Introduction
Senescence may be caused by telomere uncapping due
to replicative exhaustion, mitochondrial deterioration, oxidative
stress, severe or irreparable DNA damage or selected oncogene
expression (1). Endothelial
senescence leads to cell dysfunction, which in turn eventually
results in cardiovascular diseases, including hypertension and
atherosclerosis (2). Oxidized
low-density lipoprotein (oxLDL) is an important risk factor in
promoting the development of atherosclerosis. oxLDL stimulates
endothelial cells (ECs) to produce adhesion molecules and
proinflammatory substances. The scavenger receptor lectin-like
oxLDL receptor-1 (LOX-1) mediates the binding and internalization
of oxLDL into cells. Accordingly, LOX-1 has a critical role in
endothelial dysfunction (3), and
LOX-1 has been suggested to be an aging marker (4).
Angiotensin II (Ang II) is the primary effector of
the renin-angiotensin system (RAS) and is a multifunctional hormone
that has a major role in regulating blood pressure and
cardiovascular homeostasis (5).
Mammalian cells express 2 types of Ang II receptors, Ang II type 1
receptor (AT1R) and AT2R (6,7).
Ang II-induced reactive oxygen species production via AT1R promotes
the onset of vascular senescence as well as functional and
structural changes in blood vessels (8). Ang II signaling is considered to
contribute to the pathogenesis of atherosclerosis (9). In addition, Ang II has been reported
to upregulate LOX-1, which facilitates oxLDL uptake by ECs
(10). However, it remains
elusive whether Ang II also deteriorates EC function via a
LOX-1-independent pathway.
Sirtuin 1 (SIRT1) is implicated in anti-aging
processes and has a role in the tolerance of oxidative stress
(11). A previous study by our
group demonstrated that micro (mi)RNA let-7g increased SIRT1
expression in ECs (12). SIRT1 is
highly expressed in blood vessels, and a decrease of SIRT1
expression may be a risk factor for atherosclerosis (13–15). The insulin-like growth factor 1
(IGF1) signal transduction pathway has been demonstrated to be
associated with the aging process (16,17). Furthermore, SIRT1 expression has
been negatively correlated with the level of IGF1 (18). SIRT1 has been linked to the
insulin/IGF signaling pathway in Caenorhabditis elegans,
drosophila and mice through its ability to deacetylate fork-head
box O proteins (19–21). Treatment of cells with IGF1 may
decrease their SIRT1 levels, suggesting an inverse association
between SIRT1 and the insulin/IGF pathway (20).
MicroRNAs (miRNAs) are small non-coding RNAs that
regulate gene expression through degradation or translational
inhibition of targeted mRNA (22). The miRNA let-7 family has a
pivotal role in cell proliferation, cancer and cardiovascular
diseases (23,24). A previous study by our group
reported that let-7g directly targets and suppresses LOX-1
(25). Further studies by our
group have reported that let-7g improves several endothelial
functions by causing a decrease of senescence, inflammation and
monocyte adhesion, and an increase of angiogenesis (12,25,26). The present study aimed to
investigate whether let-7g also provides endothelial protection via
a LOX-1-independent mechanism. Let-7g was tested for its anti-aging
effect against Ang II-induced senescence in ECs while LOX-1 was
suppressed.
Materials and methods
Cell culture and treatment
Human umbilical vein endothelial cells (HUVECs) were
purchased as pooled primary cells from Lonza (Basel, Switzerland).
The HUVECs were cultured at 37°C in an atmosphere of 5%
CO2 in Clonetics Endothelial Cell Growth Media (EBM)
supplemented with the BulletKit containing bovine brain extract,
epidermal growth factor, hydrocortisone, gentamicin, amphotericin
B, fetal bovine serum (FBS) and ascorbic acid according to the
manufacturer’s instructions (Lonza).
Prior to each experiment, the cells at passage 3–5
were starved in an EGM-2 medium (EBM medium with supplement)
containing 0.5% FBS for 24 h to synchronize the cells in G0
phase.
Since Ang II at 103 nM was reported to
increase HUVEC proliferation (27), Ang II (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) at 10–104 nM was initially applied
to HUVECs for 24 h to determine the optimal dose for subsequent
experiments.
Transfection of LOX-1 small-interfering
(si)RNA
The HUVECs were synchronized via starvation for 24 h
prior to each experiment. After they were arrested, the HUVECs were
treated with culture media containing Ang II (104 nM)
for 24 h. LOX-1 siRNA (25, 50 and 100 nM; cat. no. NM_002543) or
control siRNA (100 nM) (both from Sigma-Aldrich; Merck KGaA)
(sense, 5′-GAUCAUACGUGCGAUCAGA-3′ and anti-sense,
5′-UCUGAUCGCACGUAUGAUC-3′) was transfected into the HUVECs for 24 h
using the HiPerFect transfection reagent (Qiagen, Valencia, CA,
USA). For the present time-course experiment, the HUVECs were
transfected with 50 nM LOX-1 siRNA and the cells were incubated at
37°C in 5% CO2 for set times (4, 8, 12, 24 and 48 h).
For the control group, the HUVECs were transfected with the
corresponding control-sequence siRNA (i.e. scrambled siRNA). After
incubation for 24 h, the cells were harvested and the lysates were
subjected to reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis to verify the efficacy of knockdown by
siRNA.
miRNA transfection
The Ang II-treated HUVECs were first transfected
with LOX-1 siRNA (50 nM) for 24 h, and then transfected with
different doses of let-7g (5, 10, 25 and 50 nM) or control miRNA
for 24 h using the HiPerFect transfection reagent (Qiagen). After
incubation for 24 h, the cells were harvested, and the lysates were
subjected to RT-qPCR or western blot analysis to verify the
efficacy of let-7g transfection.
Cell viability assay
Cells grown in a 96-well plate (104
cells/well) were treated with Ang II (10–104 nM) for 24
h. WST-1 reagent (Roche, Mannheim, Germany) diluted 1:10 in growth
medium was added for the last 2 h according to the manufacturer’s
instructions. The amount of viable cells was determined via optical
density measurement using a micro-plate reader at 450 nm, with 600
nm as a reference wavelength.
RNA isolation and RT-qPCR
Total RNA was extracted from the HUVECs using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), and complementary (c)DNA was produced
using 1 μg starting mRNA (Applied Biosystems; Thermo Fisher
Scientific, Inc.) and random hexamers. Reverse transcription was
performed according to the manufacturer’s protocol (Applied
Biosystems; Thermo Fisher Scientific, Inc.): 25°C for 10 min, 37°C
for 120 min and 85°C for 5 min. After the reaction was complete,
samples were cooled down to 4°C. The sequences of the PCR primers
are listed in Table I. The
relative amount of mRNA of interest was normalized to GAPDH. PCR
was performed using the ABI StepOnePlus™ Real-Time PCR system and
SYBR-Green reagent (Applied Biosystems; Thermo Fisher Scientific,
Inc.). PCR was performed in duplicate using 5 μl 2X
SYBR-Green qPCR Master Mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.), 0.2 μl primer sets, 1 μl cDNA and
3.6 μl nucleotide-free H2O in a 10-μl
reaction system. RT-qPCR cycling conditions were as follows: 95°C
for 20 sec, followed by 40 cycles each at 95°C for 1 sec and then
60°C for 20 sec. The relative expression of each mRNA was
calculated as the difference of Cq between the
expression of sample and the housekeeping gene GAPDH (28).
 | Table IPrimers used for quantitative
polymerase chain reaction analysis. |
Table I
Primers used for quantitative
polymerase chain reaction analysis.
| Gene | Primer |
|---|
| GLB1 | F:
5′-TATACTGGCTGGCTAGATCACTG-3′ |
| R:
5′-GGCAAAATTGGTCCCACCTATAA-3′ |
| SIRT1 | F:
5′-AAGTTGACTGTGAAGCTGTACG-3′ |
| R:
5′-TGCTACTGGTCTTACTTTGAGGG-3′ |
| LOX-1 | F:
5′-TTGCCTGGGATTAGTAGTGACC-3′ |
| R:
5′-GCTTGCTCTTGTGTTAGGAGGT-3′ |
| IGF1 | F:
5′-GCTCTTCAGTTCGTGTGTGGA-3′ |
| R:
5′-GCCTCCTTAGATCACAGCTCC-3′ |
| IGF1R | F:
5′-AGGATATTGGGCTTTACAACCTG-3′ |
| R:
5′-GAGGTAACAGAGGTCAGCATTTT-3′ |
| GAPDH | F:
5′-ACAACTTTGGTATCGTGGAAGG-3′ |
| R:
5′-GCCATCACGCCACAGTTTC-3′ |
Western blot analysis
The HUVECs were harvested using
radioimmunoprecipitation assay buffer (Thermo Fisher Scientific,
Inc.) supplemented with a protease inhibitor cocktail (Roche). The
protein concentrations were determined by using the Pierce BCA
Protein assay kit (Thermo Fisher Scientific, Inc.). The proteins
were then transferred onto a polyvinylidene difluoride membrane
(EMD Millipore, Billerica, MA, USA) and blocked with 5% non-fat dry
milk for 2 h at room temperature. The membranes were incubated with
primary antibodies at 4°C for 16 h, including IGF1 (cat. no.
ab9572; 0.1 μg/ml; Abcam, Cambridge, UK) and GAPDH (cat. no.
GTX10011; 1:5,000 dilution; GeneTex, Inc., Irvine, CA, USA).
Following three washes in PBST, the membranes were incubated with
the secondary rabbit immunoglobulin G antibody conjugated to
horseradish peroxidase (cat. no. GTX213110-01; 1:10,000 dilution;
GeneTex, Inc.) at room temperature for 1 h. The enhanced
chemiluminescence non-radioactive detection system was used to
detect the antibody-protein complexes with the LAS-3000 imaging
system (Fujifilm, Tokyo, Japan). Western blot band intensities were
quantitatively measured using ImageJ 1.45s software (National
Institutes of Health, Bethesda, MD, USA).
Senescence-associated β-galactosidase
(SA-β-gal) assay
The senescent cells were assayed with a SA-β-gal
staining kit (cat. no. K802-250; Biovision, Milpitas, CA, USA)
according to the manufacturer’s instructions. The HUVECs were
washed with PBS, fixed with 4% paraformaldehyde for 15 min, then
washed with PBS and incubated overnight at 37°C with a SA-β-gal
staining solution mix. Senescent cells (blue color via SA-β-gal
staining) were observed and counted using a bright-field microscope
at ×100 magnification, and the percentages of SA-β-gal positive
cells were determined. An independent investigator blinded to the
treatment of the samples counted the number of SA-β-gal-positive
cells in five randomly selected high-power fields in each
sample.
Statistical analysis
Statistical analysis was performed using Prism
software, version 5.0 (GraphPad, Inc., La Jolla, CA, USA).
Quantitative data are expressed as the mean ± standard error of the
mean. Student’s t-test was used to compare differences between two
groups. Differences between multiple groups were analyzed using
one-way analysis of variance, followed by Tukey’s post hoc multiple
comparisons test. A two-sided P<0.05 was considered to indicate
a statistically significant difference.
Results
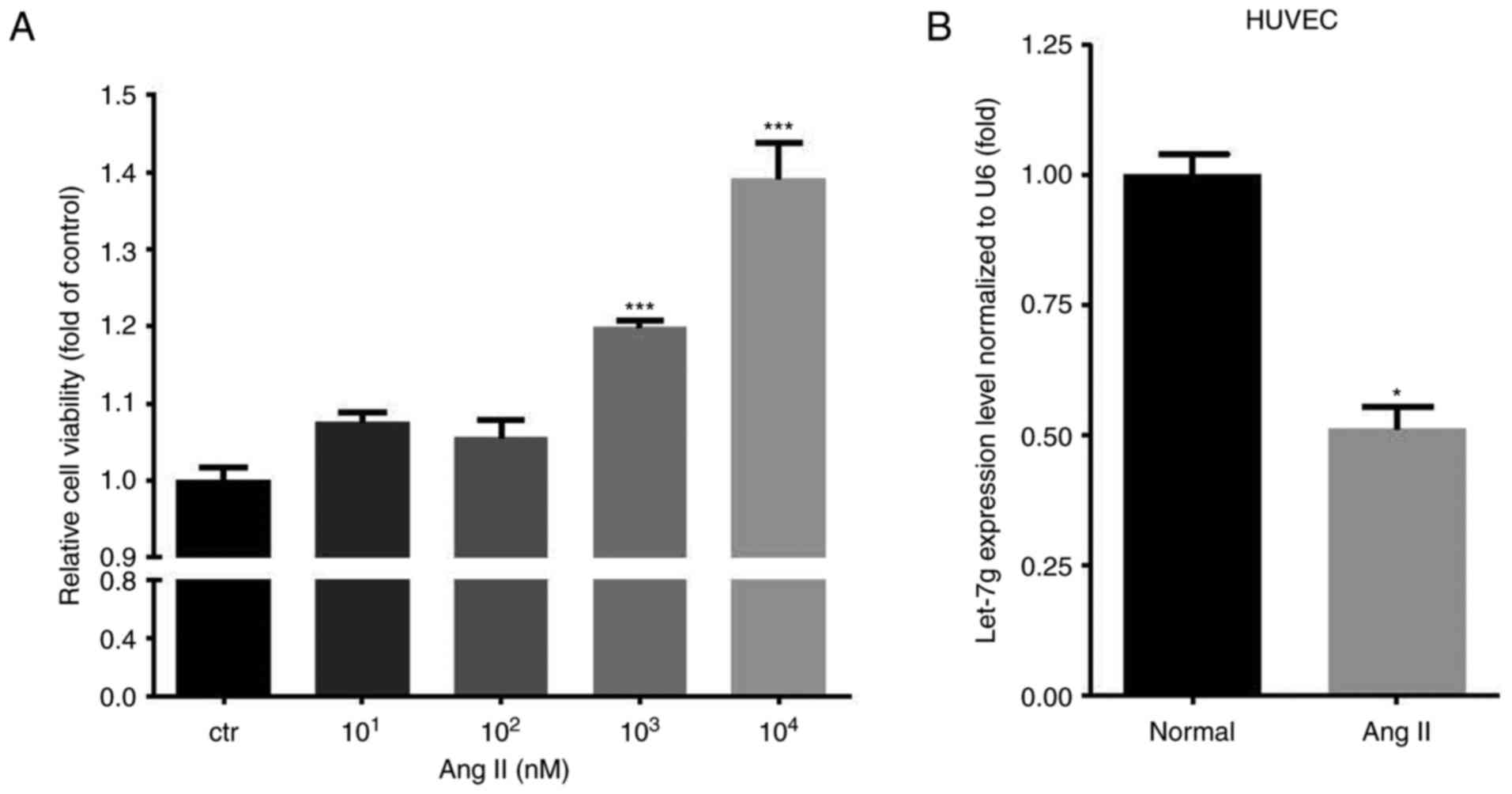
Ang II increases the viability and
decreases the let-7g level of HUVCEs
First, different doses (ranging from 10 to
104 nM) of Ang II were used to test the effect on the
amount of viable HUVECs. The results indicated that Ang II
dose-dependently increased the amount of viable HUVECs, with the
dose of 104 nM having the most significant effect
(Fig. 1A). Therefore, the
concentration of 104 nM was used for subsequent
experiments. The endogenous let-7g levels were measured prior to
and after Ang II treatment. The results indicated that Ang II
treatment reduced let-7g levels in HUVECs by ~50% (P=0.001)
(Fig. 1B).
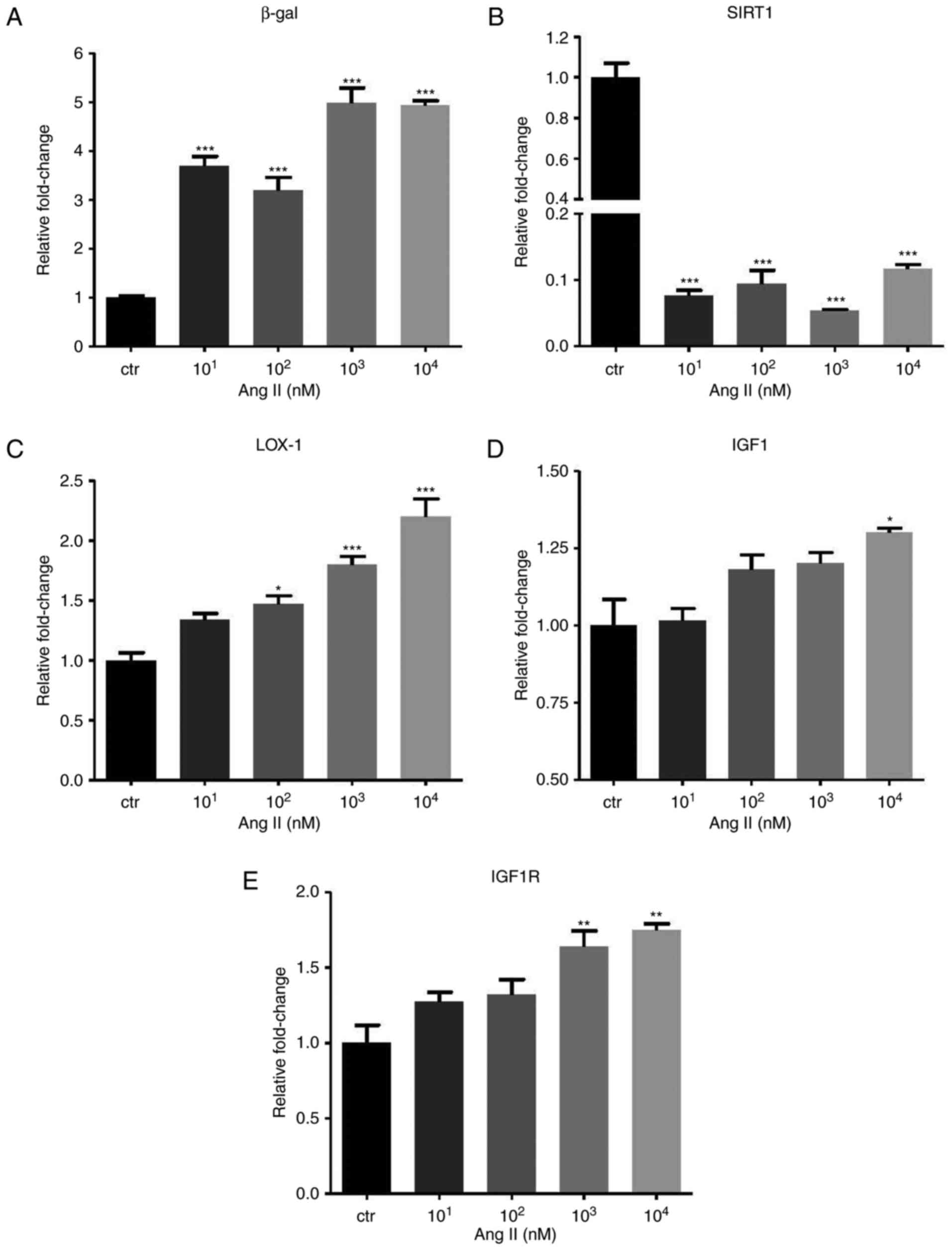
Ang II affects the expression of
β-galactosidase, SIRT1 and the IGF1 pathway
The GLB1 gene encoding β-gal and the SIRT1 gene were
identified to be positively and negatively associated with cell
senescence, respectively. In the Ang II-treated HUVECs, a
significant increase of β-gal (P<0.001) but a decrease of SIRT1
(P<0.001) mRNA expression was identified (Fig. 2A and B). In addition, IGF1
signaling was identified to be associated with the aging process.
In the HUVECs, Ang II promoted LOX-1 (P<0.001), IGF1 (P<0.05)
and IGF1 receptor (IGF1R; P<0.01) expression (Fig. 2C–E).
Knockdown of LOX-1 in Ang II-treated
HUVECs
To explore the effect of let-7g via the
LOX-1-independent mechanism, siRNA was employed to knock down LOX-1
expression. The LOX-1-specific siRNA (doses ranging from 25–100 nM)
suppressed LOX-1 levels (Fig.
3A). The concentrations of 50 and 100 nM siRNA yielded a
similar effect, and therefore, 50 nM siRNA was used in the
subsequent experiments unless a specific dose was otherwise
mentioned. To further determine the ideal incubation time for siRNA
to exert its best inhibitory effect, LOX-1 expression was measured
at 5 time-points (4, 8, 12, 24 and 48 h) after transfection of 50
nM siRNA into the HUVECs. The results indicated that 90% inhibition
occurred at 24 and 48 h (Fig.
3B). Accordingly, all of the subsequent results were obtained
at 24 h after siRNA transfection. When LOX-1 expression was knocked
down, β-gal expression also decreased in a dose-dependent manner in
the Ang II-treated HUVECs (Fig.
3C). Furthermore, SIRT1, IGF1 and IGF1R levels in the Ang
II-treated HUVECs increased along with the elevation of the dose of
LOX-1-siRNA (Fig. 3D–F).
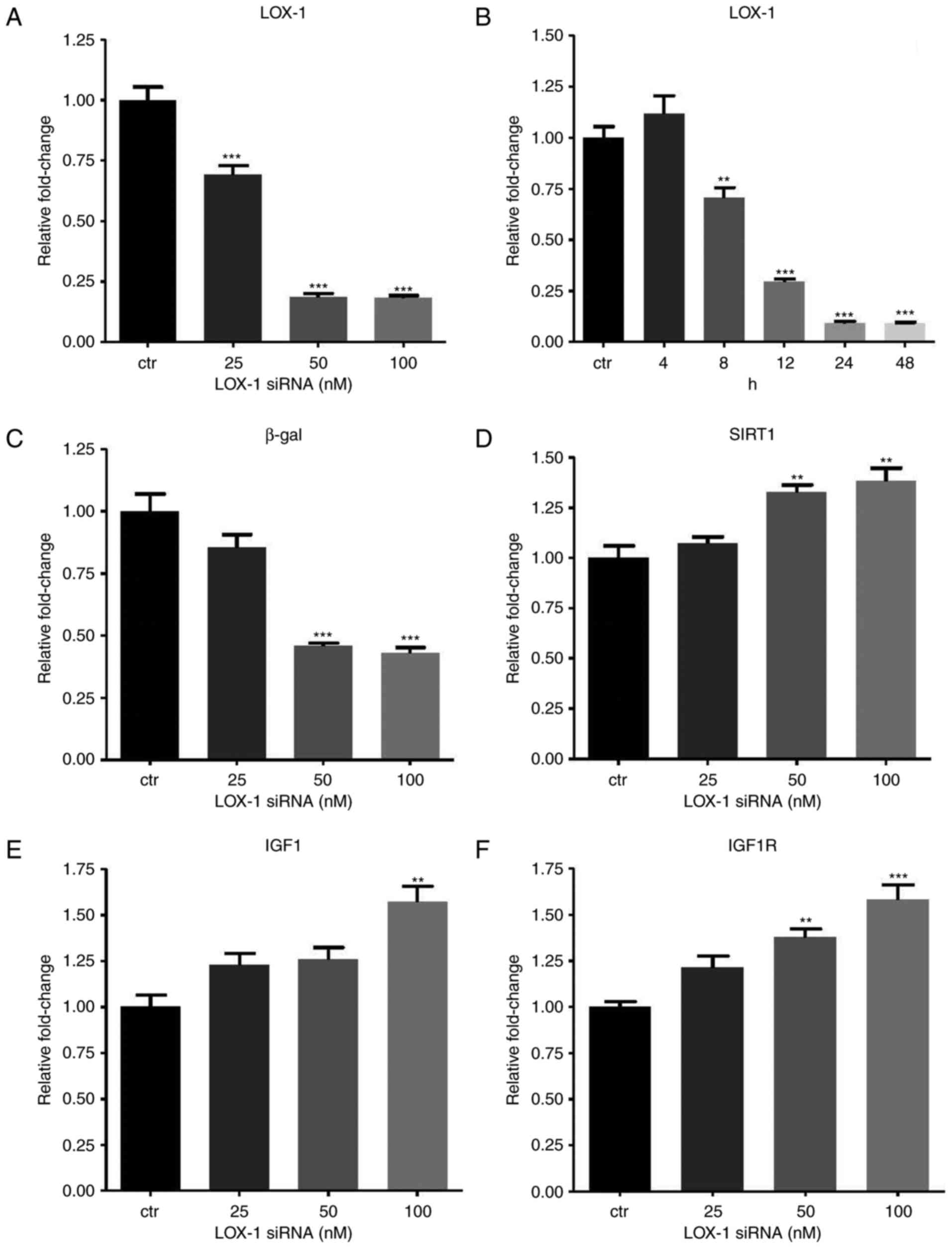 | Figure 3Effects of LOX-1 silencing on
aging-associated gene expression in Ang II-treated human umbilical
vein endothelial cells. The cells were first treated with Ang II
(104 nM) for 24 h, and then transfected with LOX-1 siRNA
for another 24 h. (A) LOX-1 mRNA expression levels were suppressed
after transfection of LOX-1 siRNA (25-100 nM). (B) To determine the
incubation time with the optimal inhibitory effect, LOX-1 mRNA
levels were measured at different time-points after transfection
with LOX-1 siRNA (50 nM). (C–F) The mRNA levels of (C) β-gal, (D)
SIRT1, (E) IGF1 and (F) IGF1R were affected by LOX-1 knockdown.
Values are expressed as the mean ± standard error of the mean.
*P<0.05, **P<0.01 and
***P<0.001 analyzed by one-way analysis of variance
and Tukey’s post hoc test. β-gal, β-galactosidase; SIRT1, sirtuin
1; LOX-1, lectin-like oxidized low-density lipoprotein receptor-1;
IGF1R, insulin-like growth factor 1 receptor; siRNA, small
interfering RNA; Ang II, angiotensin II. |
Let-7g exerts its anti-aging effect via a
LOX-1-independent mechanism
Since a previous study by our group reported that
let-7g has an anti-senescence effect on ECs (12), the present study assessed whether
let-7g also affects aging-associated gene expression and pathways
in a LOX-1-independent manner. In the Ang II-treated and
LOX-1-siRNA-transfected HUVECs, transfection of exogenous let-7g
reduced mRNA expression of IGF1, IGF1R and β-gal (Fig. 4A–C), while it increased SIRT1 mRNA
expression (Fig. 4D). Finally,
HUVECs that had been incubated under different conditions were
subjected to SA-β-gal staining (Fig.
5). The results indicated that the amount of SA-β-gal-positive
HUVECs was markedly increased by Ang II (Fig. 5A and B), while transfection of
LOX-1 siRNA partially abolished the effect of Ang II (Fig. 5C). The SA-β-gal staining result
further supported the notion that let-7g reduces senescence in Ang
II-treated and LOX-1-siRNA-transfected HUVECs (Fig. 5D and E). Previous studies have
demonstrated that let-7g suppresses the protein levels of IGF1R
(29,30) and increases the amount of SIRT1
protein (12). Therefore, the
present study only assessed the inhibitory effect of let-7g on IGF1
protein levels. When cells were treated with Ang II, western blot
analysis of IGF1 provided results that were consistent with those
on the mRNA levels of IGF1 (Fig.
4A).
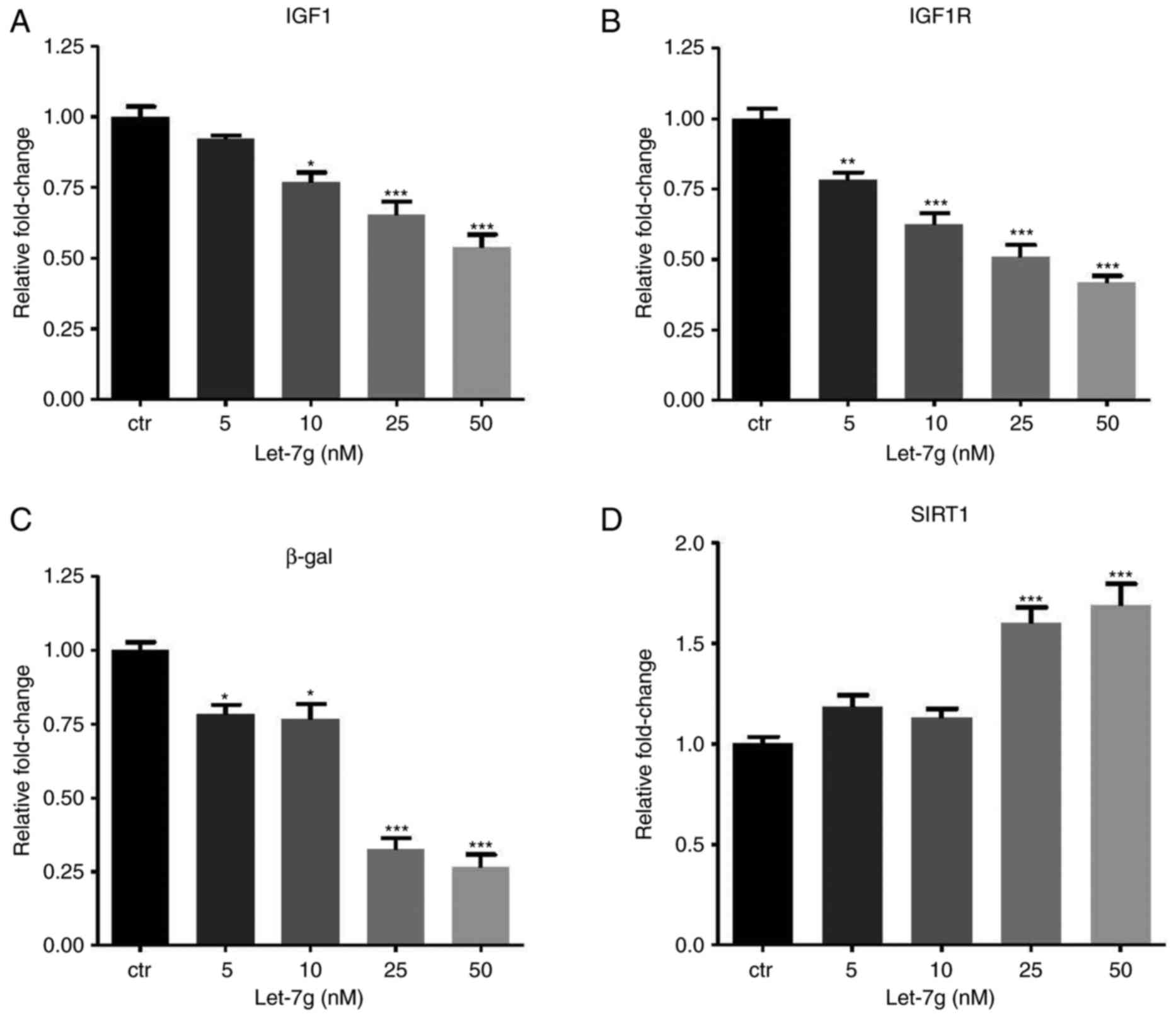 | Figure 4Let-7g affects aging-associated gene
expression via a LOX-1-independent mechanism. Human umbilical vein
endothelial cells were treated with Ang II (104 nM) for
24 h, followed by transfection with 50 nM small interfering RNA
targeting LOX-1 for another 24 h. Subsequently, the cells were
transfected with ctr microRNA or let-7g (5, 10, 25 and 50 nM) or
for 24 h. The mRNA levels of (A) IGF1, (B) IGF1R, (C) β-gal and (D)
SIRT1 were measured by reverse transcription-quantitative
polymerase chain reaction analysis. Values are expressed as the
mean ± standard error of the mean. *P<0.05,
**P<0.01 and ***P<0.001 analyzed by
one-way analysis of variance and Tukey’s post hoc test. β-gal,
β-galactosidase; SIRT1, sirtuin 1; LOX-1, lectin-like oxidized
low-density lipoprotein receptor-1; IGF1R, insulin-like growth
factor 1 receptor; Ang II, angiotensin II; ctr, control. |
 | Figure 5Let-7g suppresses Ang II-induced
HUVEC senescence and IGF protein levels. (A-D) Images of SA-β-gal
staining of HUVECs (scale bar, 100 μm). (A) HUVECs (normal),
(B) HUVECs treated with Ang II (104 nM), (C) HUVECs
treated with Ang II (104 nM) and then transfected with
LOX-1 siRNA (50 nM), (D) HUVECs treated with Ang II (50 nM),
followed by LOX-1 siRNA (50 nM) and then by let-7g (50 nM). Each
individual incubation lasted for 24 h. (E) Percentage of
SA-β-gal-positive cells. The cells from 5 randomly selected
microscopic fields of view were counted. (F) Western blot results
indicated that let-7g dose-dependently inhibited IGF1 protein
expression via a LOX-1-independent pathway. Values are expressed as
the mean ± standard error of the mean (n=3 for each group).
*P<0.05, **P<0.01 and
***P<0.001 analyzed by one-way analysis of variance
and Tukey’s post hoc test. LOX-1, lectin-like oxidized-low-density
lipoprotein receptor-1; IGF1, insulin-like growth factor 1; HUVECs,
human umbilical vein endothelial cells; SA-β-gal,
senescence-associated β-galactosidase; siRNA, small interfering
RNA; Ang II, angiotensin II. |
Discussion
Ang II has been demonstrated to be an important
signaling molecule involved in atherogenic stimuli, and has been
indicated to promote aging and cellular senescence (31). Ang II has been revealed to induce
cardiac fibrosis and downregulation of miRNA-29 expression
(32). miRNA-29 induces cellular
senescence via increasing p53, upregulating β-gal (33) and suppressing SIRT1 expression
(34). In the present study, it
was demonstrated that Ang II attenuated let-7g expression and
induced senescence in ECs. An increase of let-7g significantly
upregulated SIRT1 expression and decreased β-gal expression and
cellular senescence. Therefore, Ang II may increase
senescence-producing miRNA and suppress anti-senescence miRNA. In
Ang II-treated ECs, the expression of aging-associated markers,
including β-gal, IGF1, IGF1R and LOX-1, was increased. Conversely,
Ang II decreased the expression of the anti-aging marker SIRT1. In
addition, LOX-1 was identified to promote the aging process, as
indicated by an increase of β-gal. As the purpose of the present
study was to determine the effect of let-7g on Ang II-induced
senescence, all the cells were first treated with Ang II prior to
further experiments. Therefore, the present study does not have
data on the effect of let-7g on cells without Ang II treatment,
which is a limitation. Since a previous study by our group has
indicated that let-7g directly suppresses LOX-1 (25), the present study examined whether
let-7g exerts a LOX-1-independent anti-aging effect. In Ang
II-treated HUVECs with LOX-1 knockdown, let-7g still reduced the
expression of IGF1, IGF1R and β-gal but increased the SIRT1
expression. Furthermore, using the SA-β-gal staining as an aging
indicator, the anti-aging effect of let-7g via a LOX-1-independent
mechanism was confirmed. Taken together, the present results
suggest that anti-aging effect of let-7g on ECs proceeds via a
LOX-1-dependent and a LOX-1-independent mechanism. Along with the
previous study by our group on the effect of let-7g on LOX-1
(25), its anti-aging effect
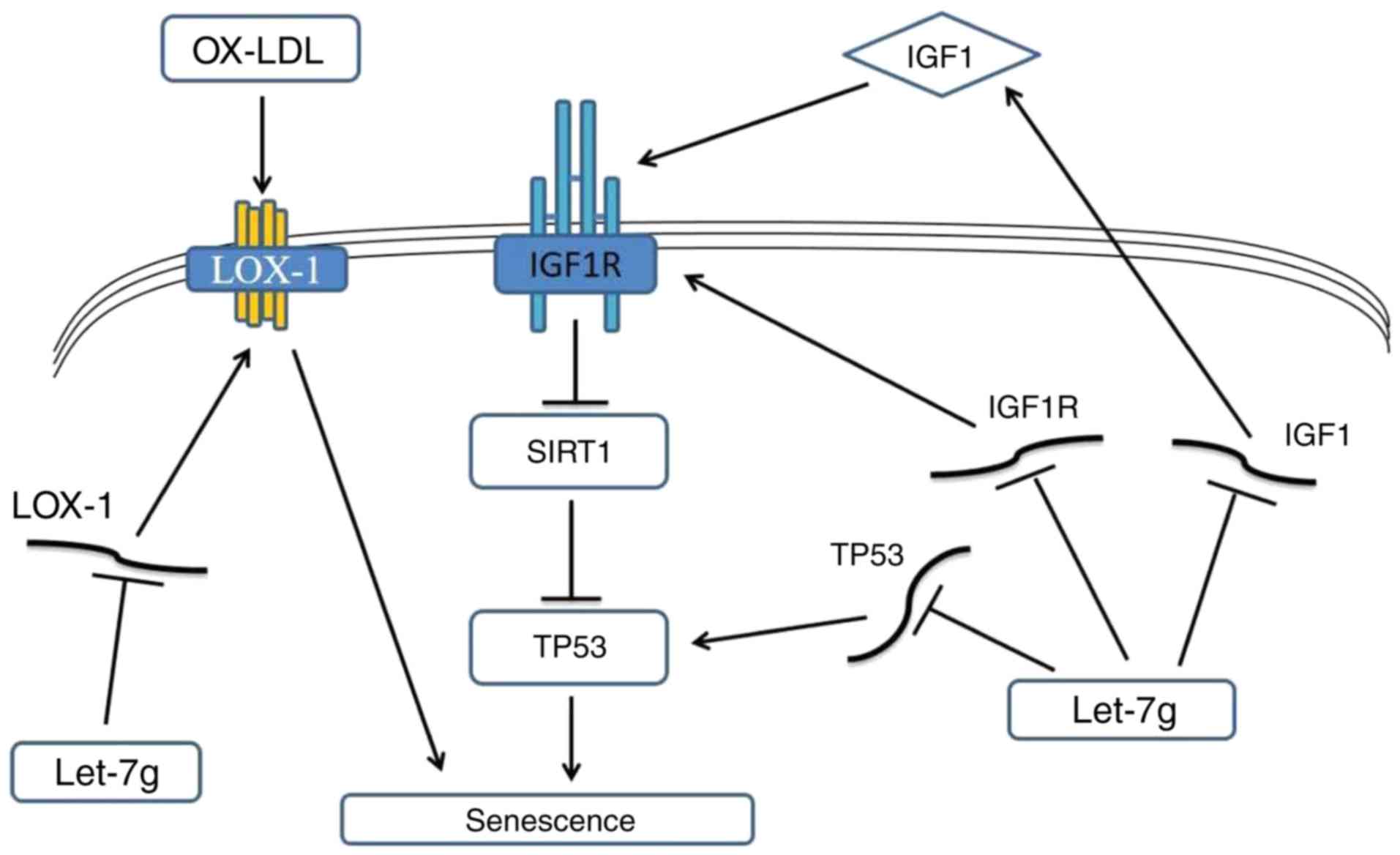
determined in the present study was summarized in a hypothetical
scheme (Fig. 6). The right side
of Fig. 6 illustrates that the
anti-senescence effect of let-7g may be mediated by suppressing the
oxLDL-LOX-1 pathway, while the left side indicates that let-7g may
also suppress IGF1 signaling to exert the anti-senescence
effect.
LOX-1 promotes cell senescence. While LOX-1 is
almost undetectable under physiological conditions, it is
upregulated after exposure to several proinflammatory and
proatherogenic stimuli (35). A
previous study by our group confirmed that let-7g reduces EC
senescence through increasing SIRT-1 protein, but did not assess
whether the anti-aging effect of let-7g is mediated through LOX-1
(12). The present study was
specifically designed to assess the anti-aging effect of let-7g
when LOX-1 was not present. The current findings imply that let-7g
exerts an anti-aging effect regardless of the presence or absence
of LOX-1, which further suggests that let-7g possesses an
anti-aging function via multiple mechanisms.
Ang II increased the levels of LOX-1, IGF1 and IGF1R
in HUVECs or microglial cells (36,37). IGF signaling has a significant
role in ECs by promoting migration, tube formation and production
of the vasodilator nitric oxide. The over-activation of IGF
signaling, however, may suppress SIRT1 expression, contributing to
the aging process. Reduced IGF signaling has also been associated
with increased longevity (38).
In fact, the insulin pathway coordinates growth, development,
metabolic homoeostasis and stress resistance, all of which affect
the lifespan.
The let-7 family has a central role in nutrient
homeostasis and insulin resistance (39,40). Previous studies have indicated
that IGF1R is a target of let-7g in mouse granulosa cells and colon
cancer cells (29,30). The present study further indicated
that let-7g reduces IGF1 expression in the presence of Ang II
stimulation, demonstrating a profound effect of let-7g on the IGF1
signaling pathway.
In conclusion, the present study provided novel
insight into the anti-aging effect of let-7g on ECs, which occurs
regardless of the presence of LOX-1. The present results also
suggest that let-7g has direct effects on multiple targets of the
IGF signaling pathway. Given that healthy and young ECs are crucial
for the cardiovascular system, blood brain barrier, and other
systemic functions, maintenance of sufficient let-7g levels may be
an important strategy to reduce the senescent process.
Acknowledgments
The present study was supported by grants from the
Ministry of Science and Technology (MOST) of Taiwan, R.O.C. (grant
nos. MOST105-2314-B-039-050, MOST103-2314-B-037-026-MY3 and
MOST104-2745-B-037-001), Academia Sinica Taiwan Biobank Stroke
Biosignature Project (grant no. BM10601010036) and a grant from the
National Health Research Institutes of Taiwan, R.O.C. (grant no.
NHRI-EX106-10605PI).
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Campisi J: Aging, cellular senescence, and
cancer. Annu Rev Physiol. 75:685–705. 2013. View Article : Google Scholar
|
|
2
|
Félétou M and Vanhoutte PM: Endothelial
dysfunction: A multifaceted disorder (The Wiggers Award Lecture).
Am J Physiol Heart Circ Physiol. 291:H985–H1002. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xu S, Ogura S, Chen J, Little PJ, Moss J
and Liu P: LOX-1 in atherosclerosis: Biological functions and
pharmacological modifiers. Cell Mol Life Sci. 70:2859–2872. 2013.
View Article : Google Scholar
|
|
4
|
Wang X, Khaidakov M, Ding Z, Dai Y,
Mercanti F and Mehta JL: LOX-1 in the maintenance of cytoskeleton
and proliferation in senescent cardiac fibroblasts. J Mol Cell
Cardiol. 60:184–190. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mehta PK and Griendling KK: Angiotensin II
cell signaling: Physiological and pathological effects in the
cardiovascular system. Am J Physiol Cell Physiol. 292:C82–C97.
2007. View Article : Google Scholar
|
|
6
|
Kambayashi Y, Bardhan S, Takahashi K,
Tsuzuki S, Inui H, Hamakubo T and Inagami T: Molecular cloning of a
novel angiotensin II receptor isoform involved in phosphotyrosine
phosphatase inhibition. J Biol Chem. 268:24543–24546.
1993.PubMed/NCBI
|
|
7
|
Sasaki K, Yamano Y, Bardhan S, Iwai N,
Murray JJ, Hasegawa M, Matsuda Y and Inagami T: Cloning and
expression of a complementary DNA encoding a bovine adrenal
angiotensin II type-1 receptor. Nature. 351:230–233. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Min LJ, Mogi M, Iwai M and Horiuchi M:
Signaling mechanisms of angiotensin II in regulating vascular
senescence. Ageing Res Rev. 8:113–121. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kunieda T, Minamino T, Nishi J, Tateno K,
Oyama T, Katsuno T, Miyauchi H, Orimo M, Okada S, Takamura M, et
al: Angiotensin II induces premature senescence of vascular smooth
muscle cells and accelerates the development of atherosclerosis via
a p21-dependent pathway. Circulation. 114:953–960. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li DY, Zhang YC, Philips MI, Sawamura T
and Mehta JL: Upregulation of endothelial receptor for oxidized
low-density lipoprotein (LOX-1) in cultured human coronary artery
endothelial cells by angiotensin II type 1 receptor activation.
Circ Res. 84:1043–1049. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feige JN and Auwerx J: Transcriptional
targets of sirtuins in the coordination of mammalian physiology.
Curr Opin Cell Biol. 20:303–309. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liao YC, Wang YS, Guo YC, Lin WL, Chang MH
and Juo SH: Let-7g improves multiple endothelial functions through
targeting transforming growth factor-beta and SIRT-1 signaling. J
Am Coll Cardiol. 63:1685–1694. 2014. View Article : Google Scholar
|
|
13
|
Gorenne I, Kumar S, Gray K, Figg N, Yu H,
Mercer J and Bennett M: Vascular smooth muscle cell sirtuin 1
protects against DNA damage and inhibits atherosclerosis.
Circulation. 127:386–396. 2013. View Article : Google Scholar
|
|
14
|
Stein S, Lohmann C, Schäfer N, Hofmann J,
Rohrer L, Besler C, Rothgiesser KM, Becher B, Hottiger MO, Borén J,
et al: SIRT1 decreases Lox-1-mediated foam cell formation in
atherogenesis. Eur Heart J. 31:2301–2309. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang QJ, Wang Z, Chen HZ, Zhou S, Zheng
W, Liu G, Wei YS, Cai H, Liu DP and Liang CC: Endothelium-specific
overexpression of class III deacetylase SIRT1 decreases
atherosclerosis in apoli-poprotein E-deficient mice. Cardiovasc
Res. 80:191–199. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kenyon C: A conserved regulatory system
for aging. Cell. 105:165–168. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Longo VD and Finch CE: Evolutionary
medicine: From dwarf model systems to healthy centenarians?
Science. 299:1342–1346. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lai CH, Ho TJ, Kuo WW, Day CH, Pai PY,
Chung LC, Liao PH, Lin FH, Wu ET and Huang CY: Exercise training
enhanced SIRT1 longevity signaling replaces the IGF1 survival
pathway to attenuate aging-induced rat heart apoptosis. Age
(Dordr). 36:97062014. View Article : Google Scholar
|
|
19
|
Brunet A, Sweeney LB, Sturgill JF, Chua
KF, Greer PL, Lin Y, Tran H, Ross SE, Mostoslavsky R, Cohen HY, et
al: Stress-dependent regulation of FOXO transcription factors by
the SIRT1 deacetylase. Science. 303:2011–2015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cohen HY, Miller C, Bitterman KJ, Wall NR,
Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R and Sinclair
DA: Calorie restriction promotes mammalian cell survival by
inducing the SIRT1 deacetylase. Science. 305:390–392. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tissenbaum HA and Guarente L: Increased
dosage of a sir-2 gene extends lifespan in Caenorhabditis elegans.
Nature. 410:227–230. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bao MH, Feng X, Zhang YW, Lou XY, Cheng Y
and Zhou HH: Let-7 in cardiovascular diseases, heart development
and cardiovascular differentiation from stem cells. Int J Mol Sci.
14:23086–23102. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiu SC, Chung HY, Cho DY, Chan TM, Liu
MC, Huang HM, Li TY, Lin JY, Chou PC, Fu RH, et al: Therapeutic
potential of microRNA let-7: Tumor suppression or impeding normal
stemness. Cell Transplant. 23:459–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen KC, Hsieh IC, His E, Wang YS, Dai CY,
Chou WW and Juo SH: Negative feedback regulation between microRNA
let-7g and the oxLDL receptor LOX-1. J Cell Sci. 124:4115–4124.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hsu PY, His E, Wang TM, Lin RT, Liao YC
and Juo SH: MicroRNA let-7g possesses a therapeutic potential for
peripheral artery disease. J Cell Mol Med. 21:519–529. 2017.
View Article : Google Scholar
|
|
27
|
Yang LL, Li DY, Zhang YB, Zhu MY, Chen D
and Xu TD: Salvianolic acid A inhibits angiotensin II-induced
proliferation of human umbilical vein endothelial cells by
attenuating the production of ROS. Acta Pharmacol Sin. 33:41–48.
2012. View Article : Google Scholar
|
|
28
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Choo KB, Soon YL, Nguyen PN, Hiew MS and
Huang CJ: MicroRNA-5p and -3p co-expression and cross-targeting in
colon cancer cells. J Biomed Sci. 21:952014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou J, Yao W, Liu K, Wen Q, Wu W, Liu H
and Li Q: MicroRNA let-7g regulates mouse granulosa cell autophagy
by targeting insulin-like growth factor 1 receptor. Int J Biochem
Cell Biol. 78:130–140. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shan H, Bai X and Chen X: Angiotensin II
induces endothelial cell senescence via the activation of
mitogen-activated protein kinases. Cell Biochem Funct. 26:459–466.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Huang XR, Wei LH, Chung AC, Yu CM
and Lan HY: miR-29b as a therapeutic agent for angiotensin
II-induced cardiac fibrosis by targeting TGF-β/Smad3 signaling. Mol
Ther. 22:974–985. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hu Z, Klein JD, Mitch WE, Zhang L,
Martinez I and Wang XH: MicroRNA-29 induces cellular senescence in
aging muscle through multiple signaling pathways. Aging (Albany
NY). 6:160–175. 2014. View Article : Google Scholar :
|
|
34
|
Xu Z, Zhang L, Fei X, Yi X, Li W and Wang
Q: The miR-29b-Sirt1 axis regulates self-renewal of mouse embryonic
stem cells in response to reactive oxygen species. Cell Signal.
26:1500–1505. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pirillo A, Norata GD and Catapano AL:
LOX-1, OxLDL, and atherosclerosis. Mediators Inflamm.
2013:1527862013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Morawietz H, Rueckschloss U, Niemann B,
Duerrschmidt N, Galle J, Hakim K, Zerkowski HR, Sawamura T and
Holtz J: Angiotensin II induces LOX-1, the human endothelial
receptor for oxidized low-density lipoprotein. Circulation.
100:899–902. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rodriguez-Perez AI, Borrajo A, Diaz-Ruiz
C, Garrido-Gil P and Labandeira-Garcia JL: Crosstalk between
insulin-like growth factor-1 and angiotensin-II in dopaminergic
neurons and glial cells: Role in neuroinflammation and aging.
Oncotarget. 7:30049–30067. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Suh Y, Atzmon G, Cho MO, Hwang D, Liu B,
Leahy DJ, Barzilai N and Cohen P: Functionally significant
insulin-like growth factor I receptor mutations in centenarians.
Proc Natl Acad Sci USA. 105:3438–3442. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dubinsky AN, Dastidar SG, Hsu CL, Zahra R,
Djakovic SN, Duarte S, Esau CC, Spencer B, Ashe TD, Fischer KM, et
al: Let-7 coordinately suppresses components of the amino acid
sensing pathway to repress mTORC1 and induce autophagy. Cell Metab.
20:626–638. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhu H, Shyh-Chang N, Segrè AV, Shinoda G,
Shah SP, Einhorn WS, Takeuchi A, Engreitz JM, Hagan JP, Kharas MG,
et al: The Lin28/let-7 axis regulates glucose metabolism. Cell.
147:81–94. 2011. View Article : Google Scholar : PubMed/NCBI
|