Introduction
Cardiac ischemia/reperfusion (I/R) injury is
associated with various etiological factors (1-3),
including primary heart conditions, and neuropathic, vascular or
systemic disorders. However, the pathological origin of I/R-induced
cardiomyocyte death remains poorly understood. The neuropathic
accumulation associated with myocardial I/R is thought to directly
or indirectly activate sensory or sympathetic fibers that innervate
the heart. However, as the underlying heart-specific neuronal
pathway and mechanism are unknown, neurogenic therapeutic
interventions often only have limited success.
It is well established that the autonomic nervous
system serves a crucial physiological role in regulating cardiac
function (4-9). Previous studies have demonstrated
that some myocardial ischemic events are triggered by the autonomic
nervous system, and a sympathetic-parasympathetic imbalance may
lead to the pathophysiological development of myocardial ischemia
(10,11). Our previous study revealed that an
injection of retrograde tracer pseudorabies virus (PRV)-614 into
the left ventricle wall in mice resulted in the retrograde
infection of neurons in the intermediolateral nucleus of the spinal
cord and the rostral ventrolateral medulla of the brainstem via the
sympathetic pathway (12). The
spinal cord has been implicated in the pathogenesis of cardiac
injury caused by I/R (4,13-16). Furthermore, there is convincing
evidence that the heart receives dense innervation from sensory
afferent fibers, which peripherally release a variety of
vasodilator neuropeptides, such as calcitonin gene-related peptide
(CGRP) (17) and substance P
(SP), in response to local stimuli (18). Despite extensive research in this
area, the mechanisms underlying cardiac I/R injury are largely
unknown. Therefore, there is an urgent requirement for more
information and a genomic approach may prove helpful.
High-throughput RNA sequencing (RNA-seq) is a
powerful tool that has been used to identify novel protein-coding
and non-coding RNA transcripts involved in the regulation of gene
expression (19-24). Recent research has focused on the
cardiac long non-coding RNAs (lncRNAs) implicated in cardiac I/R
injury (25-29); however, few studies have explored
the important role of the spinal cord during focal cardiac I/R.
Therefore, the present study was designed to identify the
expression patterns of differentially expressed lncRNAs and mRNAs
in the spinal cord under normal and cardiac I/R conditions, with
the aim to gain a better understanding of the genetic mechanisms
underlying the pathogenesis of cardiac I/R injury. The present
study also determined the expression levels of various genes in the
spinal cord at different time-points during cardiac I/R injury.
Materials and methods
Animals
A total of 24 male Sprague Dawley (SD) rats aged
8-10 weeks (200-240 g; specific pathogen-free grade; no.
42000600010250) were supplied by the Experimental Animal Research
Center of Hubei Province (Hubei, China). The present experiment
protocol was approved by the Institutional Ethical Committee of
Tongji Hospital, Tongji Medical College, Huazhong University of
Science and Technology (Hubei, China; no. TJ-A20150804). All
animals were humanely treated according to the National Institutes
of Health Guide for the Care and Use of Laboratory Animals (revised
2011) and the Guide for the Care and Use of Laboratory Animals
(National Academic Press, USA; revised 2011) (30). Animals were housed in compliance
with the Specific Pathogen-Free Animal Criteria, and maintained at
a standard temperature of 21-23°C and 65±5% humidity under a 12-h
light/dark cycle condition, with ad libitum access to
artificial feed (food and water).
Myocardial I/R injury model
Surgical procedures to establish the myocardial I/R
injury model were performed according to previously described
methods (31-33).
Experimental protocol
Experiment A
A total of 6 SD rats were randomly allocated into 2
groups. The model group (n=3) received 2 h reperfusion following
30-min occlusion of the left anterior descending coronary artery by
pulling the reversible trap (I/R group). The control group (n=3)
received the same surgical procedure, without any occlusion of the
coronary artery and reperfusion (sham group). T1-T4 spinal cord
samples were collected for RNA-seq and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis.
Experiment B
A total of 18 SD rats were randomly assigned into 3
groups: i) A control group (n=6); ii) a 0.5 h group (0.5 h
reperfusion following 30 min ischemia; n=6); and iii) a 2 h group
(2 h reperfusion following 30 min ischemia; n=6). T1-T4 spinal cord
segments were obtained for RT-qPCR analysis.
Tissue preparation and microarray gene
expression analysis
The rats were sacrificed following the completion of
the aforementioned experiments. Briefly, upon completion of the
experiments, the rats were anesthetized by intraperitoneal
injection with 100 mg/kg body weight ketamine and 10 mg/kg body
weight xylazine (34). Then the
rats were quickly decapitated to limit animal suffering and
minimize the effects on the experimental results, and the T1-T4
spinal cord segments were immediately removed and frozen with
liquid nitrogen for 1 min, then stored at −80°C until required.
Total RNA from each animal was quantified using a mirVana miRNA
Isolation kit (Ambion; Thermo Fisher Scientific, Inc., Waltham, MA,
USA), and RNA integrity was assessed according to the
manufacturer's protocol, which included standard denaturing agarose
gel electrophoresis (35-37). For RNA-seq, the micro-array work
was performed by CapitalBio Technology Co. Ltd. (Beijing, China),
whereby 6 tissue samples (3 model group samples and 3 control group
samples) were used for mRNA and lncRNA microarray analysis
(38).
The present study used an Agilent Array platform for
microarray analysis. Tissue preparation and microarray
hybridization were performed based on the manufacturer's standard
protocols (Agilent Technologies, Inc., Santa Clara, CA, USA) with
minor modifications. Briefly, the mRNA was purified from the total
RNA once the ribosomal RNA was removed using an mRNA-ONLY
Eukaryotic mRNA Isolation kit (Epicentre; Illumina, Inc., San
Diego, CA, USA). Each sample was then amplified and transcribed
into fluorescent complementary RNA (cRNA) along the entire length
of the transcripts using a random priming method (39). The labeled cRNAs were hybridized
onto the mouse lncRNA Array v2.0 (8×60 K; Arraystar, Inc.,
Rockville, MD, USA). Then the arrays were scanned using a G2505C
Scanner (Agilent Technologies, Inc.).
Bioinformatics analysis
Gene Ontology (GO) annotations were employed to
investigate the differentially expressed mRNAs and lncRNAs in the
T1-T4 spinal cord segments, according to the GO database
(www.geneontology.org/). For the
functions of genes and their products, the GO database describes 3
biological functional groups: Biological process, Cellular
compartment, and Molecular function. The present study conducted GO
functional enrichment analysis on the differentially expressed
mRNAs involved in protein-protein interaction (PPI) networks. In
addition, the key regulatory pathways in the spinal cord that
respond to I/R-induced cardiac injury were also investigated using
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis
(www.genome.jp/kegg) and STRING database
(string-db.org).
RT-qPCR analysis
The present study extracted total RNA from the upper
thoracic spinal cord segments (T1-T4) (40) using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to our
previous research (35-37). The primers for RT-qPCR were
designed based on the lncRNA sequences (Table I), and were synthesized and
purified at Invitrogen (Thermo Fisher Scientific, Inc.). The RT
reactions were performed using a iScript™ cDNA Synthesis kit
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). RT-qPCR was
performed using a ABI StepOnePlusä Multicolor system with the
SsoAdvanced™ Universal SYBR®-Green Supermix (Bio-Rad
Laboratories, Hercules, Inc.). The PCR thermocy-cling conditions
were as follows: Initial denaturation at 95°C for 1 min followed by
40 cycles of 95°C for 15 sec, 60°C for 15 sec and 72°C for 45 sec.
Compared with the averages for the housekeeping gene (GAPDH), the
data were quantified using the 2−∆∆Cq method for
relative fold-change, as described previously (41).
 | Table IPrimer sequences for reverse
transcriptase-quantitative polymerase chain reaction. |
Table I
Primer sequences for reverse
transcriptase-quantitative polymerase chain reaction.
| A, Upregulated |
|---|
|
|---|
| Gene | Length (bp) | Forward
(5′-3′) | Reverse
(5′-3′) |
|---|
| NONRATT025386 | 86 |
GGGTCTGGGGTGGGCTAA |
GGAGGTTTCTGAGTGGGATGTG |
| NONRATT016113 | 96 |
CCACAAGCGTCTCGGGATT |
AGCGAAAACAGTCATTTTAACCAA |
| NONRATT018298 | 166 |
GACAGTCAACGGAACCAAACTAA |
CGTGAACAAAAGCAAGCAAAC |
| NONRATT018300 | 178 |
GCCAACAACCAGTAAGAACCAC |
CCATACCTTTGCTACTTTGGAGA |
| NONRATT020994 | 150 |
GAACGCCACCCCACCAT |
CCTTGAAGTCTGAGGCAGGAA |
|
| B,
Downregulated | | | |
|
| Gene | Length (bp) | Forward
(5′-3′) | (5′-3′) |
| XR_590210.1 | 128 |
TTTCAGCCCATCAATGGTTTC |
TCCTCAGGAGTGCCCTTTCT |
| NONRATT002188 | 105 |
TTCCTACATACTGAGCAACGACC |
CCTACCTGTAGCTGCCACTCC |
| XR_589980.1 | 115 |
GGATGCCCACTCAAGGGTC |
GATGATAAATGCTTGCCCACC |
| XR_598701.1 | 145 |
AACAATGGGGACGGTAGTGC |
GAAATGAACCTGGGAGAAACG |
| XR_590197.1 | 92 |
ACTTCCCTGGATTCTGCTCTG |
GGGTCCCCTAACACTATTGCTT |
|
| GAPDH | 68 |
CGCTAACATCAAATGGGGTG |
TTGCTGACAATCTTGAGGGAG |
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. The data and statistical graphs were analyzed using the
GraphPad Prism v6.02 package (GraphPad Software, Inc., La Jolla,
CA, USA). Between-group counts were compared using a Student's
t-test (Mann-Whitney U), and the data from three groups were
analyzed by one-way analysis of variance followed by Dunn's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Characteristics of ischemic myocardial
tissues
With regard to the animals in the model group, there
were evident cyanotic changes in the myocardium of the occluded
area 30 min post-ischemia, and a reactive hyperemic response after
refilling of the left anterior descending coronary artery. With
regard to the samples in the model group, examination under a
dissecting microscope revealed discoloration of the occluded distal
myocardium at the early stage of reperfusion in the infarct region
(data not shown).
Expression profiling of lncRNAs in the
spinal cord 2 h post-cardiac I/R
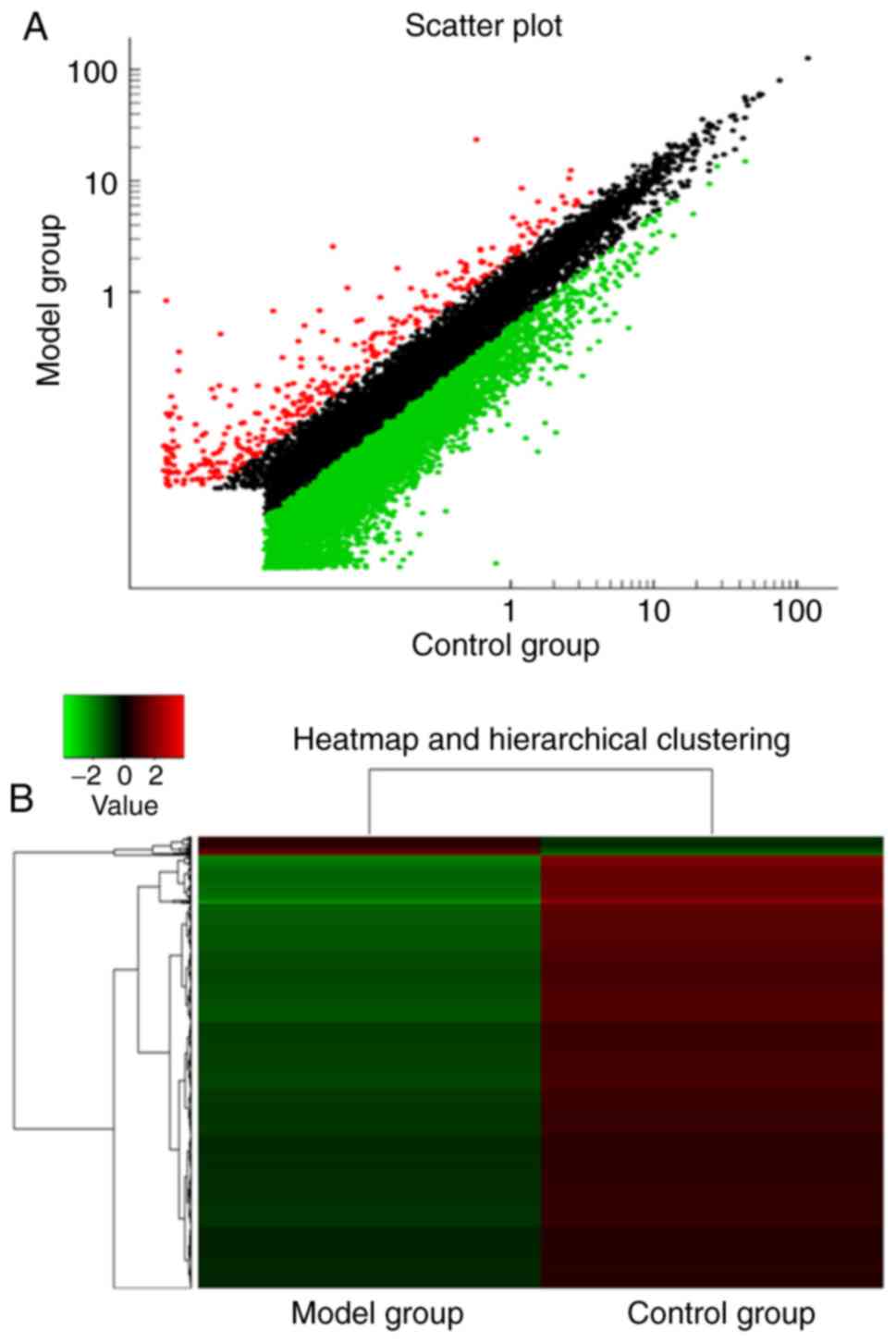
To select possible targets of lncRNAs in the model
and control groups, the present study detected up to 16,987 coding
transcripts in the T1-T4 spinal cords 2 h post-reperfusion. A total
of 3,621 upregulated and 13,366 downregulated lncRNAs were
identified in the spinal cords. On average, 234 lncRNAs were
upregulated in the spinal cords of the model group, compared with
those in the control group, whereas an average of 7,746 lncRNAs
were downregulated (>2.0-fold-change; P<0.05). The
distributions of the log2 ratios of the lncRNAs in the model and
control samples were nearly identical; Fig. 1 presents the heatmaps of the
expression ratios (log2 scale) of the lncRNAs in the spinal cords.
The top 20 up- and downregulated mRNAs are listed in Tables II and III.
| Table IITop 20 upregulated lncRNAs in the
spinal cord at 2 h post-reperfusion. |
Table II
Top 20 upregulated lncRNAs in the
spinal cord at 2 h post-reperfusion.
| LncRNAs (sequence
name) | Fold-change
(R/N) | RNA length | Chromosome
log2 |
|---|
|
gi|672017878|ref|XR_345533.2| | 83.01321 | 1,727 | Chr3 |
| NONRATT025386 | 23.99574 | 563 | Chr6 |
| NONRATT024318 | 23.93667 | 473 | Chr6 |
|
gi|672024701|ref|XR_599241.1| | 13.07902 | 728 | Chr10 |
| NONRATT025509 | 10.08656 | 553 | Chr7 |
|
gi|672017768|ref|XR_600487.1| | 9.916187 | 634 | Chr3 |
| NONRATT025839 | 9.860808 | 508 | Chr7 |
| NONRATT023339 | 9.858076 | 706 | Chr5 |
| NONRATT000120 | 9.325455 | 550 | Chr1 |
| NONRATT002260 | 8.604115 | 692 | Chr1 |
|
gi|672055933|ref|XR_592974.1| | 8.306979 | 8,102 | Chr6 |
| uc.126 | 7.209348 | 271 | - |
|
gi|672086728|ref|XR_597427.1| | 6.502431 | 4,903 | Chr20 |
| NONRATT008414 | 6.458688 | 518 | Chr13 |
| NONRATT026470 | 6.452916 | 655 | Chr7 |
| NONRATT015818 | 6.317123 | 255 | Chr2 |
|
gi|672080453|ref|XR_596511.1| | 6.286473 | 1,683 | Chr16 |
|
gi|672027556|ref|XR_340041.2| | 6.236808 | 633 | Chr13 |
|
gi|672030740|ref|XR_598338.1| | 6.169331 | 1,360 | Chr18 |
| NONRATT016237 | 6.087859 | 817 | Chr2 |
| Table IIITop 20 downregulated lncRNAs in the
spinal cord at 2 h post-reperfusion. |
Table III
Top 20 downregulated lncRNAs in the
spinal cord at 2 h post-reperfusion.
| LncRNAs (sequence
name) | Fold-change
(R/N) | RNA length | Chromosome
log2 |
|---|
| NR_130708.1 | −27.7049 | 1,379 | Chr3 |
| NONRATT028627 | −25.2873 | 410 | Chr8 |
| NONRATT021959 | −22.8492 | 709 | Chr4 |
|
gi|672034655|ref|XR_590005.1| | −21.8020 | 1,290 | Chr1 |
| NONRATT023191 | −19.8345 | 1,977 | Chr5 |
| NONRATT025830 | −19.4635 | 525 | Chr7 |
| NONRATT027253 | −19.4441 | 1,233 | Chr7 |
| NONRATT023189 | −19.2427 | 2,086 | Chr5 |
| NONRATT014248 | −18.4873 | 1,049 | Chr19 |
| NONRATT008322 | −17.5179 | 451 | Chr13 |
| NONRATT014862 | −16.4839 | 619 | Chr2 |
| NONRATT016808 | −16.4007 | 518 | Chr2 |
| NONRATT017256 | −16.2487 | 786 | Chr20 |
| NONRATT011649 | −15.8013 | 566 | Chr16 |
|
gi|672021532|ref|XR_347699.2| | −15.494 | 537 | Chr7 |
| NONRATT024978 | −14.3807 | 1,486 | Chr6 |
| NONRATT012913 | −13.9501 | 338 | Chr17 |
| NONRATT004220 | −13.7216 | 2,171 | Chr10 |
| NONRATT018550 | −13.4908 | 365 | Chr3 |
| NONRATT008489 | −13.4256 | 1,076 | Chr13 |
Expression profiling of mRNAs in the
spinal cord 2 h post-cardiac I/R
To explore the potential role of mRNAs in the T1-T4
spinal cords 2 h post-cardiac I/R, the present study determined the
expression profiles of mRNAs by high-throughput RNA-seq. Of the
26,466 mRNAs that were quantified using reads per kilobase per
million mapped reads (RPKM) values, 3,428 mRNAs were deregulated by
2-fold following I/R-induced cardiac injury, of which 767 mRNAs
were upregulated and 2,661 mRNAs were downregulated; Fig. 2 presents the heatmaps of the
expression ratios (log2 scale) of the mRNAs in the spinal cords.
The top 20 up- and downregulated mRNAs are listed in Tables IV and V.
| Table IVTop 20 upregulated mRNAs in the
spinal cord at 2 h post-reperfusion. |
Table IV
Top 20 upregulated mRNAs in the
spinal cord at 2 h post-reperfusion.
| mRNAs (sequence
name) | Fold-change
(R/N) | GENE_SYMBOL | GENE_NAME |
|---|
| A_64_P004112 | 25.97118 | - | - |
| A_64_P151353 | 25.46518 | Acsm5 | Acyl-CoA synthetase
medium-chain family member 5 |
| A_64_P181171 | 21.23528 | - | - |
| A_64_P149280 | 21.07919 | Vegfb | Vascular
endothelial growth factor B |
| A_64_P273771 | 15.23514 | - | - |
| A_64_P260129 | 14.28584 | Gzmc | Granzyme C |
| A_44_P122912 | 13.21478 | Ces1c | Carboxylesterase
1C |
| A_44_P553341 | 13.01391 | Lrfn5 | Leucine rich repeat
and fibronectin type III domain containing 5 |
| A_64_P094055 | 10.5432 | - | - |
| A_44_P401110 | 10.10015 | Prss40 | 'Protease, serine,
40' |
| A_64_P147769 | 8.814655 | Gucy1b2 | 'Guanylate cyclase
1, soluble, β2' |
| A_64_P082082 | 8.515041 | - | - |
| A_64_P045902 | 8.093776 | - | - |
| A_64_P118367 | 8.069224 | Lrrd1 | Leucine-rich
repeats and death domain containing 1 |
| A_64_P156605 | 7.85615 | - | - |
| A_64_P135295 | 7.737585 | Cd300c | CD300c
molecule |
| A_64_P042127 | 7.403613 | Akr1c3 | 'Aldo-keto
reductase family 1, member C3' |
| A_44_P445575 | 7.245778 | Irx2 | Iroquois homeobox
2 |
| A_64_P162476 | 7.220482 | Art1 |
ADP-ribosyltransferase 1 |
| A_64_P186630 | 7.156018 | Vom2r60 | 'Vomeronasal 2
receptor, 60' |
| Table VTop 20 downregulated mRNAs in the
spinal cord at 2 h post-reperfusion. |
Table V
Top 20 downregulated mRNAs in the
spinal cord at 2 h post-reperfusion.
| mRNAs (sequence
name) | Fold-change
(R/N) | GENE_SYMBOL | GENE_NAME |
|---|
| A_64_P141763 | −38.1959 | Muc16 | 'Mucin 16, cell
surface associated' |
| A_42_P473398 | −33.0016 | Cxcl1 | Chemokine (C-X-C
motif) ligand 1 |
| A_44_P550907 | −29.4102 | RGD1306750 | LOC362451 |
| A_44_P702019 | −23.3124 | Rfx4 | 'Regulatory factor
X, 4 (influences HLA class II expression)' |
| A_64_P021855 | −22.6481 | LOC499643 | Similar to
hypothetical protein FLJ25371 |
| A_44_P367541 | −21.9947 | Olr671 | Olfactory receptor
671 |
| A_64_P036090 | −20.4897 | - | - |
| A_44_P371339 | −20.0377 | IL6 | Interleukin 6 |
| A_64_P141762 | −19.8627 | - | - |
| A_64_P018554 | −19.1504 | - | - |
| A_64_P101228 | −16.1995 | Tas2r120 | 'Taste receptor,
type 2, member 120' |
| A_44_P563447 | −15.8884 | Wnt10a | 'Wingless-type MMTV
integration site family, member 10A' |
| A_44_P698466 | −15.406 | Lrrtm2 | Leucine rich repeat
transmembrane neuronal 2 |
| A_44_P461456 | −15.3963 | Prl4a1 | 'Prolactin family
4, subfamily a, member 1' |
| A_44_P378749 | −14.7924 | LOC100912608 | Homeobox protein
Hox-A10-like |
| A_44_P547892 | −14.7229 | Olr1345 | Olfactory receptor
1345 |
| A_64_P001947 | −14.5408 | - | - |
| A_64_P009237 | −14.3488 | - | - |
| A_64_P029912 | −13.3859 | Cldnd2 | Claudin domain
containing 2 |
| A_64_P074460 | −13.1395 | - | - |
Hierarchical clustering analysis of the
differentially expressed mRNAs and lncRNAs
Following the determination of the expression values
of the differentially expressed genes (DEGs), the present study
carried out hierarchical clustering analysis on the DEGs. As shown
in Figs. 1 and 2, the differentially expressed mRNAs and
lncRNAs clearly distinguished the cardiac I/R tissues from the
control samples. In the cardiac I/R tissues, there were more
downregulated genes than upregulated genes (Figs. 1 and 2).
Functional and pathway enrichment
analyses
The significantly enriched GO terms (top 30) were
comprised of 16 biological processes, 6 cellular compartments, and
8 molecular functions (Fig. 3).
It was revealed that the differentially expressed biological
processes in the spinal cords were primarily involved in the
serotonin receptor signaling pathway, regulation of protein kinase
B signaling, regulation of keratinocyte migration, and skeletal
muscle satellite cell differentiation. The significantly enriched
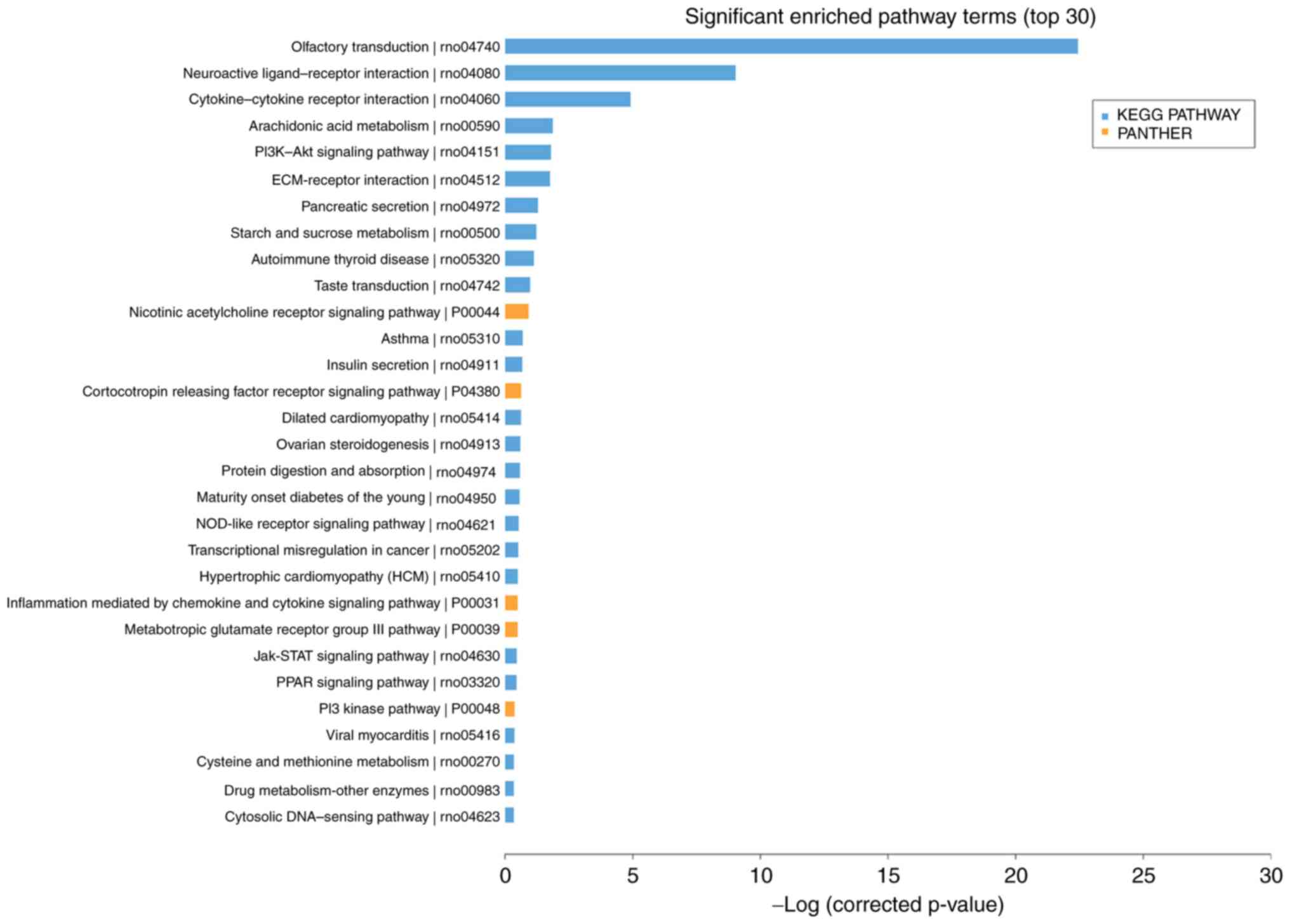
pathway terms (top 30) primarily involved KEGG pathways including
'olfactory transduction', 'arachidonic acid metabolism', the
'phosphoinositide 3-kinase-protein kinase B signaling pathway',
'extracellular matrix-receptor interaction', 'cytokine-cytokine
receptor interaction' and 'neuroactive ligand-receptor interaction'
(Fig. 4). The DEGs were analyzed
with GO background significant enrichment, which demonstrated the
number of genes associated with biological processes, cellular
compartments and molecular functions (Fig. 5).
The results of the biological process analysis
revealed that the DEGs involved in PPI networks were mainly
enriched in 'neurological system processes'
(P=1.92×10−11), 'sensory perception'
(P=1.92×10−11), the 'detection of chemical stimuli'
(P=5.10×10−10) and 'cell surface receptor signaling
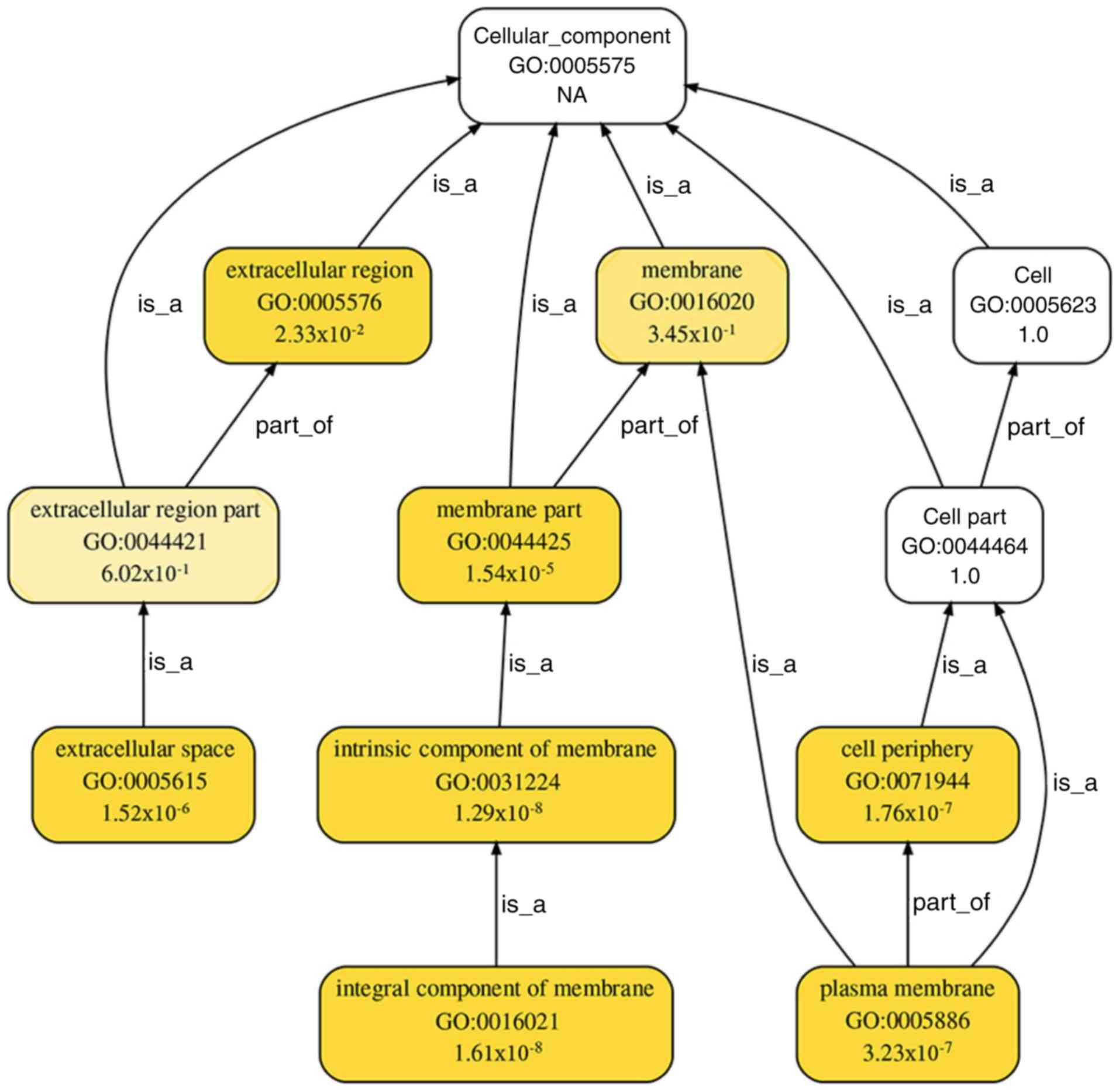
pathways' (P=4.23×10−10; Fig. 6). A total of 7 cellular components
from GO terms were significantly enriched for the DEGs involved in
PPI networks: 'Extracellular regions' (P=0.02), 'membrane parts'
(P=1.54×10−5), 'extracellular spaces'
(P=1.52×10−6), 'intrinsic components of the membrane'
(P=1.29×10−8), the 'cell periphery'
(P=1.76×10−7), 'integral components of the membrane'
(P=1.61×10−8) and the 'plasma membrane'
(P=3.22×10−7; Fig. 7).
A total of 10 molecular functions from GO terms were significantly
enriched for the DEGs involved in PPI networks: 'Molecular
transducer activity' (P=6.13×10−14), 'signal transducer
activity' (P=9.42×10−14), 'receptor activity' (P=
4.77×10−14), 'signal receptor activity'
(P=4.77×10−15), 'transmembrane signal receptor activity'
(P=1.31×10−15), 'serine-type peptidase activity'
(P=9.57×10−5), 'endopeptidase activity' (P=0.02),
'olfactory receptor activity' (P=5.07×10−9),
'G-protein-coupled receptor activity' (P=3.03×10−14) and
'serine-type endopeptidase activity' (P=1.08×10−5)
(Fig. 8).
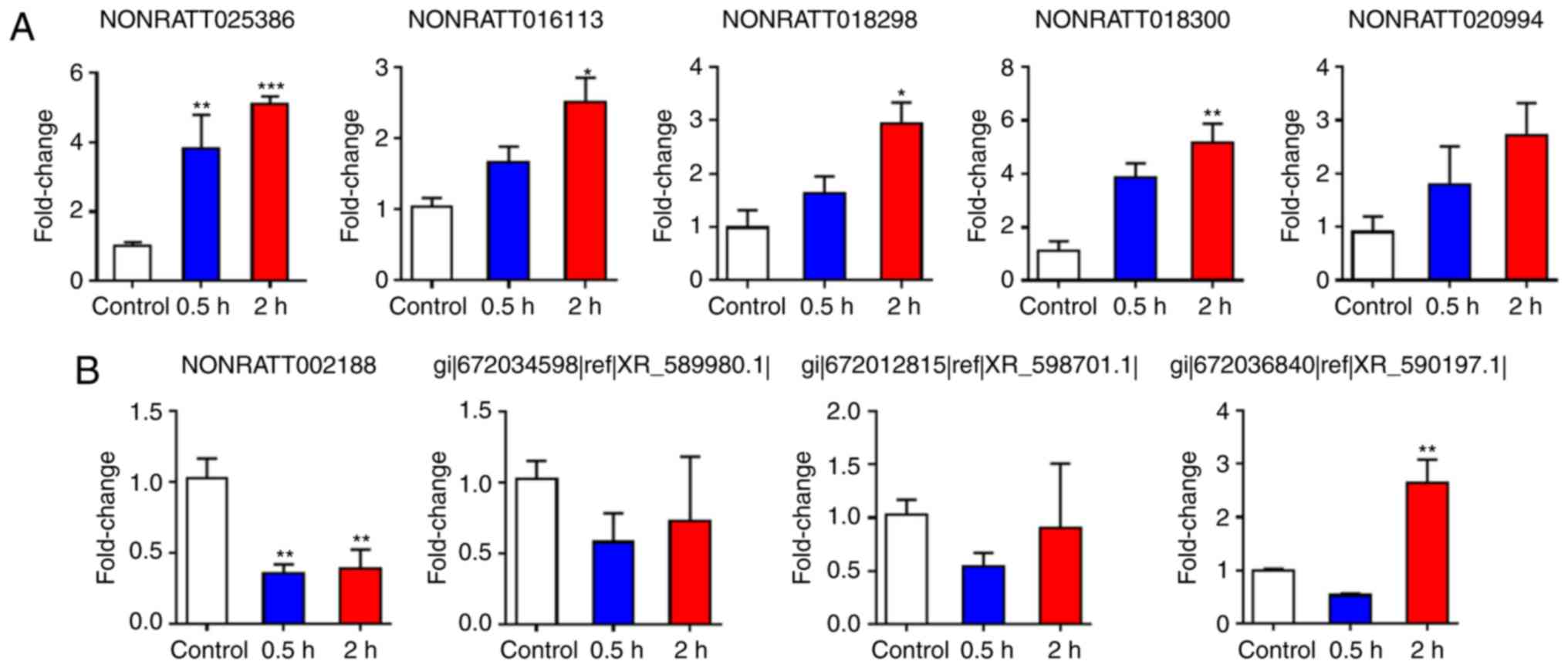
RT-qPCR validation of lncRNA expression
in the spinal cords 2 h post-cardiac I/R injury
To validate the reliability of the RNA sequencing
results in the rats, the present study analyzed the differentially
expressed lncRNAs, including 5 upregulated lncRNAs and 4
downregulated lncRNAs, by RT-qPCR analysis. T1-T4 spinal cord
tissues were collected from the control and I/R groups 2 h
post-reperfusion. The expression levels of 4 upregulated lncRNAs
(NONRATT025386, NONRATT016113, NONRATT018298 and NONRATT018300)
increased significantly in the I/R group when compared with those
in the control group, whereas the expression level of one
downregulated lncRNA (NONRATT002188) decreased significantly
(Fig. 9). The RT-qPCR results for
3 lncRNAs (XR_589980.1, XR_598701.1 and XR_590197.1) were not
consistent with the data from the RNA sequencing (XR_589980.1
decreased, and XR_598701.1 and XR_590197.1 increased
post-reperfusion when compared with the control; Fig. 10).
Expression levels of 4 lncRNAs in the
spinal cord 0.5 h post-cardiac I/R injury
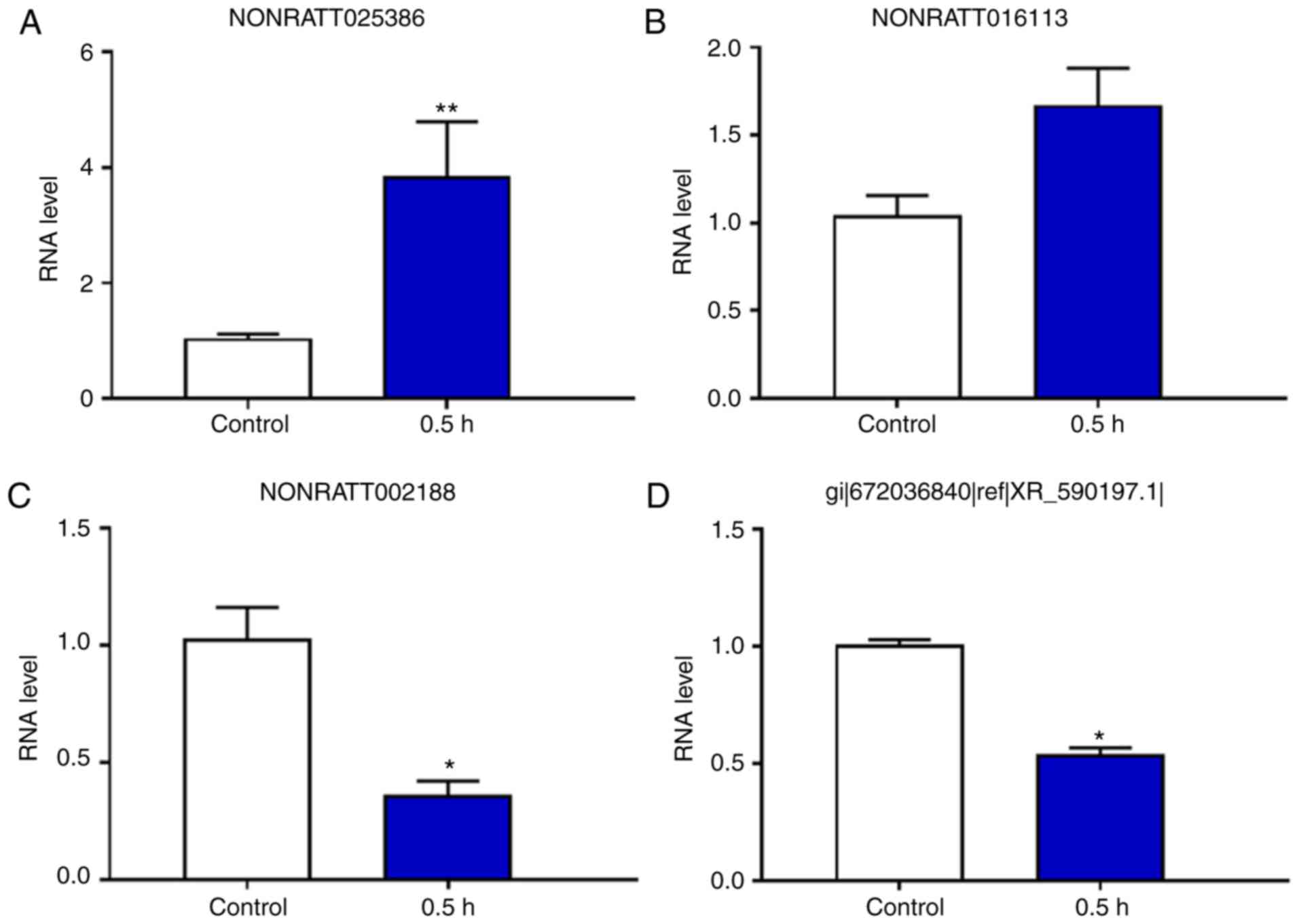
The present results indicated that the expression
levels of the lncRNA NONRATT025386 were significantly upregulated
in the 0.5 h group when compared with the control group, whereas
the expression levels of the lncRNAs NONRATT002188 and XR_590197.1
were signifi-cantly downregulated in the 0.5 h group compared with
the control group. Furthermore, the lncRNA NONRATT016113 showed no
significant difference between the two groups (Figs. 10 and 11).
Expression levels of 9 lncRNAs in the
spinal cord at different time-points (0.5 and 2 h) following
cardiac I/R injury
The present study collected samples from spinal cord
tissues at 0.5 and 2 h post-cardiac I/R injury for RT-qPCR
validation. The results indicated that the expression levels of the
lncRNA NONRATT025386 were significantly upregulated in the 0.5 and
2 h groups compared with the control group. In addition, the
expression levels of NONRATT016113, NONRATT018298 and NONRATT018300
were also significantly increased in the 2 h group, but not at 0.5
h, compared with the control group (Figs. 9–11). By contrast, the expression levels
of the lncRNA NONRATT002188 were significantly downregulated in the
0.5 and 2 h groups when compared with the control group (Figs. 9–11).
Discussion
With regard to ischemic cardiac tissues, previous
research has focused on multiple signaling pathways that regulate
the critical balance between cell death and survival during cardiac
I/R injury (42-47). The present study, to the best of
our knowledge, for the first time provides evidence that suggests
that many DEGs, pathways and biological processes of the T1-T4
spinal cord are implicated in myocardial I/R. Based on
high-throughput RNA seq, 16,987 lncRNAs in the T1-T4 spinal cord
tissues were identified, of which 3,621 were upregulated and 13,366
were downregulated (>2.0-fold-change; P<0.05). Among the
26,466 mRNAs that were quantified using RPKM values, 3,428 were
deregulated by 2-fold following I/R-induced cardiac injury, of
which 767 were upregulated and 2,661 were downregulated. According
to these results, some differentially expressed lncRNAs were
verified by RT-qPCR analyses.
Previous studies have shown that the spinal cord
serves an important role in the pathogenesis of cardiac disease
(48-51). The present study used a virally
mediated trans-synaptic tracing method, by injecting the PRV virus
into the rat heart and kidney, and these viruses were subsequently
found in the lateral medial column of the spinal cord in the
corresponding segment (12,52). This revealed the characteristics
of the transcriptome in the T1-T4 spinal cord following cardiac I/R
injury and the specific spinal segment that innervates the heart
serves a significant role in cardiac I/R injury. Cheng et al
(53) reported that melatonin
regulation of the transcriptome was associated with the reversal of
morphine tolerance by transcriptomic analysis of the L5-S3
segmental spinal cord. Mohrman et al (54) revealed the spinal cord
transcriptomic and metabolic patterns in a excitotoxic injection
injury model of syringomyelia. Niu et al (55) demonstrated the upregulation of
tumor necrosis factor (TNF)-α in spinal cord neurons during
coronary artery occlusion in rats, suggesting that TNF-α in the
spinal cord may be associated with the nociception initiated by
acute myocardial ischemia/infarction. Schulz et al (56) reported that connexin 43 in the
spinal cord serves an important role in providing protection from
cardiac I/R injuries. It is well known that myocardial ischemia
creates an autonomic nervous system imbalance, and can trigger
cardiac arrhythmias (57,58). Howard-Quijano et al
(59) indicated that
neuromodulation by spinal cord stimulation (SCS), in which a 4-pole
lead was placed percutaneously in the T1-T4 epidural space,
attenuated local cardiac sympathoexcitation from ischemia-induced
increases in afferent signaling, reduced ventricular arrhythmias
and improved myocardial function during acute ischemia. Liao et
al (60) reported that
chronic thoracic SCS at the T1-T3 level induced significant
remodeling of cardiac sympathetic innervation over the peri-infarct
and infarct regions, and was associated with improved left
ventricular function and reduced myocardial norepinephrine
spillover. The present study revealed that the differential
expression of certain mRNAs and lncRNAs in the spinal cord affects
the myocardial ischemic regions, suggesting that in the spinal
T1-T4 segment these genes are involved in the response to cardiac
injury. Although the functions of mRNAs and lncRNAs in the spinal
cord are unclear, the present findings provide a novel paradigm for
cardioprotection against I/R-induced myocardial injury.
The present results also revealed that some mRNAs in
the spinal cord, including chemokine C-X-C motif ligand 1 (CXCL1),
regulatory factor X4 (RFX4), WNT10a and interleukin (IL)-6, were
differentially expressed following cardiac I/R. Haider et al
(61) reported that the
angiogenic potential of the mononuclear cell (MNC) secretome is
regulated by CXCL-1 upregulation in spinal cord tissue, and factors
in the MNC secretome may mitigate the pathophysiological processes
of secondary damage following spinal cord injury, and may also
improve functional outcomes in rats. Ashique et al (62) reported that the spinal cords of
RFX4 mutants were correlated with defects in patterning and cilia
formation, suggesting that RFX4 is a regionally specific
transcriptional regulator of Sonic hedgehog signaling during the
development of the central nervous system. Zhao et al
(63) demonstrated that
hyperbaric oxygen (HBO) reverses Wnt-10a upregulation induced by
chronic constriction (CCI) injury in the dorsal root ganglion
(DRG), spinal cord and hippocampus, suggesting that HBO attenuates
CCI-induced rat neuropathic pain and inflammatory responses,
potentially through regulation of the Kindlin-1/Wnt-10a signaling
pathway. Although the detailed functions of many mRNAs from spinal
cords are not fully understood, the present results provide novel
insight into the molecular mechanisms underlying cardiac I/R
injury.
It is well known that regulations of gene expression
are varied over the time course (64-66). Our previous research associated
with Itchy E3 ubiquitin protein ligase demonstrated that gene
expression was significantly different in the C5-C8 spinal cord at
0.5 and 2 h following compound 48/80 injection when compared with
the control group (36). Similar
to the above method, we also screened key genes in myocardial
tissues under 30 min cardiac ischemia following 2 h reperfusion
compared with the sham group (67). Li et al (68) also indicated that impairment of
sensory nerves with significant reductions in CGRP and SP in the
DRG, ventricular myocardium and serum may be associated with an
increase in myocardial vulnerability in acute cardiac I/R injury in
diabetic rats. It was revealed that the injury was relatively
evident at 2 h post-myocardial reperfusion, which may be considered
as an acute cardiac I/R injury. Notably, this time point could be
equivalent to the patients who received early percutaneous coronary
intervention following myocardial infarction (69,70). Therefore, the present study chose
0.5 and 2 h post-reperfusion as the different time points of
cardiac reperfusion injury. The present study revealed that there
are significant differences in lncRNA and mRNA expression patterns
at different time points of myocardial I/R, suggesting that the
neural modulation of cardiac I/R injury may be temporal- and
spatial-dependent.
In particular, our previous study also demonstrated
that proton magnetic resonance spectroscopy was able to
simultaneously detect and quantify the absolute concentrations of
multiple metabolites within the spinal cord underlying
α-Me-5-HT-evoked pruritus (71).
Using the above method, we can also detect the changes of various
metabolites in the spinal cord following cardiac I/R injury, which
may deepen our understandings of the pathophysiology and
pharmacological therapies for acute myocardial infarction.
The present study screened several differentially
expressed mRNAs under cardiac I/R injury. In the future, whether
the proteins encoded by the mRNAs are consistent with these mRNAs
will be verified by immunoblotting. If so, the effects on cardiac
I/R injury may be observed by activating or silencing the
expression of one specific mRNA. Li et al (68) revealed that the regulation of the
spinal cord served a significant role under cardiac I/R injury in
diabetic neuropathic rats. Thus, it can be hypothesized that
intervention on the spinal cord may have an important influence on
cardioprotection in the future.
The present study used high-throughput RNA seq,
coupled with RT-qPCR analysis, to demonstrate that the expression
profiles of lncRNAs and mRNAs in spinal cords differed markedly
between the control and 2 h groups, and ultimately identified 7,980
differentially expressed (>2-fold) lncRNAs (234 upregulated and
7,746 downregulated) and 3,428 mRNAs (767 upregulated and 2,661
downregulated). The expression patterns of several lncRNAs were
confirmed by RT-qPCR. The results also indicated that the
expression levels of the lncRNA NONRATT025386 were significantly
upregulated in the 0.5 and 2 h groups compared with the control
group, whereas the expression levels of NONRATT016113,
NONRATT018298 and NONRATT018300 were significantly increased in the
2 h group when compared with the control group, although there was
no statistically significant difference between the expression
levels in the 0.5 h and control groups. Furthermore, the expression
levels of the lncRNA NONRATT002188 were significantly downregulated
in the 0.5 and 2 h groups compared with the control group.
In conclusion, this study revealed that
high-throughput RNA seq can facilitate the systematic exploration
of gene expression on a genome-wide scale, and can be used to
investigate DEGs and lncRNA expression patterns in the spinal cords
of rats during I/R-induced cardiac injury. In the search for better
treatments for cardiac I/R injury, expanded sets of differentially
expressed mRNAs and lncRNAs may prove very useful for identifying
novel therapeutic targets.
Funding
The present study was supported by grants from
National Natural Science Foundation of P.R. China (grant nos.
81670240, 81873467 and 81770283), National Natural Science
Foundation of Hubei Province (grant no. 2016CFB625), Key Research
and Development Project of Hainan Province of China (grant no.
ZDYF2018115) and Medical Innovation Project in Fujian Province
(grant no. 2017-CX-48).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HX and DW conceived and designed the study. QW and
ZL performed the surgical procedures. YL and ZH participated in the
experimental design. ZL and YC performed the experiments. MF and SL
analyzed the data. HX and DW wrote the manuscript and all authors
contributed to the final manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Ethical Committee of Tongji Hospital, Tongji Medical College,
Huazhong University of Science and Technology (Hubei, China; no.
TJ-A20150804).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
The authors would like to thank CapitalBio
Technology Co., Ltd. (Beijing, China) for their technical advice
and Dr Taotao Liu (Department of Anesthesiology, Peking University
Third Hospital, Peking University, Beijing, China) for his
contribution to image acquisition.
References
|
1
|
Shi J, Bei Y, Kong X, Liu X, Lei Z, Xu T,
Wang H, Xuan Q, Chen P, Xu J, et al: miR-17-3p contributes to
exercise-induced cardiac growth and protects against myocardial
ischemia-reperfusion injury. Theranostics. 7:664–676. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu X, Xiao J, Zhu H, Wei X, Platt C,
Damilano F, Xiao C, Bezzerides V, Boström P, Che L, et al: miR-222
is necessary for exercise-induced cardiac growth and protects
against pathological cardiac remodeling. Cell Metab. 21:584–595.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Burley DS, Ferdinandy P and Baxter GF:
Cyclic GMP and protein kinase-G in myocardial
ischaemia-reperfusion: Opportunities and obstacles for survival
signaling. Br J Pharmacol. 152:855–869. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wong GT, Yao L, Xia Z and Irwin MG:
Intrathecal morphine remotely preconditions the heart via a neural
pathway. J Cardiovasc Pharmacol. 60:172–178. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ding X, Ardell JL, Hua F, McAuley RJ,
Sutherly K, Daniel JJ and Williams CA: Modulation of cardiac
ischemia-sensitive afferent neuron signaling by preemptive C2
spinal cord stimulation: Effect on substance P release from rat
spinal cord. Am J Physiol Regul Integr Comp Physiol. 294:R93–R101.
2008. View Article : Google Scholar
|
|
6
|
Sroka K: On the genesis of myocardial
ischemia. Z Kardiol. 93:768–783. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zipes DP: Heart-brain interactions in
cardiac arrhythmias: Role of the autonomic nervous system. Cleve
Clin J Med. 75(Suppl 2): S94–S96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Armour JA: Cardiac neuronal hierarchy in
health and disease. Am J Physiol Regul Integr Comp Physiol.
287:R262–R271. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Armour JA, Linderoth B, Arora RC,
DeJongste MJ, Ardell JL, Kingma JG Jr, Hill M and Foreman RD:
Long-term modulation of the intrinsic cardiac nervous system by
spinal cord neurons in normal and ischaemic hearts. Auton Neurosci.
95:71–79. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pozzati A, Pancaldi LG, Di Pasquale G,
Pinelli G and Bugiardini R: Transient sympathovagal imbalance
triggers 'ischemic' sudden death in patients undergoing
electrocardio-graphic Holter monitoring. J Am Coll Cardiol.
27:847–852. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Airaksinen KE: Autonomic mechanisms and
sudden death after abrupt coronary occlusion. Ann Med. 31:240–245.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu LJ, Liu TT, He ZG, Hong QX and Xiang
HB: Hypothesis: CeM-RVLM circuits may be implicated in sudden
unexpected death in epilepsy by melanocortinergic-Sympathetic
signaling. Epilepsy Behav. 45:124–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li R, Wong GT, Wong TM, Zhang Y, Xia Z and
Irwin MG: Intrathecal morphine preconditioning induces
cardioprotection via activation of delta, kappa, and mu opioid
receptors in rats. Anesth Analg. 108:23–29. 2009. View Article : Google Scholar
|
|
14
|
Lu Y, Hu J, Zhang Y, Dong CS and Wong GT:
Remote intra-thecal morphine preconditioning confers
cardioprotection via spinal cord nitric oxide/cyclic guanosine
monophosphate/protein kinase G pathway. J Surg Res. 193:43–51.
2015. View Article : Google Scholar
|
|
15
|
Jiang L, Hu J, He S, Zhang L and Zhang Y:
Spinal neuronal NOS signaling contributes to morphine
cardioprotection in ischemia reperfusion injury in rats. J
Pharmacol Exp Ther. 358:450–456. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mei B, Li W, Cheng X, Liu X, Gu E and
Zhang Y: Activating mu-opioid receptors in the spinal cord mediates
the cardioprotective effect of remote preconditioning of trauma.
Cardiol J. 24:314–323. 2017. View Article : Google Scholar
|
|
17
|
Wimalawansa SJ: Calcitonin gene-related
peptide and its receptors: Molecular genetics, physiology,
pathophysiology, and therapeutic potentials. Endocr Rev.
17:533–585. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Y, Chen AF and Wang DH: ET(A)
receptor blockade prevents renal dysfunction in salt-sensitive
hypertension induced by sensory denervation. Am J Physiol Heart
Circ Physiol. 289:H2005–H2011. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu S, Marie Lutz B, Miao X, Liang L, Mo K,
Chang YJ, Du P, Soteropoulos P, Tian B, Kaufman AG, et al: Dorsal
root ganglion transcriptome analysis following peripheral nerve
injury in mice. Mol Pain. 12:2016. View Article : Google Scholar
|
|
20
|
Wang S, Xu H, Zou L, Xie J, Wu H, Wu B, Yi
Z, Lv Q, Zhang X, Ying M, et al: LncRNA uc.48+ is involved in
diabetic neuro-pathic pain mediated by the P2X receptor in the
dorsal root ganglia. Purinergic Signal. 12:138–148. 2015.
|
|
21
|
Boon RA, Jae N, Holdt L and Dimmeler S:
Long noncoding RNAs: From clinical genetics to therapeutic targets.
J Am Coll Cardiol. 67:1214–1226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao X, Tang Z, Zhang H, Atianjoh FE, Zhao
JY, Liang L, Wang W, Guan X, Kao SC, Tiwari V, et al: A long
noncoding RNA contributes to neuropathic pain by silencing Kcna2 in
primary afferent neurons. Nat Neurosci. 16:1024–1031. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang BC, Sun WX, He LN, Cao DL, Zhang ZJ
and Gao YJ: Identification of lncRNA expression profile in the
spinal cord of mice following spinal nerve ligation-induced
neuropathic pain. Mol Pain. 11:432015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Q, Li ZX, Liu BW, He ZG, Liu C, Chen
M, Liu SG, Wu WZ and Xiang HB: Altered expression of differential
gene and lncRNA in the lower thoracic spinal cord on different time
courses of experimental obstructive jaundice model accompanied with
altered peripheral nociception in rats. Oncotarget.
8:106098–106112. 2017.PubMed/NCBI
|
|
25
|
Liu Y, Li G, Lu H, Li W, Li X, Liu H, Li
X, Li T and Yu B: Expression profiling and ontology analysis of
long noncoding RNAs in post-ischemic heart and their implied roles
in ischemia/reperfusion injury. Gene. 543:15–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu Y, Zhou D, Li G, Ming X, Tu YF, Tian
J, Lu H and Yu B: Long non coding RNA-UCA1 contributes to
cardiomyocyte apoptosis by suppression of p27 expression. Cell
Physiol Biochem. 35:1986–1998. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vausort M, Wagner DR and Devaux Y: Long
noncoding RNAs in patients with acute myocardial infarction. Circ
Res. 115:668–677. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Grote P, Wittler L, Hendrix D, Koch F,
Währisch S, Beisaw A, Macura K, Bläss G, Kellis M, Werber M and
Herrmann BG: The tissue-specific lncRNA Fendrr is an essential
regulator of heart and body wall development in the mouse. Dev
Cell. 24:206–214. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Klattenhoff CA, Scheuermann JC, Surface
LE, Bradley RK, Fields PA, Steinhauser ML, Ding H, Butty VL, Torrey
L, Haas S, et al: Braveheart, a long noncoding RNA required for
cardiovascular lineage commitment. Cell. 152:570–583. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
31
|
Murry CE, Jennings RB and Reimer KA:
Preconditioning with ischemia: A delay of lethal cell injury in
ischemic myocardium. Circulation. 74:1124–1136. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang CH, Lai CC, Yang AH and Chiang SC:
Myocardial preconditioning reduces kidney injury and apoptosis
induced by myocardial ischaemia and reperfusion. Eur J Cardthorac
Surg. 48:382–391. 2015. View Article : Google Scholar
|
|
33
|
Li ZX, Lin Q, He ZG, Wang Q, Chen YL, Feng
MH, Li SY and Xiang HB: Altered myocardial gene expression
profiling in the ischemic tissues at different time points after
cardiac ischemia/reperfusion in rats. Oncotarget. 9:2018.
|
|
34
|
Flecknell PA: Anaesthesia of animals for
biomedical research. Br J Anaesth. 71:885–894. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu QQ, Liu H, He ZG, Zhang SJ, Liu BW,
Wang L, Qiu WH, Xu Q, Xiang HB and Lv YM: Differential gene and
lncRNA expression in the lower thoracic spinal cord following
isch-emia/reperfusion-induced acute kidney injury in rats.
Oncotarget. 8:53465–53481. 2017.PubMed/NCBI
|
|
36
|
Liu BW, Li ZX, He ZG, Liu C, Xiong J and
Xiang HB: Altered expression of target genes of spinal cord in
different itch models compared with capsaicin assessed by RT-qPCR
validation. Oncotarget. 8:74423–74433. 2017.PubMed/NCBI
|
|
37
|
He ZG, Liu BW, Li ZX, Liu C and Xiang HB:
Altered expression profiling of spinal genes modulated by compound
48/80 in a mouse itch model. J Anesth Perioper Med. 4:220–224.
2017. View Article : Google Scholar
|
|
38
|
Orom UA, Derrien T, Beringer M, Gumireddy
K, Gardini A, Bussotti G, Lai F, Zytnicki M, Notredame C, Huang Q,
et al: Long noncoding RNAs with enhancer-like function in human
cells. Cell. 143:46–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bottomly D, Walter NA, Hunter JE,
Darakjian P, Kawane S, Buck KJ, Searles RP, Mooney M, McWeeney SK
and Hitzemann R: Evaluating gene expression in C57BL/6J and DBA/2J
mouse striatum using RNA-Seq and microarrays. PLoS One.
6:e178202011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu Y, Zhang XH and Pang YZ: Association of
tyrosinase (TYR) and tyrosinase-related protein 1 (TYRP1) with
melanic plumage color in korean quails (Coturnix coturnix).
Asian-Australas J Anim Sci. 26:1518–1522. 2013. View Article : Google Scholar
|
|
41
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Peng J, Lu R, Ye F, Deng HW and Li YJ: The
heme oxygenase-1 pathway is involved in calcitonin gene-related
peptide-mediated delayed cardioprotection induced by monophosphoryl
lipid A in rats. Regul Pept. 103:1–7. 2002. View Article : Google Scholar
|
|
43
|
Kawai S, Yamada T, Matsuura T, Funao T and
Nishikawa K: Neuropathic pain attenuates ischemia reperfusion
injury through beta2-adrenergic pathway. Life Sci. 187:9–16. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Redington KL, Disenhouse T, Strantzas SC,
Gladstone R, Wei C, Tropak MB, Dai X, Manlhiot C, Li J and
Redington AN: Remote cardioprotection by direct peripheral nerve
stimulation and topical capsaicin is mediated by circulating
humoral factors. Basic Res Cardiol. 107:2412012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chiu JH, Cheng YF, Wang JY and Hsu CF:
Remote pharmacological preconditioning on median nerve territory
increases Hsp32 expression and attenuates ischemia-reperfusion
injury in rat heart. Life Sci. 90:629–636. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Polhemus DJ, Gao J, Scarborough AL,
Trivedi R, McDonough KH, Goodchild TT, Smart F, Kapusta DR and
Lefer DJ: Radiofrequency renal denervation protects the isch-emic
heart via inhibition of GRK2 and increased nitric oxide signaling.
Circ Res. 119:470–480. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Miura T, Kawamura S, Tatsuno H, Ikeda Y,
Mikami S, Iwamoto H, Okamura T, Iwatate M, Kimura M, Dairaku Y, et
al: Ischemic preconditioning attenuates cardiac sympathetic nerve
injury via ATP-sensitive potassium channels during myocardial
ischemia. Circulation. 104:1053–1058. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Foreman RD, Garrett KM and Blair RW:
Mechanisms of cardiac pain. Compr Physiol. 5:929–960. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Santos SF, Rebelo S, Derkach VA and
Safronov BV: Excitatory interneurons dominate sensory processing in
the spinal substantia gelatinosa of rat. J Physiol. 581:241–254.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Franco-Cereceda A, Kallner G and Lundberg
JM: Capsazepine-sensitive release of calcitonin gene-related
peptide from C-fibre afferents in the guinea-pig heart by low pH
and lactic acid. Eur J Pharmacol. 238:311–316. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Steagall RJ, Sipe AL, Williams CA, Joyner
WL and Singh K: Substance P release in response to cardiac ischemia
from rat thoracic spinal dorsal horn is mediated by TRPV1.
Neuroscience. 214:106–119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ye DW, Li RC, Wu W, Liu C, Ni D, Huang QB,
Ma X, Li HZ, Yang H, Xiang HB and Zhang X: Role of spinal cord in
regulating mouse kidney: A virally mediated trans-synaptic tracing
study. Urology. 79:745e741–744. 2012. View Article : Google Scholar
|
|
53
|
Cheng YC, Tsai RY, Sung YT, Chen IJ, Tu
TY, Mao YY and Wong CS: Melatonin regulation of transcription in
the reversal of morphine tolerance: Microarray analysis of
differential gene expression. Int J Mol Med. 43:791–806. 2019.
|
|
54
|
Mohrman AE, Farrag M, Huang H, Ossowski S,
Haft S, Shriver LP and Leipzig ND: Spinal cord transcriptomic and
metabolomic analysis after excitotoxic injection injury model of
syringomyelia. J Neurotrauma. 34:720–733. 2017. View Article : Google Scholar
|
|
55
|
Niu YL, Guo Z and Zhou RH: Up-regulation
of TNF-alpha in neurons of dorsal root ganglia and spinal cord
during coronary artery occlusion in rats. Cytokine. 47:23–29. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Schulz R, Gorge PM, Gorbe A, Ferdinandy P,
Lampe PD and Leybaert L: Connexin 43 is an emerging therapeutic
target in ischemia/reperfusion injury, cardioprotection and
neuroprotection. Pharmacol Ther. 153:90–106. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ding X, Hua F, Sutherly K, Ardell JL and
Williams CA: C2 spinal cord stimulation induces dynorphin release
from rat T4 spinal cord: Potential modulation of myocardial
ischemia-sensitive neurons. Am J Physiol Regul Integr Comp Physiol.
295:R1519–R1528. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hua F, Ardell JL and Williams CA: Left
vagal stimulation induces dynorphin release and suppresses
substance P release from the rat thoracic spinal cord during
cardiac ischemia. Am J Physiol Regul Integr Comp Physiol.
287:R1468–R1477. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Howard-Quijano K, Takamiya T, Dale EA,
Kipke J, Kubo Y, Grogan T, Afyouni A, Shivkumar K and Mahajan A:
Spinal cord stimulation reduces ventricular arrhythmias during
acute ischemia by attenuation of regional myocardial excitability.
Am J Physiol Heart Circ Physiol. 313:H421–H431. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Liao SY, Liu Y, Zuo M, Zhang Y, Yue W, Au
KW, Lai WH, Wu Y, Shuto C, Chen P, et al: Remodelling of cardiac
sympathetic re-innervation with thoracic spinal cord stimulation
improves left ventricular function in a porcine model of heart
failure. Europace. 17:1875–1883. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Haider T, Hoftberger R, Ruger B, Mildner
M, Blumer R, Mitterbauer A, Buchacher T, Sherif C, Altmann P, Redl
H, et al: The secretome of apoptotic human peripheral blood
mononuclear cells attenuates secondary damage following spinal cord
injury in rats. Exp Neurol. 267:230–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ashique AM, Choe Y, Karlen M, May SR,
Phamluong K, Solloway MJ, Ericson J and Peterson AS: The Rfx4
transcription factor modulates Shh signaling by regional control of
ciliogen-esis. Sci Signal. 2:ra702009. View Article : Google Scholar
|
|
63
|
Zhao B, Pan Y, Xu H and Song X: Hyperbaric
oxygen attenuates neuropathic pain and reverses inflammatory
signaling likely via the Kindlin-1/Wnt-10a signaling pathway in the
chronic pain injury model in rats. J Headache Pain. 18:12017.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Harrison BJ, Venkat G, Hutson T, Rau KK,
Bunge MB, Mendell LM, Gage FH, Johnson RD, Hill C, Rouchka EC, et
al: Transcriptional changes in sensory ganglia associated with
primary afferent axon collateral sprouting in spared dermatome
model. Genom Data. 6:249–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Ke C, Gao F, Tian X, Li C, Shi D, He W and
Tian Y: Slit2/Robo1 mediation of synaptic plasticity contributes to
bone cancer pain. Mol Neurobiol. 54:295–307. 2017. View Article : Google Scholar
|
|
66
|
Knowlton WM, Palkar R, Lippoldt EK, McCoy
DD, Baluch F, Chen J and McKemy DD: A sensory-labeled line for
cold: TRPM8-expressing sensory neurons define the cellular basis
for cold, cold pain, and cooling-mediated analgesia. J Neurosci.
33:2837–2848. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang Q, He ZG, Li ZX, Li SY, Chen YL, Feng
MH, Hong QX and Xiang HB: Bioinformatics analysis of gene
expression profile data to screen key genes involved in cardiac
ischemia-reperfusion injury. Int J Clin Exp Med. 11:4955–4966.
2018.
|
|
68
|
Li TP, Guo Z, Liu CJ, Sun T, Chen L and
Zhao X: Association of down-regulation of calcitonin gene-related
peptide and substance P with increase of myocardial vulnerability
in diabetic neuropathic rats. Peptides. 96:1–7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Cheng YF, Chang YT, Chen WH, Shih HC, Chen
YH, Shyu BC and Chen CC: Cardioprotection induced in a mouse model
of neuropathic pain via anterior nucleus of paraventricular
thalamus. Nat Commun. 8:8262017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Liu T, He Z, Tian X, Kamal GM, Li Z, Liu
Z, Liu H, Xu F, Wang J and Xiang H: Specific patterns of spinal
metabolites underlying alpha-Me-5-HT-evoked pruritus compared with
histamine and capsaicin assessed by proton nuclear magnetic
resonance spectroscopy. Biochim Biophys Acta Mol Basis Dis.
1863:1222–1230. 2017. View Article : Google Scholar : PubMed/NCBI
|