Introduction
Age-related cataract (ARC) is one of the most common
chronic diseases and a main cause of blindness worldwide (1). At present, surgery is the most
effective method of treating cataracts. However, surgery is an
economic burden to society, and has associated risks and
complications (2). On the basis
of the location of the opacity in the lens, ARC is classified as
cortical, nuclear or posterior sub-capsular cataract (3,4).
In the present study, the main focus was on nuclear cataract (S2)
and posterior capsule cataract (S3). ARC is associated with a
variety of factors, including age, sex, radiation (visible light,
ultraviolet and X-ray), oxidation, trauma, diet and drugs (5). ARC may also be associated with the
immune response and growth factors (6,7).
With the emergence of high-throughput sequencing
technology, long non-coding RNA (lncRNA) has emerged as an
alternative to microRNA (miRNA) as a research topic of great
interest. LncRNAs are transcripts that are >200 nucleotides in
length, and are similar in structure to mRNA but have little or no
protein-coding potential (8,9).
According to their genomic locations, lncRNAs may be divided into
several types, one of which is intronic lncRNAs (lincRNA), which
refers to lncRNA located within an intergenic region of the genome
(10). lncRNAs regulate gene
expression at the transcriptional, epigenetic or translational
level, and thereby alter cellular responses to various stresses
(11). Aberrant lncRNA expression
is implicated in several human diseases, including tumorigenesis,
neurological diseases and cardiovascular diseases (12–14). Certain lncRNAs have been
demonstrated to serve important roles in the development of the eye
and ocular diseases. Ventral anterior homeobox 2, opposite strand
(Vax2os), retinal non-coding RNA 2 (RNCR2), six3 opposite strand
(Six3OS) and taurine upregulated 1 (Tug1) have been indicated to
have an association with eye development: Vax2os1 controls the cell
cycle progression of photoreceptor progenitors in the mouse retina,
whereas RNCR2, Six3OS and Tug1 play key roles in the management of
retinal cell-specific differentiation (9,15).
In addition, metastasis associated lung adenocarcinoma transcript 1
(MALAT1) has been identified to have significantly increased
expression in the retinal tissue and aqueous humor of diabetic
mice, and in the vascular fiber membranes of diabetic patients
(16,17). However, the role of lncRNA in the
lens is unclear. Therefore, the present study was designed to
explore the possible regulation mechanism of lncRNAs in ARC.
Materials and methods
Clinical sample collection
The lens samples for the lncRNA microarray were
collected from postmortem donors (free from ocular diseases) (S1)
and ARC patients (9 patients, free of other ocular diseases; these
were divided into 2 groups: the S2 group included 3 patients with
nuclear cataract, and the S3 group included 6 patients with
posterior capsule cataract) at the Second Affiliated Hospital of
Harbin Medical University (Harbin, China) from November, 2014 to
January, 2015. All samples were obtained from male donors. The
average age of the normal control group was 36, and the average age
of the cataract patients was 65. The present study was approved by
the Human Ethics Review Board of the Second Affiliated Hospital of
Harbin Medical University (Harbin, China) and informed consent was
provided by all 9 cataract patients. All lens samples were obtained
by intact continuous curvilinear capsulorhexis, without vascular
contacting or damage to the iris or any other intraocular
structures. Since the RNA of a single lens was not sufficient for
microarray analysis, 3 cataract lenses were pooled together as a
biological repeat to obtain enough RNA. The degree of lenticular
opacification was determined using the Lens Opacities
Classification System II (18).
Tissue collection and RNA extraction
The collected samples were stored in liquid
nitrogen. Total RNAs were extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Waltham, MA, USA). The yield
of RNA was determined using a NanoDrop 2000 spectrophotometer
(Thermo Fisher Scientific, Inc.). LncRNA high throughput sequencing
analysis, including labelling, hybridization, scanning,
normalization and data analysis, was performed by Annoroad Geomics
(Beijing, China) (16,17).
RNA sequencing
A quality check of the input total RNA was performed
to confirm its integrity by running an aliquot on an Agilent
Bioanalyzer (Agilent Technologies, Inc., Santa Clara, CA, USA), and
the concentration of the RNA was measured using an ultra-micro
spectrophotometer. A 3-μg quantity of RNA was selected as
the initial amount of each sample from which to construct a lncRNA
library, following removal of the ribosomal RNA (rRNA) in the
sample using Ribo-Zero™ Gold kits (human/mouse/rat) (Epicentre,
Madison, WI, USA). Different index tags were selected according to
the NEB Next Ultra database direct operating protocol (NEB Next
Ultra Directional RNA Library Prep kit for Illumina;NEB Ipswich,
MA, USA; all reagents used here were provided in the kit). The
specific steps of library construction were as follows: Firstly,
the Ribo-Zero™ Gold kit was used to remove rRNA. Then,
fragmentation buffer was added to the reaction system to fragment
RNA into short segments, and the short fragments were used as
templates for first cDNA chain synthesis using six base random
primers (random hexamers). This was followed by second-strand cDNA
synthesis using DNA polymerase I and RNaseH. The cDNA fragments
were then subjected to an end-repair process, the addition of a
single 'A' base, and ligation of the Illumina multiplexing
adapters. The products were purified and enriched using polymerase
chain reaction (PCR) to create the final cDNA sequencing library,
as previously described (19–21). For PCR, the thermocycling
conditions were as follows: step 1, 65°C for 15 min; step 2, 30°C
for 10 min, 42–50°C for 45 min and 95°C for 5 min.
Data processing
The raw data was filtered to provide high-quality,
cleaned reads, and a follow-up analysis was then conducted. The
data processing steps were as follows: i) Removal of contamination
reads, ii) removal of low-quality reads and iii) removal of the
reads for bases having a nitrogen content >5% (5,22).
LncRNA analysis
Only lincRNA was analyzed in the present study.
According to the characteristics of the lincRNA, a series of strict
filters were used. The filter conditions were a length of ≥200 bp,
exon number of ≥2 and minimum coverage of ≥3 transcription reads.
Known mRNA transcripts, known non-coding RNA transcripts,
homologous transcripts and transcripts with protein-coding
capability were removed.
Read comparison
In this study, we used the TopHat software version
2.0.12 and selected the default parameters, and compared the
reference sequence to hg19 (31).
Differential expression analysis
The cataract samples and normal samples in each
group were subjected to differential expression analysis using
DESeq2. Differential expression analysis was then carried out
following standardization and variance estimation. The standards
for screening the expression differences of mRNA and lncRNA were
P<0.001, a false discovery rate (FDR) of <0.001 and a
|log2 ratio| >2 (21).
Gene set enrichment analysis
Gene Ontology (GO) enrichment and Kyoto Encyclopedia
of genes and genomes (KEGG) pathway enrichment analyses were
performed using the DAVID functional annotation web server
(http://david.abcc.ncifcrf.gov) using
default parameters. Enriched GO terms (FDR <0.05) and KEGG
pathways (P<0.1) were obtained (21–24).
Screening target genes
Cis target prediction was based on the distance
between the lncRNA its linked protein-coding gene and to forecast
the targets. Trans target prediction was according to the sequence
of lncRNA and mRNA sequence, and the evaluation of the combination
of free energy, which is a combination of the stability (25).
During the construction of all of the networks,
Cytoscape software was used. Cytoscape is an open source software
platform for visualizing molecular interaction networks and
biological pathways and integrating these networks with
annotations, gene expression profiles and other state data
(http://www.cytoscape.org/).
Conservation analysis
The conservation of the corresponding sequence and
site of the identified novel lincRNA was analyzed in order to
evaluate the extent of its variation. Conservation scores
(mammalian phastCons scores) were used to carry out the
conservation analysis, as previously described (26).
Reverse transcription-quantitative PCR
(RT-qPCR) validation of differentially expressed lncRNAs
Total RNA was extracted for use in microarray
experiments from cataract patients and postmortem eyes as described
above. Agarose gel electrophoresis was used to determine the
integrity of the RNA, and its concentration was determined using a
UV spectrophotometer. Quantification was performed with a two-step
reaction process: RT and qPCR. Each RT reaction consisted of 0.5 mg
RNA, 2 μl 5X PrimeScript buffer, 0.5 μl oligo(dT),
0.5 μl random 6 mers and 0.5 μl PrimeScript RT Enzyme
Mix I (Takara Bio, Inc., Otsu, Japan), with RNase Free
dH2O added to a total volume of 10 μl. Reactions
were performed using a GeneAmp PCR system 9700 (Applied Biosystems;
Thermo Fisher Scientific, Inc.) for 15 min at 37°C, followed by
heat inactivation of RT for 5 sec at 85°C. The 10-μl RT
reaction mix was then diluted 10-fold with nuclease-free water and
held at −20°C. qPCR was performed using a Stratagene Mx3000P
Real-time PCR instrument (Agilent Technologies, Inc., Santa Clara,
CA, USA) with 20 μl PCR reaction mixture comprising 100 ng
template DNA, 10 μl 2X GreenStar Master mix, 1 μl
forward primer, 1 μl reverse primer and PCR grade water to a
volume of 20 μl (Bioneer Corporation, Daejeon, Korea).
Reactions were incubated in a 384-well optical plate at 95°C for 10
min, followed by 40 cycles of 95°C for 10 sec, 60°C for 30 sec.
Each sample was run and analyzed in triplicate. Data quantification
was carried out using the 2-ΔΔCq method (27).
Results
Sequencing data analysis
The base distribution was analyzed, in order to
investigate the presence of the AT and GC separation phenomenon;
this phenomenon may arise during the sequencing or building of the
library, and will affect the subsequent quantitative analysis. It
was found that the frequencies of AT and Cg was almost the same,
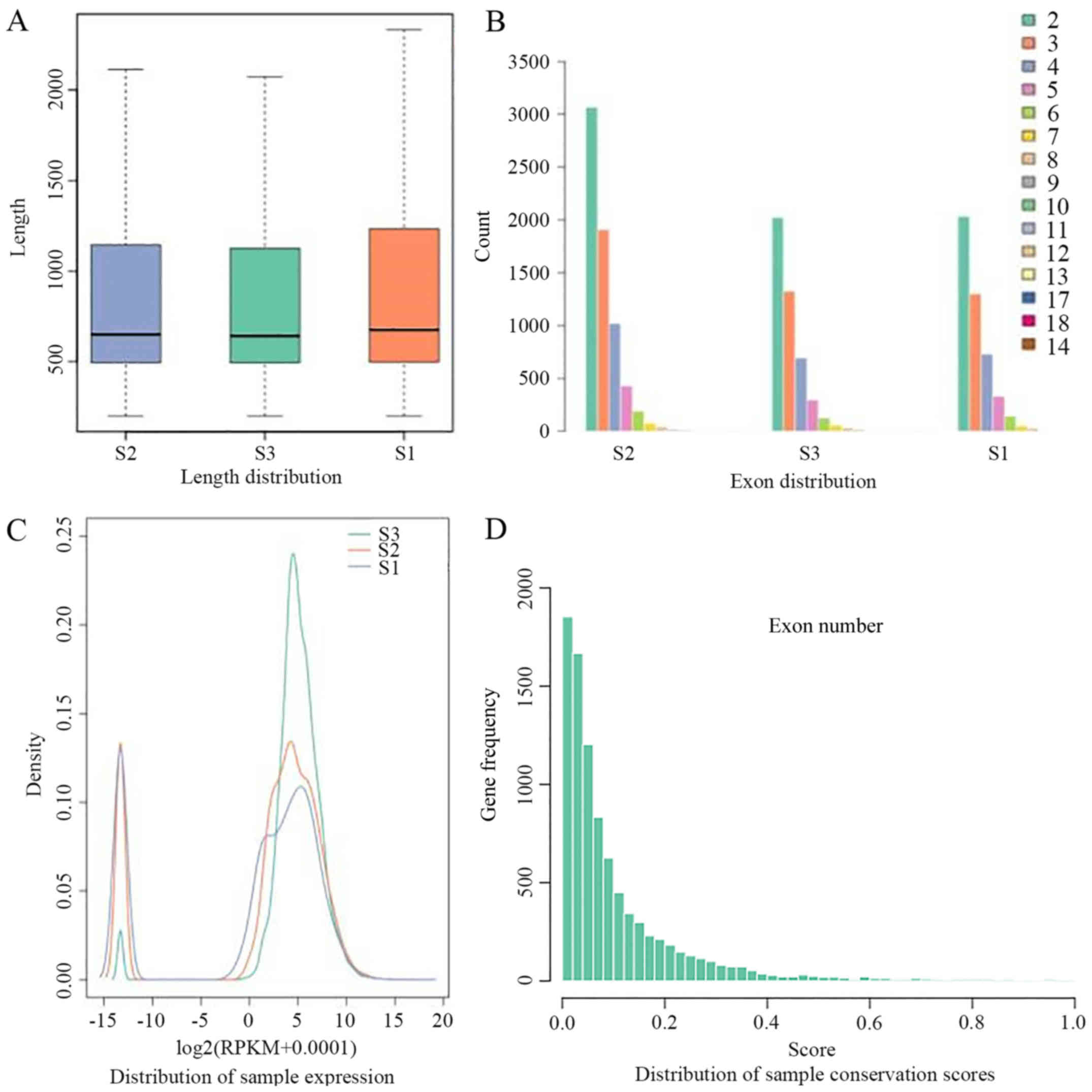
which confirmed the reliability of the obtained data (Fig. 1A and B).
Read comparison
TopHat software was used to compare the filtered
samples with the reference genome, and to locate the sequence
within the genome. When the reference genome selection is
appropriate and assembly is complete, in samples without exogenous
species contamination, the matching rate of the sample is usually
>80% (28–30) (Table
I). The total number of reads in S2 was 126,905,134, including
92% mapped reads and 4% multiMap reads. The total number of reads
in S3 was 102,287,578, including 96% mapped reads and 7% multiMap
reads. The number and proportion of matching sequences of three
functional components (exon, intron and intergenic) were confirmed.
The majority of the matched sequences in the three samples were in
the exon (Fig. 1C). In general,
if the species of annotation information comprehensive, the
majority of the sequence should match with the exon region
(31,32).
 | Table IComparison of samples [n (%)]. |
Table I
Comparison of samples [n (%)].
| Reads | S2 | S3 |
|---|
| Total reads | 126,905,134 | 102,287,578 |
| Mapped reads | 117,262,466
(92) | 98,284,753
(96) |
| MultiMap reads | 4,738,594 (4) | 6,924,023 (7) |
Estimation of mRNA expression
Gene expression levels are generally measured on the
basis of the mRNA transcription number of the gene. The units reads
per kilobase million mapped reads (RPKM) were used as a measure of
gene expression. It was observed that the number of differentially
expressed genes accounted for only a small proportion of all genes,
and so these genes are likely to have little impact on the overall
distribution of genes in the samples, and all samples should have a
similar distribution of expression (Fig. 1D) (33).
Characteristics of the novel lincRNA
In order to better define the characteristics of the
novel lincRNA, its length (Fig.
2A), the number of exons (Fig.
2B) and expression quantity (Fig.
2C) were analyzed. The distribution of lincRNA expression was
calculated according to the gene expression in all groups, and it
was observed that all samples were similar in expression. The
differentially expressed genes account for only a small fraction of
the overall gene (34).
Differential expression and functional
analysis of mRNA
DEseq was used to conduct a differential expression
analysis between S2 and S1, and between S3 and S1 using q<0.05
(35). The analysis identified
27,447 and 15,019 candidate differentially expressed mRNAs,
respectively, in these two comparisons. Hierarchical cluster
analysis was conducted for the classification of candidate mRNAs in
each group. Then, fold change values were used to classify the
candidate mRNAs into upregulated mRNAs (fold change ≥2) and
downregulated mRNAs (fold change ≤0.05). For S2 vs. S1 and S3 vs.
S1, 24,947 and 10,478 upregulated mRNAs and 2,500 and 4,631
downregulated mRNAs, respectively, were identified (Table II). Further analysis of the
differential expression is presented in Fig. 3. Intersections of the
differentially expressed mRNAs were detected, and it was found that
there were 11,348 differentially expressed mRNAs, 9,261 upregulated
mRNAs and 2,087 downregulated mRNAs (Fig. 3C). Furthermore, an enrichment
analysis was performed to investigate the functions of these
upregulated and downregulated mRNAs. These mRNAs, whether
upregulated or downregulated, were enriched in several basic
biological or nerve-related terms and pathways, such as the
phenylalanine metabolism pathway (Fig. 3A and B). Notable GO terms included
cytoskeletal protein binding and actin binding. The phenylalanine
metabolism pathway and cytoskeletal protein binding and actin
binding are the biological processes of the marked enrichment of
all data after processing (data not shown). In particular, the
mRNAs were significantly rich in GO terms associated with
structural constituents of the eye lens, which was related to the
function and nerves of eyes. Some researchers considered ARC to be
a structural disease (36). The
lens is composed of >90% crystalline protein, with the remainder
comprising skeletal, membrane, connective and water channel
proteins (37). A large number of
studies have indicated that the occurrence of cataract is directly
associated with the apoptosis of lens epithelial cells induced by
the modification of lens proteins, such as changes in the ratio of
proteins or protein structure (38–40). The results of the present study
revealed various differentially expressed mRNAs that were strongly
associated with the function of the eyes. In the KEGG pathway
analysis (data not shown), downregulated mRNA expression was
particularly enriched in the tyrosine metabolism pathway, which is
closely associated with cataract. The abnormal metabolism of
tyrosine in the lens results in the production of quinones, which
may be oxidized and thereby damage the protein of the lens, causing
lens opacity and ultimately the occurrence of cataract (40–42). In addition, tyrosine is able to
bind to lens proteins through phosphorylation, which may also have
an association with cataract (43).
 | Figure 3GO analysis of the differentially
expressed genes for (A) S2 vs. S1 and (B) S3 vs. S1. The horizontal
coordinates for the large classes under the GO annotation represent
a variety of biological processes, cellular components and
molecular functions. The x axis is a variety of biological
processes, cell components, and molecular functions of GO. The left
vertical coordinate is the proportion of the class, and the right
vertical is the specific gene number of the class. Different colors
represent different groups. Venn diagrams showing the intersection
of the differentially expressed (C) mRNAs (upper diagrams,
upregulated mRNAs; lower diagrams, downregulated mRNAs) and (D)
long non-coding RNAs (upper diagrams, upregulated lncRNAs; lower
diagrams, downregulated lncRNAs). GO, gene ontology; S1, normal
eyes; S2, nuclear cataract; S3, posterior capsule cataract. |
| Table IIDifferential expression of mRNAs. |
Table II
Differential expression of mRNAs.
| Expression | S2 vs. S1 | S3 vs. S1 |
|---|
| Upregulated | 24,947 | 10,478 |
| Downregulated | 2,500 | 4,613 |
| Total | 27,447 | 15,109 |
Differential expression and functional
analysis of lncRNA
Due to the low expression feature of lncRNAs,
q<1.1 was used to identify the differentially expressed lncRNAs
from the novel lincRNAs between S2 and S1, and between S3 and S1.
This revealed that there were 4,146 and 2,895 differentially
expressed lncRNAs, respectively (Table III). Fold-change values were
used to classify the candidate lncRNAs into upregulated lncRNAs
(fold change ≥2) and downregulated lncRNAs (fold change ≤0.05), and
3,477 and 2,051 upregulated lncRNAs and 669 and 844 downregulated
lncRNAs were identified, respectively. Intersections of the lncRNAs
were also obtained for each group. There were found to be 2,269
common differentially expressed lncRNAs, 1,701 upregulated lncRNAs
and 568 downregulated lncRNAs (Fig.
3D). Hierarchical cluster analysis was also performed, to
classify the candidate lncRNAs for each group.
| Table IIIDifferential expression of
lncRNAs. |
Table III
Differential expression of
lncRNAs.
| Expression | S2 vs. S1 | S3 vs. S1 |
|---|
| Upregulated | 3,477 | 2,051 |
| Downregulated | 669 | 844 |
| Total | 4,146 | 2,895 |
LncRNA-mRNA network
The 12,097 and 2,332 differential mRNAs and lncRNAs
were further analyzed by integrating all differential mRNAs and
lncRNAs for each group using intersection sets. Firstly,
comprehensive mRNA-lncRNA interactions were obtained by integrating
cis-targets and trans-targets. The interactions were also
classified into upregulated and downregulated interactions using
the dysregulated mRNAs. The functions of the dysregulated lncRNAs
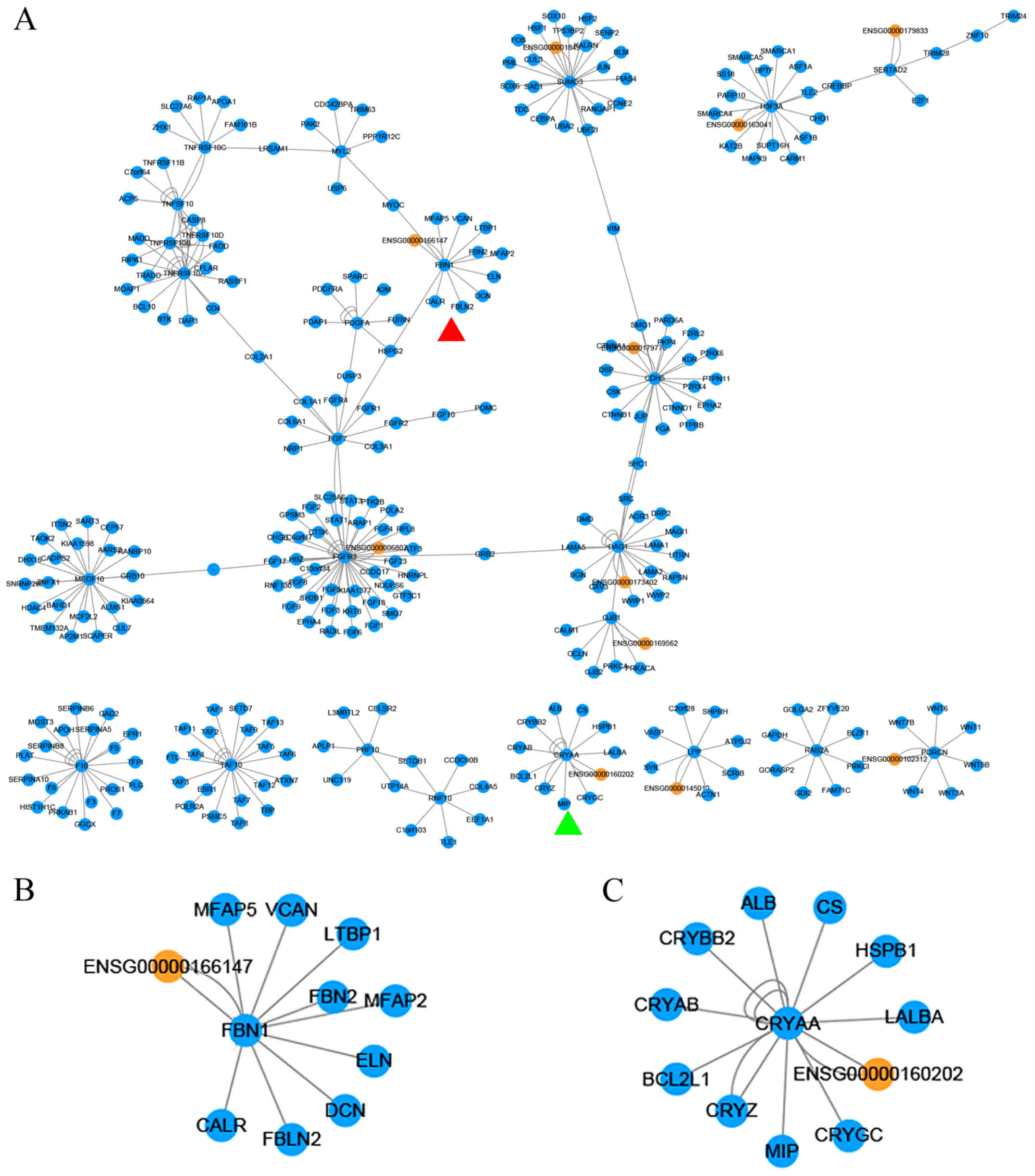
were analyzed on the basis of their target mRNAs (Figs. 4 and 5). It was observed that these
dysregulated lncRNAs were enriched in certain basic pathways and GO
terms, including regulation of the actin cytoskeleton pathway and
protein binding terms. The result was coincident with the
dysregulated mRNAs. Genes such as paired box 6 and chaperone-like
activity of αA-crystallin (CRYAA), have similar functions. The
lncRNA target gene major intrinsic protein of lens fiber (MIP,
ENSG00000135517) encodes the most abundant junctional membrane
protein in the mature lens and this protein serves a critical role
in the maintenance of the normal structure and internal circulation
of the lens (44,45). In ARC, CRYAA is considered to be
critical for the maintenance of lens transparency (46,47). In the GO analysis, it was found
that ENSG00000160202 and CRYAA were linked nodes (Fig. 4C), indicating that they have
similar biological functions (48). ENSG00000166147 and fibrillin-1
(FBN1) were also linked nodes (Fig.
4B). FBN1 is the most common pathogenic gene for Marfan
syndrome, and encodes the fibrillin-1 protein. Eye diseases
including lens dislocation or subluxation, and cataract, are
observed in typical Marfan syndrome patients. Studies have
suggested that fibrillin-1 is not a component of the normal lens,
but may serve as a connecter between latent-transforming growth
factor β-binding protein 1 and the extracellular matrix (ECM) in
capsular opacification. In addition, FBN1 is an important gene in
eye development (49,50).
Furthermore, a human protein-protein interaction
(PPI) network based on the HPRD data in the present study was
constructed. The largest sub-network of the upregulated and
downregulated mRNAs was then obtained from the PPI network. During
the construction of all of the networks, the software Cytoscape was
used. The upregulated network included 359 nodes and 395 edges, and
the downregulated network included 305 nodes and 346 edges. The two
networks approximated the scale-free network topology of
dysregulated networks. The connectivity and betweenness of nodes
were also analyzed. Some other networks also have similar
topological features (51,52)
(data not shown).
Conservation analysis
The PhastCons score of each chromosome was
downloaded, which contains all loci on each gene from the
University of California, Santa Cruz database, in order to analyze
the conservative of lncRNA. The conservation scores of the majority
of the lncRNAs in the present study (Fig. 2D) were at a low level (53).
RT-qPCR validation
To validate the results of sequencing, 30
differentially expressed lncRNA transcripts were selected for
validation using RT-qPCR. The RT-qPCR results were consistent with
the sequencing results for 28 of the lncRNA transcripts, showing
the same trends of differential expression (P<0.05). Of note,
due to individual differences or other reasons, two were
inconsistent with the sequencing results (Table IV). The specific mechanisms need
to be further explored.
| Table IVValidation of sequencing results by
RT-qPCR. |
Table IV
Validation of sequencing results by
RT-qPCR.
| Target of
lncRNA | High-throughput
sequencing | RT-qPCR |
|---|
|
ENSG00000188536 | Upregulated | Upregulated |
|
ENSG00000206172 | Upregulated | Upregulated |
|
ENSG00000225282 | Upregulated | Upregulated |
|
ENSG00000175147 | Upregulated | Upregulated |
|
ENSG00000237988 | Upregulated | Upregulated |
|
ENSG00002225282 | Upregulated | Upregulated |
|
ENSG00000008517 | Upregulated | Upregulated |
|
ENSG00000077080 | Upregulated | Downregulated |
|
ENSG00000244734 | Upregulated | Upregulated |
|
ENSG00000196361 | Upregulated | Upregulated |
|
ENSG00000197414 | Upregulated | Upregulated |
|
ENSG00000171346 | Upregulated | Upregulated |
|
ENSG00000253418 | Upregulated | Upregulated |
|
ENSG00000262516 | Upregulated | Upregulated |
|
ENSG00000118231 | Upregulated | Upregulated |
|
ENSG00000146166 | Downregulated | Downregulated |
|
ENSG00000100122 | Downregulated | Downregulated |
|
ENSG00000135517 | Downregulated | Downregulated |
|
ENSG00000196431 | Downregulated | Downregulated |
|
ENSG00000122986 | Downregulated | Downregulated |
|
ENSG00000233797 | Downregulated | Upregulated |
|
ENSG00000106868 | Downregulated | Downregulated |
|
ENSG00000082515 | Downregulated | Downregulated |
|
ENSG00000138376 | Downregulated | Downregulated |
|
ENSG00000105325 | Downregulated | Downregulated |
|
ENSG00000136158 | Downregulated | Downregulated |
|
ENSG00000186314 | Downregulated | Downregulated |
|
ENSG00000134258 | Downregulated | Downregulated |
|
ENSG00000213949 | Downregulated | Downregulated |
|
ENSG00000182218 | Downregulated | Downregulated |
Discussion
Cataract is the leading cause of blindness globally,
and its incidence increases yearly (1). To date, the pathogenesis of cataract
has mainly been investigated at the cellular and molecular levels,
including the post-translational modification, physical and
chemical factors of lens proteins. miRNA has tissue and cell
specificity in the eye. It serves an important regulatory role in
the regulation of cell proliferation, differentiation and apoptosis
processes (54).
With the rapid development of high-throughput
sequencing and bioinformatics technology, lncRNAs have been
identified as a class of non-encoding RNAs abundantly expressed in
mammalian genomes; however, their functions are mostly unknown. The
study of lncRNA has revealed that although their sequences are less
conservative than those of mRNAs, the promoter and functional
components are highly conservative. This indicates that lncRNA may
have a specific function (55).
There is evidence to support the hypothesis that lncRNAs are able
to regulate the expression of protein-encoding genes, including the
stability and subcellular localization of proteins. This is
involved in a wide variety of biological processes, such as
imprinting control, cell differentiation and immune response
(56). The role of lncRNA in the
eye has attracted attention. Several lncRNAs including Vax2os,
RNCR2, Six3OS and Tug1, have been confirmed to be associated with
the development of the eye. Vax2os controls the cell cycle
progression of receptors in mouse retina; RNCR2, Six3OS and Tug1
serve key roles in the differentiation of retinal cells (57,58). Another lncRNA, MIAT, has an effect
on the proliferation and differentiation of lens epithelial cells
(59). However, the function of
lncRNA in cataract is not yet clear.
In the present study, high-throughput sequencing
revealed a large number of differentially expressed lncRNAs and
mRNAs in ARC compared with ocular disease-free eyes (from
postmortem donors). Construction of an lncRNA-mRNA network was used
to analyze the functions of the lncRNA. The reliability of the
differential expression of the lncRNAs was evaluated using RT-qPCR.
The RT-qPCR results for 28 of the 30 selected differentially
expressed lncRNAs were consistent with the sequencing results.
LncRNA is RNA characterized by abundant expression but low
conservation, low cell expression specificity and low transcription
characteristics (60). As a
result, it is difficult to directly identified the function of
lncRNA via the nucleic acid sequences with current technologies.
Therefore, the present study focused on their biological functions
and the regulation pathways of differentially expressed genes using
bioinformatics, to provide a theoretical basis for further study of
the roles of lncRNAs in cataract. The lncRNA-mRNA expression
network was constructed and GO analysis of the differentially
expressed mRNA was conducted. In order to promote the accuracy of
analysis, the intersections of the differentially expressed mRNAs
were determined. It was found there were 12,097 differentially
expressed mRNAs, 9,261 upregulated mRNAs and 2,087 downregulated
mRNAs. These mRNAs were enriched in several basic biological or
nerve-related terms and pathways, such as sodium symporter
activity, neurotransmitter transporter activity and cytoskeletal
protein binding. Notably, some were significantly enriched in GO
terms relating to structural constituents of the eye lens. This
demonstrated the complexity of the regulatory mechanism. KEGG
pathway analysis of differentially expressed mRNAs in the
lncRNA-mRNA expression networks was also conducted. The results
reveal that genes involved in biological pathways including
tyrosine metabolism, phenylalanine metabolism and regulation of the
actin cytoskeleton. These signaling pathways are associated with
numerous pathological processes. Thus, it appears that lncRNAs and
the differentially expressed gene network are involved in the
regulation of multiple cellular processes. Therefore, they may be a
novel target for the treatment of cataract and associated diseases,
which may have potential clinical significance.
It is worthy of note that the calcium signaling
pathway, and regulation of the actin cytoskeleton, are closely
associated with the underlying pathological processes of a number
of diseases. The calcium ion is the most common signal transduction
factor in the human body, and plays an important role in cell
division, growth and death. Calcium ions act as the second
messenger in the regulation of cell apoptosis, which is known to be
one of the main causes of ARC (61,62). It may be speculated that the
involvement of the lncRNAs in the pathogenesis of ARC involves
regulation of the calcium signaling pathway, which provides new
opportunities for the diagnosis and treatment of ARC. The lens is a
fascinating and unique transparent tissue that grows continuously
throughout life. During the process of differentiation into fiber
cells, lens epithelial cells undergo marked morphological changes,
membrane remodeling, polarization, transcriptional activation and
the elimination of cellular organelles, including nuclei,
concomitant with migration towards the lens interior (63,64). The majority of these events are
considered to be influenced by dynamic reorganization of the
cellular actin cytoskeleton and by intercellular and cell (65). These observations suggest a number
of new directions for exploration with regard to the pathogenic
effects of lncRNA.
Acknowledgments
The authors are grateful to all patients and normal
volunteers for their participation in this study. This study was
supported by the National Natural Science Foundation of China,
Harbin, Y.Q. (grant no. 81170833).
References
|
1
|
Resnikoff S and Keys TU: Future trends in
global blindness. Indian J Ophthalmol. 60:387–395. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pescosolido N, Barbato A, Giannotti R,
Komaiha C and Lenarduzzi F: Age-related changes in the kinetics of
human lenses: Prevention of the cataract. Int J Ophthalmol.
9:1506–1517. 2016.PubMed/NCBI
|
|
3
|
Hammond CJ, Duncan DD, Snieder H, de Lange
M, West SK, Spector TD and Gilbert CE: The heritability of
age-related cortical cataract: The twin eye study. Invest
Ophthalmol Vis Sci. 42:601–605. 2001.PubMed/NCBI
|
|
4
|
Tsai SY, Hsu WM, Cheng CY, Liu JH and Chou
P: Epidemiologic study of age-related cataracts among an elderly
Chinese population in Shih-Pai, Taiwan. Ophthalmology.
110:1089–1095. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sheng Y, He F, Lin JF, Shen W and Qiu YW:
Tea and risk of age-related cataracts: A cross-sectional study in
Zhejiang Province, China. J Epidemiol. 26:587–592. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen W, Lin H, Zhong X, Liu Z, Geng Y, Xie
C and Chen W: Discrepant expression of cytokines in inflammation-
and age-related cataract patients. PloS One. 9:e1096472014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hwang HB, Yim HB, Cho YK and Choi JA: The
Association Between Aqueous Connective Tissue growth Factor and the
Severity of Age-related Cataracts as Graded by the Lens Opacities
Classification System III. Curr Eye Res. 41:350–356. 2016.
View Article : Google Scholar
|
|
8
|
Wilusz JE, Sunwoo H and Spector DL: Long
noncoding RNAs: Functional surprises from the RNA world. Genes Dev.
23:1494–1504. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yan B, Tao ZF, Li XM, Zhang H, Yao J and
Jiang Q: Aberrant expression of long noncoding RNAs in early
diabetic retinopathy. Invest Ophthalmol Vis Sci. 55:941–951. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li J, Xuan Z and Liu C: Long non-coding
RNAs and complex human diseases. Int J Mol Sci. 14:18790–18808.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Uchida S and Dimmeler S: Long noncoding
RNAs in cardiovascular diseases. Circ Res. 116:737–750. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao Q, Li T, Qi J, Liu J and Qin C: The
miR-545/374a cluster encoded in the Ftx lncRNA is overexpressed in
HBV-related hepatocellular carcinoma and promotes tumorigenesis and
tumor progression. PloS One. 9:e1097822014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roberts TC, Morris KV and Wood MJ: The
role of long non-coding RNAs in neurodevelopment, brain function
and neurological disease. Philos Trans R Soc Lond B Biol Sci.
369:pii: 20130507. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meola N, Pizzo M, Alfano G, Surace EM and
Banfi S: The long noncoding RNA Vax2os1 controls the cell cycle
progression of photoreceptor progenitors in the mouse retina. RNA.
18:111–123. 2012. View Article : Google Scholar :
|
|
16
|
Shen Y, Dong LF, Zhou RM, Yao J, Song YC,
Yang H, Jiang Q and Yan B: Role of long non-coding RNA MIAT in
proliferation, apoptosis and migration of lens epithelial cells: A
clinical and in vitro study. J Cell Mol Med. 20:537–548. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao J, Wang XQ, Li YJ, Shan K, Yang H,
Wang YN, Yao MD, Liu C, Li XM, Shen Y, et al: Long non-coding RNA
MALAT1 regulates retinal neurodegeneration through CREB signaling.
EMBO Mol Med. 8:346–362. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chylack LT Jr, Leske MC, McCarthy D, Khu
P, Kashiwagi T and Sperduto R: Lens opacities classification system
II (LOCS II). Arch Ophthalmol. 107:991–997. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kissopoulou A, Jonasson J, Lindahl TL and
Osman A: Next generation sequencing analysis of human platelet
PolyA+ mRNAs and rRNA-depleted total RNA. PLoS One.
8:e818092013. View Article : Google Scholar
|
|
20
|
Lin Y, Golovnina K, Chen ZX, Lee HN,
Negron YL, Sultana H, Oliver B and Harbison ST: Comparison of
normalization and differential expression analyses using RNA-Seq
data from 726 individual Drosophila melanogaster. BMC Genomics.
17:282016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang ZH, Jhaveri DJ, Marshall VM, Bauer
DC, Edson J, Narayanan RK, Robinson GJ, Lundberg AE, Bartlett PF,
Wray NR, et al: A comparative study of techniques for differential
expression analysis on RNA-Seq data. PLoS One. 9:e1032072014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu Z, Fu Y, Cao J, Yu M, Tang X and Zhao
S: Identification of differentially expressed miRNAs between white
and black hair follicles by RNA-sequencing in the goat (Capra
hircus). Int J Mol Sci. 15:9531–9545. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shah DH: RNA sequencing reveals
differences between the global transcriptomes of Salmonella
enterica serovar enteritidis strains with high and low
pathogenicities. Appl Environ Microbiol. 80:896–906. 2014.
View Article : Google Scholar :
|
|
24
|
Huang S, Feng C, Chen L, Huang Z, Zhou X,
Li B, Wang LL, Chen W, Lv FQ and Li TS: Identification of potential
key long non-coding RNAs and target genes associated with pneumonia
using long non-coding RNA sequencing (lncRNA-Seq): A preliminary
study. Med Sci Monit. 22:3394–3408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wainberg M, Alipanahi B and Frey B: Does
conservation account for splicing patterns? BMC Genomics.
17:7872016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang CH, Wang YT, Tsai CF, Chen YJ, Lee
JS and Chiou SH: Phosphoproteomics characterization of novel
phosphorylated sites of lens proteins from normal and cataractous
human eye lenses. Mol Vis. 17:186–198. 2011.PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Finotello F and Di Camillo B: Measuring
differential gene expression with RNA-seq: challenges and
strategies for data analysis. Brief Funct Genomics. 14:130–142.
2015. View Article : Google Scholar
|
|
29
|
Kumar R, Ichihashi Y, Kimura S, Chitwood
DH, Headland LR, Peng J, Maloof JN and Sinha NR: A High-Throughput
Method for Illumina RNA-Seq Library Preparation. Front Plant Sci.
3:2022012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yu J, He K, Ren T, Lou Y and Zhao A:
High-throughput sequencing reveals differential expression of
miRNAs in prehierarchal follicles of laying and brooding geese.
Physiol Genomics. 48:455–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Langmead B, Trapnell C, Pop M and Salzberg
SL: Ultrafast and memory-efficient alignment of short DNA sequences
to the human genome. Genome Biol. 10:R252009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wagner GP, Kin K and Lynch VJ: Measurement
of mRNA abundance using RNA-seq data: RPKM measure is inconsistent
among samples. Theory Biosci. 131:281–285. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X and Yan GY: Novel human
lncRNA-disease association inference based on lncRNA expression
profiles. Bioinformatics. 29:2617–2624. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang L, Feng Z, Wang X, Wang X and Zhang
X: DEGseq: An R package for identifying differentially expressed
genes from RNA-seq data. Bioinformatics. 26:136–138. 2010.
View Article : Google Scholar
|
|
36
|
Panda AK, Nandi SK, Chakraborty A, Nagaraj
RH and Biswas A: Differential role of arginine mutations on the
structure and functions of alpha-crystallin. Biochim Biophys Acta.
1860:199–210. 2016. View Article : Google Scholar
|
|
37
|
Antosova B, Smolikova J, Borkovcova R,
Strnad H, Lachova J, Machon O and Kozmik Z: Ectopic activation of
Wnt/beta-catenin signaling in lens fiber cells results in cataract
formation and aberrant fiber cell differentiation. PloS One.
8:e782792013. View Article : Google Scholar
|
|
38
|
Li Y, Jia Y, Zhou J and Huang K: Effect of
methionine sulfoxide reductase B1 silencing on high-glucose-induced
apoptosis of human lens epithelial cells. Life Sci. 92:193–201.
2013. View Article : Google Scholar
|
|
39
|
Linetsky M, Raghavan CT, Johar K, Fan X,
Monnier VM, Vasavada AR and Nagaraj RH: UVA light-excited
kynurenines oxidize ascorbate and modify lens proteins through the
formation of advanced glycation end products: implications for
human lens aging and cataract formation. J Biol Chem.
289:17111–17123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Staniszewska MM and Nagaraj RH:
3-hydroxykynurenine-mediated modification of human lens proteins:
structure determination of a major modification using a monoclonal
antibody. J Biol Chem. 280:22154–22164. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ipson BR and Fisher AL: Roles of the
tyrosine isomers meta-tyrosine and ortho-tyrosine in oxidative
stress. Ageing Res Rev. 27:93–107. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Srivastava SK and Beutler E: Cataract
produced by tyrosinase and tyrosine systems in rabbitens in vitro.
Biochem J. 112:421–425. 1969. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu X, Zhou P, Fan F, Li D, Wu J, Lu Y and
Luo Y: CpG site methylation in CRYAA promoter affect transcription
factor Sp1 binding in human lens epithelial cells. BMC Ophthalmol.
16:1412016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang J, Zhou S, Guo M, Li Y and Gu J:
Different alpha crystallin expression in human age-related and
congenital cataract lens epithelium. BMC Ophthalmol. 16:672016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou Z, Wang B, Luo Y, Zhou G, Hu S, Zhang
H, Ma X and Qi Y: Major intrinsic protein (MIP) polymorphism is
associated with age-related cataract in Chinese. Mol Vis.
17:2292–2296. 2011.PubMed/NCBI
|
|
46
|
Takemoto L and Sorensen CM:
Protein-protein interactions and lens transparency. Exp Eye Res.
87:496–501. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ma X, Jiao X, Ma Z and Hejtmancik JF:
Polymorphism rs7278468 is associated with Age-related cataract
through decreasing transcriptional activity of the CRYAA promoter.
Sci Rep. 6:232062016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bhagyalaxmi SG, Srinivas P, Barton KA,
Kumar KR, Vidyavathi M, Petrash JM, Reddy GB and Padma T: A novel
mutation (F71L) in αA-crystallin associated with age-related
cataract results in defective chaperone-like function despite
unaltered structure. Biochim Biophys Acta. 1792:974–981. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hubmacher D, Reinhardt DP, Plesec T,
Schenke-Layland K and Apte SS: Human eye development is
characterized by coordinated expression of fibrillin isoforms.
Invest Ophthalmol Vis Sci. 55:7934–7944. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Saika S, Miyamoto T, Tanaka T, Ishida I,
Ohnishi Y and Ooshima A: Latent TGFbeta binding protein-1 and
fibrillin-1 in human capsular opacification and in cultured lens
epithelial cells. Br J Ophthalmol. 85:1362–1366. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ning S, Gao Y, Wang P, Li X, Zhi H, Zhang
Y, Liu Y, Zhang J, Guo M, Han D, et al: Construction of a
lncRNA-mediated feed-forward loop network reveals global
topological features and prognostic motifs in human cancers.
Oncotarget. 7:45937–45947. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu J, Feng L, Han Z, Li Y, Wu A, Shao T,
Ding N, Li L, Deng W, Di X, et al: Extensive ceRNA-ceRNA
interaction networks mediated by miRNAs regulate development in
multiple rhesus tissues. Nucleic Acids Res. 44:9438–9451.
2016.PubMed/NCBI
|
|
53
|
Marques AC and Ponting CP: Catalogues of
mammalian long noncoding RNAs: Modest conservation and
incompleteness. Genome Biol. 10:R1242009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu C, Lin H, Wang Q, Chen W, Luo H, Chen W
and Zhang H: Discrepant expression of microRNAs in transparent and
cataractous human lenses. Invest Ophthalmol Vis Sci. 53:3906–3912.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
McLaughlin PA, Nirmalan GP, Tam KH and
Robinson GA: Release of 131I from the ovary of the laying Japanese
quail after injection of perchlorate, thiocyanate or iodide. J
Endocrinol. 63:337–342. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
McIllmurray MB, Price MR and Langman MJ:
Inhibition of leucocyte migration in patients with large intestinal
cancer by extracts prepared from large intestinal tumours and from
normal colonic mucosa. Br J Cancer. 29:305–311. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Young TL, Matsuda T and Cepko CL: The
noncoding RNA taurine upregulated gene 1 is required for
differentiation of the murine retina. Curr Biol. 15:501–512. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cumming GR, Dufresne C, Kich L and Samm J:
Exercise electrocardiogram patterns in normal women. Br Heart J.
35:1055–1061. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Gosak M, Markovič R, Fajmut A, Marhl M,
Hawlina M and Andjelić S: The analysis of intracellular and
intercellular calcium signaling in human anterior lens capsule
epithelial cells with regard to different types and stages of the
cataract. PLoS One. 10:e01437812015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Schmitz SU, Grote P and Herrmann BG:
Mechanisms of long noncoding RNA function in development and
disease. Cell Mol Life Sci. 73:2491–2509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sundararajan M, Thomas PA, Teresa PA,
Anbukkarasi M and Geraldine P: Regulatory effect of chrysin on
expression of lenticular calcium transporters, calpains, and
apoptotic-cascade components in selenite-induced cataract. Mol
Vision. 22:401–423. 2016.
|
|
62
|
Cekic O: Copper, lead, cadmium and calcium
in cataractous lenses. Ophthalmic Res. 30:49–53. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Fischer RS, Lee A and Fowler VM:
Tropomodulin and tropomyosin mediate lens cell actin cytoskeleton
reorganization in vitro. Invest Ophthalmol Vis Sci. 41:166–174.
2000.PubMed/NCBI
|
|
64
|
Zhou CJ and Lo WK: Association of
clathrin, AP-2 adaptor and actin cytoskeleton with developing
interlocking membrane domains of lens fibre cells. Exp Eye Res.
77:423–432. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gostishchev VK and Khokhlov AM:
Pathogenesis of trophic ulcers in varicose veins of the lower
extremities. Khirurgiia (Mosk). 10:100–105. 1991.
|