 1993.PubMed/NCBI View Article : Google Scholar
|
|
77
|
Briganti S, Camera E and Picardo M:
Chemical and instrumental approaches to treat hyperpigmentation.
Pigment Cell Res. 16:101–110. 2003.PubMed/NCBI View Article : Google Scholar
|
|
78
|
Kang WH, Chun SC and Lee S: Intermittent
therapy for melasma in Asian patients with combined topical agents
(retinoic acid, hydroquinone and hydrocortisone): Clinical and
histological studies. J Dermatol. 25:587–596. 1998.PubMed/NCBI View Article : Google Scholar
|
|
79
|
Guevara IL and Pandya AG: Melasma treated
with hydroquinone, tretinoin and a fluorinated steroid. Int J
Dermatol. 40:212–215. 2001.PubMed/NCBI View Article : Google Scholar
|
|
80
|
Badreshia-Bansal S and Draelos ZD: Insight
into skin lightening cosmeceuticals for women of color. J Drugs
Dermatol. 6:32–39. 2007.PubMed/NCBI
|
|
81
|
Parvez S, Kang M, Chung HS, Cho C, Hong
MC, Shin MK and Bae H: Survey and mechanism of skin depigmenting
and lightening agents. Phytother Res. 20:921–934. 2006.PubMed/NCBI View Article : Google Scholar
|
|
82
|
Haddad AL, Matos LF, Brunstein F, Ferreira
LM and Silva A and Costa D Jr: A clinical, prospective, randomized,
double-blind trial comparing skin whitening complex with
hydroquinone vs. placebo in the treatment of melasma. Int J
Dermatol. 42:153–156. 2003.PubMed/NCBI View Article : Google Scholar
|
|
83
|
Gupta AK, Gover MD, Nouri K and Taylor S:
The treatment of melasma: A review of clinical trials. J Am Acad
Dermatol. 55:1048–1065. 2006.PubMed/NCBI View Article : Google Scholar
|
|
84
|
Kim DS, Park SH, Kwon SB, Li K, Youn SW
and Park KC: (-)-Epigallocatechin-3-gallate and hinokitiol reduce
melanin synthesis via decreased MITF production. Arch Pharm Res.
27:334–339. 2004.PubMed/NCBI View Article : Google Scholar
|
|
85
|
Fan M, Zhang G, Hu X, Xu X and Gong D:
Quercetin as a tyrosinase inhibitor: Inhibitory activity,
conformational change and mechanism. Food Res Int. 100:226–233.
2017.PubMed/NCBI View Article : Google Scholar
|
|
86
|
Jones K, Hughes J, Hong M, Jia Q and
Orndorff S: Modulation of melanogenesis by aloesin: A competitive
inhibitor of tyrosinase. Pigment Cell Res. 15:335–340.
2002.PubMed/NCBI View Article : Google Scholar
|
|
87
|
Jin YH, Lee SJ, Chung MH, Park JH, Park
YI, Cho TH and Lee SK: Aloesin and arbutin inhibit tyrosinase
activity in a synergistic manner via a different action mechanism.
Arch Pharm Res. 22:232–236. 1999.PubMed/NCBI View Article : Google Scholar
|
|
88
|
Solano F, Briganti S, Picardo M and Ghanem
G: Hypopigmenting agents: An updated review on biological, chemical
and clinical aspects. Pigment Cell Res. 19:550–571. 2006.PubMed/NCBI View Article : Google Scholar
|
|
89
|
Satooka H and Kubo I: Resveratrol as a
kcat type inhibitor for tyrosinase: Potentiated melanogenesis
inhibitor. Bioorg Med Chem. 20:1090–1099. 2012.PubMed/NCBI View Article : Google Scholar
|
|
90
|
Lee TH, Seo JO, Baek SH and Kim SY:
Inhibitory effects of resveratrol on melanin synthesis in
ultraviolet B-induced pigmentation in Guinea pig skin. Biomol Ther
(Seoul). 22:35–40. 2014.PubMed/NCBI View Article : Google Scholar
|
|
91
|
Chen J, Yu X and Huang Y: Inhibitory
mechanisms of glabridin on tyrosinase. Spectrochim Acta A Mol
Biomol Spectrosc. 168:111–117. 2016.PubMed/NCBI View Article : Google Scholar
|
|
92
|
Lin Y, Kuang Y, Li K, Wang S, Song W, Qiao
X, Sabir G and Ye M: Screening for bioactive natural products from
a 67-compound library of Glycyrrhiza inflata. Bioorg Med Chem.
25:3706–3713. 2017.PubMed/NCBI View Article : Google Scholar
|
|
93
|
Fu B, Li H, Wang X, Lee FS and Cui S:
Isolation and identification of flavonoids in licorice and a study
of their inhibitory effects on tyrosinase. J Agric Food Chem.
53:7408–7414. 2005.PubMed/NCBI View Article : Google Scholar
|
|
94
|
Nerya O, Vaya J, Musa R, Izrael S,
Ben-Arie R and Tamir S: Glabrene and isoliquiritigenin as
tyrosinase inhibitors from licorice roots. J Agric Food Chem.
51:1201–1207. 2003.PubMed/NCBI View Article : Google Scholar
|
|
95
|
Yang SH, Tsatsakis AM, Tzanakakis G, Kim
HS, Le B, Sifaki M, Spandidos DA, Tsukamoto C, Golokhvast KS,
Izotov BN, et al: Soyasaponin Ag inhibits αMSHinduced melanogenesis
in B16F10 melanoma cells via the downregulation of TRP2. Int J Mol
Med. 40:631–636. 2017.PubMed/NCBI View Article : Google Scholar
|
|
96
|
Wang Y, Curtis-Long MJ, Lee BW, Yuk HJ,
Kim DW and Tan XF and Park KH: Inhibition of tyrosinase activity by
polyphenol compounds from Flemingia philippinensis roots. Bioorg
Med Chem. 22:1115–1120. 2014.PubMed/NCBI View Article : Google Scholar
|
|
97
|
Takahashi M, Takara K, Toyozato T and Wada
K: A novel bioactive chalcone of Morus australis inhibits
tyrosinase activity and melanin biosynthesis in B16 melanoma cells.
J Oleo Sci. 61:585–592. 2012.PubMed/NCBI View Article : Google Scholar
|
|
98
|
Roh JS, Han JY, Kim JH and Hwang JK:
Inhibitory effects of active compounds isolated from safflower
(Carthamus tinctorius L.) seeds for melanogenesis. Biol Pharm Bull.
27:1976–1978. 2004.PubMed/NCBI View Article : Google Scholar
|
|
99
|
Liang CP, Chang CH, Liang CC, Hung KY and
Hsieh CW: In vitro antioxidant activities, free radical scavenging
capacity and tyrosinase inhibitory of flavonoid compounds and
ferulic acid from Spiranthes sinensis (Pers.) Ames. Molecules.
19:4681–4694. 2014.PubMed/NCBI View Article : Google Scholar
|
|
100
|
Jhan JK, Chung YC, Chen GH, Chang CH, Lu
YC and Hsu CK: Anthocyanin contents in the seed coat of black soya
bean and their anti-human tyrosinase activity and antioxidative
activity. Int J Cosmet Sci. 38:319–324. 2016.PubMed/NCBI View Article : Google Scholar
|
|
101
|
Jeong HS, Gu GE, Jo AR, Bang JS, Yun HY,
Baek KJ, Kwon NS, Park KC and Kim DS: Baicalin-induced Akt
activation decreases melanogenesis through downregulation of
microphthalmia-associated transcription factor and tyrosinase. Eur
J Pharmacol. 761:19–27. 2015.PubMed/NCBI View Article : Google Scholar
|
|
102
|
Hwang JA, Park NH, Na YJ, Lee HK, Lee JH,
Kim YJ and Lee CS: Coumestrol down-regulates melanin production in
melan-a murine melanocytes through degradation of tyrosinase. Biol
Pharm Bull. 40:535–539. 2017.PubMed/NCBI View Article : Google Scholar
|
|
103
|
de Freitas MM, Fontes PR, Souza PM,
William Fagg C, Neves Silva Guerra E, de Medeiros Nóbrega YK,
Silveira D, Fonseca-Bazzo Y, Simeoni LA, Homem-de-Mello M, et al:
Extracts of Morus nigra L. leaves standardized in chlorogenic acid,
rutin and isoquercitrin: Tyrosinase inhibition and cytotoxicity.
PLoS One. 11(e0163130)2016.PubMed/NCBI View Article : Google Scholar
|
|
104
|
Chen YS, Lee SM, Lin CC, Liu CY, Wu MC and
Shi WL: Kinetic study on the tyrosinase and melanin formation
inhibitory activities of carthamus yellow isolated from Carthamus
tinctorius L. J Biosci Bioeng. 115:242–245. 2013.PubMed/NCBI View Article : Google Scholar
|
|
105
|
Nihei KI and Kubo I: Substituent effect of
benzaldehydes on tyrosinase inhibition. Plant Physiol Biochem.
112:278–282. 2017.PubMed/NCBI View Article : Google Scholar
|
|
106
|
Moghrovyan A, Sahakyan N, Babayan A,
Chichoyan N, Petrosyan M and Trchounian A: Essential oil and
ethanol extract of oregano (Origanum vulgare L.) from Armenian
flora as a natural source of terpenes, flavonoids and other
phytochemicals with antiradical, antioxidant, metal chelating,
tyrosinase inhibitory and antibacterial activity. Curr Pharm Des.
25:1809–1816. 2019.PubMed/NCBI View Article : Google Scholar
|
|
107
|
Park HJ, Cho JH, Hong SH, Kim DH, Jung HY,
Kang IK and Cho YJ: Whitening and anti-wrinkle activities of
ferulic acid isolated from Tetragonia tetragonioides in B16F10
melanoma and CCD-986sk fibroblast cells. J Nat Med. 72:127–135.
2018.PubMed/NCBI View Article : Google Scholar
|
|
108
|
Rao AR, Sindhuja HN, Dharmesh SM, Sankar
KU, Sarada R and Ravishankar GA: Effective inhibition of skin
cancer, tyrosinase and antioxidative properties by astaxanthin and
astaxanthin esters from the green alga Haematococcus pluvialis. J
Agric Food Chem. 61:3842–3851. 2013.PubMed/NCBI View Article : Google Scholar
|
|
109
|
Niwano T, Terazawa S, Nakajima H,
Wakabayashi Y and Imokawa G: Astaxanthin and withaferin A block
paracrine cytokine interactions between UVB-exposed human
keratinocytes and human melanocytes via the attenuation of
endothelin-1 secretion and its downstream intracellular signaling.
Cytokine. 73:184–197. 2015.PubMed/NCBI View Article : Google Scholar
|
|
110
|
Tu CX, Lin M, Lu SS, Qi XY, Zhang RX and
Zhang YY: Curcumin inhibits melanogenesis in human melanocytes.
Phytother Res. 26:174–179. 2012.PubMed/NCBI View Article : Google Scholar
|
|
111
|
Cabanes J, Chazarra S and Garcia-Carmona
F: Kojic acid, a cosmetic skin whitening agent, is a slow-binding
inhibitor of catecholase activity of tyrosinase. J Pharm Pharmacol.
46:982–985. 1994.PubMed/NCBI View Article : Google Scholar
|
|
112
|
Picardo M and Carrera M: New and
experimental treatments of cloasma and other hypermelanoses.
Dermatol Clin. 25353–362. (ix)2007.PubMed/NCBI View Article : Google Scholar
|
|
113
|
Dooley TP, Gadwood RC, Kilgore K and
Thomasco LM: Development of an in vitro primary screen for skin
depigmentation and antimelanoma agents. Skin Pharmacol. 7:188–200.
1994.PubMed/NCBI View Article : Google Scholar
|
|
114
|
Curto EV, Kwong C, Hermersdörfer H, Glatt
H, Santis C, Virador V and Hearing VJ Jr and Dooley TP: Inhibitors
of mammalian melanocyte tyrosinase: In vitro comparisons of alkyl
esters of gentisic acid with other putative inhibitors. Biochem
Pharmacol. 57:663–672. 1999.PubMed/NCBI View Article : Google Scholar
|
|
115
|
Hsu KD, Chen HJ, Wang CS, Lum CC, Wu SP,
Lin SP and Cheng KC: Extract of ganoderma formosanum mycelium as a
highly potent tyrosinase inhibitor. Sci Rep.
6(32854)2016.PubMed/NCBI View Article : Google Scholar
|
|
116
|
Peng CC, Sun HT, Lin IP, Kuo PC and Li JC:
The functional property of royal jelly 10-hydroxy-2-decenoic acid
as a melanogenesis inhibitor. BMC Complement Altern Med.
17(392)2017.PubMed/NCBI View Article : Google Scholar
|
|
117
|
Chen WC, Tseng TS, Hsiao NW, Lin YL, Wen
ZH, Tsai CC, Lee YC, Lin HH and Tsai KC: Discovery of highly potent
tyrosinase inhibitor, T1, with significant anti-melanogenesis
ability by zebrafish in vivo assay and computational molecular
modeling. Sci Rep. 5(7995)2015.PubMed/NCBI View Article : Google Scholar
|
|
118
|
Chang TS and Chen CT: Inhibitory effect of
homochlorcyclizine on melanogenesis in α-melanocyte stimulating
hormone-stimulated mouse B16 melanoma cells. Arch Pharm Res.
35:119–127. 2012.PubMed/NCBI View Article : Google Scholar
|
|
119
|
Newton RA, Cook AL, Roberts DW, Leonard JH
and Sturm RA: Post-transcriptional regulation of melanin
biosynthetic enzymes by cAMP and resveratrol in human melanocytes.
J Invest Dermatol. 127:2216–2227. 2007.PubMed/NCBI View Article : Google Scholar
|
|
120
|
Ando H, Wen ZM, Kim HY, Valencia JC,
Costin GE, Watabe H, Yasumoto K, Niki Y, Kondoh H, Ichihashi M, et
al: Intracellular composition of fatty acid affects the processing
and function of tyrosinase through the ubiquitin-proteasome
pathway. Biochem J. 394:43–50. 2006.PubMed/NCBI View Article : Google Scholar
|
|
121
|
Park SH, Kim DS, Kim WG, Ryoo IJ, Lee DH,
Huh CH, Youn SW, Yoo ID and Park KC: Terre in: A new melanogenesis
inhibitor and its mechanism. Cell Mol Life Sci. 61:2878–2885.
2004.PubMed/NCBI View Article : Google Scholar
|
|
122
|
Lee S, Kim WG, Kim E, Ryoo IJ, Lee HK, Kim
JN, Jung SH and Yoo ID: Synthesis and melanin biosynthesis
inhibitory activity of (+/-)-terrein produced by Penicillium sp.
20135. Bioorg Med Chem Lett. 15:471–473. 2005.PubMed/NCBI View Article : Google Scholar
|
|
123
|
Cheung FW, Guo J, Ling YH, Che CT and Liu
WK: Anti-melanogenic property of geoditin A in murine B16 melanoma
cells. Mar Drugs. 10:465–476. 2012.PubMed/NCBI View Article : Google Scholar
|
|
124
|
Minwalla L, Zhao Y, Cornelius J, Babcock
GF, Wickett RR, Le Poole IC and Boissy RE: Inhibition of melanosome
transfer from melanocytes to keratinocytes by lectins and
neoglycoproteins in an in vitro model system. Pigment Cell Res.
14:185–194. 2001.PubMed/NCBI View Article : Google Scholar
|
|
125
|
Seiberg M: Keratinocyte-melanocyte
interactions during melanosome transfer. Pigment Cell Res.
14:236–242. 2001.PubMed/NCBI View Article : Google Scholar
|
|
126
|
Hakozaki T, Minwalla L, Zhuang J, Chhoa M,
Matsubara A, Miyamoto K, Greatens A, Hillebrand GG, Bissett DL and
Boissy RE: The effect of niacinamide on reducing cutaneous
pigmentation and suppression of melanosome transfer. Br J Dermatol.
147:20–31. 2002.PubMed/NCBI View Article : Google Scholar
|
|
127
|
Paine C, Sharlow E, Liebel F, Eisinger M,
Shapiro S and Seiberg M: An alternative approach to depigmentation
by soybean extracts via inhibition of the PAR-2 pathway. J Invest
Dermatol. 116:587–595. 2001.PubMed/NCBI View Article : Google Scholar
|
|
128
|
Wallo W, Nebus J and Leyden JJ: Efficacy
of a soy moisturizer in photoaging: A double-blind,
vehicle-controlled, 12 week study. J Drugs Dermatol. 6:917–922.
2007.PubMed/NCBI
|
|
129
|
Lee CS, Nam G, Bae IH and Park J:
Whitening efficacy of ginsenoside F1 through inhibition of melanin
transfer in cocultured human melanocytes-keratinocytes and
three-dimensional human skin equivalent. J Ginseng Res. 43:300–304.
2019.PubMed/NCBI View Article : Google Scholar
|
|
130
|
Kuroda TS and Fukuda M: Rab27A-binding
protein Slp2-a is required for peripheral melanosome distribution
and elongated cell shape in melanocytes. Nat Cell Biol.
6:1195–1203. 2004.PubMed/NCBI View Article : Google Scholar
|
|
131
|
Wu XS, Rao K, Zhang H, Wang F, Sellers JR,
Matesic LE, Copeland NG, Jenkins NA and Hammer JA III:
Identification of an organelle receptor for myosin-Va. Nat Cell
Biol. 4:271–278. 2002.PubMed/NCBI View
Article : Google Scholar
|
|
132
|
Kudo M, Kobayashi-Nakamura K and
Tsuji-Naito K: Bifunctional effects of O-methylated flavones from
Scutellaria baicalensis Georgi on melanocytes: Inhibition of
melanin production and intracellular melanosome transport. PLoS
One. 12(e0171513)2017.PubMed/NCBI View Article : Google Scholar
|
|
133
|
Lee HY, Jang EJ, Bae SY, Jeon JE, Park HJ,
Shin J and Lee SK: Anti-melanogenic activity of gagunin D, a Highly
Oxygenated Diterpenoid from the Marine Sponge Phorbas sp., via
Modulating Tyrosinase Expression and Degradation. Mar Drugs.
14(pii: E212)2016.PubMed/NCBI View Article : Google Scholar
|
|
134
|
Ando H, Ryu A, Hashimoto A, Oka M and
Ichihashi M: Linoleic acid and alpha-linolenic acid lightens
ultraviolet-induced hyperpigmentation of the skin. Arch Dermatol
Res. 290:375–381. 1998.PubMed/NCBI View Article : Google Scholar
|
|
135
|
Yoshimura K, Tsukamoto K, Okazaki M,
Virador VM, Lei TC, Suzuki Y, Uchida G, Kitano Y and Harii K:
Effects of all-trans retinoic acid on melanogenesis in pigmented
skin equivalents and monolayer culture of melanocytes. J Dermatol
Sci. 27((Suppl 1): S68-S75)2001.PubMed/NCBI View Article : Google Scholar
|
|
136
|
Yoshimura K, Harii K, Aoyama T, Shibuya F
and Iga T: A new bleaching protocol for hyperpigmented skin lesions
with a high concentration of all-trans retinoic acid aqueous gel.
Aesthetic Plast Surg. 23:285–291. 1999.PubMed/NCBI View Article : Google Scholar
|
|
137
|
Ramos-e-Silva M, Hexsel DM, Rutowitsch MS
and Zechmeister M: Hydroxy acids and retinoids in cosmetics. Clin
Dermatol. 19:460–466. 2001.PubMed/NCBI View Article : Google Scholar
|
|
138
|
Gupta AK, Gover MD, Nouri K and Taylor S:
The treatment of melasma: A review of clinical trials. J Am Acad
Dermatol. 55:1048–1065. 2006.PubMed/NCBI View Article : Google Scholar
|
|
139
|
Amer M and Metwalli M: Topical liquiritin
improves melasma. Int J Dermatol. 39:299–301. 2000.PubMed/NCBI View Article : Google Scholar
|
|
140
|
Virador VM, Kobayashi N, Matsunaga J and
Hearing VJ: A standardized protocol for assessing regulators of
pigmentation. Anal Biochem. 270:207–219. 1999.PubMed/NCBI View Article : Google Scholar
|
|
141
|
Lei TC, Virador VM, Vieira WD and Hearing
VJ: A melanocyte-keratinocyte coculture model to assess regulators
of pigmentation in vitro. Anal Biochem. 305:260–268.
2002.PubMed/NCBI View Article : Google Scholar
|
|
142
|
Hermanns JF, Petit L, Piérard-Franchimont
C, Paquet P and Piérard GE: Assessment of topical hypopigmenting
agents on solar lentigines of asian women. Dermatology.
204:281–286. 2002.PubMed/NCBI View Article : Google Scholar
|
|
143
|
Tengamnuay P, Pengrungruangwong K,
Pheansri I and Likhitwitayawuid K: Artocarpus lakoocha heartwood
extract as a novel cosmetic ingredient: Evaluation of the in vitro
anti-tyrosinase and in vivo skin whitening activities. Int J Cosmet
Sci. 28:269–276. 2006.PubMed/NCBI View Article : Google Scholar
|
|
144
|
Alexis AF and Blackcloud P: Natural
ingredients for darker skin types: Growing options for
hyperpigmentation. J Drugs Dermatol. 12((9 Suppl):
s123-s127)2013.PubMed/NCBI
|