Introduction
Endothelin (ET) plays important roles on various
physiological functions including vascular constriction (1–4). ET
family comprises three isoforms, ET-1, ET-2, and ET-3, that bind to
two receptor subtypes, endothelin A (ETA) and endothelin
B (ETB) receptors (1–4).
Recent studies reported that ETA and ETB were
highly expressed on lung, colon and skin cancers (5–7). In
addition, several reports suggested that ET-1 plays important roles
in tumorigenesis, tumor progression, and metastasis (8–10).
Thus, the ET receptors and their signalling pathways may be a
therapeutic target in cancer therapy (11). However, little is known about the
role of ET signalling on tumor cell proliferation of oral squamous
cell carcinoma (SCC).
Human SCC is major neoplasm in esophagus or oral
cavity and the incidence has recently been increasing (12–14).
The optimal treatment for early carcinoma of oral cavity is
surgical operation. However, overall survival remains largely
unchanged (12–14). In addition, the decrease in quality
of life (QOL) after wide excision of tongue is also important issue
for patients. Therefore, different therapies are required. In our
previous studies, we investigated the whole genome analysis using
DNA microarray to find the potential target genes involved in tumor
cell growth, and reported the critical role of several important
molecules on the cell growth of SCCs (15–19).
According to the results of DNA microarray, we found increased
expression of ET receptor mRNA in cell lines of oral SCCs and the
alteration of expression level on SCC growth (15,18,19).
Therefore, we have examined whether ET receptors may be expressed
in primary oral SCC tissues, and whether ET receptor-signalling may
play a critical role of SCC growth. Our results imply a potentially
important and novel role of ET function on SCC growth, and suggest
that ET receptor-signalling might be useful target in the therapy
of SCCs.
Materials and methods
Tissue samples
All of clinical studies were approved by the Ethics
Committee of Osaka University Dental Hospital. Twenty-three samples
of squamous cell carcinoma (SCC) located in the tongue were
obtained from surgical resection tissue specimens at Osaka
University Dental Hospital after informed consent was obtained. The
patients, who received no preoperative therapy including
chemotherapy and irradiation therapy, were randomly selected
(Table I). The age range was 33–92
years (average: 62.0±13.9 years, mean ± SD).
 | Table IProfile of lingual squamous cell
carcinoma patients, their histological diagnosis and expression of
ETA and ETB in tissue sections. |
Table I
Profile of lingual squamous cell
carcinoma patients, their histological diagnosis and expression of
ETA and ETB in tissue sections.
| Case | Age/Gender |
Differentiation | ETA
expression | ETB
expression |
|---|
| Tumor area | Non-tumor area | Tumor area | Non-tumor area |
|---|
| 1 | 71/F | Well differentiated
SCC | +++ | + | +++ | ++ |
| 2 | 57/M | Well differentiated
SCC | + | N/A | ++ | N/A |
| 3 | 69/M | Well differentiated
SCC | ++ | N/A | +++ | N/A |
| 4 | 64/M | Well differentiated
SCC | +++ | + | +++ | ++ |
| 5 | 46/F | Well differentiated
SCC | ++ | + | +++ | + |
| 6 | 61/M | Well differentiated
SCC | + | N/A | + | N/A |
| 7 | 48/M | Well differentiated
SCC | ++ | N/A | +++ | N/A |
| 8 | 72/M | Well differentiated
SCC | ++ | N/A | +++ | N/A |
| 9 | 79/M | Well differentiated
SCC | ++ | N/A | ++ | N/A |
| 10 | 68/M | Moderately
differentiated SCC | +++ | N/A | +++ | N/A |
| 11 | 64/M | Moderately
differentiated SCC | ++ | + | +++ | ++ |
| 12 | 58/F | Moderately
differentiated SCC | +++ | N/A | +++ | N/A |
| 13 | 92/F | Moderately
differentiated SCC | ++ | − | +++ | − |
| 14 | 86/F | Moderately
differentiated SCC | ++ | + | + | + |
| 15 | 57/M | Moderately
differentiated SCC | +++ | N/A | +++ | N/A |
| 16 | 62/M | Moderately
differentiated SCC | ++ | N/A | +++ | N/A |
| 17 | 52/F | Poor-moderately
differentiated SCC | ++ | N/A | ++ | N/A |
| 18 | 38/M | Poorly
differentiated SCC | + | − | ++ | + |
| 19 | 51/F | Poorly
differentiated SCC | ++ | − | ++ | + |
| 20 | 67/M | Poorly
differentiated SCC | ++ | N/A | ++ | N/A |
| 21 | 66/M | Poorly
differentiated SCC | ++ | N/A | +++ | N/A |
| 22 | 65/M | Poorly
differentiated SCC | ++ | N/A | +++ | N/A |
| 23 | 33/M | Poorly
differentiated SCC | ++ | − | +++ | + |
Chemicals and antibodies
ET receptor specific antagonists, BQ123 for
ETA and BQ788 for ETB were purchased from
Sigma-Aldrich Japan (Tokyo, Japan). Anti-ETA or
ETB polyclonal antibody was from Acris (Acris, Herford,
Germany). Antibodies against Focal adhesion kinase (FAK),
phosphorylated FAK, phosphorylated MEK1/2, p44/42 MAPK (pErk1/2)
phosphorylated p44/42 MAPK and anti-rabbit IgG (HRP-linked) for
secondary antibody are from Cell Signalling Technologies (Beverly,
MA). Cisplatin was from Wako Pure Chemical Industries, Ltd. (Osaka,
Japan).
Immunohistochemical staining of
ETA and ETB
The expression of ETA or ETB
in tissues was detected by anti-ETA or ETB
specific polyclonal antibody using standard immunohistochemical
techniques on formalin-fixed and paraffin-embedded continuous
sections. Incubation with anti-ETA or ETB
polyclonal antibody was performed at 4°C for 16 h, then the
sections were washed out. After the application with secondary
antibody, the Vectastain ABC kit (Vector Laboratories, Burlingame,
CA) was used with a 3,3′-diaminobenzidine (DAB) substrate kit,
according to the manufacturer’s instructions. The staining endpoint
was determined when the standard tissue sections were constantly
stained to the intensity as described previously (18,19).
The intensity of the immunohistochemical staining
with anti-ETA or ETB antibody was evaluated
by scoring the staining reaction in four groups: (−), none/weak;
(+), weak/moderate; (++), moderate/strong, and (+++), very strong
cytoplasmic staining intensity, respectively (18,19).
To check the reproducibility of the evaluation system concerning
the immunohistochemical staining for the ETA and
ETB proteins, another oral surgeon and pathologist who
were unaware of the original assessment re-evaluated the results of
staining according to the system above. Tumor areas were confirmed
by both of the pathologist and surgeon under the microscopy.
Non-tumor areas were selected, the comparatively normal areas were
separated away from the tumor areas, and confirmed by the
pathologist.
Cell culture
We used human oral SCC cell line (SAS) and human
esophageal SCC cell line (KYSE70). SAS was established as tongue
SCC and KYSE70 was established as esophageal SCC (15,16).
SAS was maintained in DMEM containing 10% fetal bovine serum (FBS),
and KYSE70 was maintained in DMEM containing 2% FBS at 37°C under
0.5% CO2. For cell growth experiment, cells were
trypsinized and replated onto culture dishes (15–19).
Cell survival assay using ET receptor
antagonists
SCC cells were treated with ET receptor antagonists,
BQ123 for ETA and BQ788 for ETB for 24 and 48
h in culture medium. Then, cell viability was measured 24 and 48 h
after the treatment using Countess Automated Cell Counter
(Invitrogen, Eugene, OR). The inhibition of cell growth was
compared to vehicle-treated control.
RNA interference approach
SAS and KYSE70 were trypsinized and resuspended in
DMEM without FBS, and the cells were separated approximately
1×105 cells for each dish. The ETA and
ETB-specific siRNA (Stealth siRNA) were purchased from
Invitrogen Japan (Tokyo, Japan). The sequence of the sense strand
of ETA-siRNA is 5′-UUUGAUGUGGCAUUGAGCAUACAGG-3′, and
antisense is 5′-CCUGUAUGCUCAAUGCCACAUCAAA-3′, respectively. The
sequence of the sense strand of ETB-siRNA is
5′-UAAUUCUACUCCAAGAAGCAACAGC-3′, and antisense is
5′-GCUGUUGCUUCUUGGAGUAGAAUUA-3′, respectively. For the
transfection, ETA, ETB-siRNA (40 nM) or
negative control (40 nM Stealth RNAi Negative Control Duplexes,
Invitrogen Japan Inc.) solution was added to DMEM medium containing
Lipofectamine RNAiMax (Invitrogen Japan) and allowed to incubate
for 20 min at room temperature to create the transfection mixture.
The transfection mixture was then added to the cells at the
indicated final concentration of siRNA. Twenty-four hours after the
transfection, the medium was changed to DMEM containing 10% FBS for
SAS and 2% FBS for KYSE70. Then, viable cell number was measured 24
and 48 h after the medium change using Countess Automated Cell
Counter. The cell growth was expressed as the percentage to that of
vehicle control.
Western blot analysis
Adherent or suspended cells were washed in PBS, and
cell extracts were prepared by lysing cells in lysis buffer. The
proteins were separated by electrophoresis using 10% SDS-PAGE, and
transferred to nitrocellulose membrane (Millipore, Bedford, MA).
Detection of proteins were performed by each polyclonal antibody
and visualized by using the ECL detection kit (Amersham, London,
UK) following the manufacturer’s suggested procedure.
Combination of ETA,
ETB-siRNA and anti-tumor drug
Com- bined treatment of ETA,
ETB-siRNA with anti-tumor drug, cisplatin, was
performed. Briefly, after the low concentration of ETA
or ETB-siRNA (20 nM) treatment, 2.5 μM of cisplatin that
was a concentration slightly effective on cell growth inhibition
was treated for 48 h. Cell growth was measured by Countess
Automated Cell Counter and expressed as percentage.
Statistical analysis
All results are expressed as mean ± SEM. Statistical
comparisons were made using the Student-t test or Scheffe’s method
after analysis of variances (ANOVA). The results were considered
significantly different at P<0.05.
Results
Lingual SCCs in tumor tissues express
ETA and ETB
Lingual SCC primary tissues were stained using
anti-ETA or anti-ETB specific antibody,
respectively. Positive staining of ETA was observed in
tumor area (Fig. 1A, right). In
contrast, none of staining of ETA was observed in
non-tumor area in the same tissue section (Fig. 1A, left). Similar staining pattern
was also observed in other tumor tissue sections (Table I). Statistically significance of
the ETA expression between tumor and non-tumor area was
observed (Fig. 1B). In addition,
positive staining of ETB in tumor area, but not
non-tumor area, was also observed in the same tissue section
(Fig. 1C). Statistical
significance of the ETB expression between tumor and
non-tumor areas was also observed (Fig. 1D and Table I). These results are similar to
that of ETA. Good correlation between ETA and
ETB expression was observed (Fig. 1E).
ET receptor antagonists suppress cell
growth of lingual and esophageal SCC
According to the data of ET receptor expression in
SCCs, we hypothesized that ET receptor-signalling might play an
important role on the cell growth of SCCs. To investigate the
hypothesis, we used ET receptor antagonists, BQ123 for
ETA and BQ788 for ETB. As shown in Fig. 2A and B, ET receptor antagonists,
BQ123 and BQ788 suppressed the cell growth of lingual SCC cell
line, SAS. The suppression by the antagonists was concentration-
and time-dependent (data not shown).
In addition to the results of growth suppression of
lingual SCC by the inhibition of ET receptors, both antagonists
also suppressed the cell growth of esophageal SCC cell line, KYSE70
(Fig. 2C and D). These results
indicate that ET receptor-signalling is required for the growth of
SCCs.
ETA and ETB-siRNA
suppress cell growth of lingual and esophageal SCC
To clarify the exact function of ET receptors on the
growth of SCCs, we used small interfering RNA (siRNA) for
ETA and ETB. ETA and
ETB-siRNA effectively decreased the ET receptor protein
levels in SCCs. The inhibition of cell growth on SAS was clearly
observed when ETA or ETB was knocked down by
the treatment with siRNA (Fig. 3A and
B). Similar suppression of cell growth by the knockdown of
ETA or ETB was also observed when esophageal
SCC cell line, KYSE70 was treated with siRNA for ETA or
ETB (Fig. 3C and D).
These results clearly indicate that ET receptor-signalling is
required for the growth of SCCs.
Investigation of potential
mechanisms
We next investigated the mechanisms of inhibition of
cell growth induced by the suppression of ET receptor-signalling.
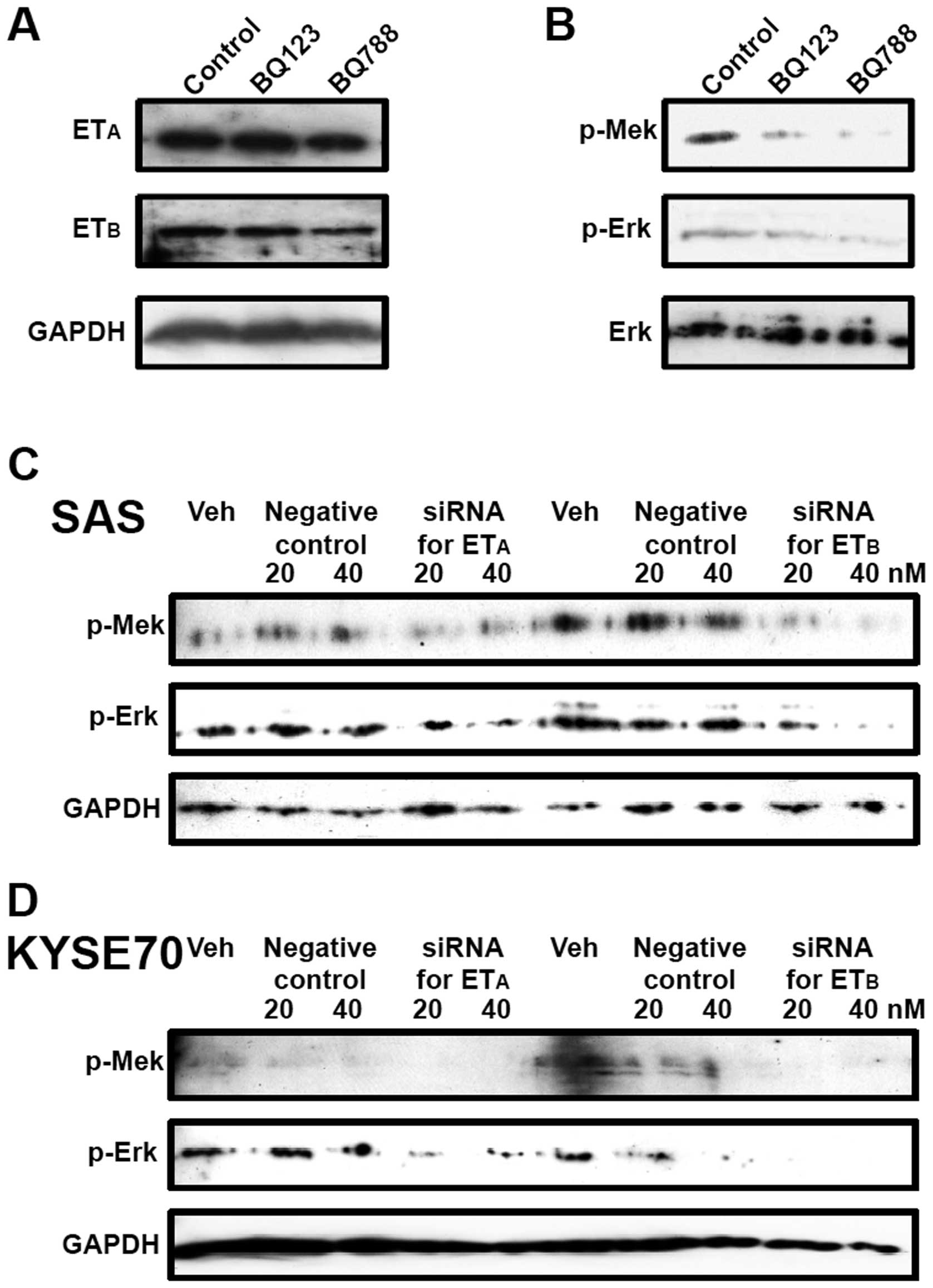
Western blot analysis showed the expression of ETA and
ETB proteins on the lingual SCC cell line SAS (Fig. 4A). Although the specific
antagonists blocked the ETA or ETB
signalling, no alterations of receptor protein expression levels
were observed (Fig. 4A). In
contrast, blockade of ET receptor-signalling by the treatment with
antagonists caused the suppression of phosphorylation of MEK and
Erk (mitogen-activated protein kinase), the important members of
MAPK pathway (Fig. 4B). In
addition, similar suppression of MAPK pathway by knockdown of ET
receptors was observed when SAS and KYSE70 were treated with
ETA or ETB-siRNA (Fig. 4C and D). These results indicate the
involvement of MAPK pathway on the ET receptor-signalling mediated
cell growth of SCCs.
In contrast, no inhibition of phosphorylation of
focal adhesion kinase (FAK), a 125 kDa non-receptor tyrosine kinase
(20,21), by the suppression of ET receptor
signalling was observed (data not shown).
We also investigated the effect of blockade of ET
receptor-signalling on expression of integrins such as integrin α5
and β1 (22,23). However, no alterations of integrin
α5 and β1 expressions were observed (data not shown). These results
suggest that the cell growth suppression of SCCs by the knockdown
or blockade of ET receptors is mediated through the direct
inhibition of MAPK signalling pathway.
Combination therapy of ETA or
ETB-siRNA and anti-tumor drugs
Reduction of dosage of anti-tumor drugs for cancer
chemotherapy is clinically important to minimize the side effects,
although the complete tumor cell death is required. Combined
treatment of ETB-siRNA (20 nM) with anti-tumor drug,
cisplatin (2.5 μM), drastically inhibited the cell growth of SAS in
comparison to that in each single treatment (Fig. 5A). Similar results were also
observed in the combined treatment of ETA-siRNA (20 nM)
with cisplatin (data not shown). These results indicate that
combination therapy of ETA or ETB-siRNA and
ordinal anti-tumor drugs may be a novel and useful therapy for
SCCs.
Discussion
There have been several reports on the expression of
ET receptors in various human cancers (5–7), and
it is considered to be the relationship between ET
receptor-signalling and tumor cell growth. There are, however, few
reports on the evaluation and investigation of the exact role of ET
receptor-signalling using human SCC tissues and cultured cell lines
of oral and esophageal carcinomas.
In the present study, using an immunohistochemical
method, we demonstrated significantly higher levels of expression
of ETA and ETB protein in human lingual
cancer tissues than in non-tumor areas in the same tissue samples.
Similar results were also observed on the cultured SCC cell lines
such as SAS, lingual SCC, and KYSE70, esophageal SCC. These results
indicate the involvement of ET receptor-signalling on SCC growth.
Furthermore, we showed that the suppression of ET receptor protein
by siRNA or the blockade by antagonists caused the inhibition of
SCC growth. In our experimental conditions, both the treatment with
ETA and ETB antagonists and siRNA strongly
inhibited the cell growth of SCCs. These results strongly suggest
the important role for ET receptor-signalling in SCC cell survival.
In fact, recent reports strongly indicated the involvement of ET
and its receptor on oral cancer (24,25).
In addition, it was also reported that suppression of
endothelin-converting enzyme-1 caused the inhibition of SCC
proliferation (26). Our results,
together with those reports, strongly suggest the importance of ET
synthesis and its receptor-signalling pathway on oral SCC
proliferation.
It is reported that phosphorylation of FAK is
involved in the inhibition of apoptosis and promote cell growth in
SCC cell lines (15,18). FAK is a 125 kDa non-receptor
tyrosine kinase and an important regulator of cell survival,
invasion, migration, and cell cycle progression (15,18,20,21).
FAK is functionally important in transducing intracellular messages
that are associated with growth factor signalling (15,18,20,21,27).
The intracellular messages link p-FAK at Tyr925 to
signalling pathways that activate MAPK cascades. In our present
study, however, the inhibition of phosphorylation of FAK in SCCs
treated with ET antagonists and siRNAs was not observed. In
contrast, the inhibition of the phosphorylation of MEK and Erk by
the treatment with ET antagonists and siRNAs was clearly observed.
These results indicate that the inhibition of MAPK pathway by the
suppression of ET receptor-signalling is due to the direct
inhibition of MAPK pathway, but not through FAK pathway (Fig. 5B). Several reports have indicated
the coupling of ET receptor-signalling and MAPK pathway (28,29).
Our results agree with those reports and indicate that the
mechanisms of the inhibition of cell growth by ET receptor-siRNAs
and antagonists are, in part, due to the inhibition of MAPK
pathway.
Reduction of dosage of anti-tumor drugs for cancer
chemotherapy is clinically important to minimize the side effects,
although the complete tumor cell death is required. Combined
treatment of low concentration of ET receptor-siRNA (20 nM) with
low concentration of anti-tumor drug, cisplatin (2.5 μM),
drastically inhibited the cell growth of SAS in comparison to that
in each single treatment. Cisplatin is extensively characterized as
DNA damaging agent and the cytotoxicity of cisplatin is attributed
to the ability to form inter and intra-strand nuclear DNA
crosslinks (30,31). In contrast, inhibition of cell
growth by ET receptor-siRNAs presented in our study was mainly due
to the direct inhibition of MAPK pathway. Therefore, those two
pathways on growth inhibition are different. This difference of
mechanisms between ET receptor-siRNA and cisplatin may lead to show
synergistic effect on the inhibition of tumor cell growth (Fig. 5A and B). Our results indicate that
the decrease in ET receptor levels in SCCs that strongly express ET
receptors increases the sensitivity against chemotherapy, and that
the siRNA for ET receptors combined with anti-tumor drugs might be
a useful therapy to reduce the dosage of anti-tumor drugs.
In summary, we showed the overexpression of
ETA and ETB in tumor cells of human primary
lingual SCC tissues and cultured SCC cell lines, and suggest a
potentially important role for ET receptor-signalling on the cell
growth of human SCCs.
Acknowledgements
This study was supported in part by grants (21592357
to K.W., 20390471 to Y.K., and 21390535 to M.K.) from the Japanese
Society for the Promotion of Science, and was supported by The
Osaka Medical Research foundation for Intractable Diseases.
References
|
1
|
Yanagisawa M, Kurihara H, Kimura S, et al:
A novel potent vasoconstrictor peptide produced by vascular
endothelial cells. Nature. 332:411–415. 1988.
|
|
2
|
Inoue A, Yanagisawa M, Kimura S, et al:
The human endothelin family: three structurally and
pharmacologically distinct isopeptides predicted by three separate
genes. Proc Natl Acad Sci USA. 86:2863–2867. 1989.
|
|
3
|
Davenport AP: International union of
pharmacology. XXIX. Update on endothelin receptor nomenclature.
Pharmacol Rev. 54:219–226. 2002.
|
|
4
|
Kusserow H and Unger T: Vasoacitive
peptides, their receptors and drug development. Basic Clin
Pharmacol Toxicol. 94:5–12. 2004.
|
|
5
|
Ahmed SI, Thompson J, Coulson JM and Woll
PJ: Studies on the expression of endothelin, its receptor subtypes,
and converting enzymes in lung cancer and in human bronchial
epithelium. Am J Respir Cell Mol Biol. 22:422–431. 2000.
|
|
6
|
Asham E, Shankar A, Loizidou M, et al:
Increased endothelin-1 in colorectal cancer and reduction of tumor
growth by ET(A) receptor antagonism. Br J Cancer. 85:1759–1763.
2001.
|
|
7
|
Zhang Y, Tang L, Su M, et al: Expression
of endothelines and their receptors in nonmelanoma skin cancers. J
Cutan Med Surg. 10:269–276. 2006.
|
|
8
|
Spinella F, Rosano L, DiCastro V, Natali
PG and Bagnato A: Endothelin-1 induces vascular endothelial growth
factor by increasing hypoxia-inducible factor-1 alpha in ovarian
carcinoma cells. J Biol Chem. 277:27850–27855. 2002.
|
|
9
|
Wulfing P, Kersting C, Tio J, et al:
Endothelin-1-, endothelin-A-, and endothelin-B-receptor expression
is correlated with vascular endothelial growth factor expression
and angiogenesis in breast cancer. Clin Cancer Res. 10:2393–2400.
2004.
|
|
10
|
Boldrin L, Gisfredi S, Ursino S, et al:
Expression of endothelin-1 is related to poor prognosis in
non-small cell lung carcinoma. Eur J Cancer. 41:2828–2835.
2005.
|
|
11
|
Kandalaft LE, Facciabene A, Buckanovich RJ
and Coukos G: Endothelin B receptor, a new target in cancer immune
therapy. Clin Cancer Res. 15:4521–4528. 2009.
|
|
12
|
Goepfert H: Squamous cell carcinoma of the
head and neck: past progress and future promise. CA Cancer J Clin.
48:195–198. 1998.
|
|
13
|
Okura M, Hiranuma T, Adachi T, et al:
Induction chemotherapy is associated with an increase in the
incidence of locoregional recurrence in patients with carcinoma of
the oral cavity: results from a single institution. Cancer.
82:804–815. 1998.
|
|
14
|
Prince S and Bailey BM: Squamous carcinoma
of the tongue: review. Br J Oral Maxillofac Surg. 37:164–174.
1999.
|
|
15
|
Masuda T, Wada K, Nakajima A, et al:
Critical role of peroxisome proliferator-activated receptor γ on
anoikis and invasion of squamous cell carcinomas. Clin Cancer Res.
11:4012–4021. 2005.
|
|
16
|
Takahashi H, Fujita K, Fujisawa T, et al:
Inhibition of peroxisome proliferator-activated receptor gamma
activity in esophageal carcinoma cells results in a drastic
decrease of invasive properties. Cancer Sci. 97:854–860. 2006.
|
|
17
|
Ishida H, Wada K, Masuda T, et al:
Critical role of estrogen receptor on anoikis and invasion of
squamous cell carcinoma. Cancer Sci. 98:636–643. 2007.
|
|
18
|
Nagata M, Wada K, Nakajima A, et al: Role
of myeloid cell leukemia-1 on cell growth of squamous cell
carcinoma. J Pharmacol Sci. 110:344–353. 2009.
|
|
19
|
Kusayama M, Wada K, Nagata M, et al:
Critical role of aquaporin 3 on growth of human esophageal and oral
squamous cell carcinoma. Cancer Sci. 102:1128–1136. 2011.
|
|
20
|
Chen HC, Appeddu PA, Parsons JT,
Hildebrand JD, Schaller MD and Guan JL: Interaction of focal
adhesion kinase with cytoskeletal protein talin. J Biol Chem.
270:16995–16999. 1995.
|
|
21
|
Sieg DJ, Hauck CR, Ilic D, et al: FAK
integrates growth-factor and integrin signals to promote cell
migration. Nat Cell Biol. 2:249–256. 2000.
|
|
22
|
Zhang Z, Vuori K, Reed JC and Ruoslahti E:
The alpha 5 beta 1 integrin supports survival of cells on
fibronectin and up-regulates Bcl-2 expression. Proc Natl Acad Sci
USA. 92:6161–6165. 1995.
|
|
23
|
Yamada KM and Geiger B: Molecular
interactions in cell adhesion complexes. Curr Opin Cell Biol.
9:76–85. 1997.
|
|
24
|
Hoffman RR, Yurgel LS and Campos MM:
Endothelins and their receptors as biological markers for oral
cancer. Oral Oncol. 46:644–647. 2010.
|
|
25
|
Hoffman RR, Yurgel LS and Campos MM:
Evaluation of salivary endothelin-1 levels in oral squamous cell
carcinoma and oral leukoplakia. Regulatory Peptide. 166:55–58.
2011.
|
|
26
|
Awano S, Dawson LA, Hunter AR, Turner AJ
and Usmani BA: Endothelin system in oral squamous carcinoma cells:
Specific siRNA targeting of ECE-1 blocks cell proliferation. Int J
Cancer. 118:1645–1652. 2006.
|
|
27
|
Frish SM and Francis H: Disruption of
epithelial cell-matrix interactions induces apoptosis. J Cell Boil.
124:619–626. 1994.
|
|
28
|
Bogoyevitch MA, Glennon PE, Andersson MB,
et al: Endothelin-1 and fibroblast growth factors stimulate the
mitogen-activated protein kinase signalling cascade in cardiac
myocytes. J Biol Chem. 269:1110–1119. 1994.
|
|
29
|
Cramer H, Schmenger K, Heinrich K, et al:
Coupling of endothelin receptors to the ERK/MAP kinase pathway. Eur
J Biochem. 268:5449–5459. 2001.
|
|
30
|
Meyers M, Hwang A, Wagner MW and Boothman
DA: Role of DNA mismatch repair in apoptotic responses to
therapeutic agents. Environ Mol Mutagen. 44:249–264. 2004.
|
|
31
|
Psyrri A and Fountzilas G: Advances in the
treatment of locally advanced non-nasopharyngeal squamous cell
carcinoma of the head and neck region. Med Oncol. 23:1–15.
2006.
|