Introduction
Global cancer burden was reported as 14.1 million
new cases in 2012; breast cancer is the most common cancer in women
worldwide, with nearly 1.7 million new cases were diagnosed in
2012. This represents ~12% of all new cancer cases and 25% of all
cancers in women (1).
Curcumin is the major bioactive ingredient extracted
from the rhizome of the plant Curcuma longa Linn (turmeric).
Curcumin has chemopreventive properties against various
malignancies, and extensive biological and pharmacological
function, for example, anticancer, antioxidant, anti-inflammatory,
antibacterial, antispasmodic and anticoagulant, without any major
side effects (2,3). As a pharmacologically safe agent,
curcumin could be used alone to prevent cancer and in combination
with chemotherapy to treat cancer. Different molecular targets are
modulated by curcumin. It induces apoptosis by means of the
inhibition of NF-κB activity through downregulation of upstream
kinases, IKK-β and IKK-α (4).
Paclitaxel is a microtubule-stabilizing agent
originally isolated from the bark of Taxus brevifolia. It is
a potent agent which inhibits neoplastic growth in several
malignancies including ovarian, breast, non-small cell lung cancer,
head and neck cancers (5–7). As with the majority of anticancer
agents, paclitaxel causes cancer cell death with signs of apoptosis
and also it is supposedly associated with mitotic catastrophe
(8,9).
c-Ha-ras is an oncoprotein corresponding to a
GTP-binding protein (10). DNA
transfection studies have suggested that an activated c-Ha-ras
oncogene can convert human breast cancer cells to a more
aggressive, estrogen-independent phenotype (11). Ras homolog gene family member A
(Rho-A) is a small (~22 kDa) G protein/guanosine triphosphatase
that is part of the Ras-related C3 botulinum toxin substrate (Rac)
subfamily of the Rho family (12).
Rho-A can reorganize the cell cytoskeleton and regulate cell
migration by activating effector proteins such as Rho-associated
coiled-coil kinase (ROCK) (13);
such changes are associated with tumor invasion and migration in
several types of carcinoma cells (14,15).
p53, the tumor suppressor protein plays a key role with respect to
apoptosis but also senescence, growth arrest and DNA repair
(16,17).
Apoptosis is a process of programmed cell death that
occurs in response to environmental stimuli and appropriated
strategy for prevention and treatment of cancer (18). Gene and protein expression of Bax
in breast cancer cells increase sensitivity to apoptotic stimuli
and decrease tumor growth. Apoptosis is a phenomenon usually linked
to the presence of functional p53 (19). Bcl-2 protein family plays an
important role in the survival or death of a cell and it is a
target of many antitumor drugs (20,21).
The Bcl-2 protein family, of which Bax is a member, plays a
critical role in determining either cell death or survival
(22,23). Bax is a pro-apoptotic protein that
acts as an enhancer of apoptosis while Bcl-2 has anti-apoptotic
properties as Bcl-xL that inhibits apoptosis and stimulates the
progress of breast cancer (20,24).
In particular, the balance between Bax (pro-apoptotic) and Bcl-2
(anti-apoptotic) protein levels is important for the regulation of
apoptosis (25). Overexpression of
Bax leads to apoptosis in the absence of any stimulus, suggesting
that tight regulation of Bax, from transcription to
post-translation, is necessary for cell survival (26). Anti-apoptotic Bcl-2 family members
are overexpressed in a variety of cancers through genetic
alterations, such as chromosomal translocation (Bcl-2) or
amplification (Bcl-xL and Mcl-1) (27–29).
The nuclear factor κ-light-chain-enhancer of
activated B cells (NF-κB) is a pro-inflammatory and pro-survival
transcription factor and it is known to be highly involved in the
initiation and progression of breast cancer (30). NF-κB deregulation of activity
alters the expression of cell death-regulating genes, leading to
the upregulation of anti-apoptotic and pro-survival genes, such as
members of the Bcl-2 family (31).
Protein inhibitors of κB (IκB) suppress activation of NF-κB
including IκBα, IκBβ, IκBɛ and IκBζ (32–40).
The aim of this study is to demonstrate the effect of curcumin and
paclitaxel on gene and protein expression related to apoptosis in
human breast cancer cell lines to provide new therapeutic potential
of curcumin combined with paclitaxel on human breast cancer and to
establish whether paclitaxel acquired chemoresistance in breast
cancer cells.
Materials and methods
Cell culture conditions
Human breast carcinoma cell lines MDA-MB-231
(ATCC® HTB26™) and MCF-7 (ATCC® HTB22™) were
maintained in a culture medium at 37°C in a humidified atmosphere
of 5% CO2 in air. The culture medium consisted of basic
medium supplemented with 10% fetal bovine serum (Life Technologies,
Grand Island, NY, USA). The basic medium was RPMI-1640 for
MDA-MB-231 and minimum essential media (Life Technologies) for
MCF-7 with 10 μg/ml of human insulin (Sigma-Aldrich, St. Louis, MO,
USA), both supplemented with 1% penicillin-streptomycin (Life
Technologies). Curcumin and paclitaxel were obtained from
Sigma-Aldrich. Both substances were dissolved in dimethylsulfoxide
(0.1%). Cells were seeded after 24 h prior to treatment with
curcumin (15 and 30 μM), paclitaxel (10–240 nM) and combination of
these two drugs. All the treatments were performed at 37°C. Cells
were harvested and seeded at 4×104 cells/ml of culture
medium into 24-well plates (Corning Costar, Corning, NY, USA).
After a 24-h pre-incubation period allowing cells to attach, the
culture medium was replaced by either culture medium without drugs
or medium with one of tested treatments of curcumin and/or
paclitaxel at desired concentrations for 48 h and it was replaced
by culture medium without drugs for 96 h. Cells were stained with
trypan blue (Sigma-Aldrich) with a 1:9 ratio of trypan blue for
cell suspension. Cells were counted in TC20™ Automated Cell Counter
(Bio-Rad Laboratories, Hercules, CA, USA). Dark blue cells were
evaluated as dead cells.
Doubling time (DT)
Cell growth was observed and counted at 0, 24, 48
and 96 h to determine doubling time (h). Upon treatment, both
control and treated cells were replated as described above. Their
growth was measured by determining the total number of doublings by
applying the formula: Total doublings: Td = ln2/μ where μ = ln
(N1/N0)/(t1-t0) where
N1 and N0 are the number of cells at times
t1 and t0. N1 is the total number
of cells counted at confluence and N0 is the initial
plating density. Total doublings were evaluated over multiple
passages. The number of doublings was determined by dividing the
total number of doublings by the number of days which each
particular sample was followed. Based upon these values, the time
required for the total cell number to double from any reference
point, or doubling time, was calculated (41).
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted with TRIzol (Invitrogen,
Carlsbad, CA, USA), and the concentration and purity of RNA were
determined using a UV spectrophotometer (Thermo Scientific,
Rochester, NY, USA). Total RNA was reverse transcribed into cDNA
using high capacity cDNA Reverse Transcription kit (Applied
Biosystems, Carlsbad, CA, USA) and 10 U of RNase inhibitor (Applied
Biosystems) according to the manufacturer’s protocol. A CFX 96
Touch Real-Time PCR Detection systems (Bio-Rad Laboratories) was
used with an aliquot of cDNA (2 μl) in 20 μl qPCR reaction
containing SYBR Green PCR Master Mix (Agilent, La Jolla, CA, USA)
for measurement of target genes such as c-Ha-ras, Rho-A, p53,
Bax, Bcl-xL, CCND1, NF-κB, IκBα, Stat-3, Bid, caspase-3 and
caspase-8; and β-actin as reference to obtain the
relative fold change for target genes using the comparative Ct
method and using Bio-Rad CFX Manager 2.1 software. Table I shows the primers for the genes
selected to develop cDNA probes. Relative expression was normalized
to the average of controls.
 | Table ISelected primers for target genes to
develop cDNA probes. |
Table I
Selected primers for target genes to
develop cDNA probes.
| Gene name | Product length
(bp)a | Primer
sequenceb |
|---|
|
c-Ha-ras | 112 | F:
CACCAGTACAGGGAGCAGAT
R: GAGCCTGCCGAGATTCCACA |
| Rho-A | 140 | F:
CCATCATCCTGGTTGGGAAT
R: CATGTACCCAAAAGCGCCA |
| p53 | 128 | F:
CCTCAGCATCTTATCCGAGTGG
R: TGGATGGTGGTACAGTCAGAGC |
| Bax | 143 | F:
GCGAGTGTCTCAAGCGCATC
R: CCAGTTGAAGTTGCCGTCAGAA |
| Bcl-xL | 211 | F:
CTGAATCGGAGATGGAGACC
R: TGGGATGTCAGGTCACTGAA |
| CCND1 | 60 | F:
GTGGCCTCTAAGATGAAGGA
R: GGTGTAGATGCACAGCTTCT |
| NF-κB | 114 | F:
ATCTGCCGAGTGAACCGAAACT
R: CCAGCCTGGTCCCGTGAAA |
| IκBα | 135 | F:
CTCCGAGACTTTCGAGGAAATAC
R: GCCATTGTAGTTGGTAGCCTTCA |
| Stat-3 | 163 | F:
GGTTGGACATGATGCACACTAT
R: AGGGCAGACTCAAGTTTATCAG |
| Bid | 199 | F:
GCTTCCAGTGTAGACGGAGC
R: GTGCAGATTCATGTGTGGATG |
|
Caspase-3 | 192 | F:
CAGAACTGGACTGTGGCATTG
R: GCTTGTCGGCATACTGTTTCA |
|
Caspase-8 | 128 | F:
CATCCAGTCACTTTGCCAGA
R: GCATCTGTTTCCCCATGTTT |
| β-actin | 569 | F:
ACTACCTCATGAAGATCCTC
R:TAGAAGCATTTGCGGTGGACGATGG |
Western blot analysis
Cells were lysed with 1 ml lysis buffer (pH 7.2)
[Tris Base (50 mM), NaCl (100 mM), EDTA (1 mM), Ortovanadate (1
mM), PMSF (1 mM), Triton X-100 (0.1%) and centrifuged (13,200 rpm ×
15 min)]. The supernatant with cellular proteins was dissolved in
SDS-PAGE sample solution (60 mM) Tris, pH 6.5, 10% (w/v) glycerol,
5% (w/v) β-mercaptoethanol, 20% (w/v) SDS, and 0.025% (w/v)
bromophenol blue and denatured by boiling (2×5 min), and vortex
mixing (2×30 seg). The total amount of protein was 30 μg in each
lane with standard protein markers (Bio-Rad Laboratories). After
fractionation by SDS-PAGE on gels (7×14 cm), proteins were
electro-blotted onto PVDF membrane (Amersham, Biosciences,
Amersham, UK) using a blotting apparatus (Bio-Rad Laboratories).
Prestained SDS-PAGE (Standards) blots were blocked for 2 h in 10%
defatted dry milk-TBS-0.1% Tween and then incubated for 2 h at room
temperature with corresponding primary antibodies [1:200, Bcl-2
(sc-7382), Bax (sc-7480), and β-actin (Sigma A-5316) followed by
incubation with secondary peroxidase-conjugated mouse IgG (1:5,000)
(Cell Signaling, CA, USA) in 5% defatted dry milk-TBS-0.1% Tween].
All steps were performed at room temperature, and blots were rinsed
between incubation steps with TBS-0.1% Tween. Cell blots were
probed with mouse anti-β-actin antibody as control. Immunoreactive
bands were visualized by using the ECL™ Western Blotting Detection
Reagent detection method (Amersham, Dübendorf, Switzerland) and
exposure of the membrane to X-ray film (Kodak, Rochester, NY, USA).
Protein determination was performed using the Bicinchoninic Acid
Method (Bio-Rad Laboratories) and BSA as the standards. Experiments
were performed in triplicates.
Statistical analysis
Data were expressed as the average ± standard error
of the mean (SEM). Comparisons of multiple groups were performed
between treated groups and controls carried out by ANOVA and
Dunnet’s test with GraphPad Prism 5 software. A P<0.05 was
considered to be significant. Assays were performed at least three
times independently.
Results
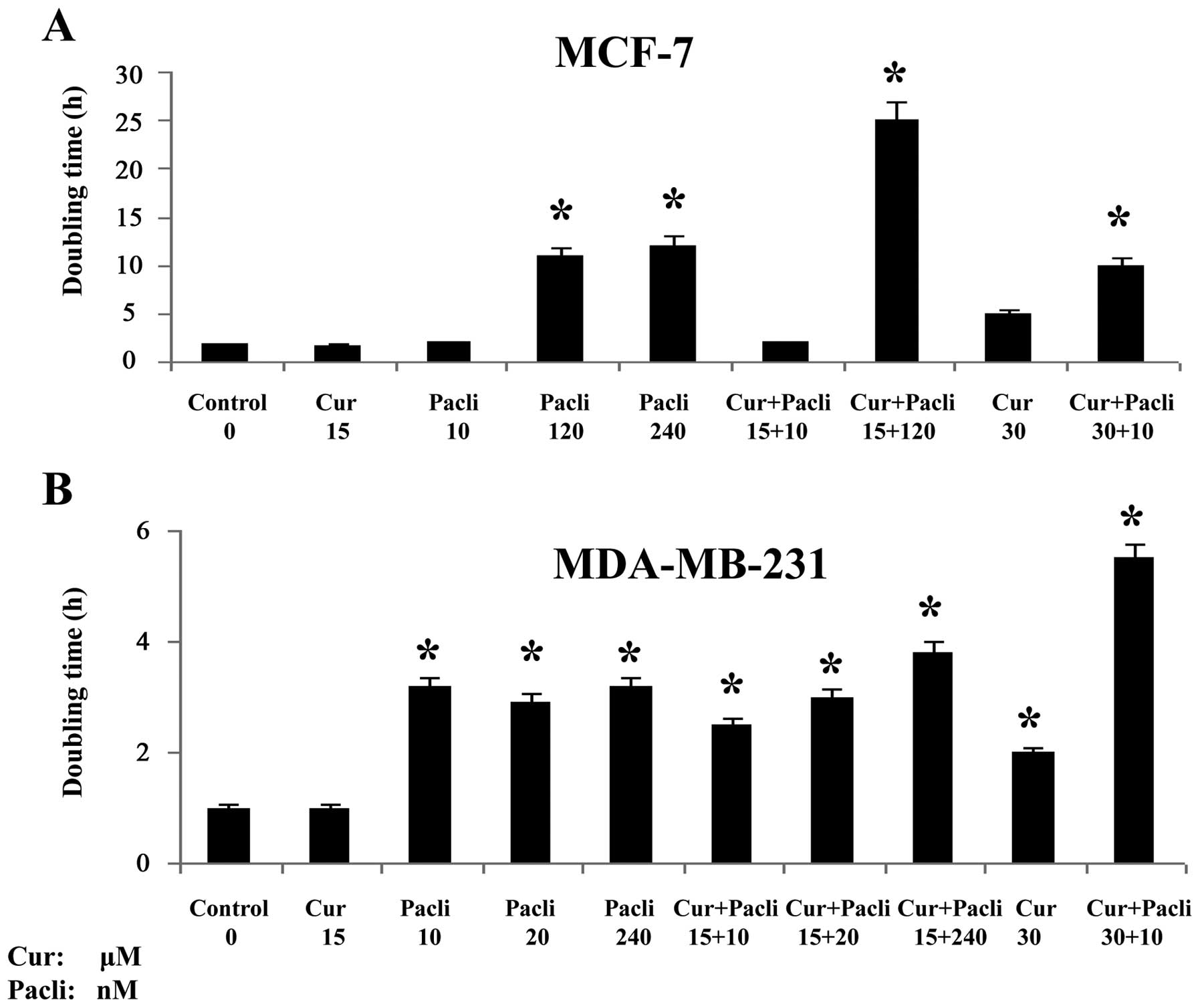
Effect of curcumin and paclitaxel on DT
of tumor cell lines
The DT of curcumin and paclitaxel was evaluated with
two cell lines that are either positive or negative for hormonal
receptors (ER+ PR+ and HER2+) as
the luminal MCF-7 and the basal-like MDA-MB-231, respectively.
Fig. 1 indicates that curcumin at
15 μM and paclitaxel at 120 nM had significantly (P<0.05) longer
DT than control and substances alone with other combinations in
MCF-7 cell line. On the other hand, these results showed that
MDA-MB-231 cell line had significantly (P<0.05) longer DT with
curcumin at 15 μM combined with paclitaxel at 10, 20 and 240 nM as
well as curcumin at 30 μM with paclitaxel at 10 nM than control and
curcumin at 15 μM alone. Therefore, these combinations were used
for determining several important gene targets in malignancy and
apoptosis. All the doses used significantly decreased the rate of
growth of cells above its controls as indicated by the DT after
several passages.
Effect of curcumin and paclitaxel on gene
expression in MCF-7 breast cancer cells
Gene expression analyzed by RT-qPCR indicated that
curcumin combined (15 μM) with paclitaxel (120 nM) significantly
(P<0.05) decreased c-Ha-Ras, Rho-A, p53 and Bcl-xL
gene expression in comparison to control and substances alone in
MCF-7 cells as seen in Fig. 2A–D.
Gene expression indicated that curcumin (15 μM) and paclitaxel (120
nM) alone significantly (P<0.05) increased Bax expression
in comparison to control and substances alone in MCF-7 as seen in
Fig. 2D.
Curcumin and paclitaxel alone and both combined
significantly (P<0.05) decreased CCND1 (Fig. 3A) and NF-κB (Fig. 3B), whereas curcumin alone decreased
IκBα and Stat-3 gene expression (Fig. 3C) and paclitaxel alone and combined
increased them (Fig. 3D) in MCF-7
cells in comparison to control.
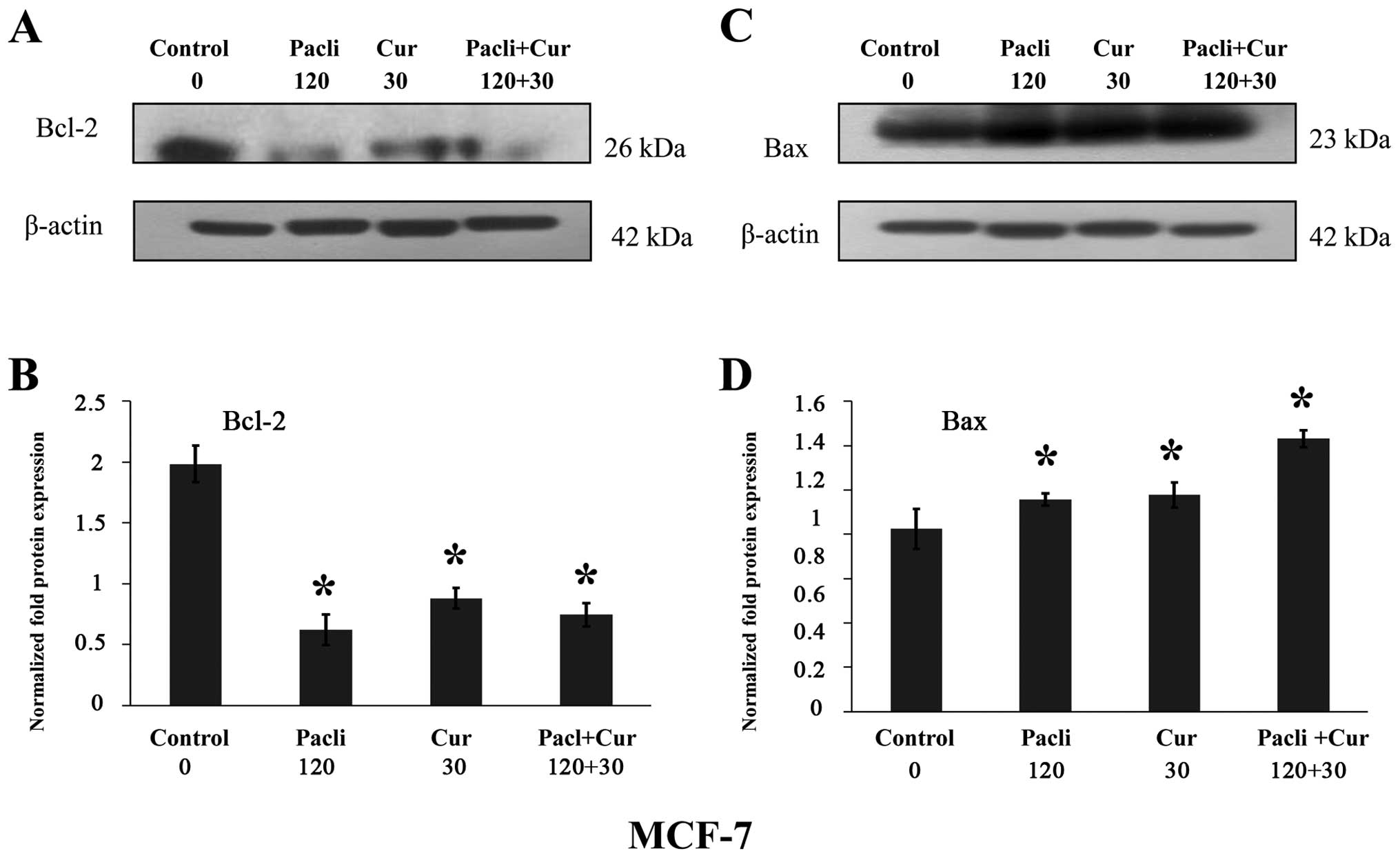
Effect of curcumin and paclitaxel on
protein expression by western blot analysis in MCF-7 breast cancer
cell line
Results indicated that either paclitaxel at 120 nM
or curcumin at 30 μM alone and combined (Fig. 4A and B) significantly decreased
Bcl-2 and increased Bax protein expression (Fig. 4C and D) in MCF-7, in comparison to
control corroborating the apoptotic effect of these substances.
Normalized fold protein expression is presented in graphs of
Fig. 4B and D.
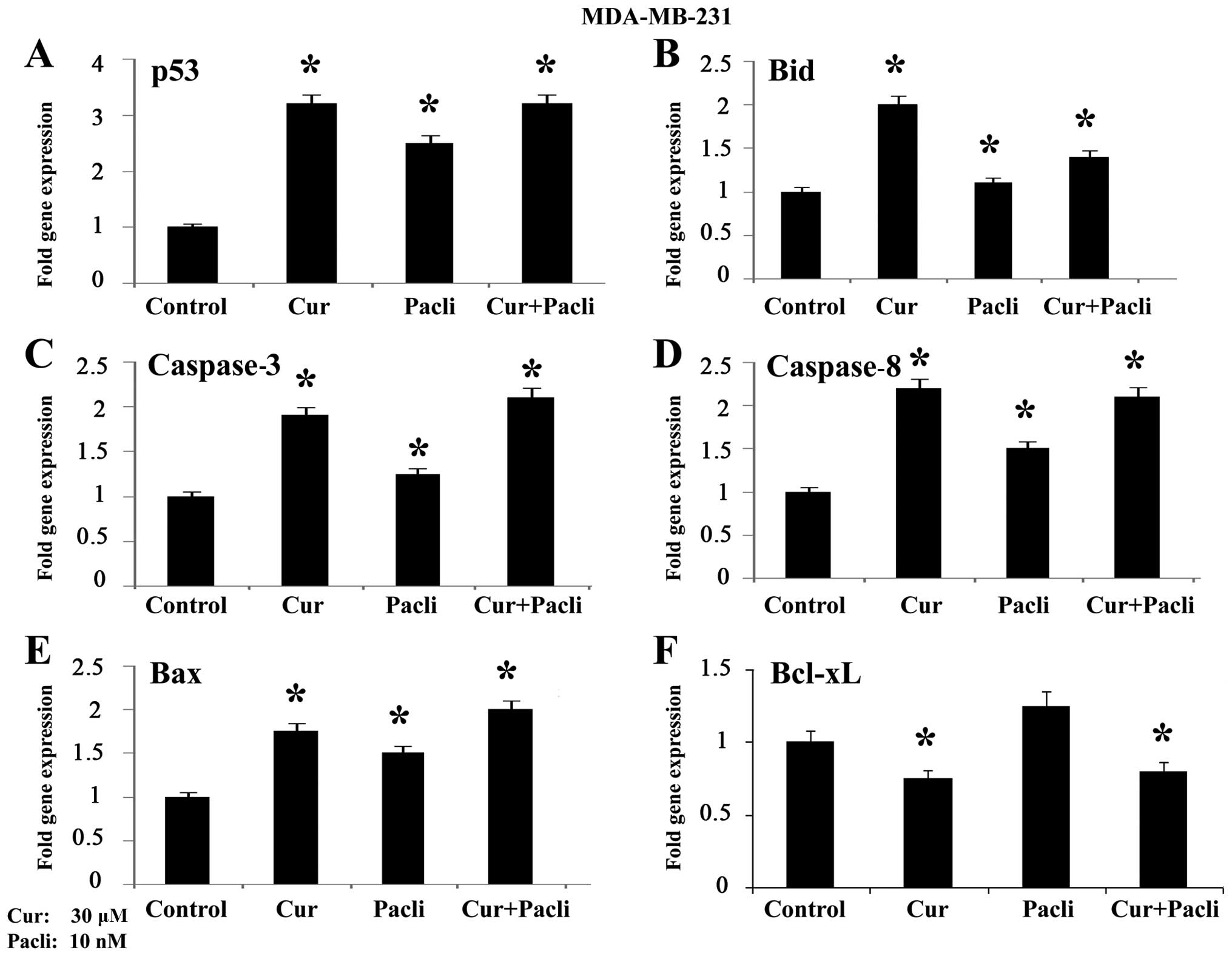
Effect of curcumin and paclitaxel on gene
expression in MDA-MB-231 breast cancer cell line
Gene expression analyzed by RT-qPCR in MDA-MB-231
cells is shown in Fig. 5 where
results indicate that curcumin at 30 μM alone and combined with
paclitaxel at 10 nM significantly (P<0.05) increased p53
(Fig. 5A), Bid (Fig. 5B), caspase-3 (Fig. 5C), caspase-8 (Fig. 5D) and Bax (Fig. 5E) gene expression, whereas Bcl-xL
decreased such expression (Fig.
5F) in comparison to control.
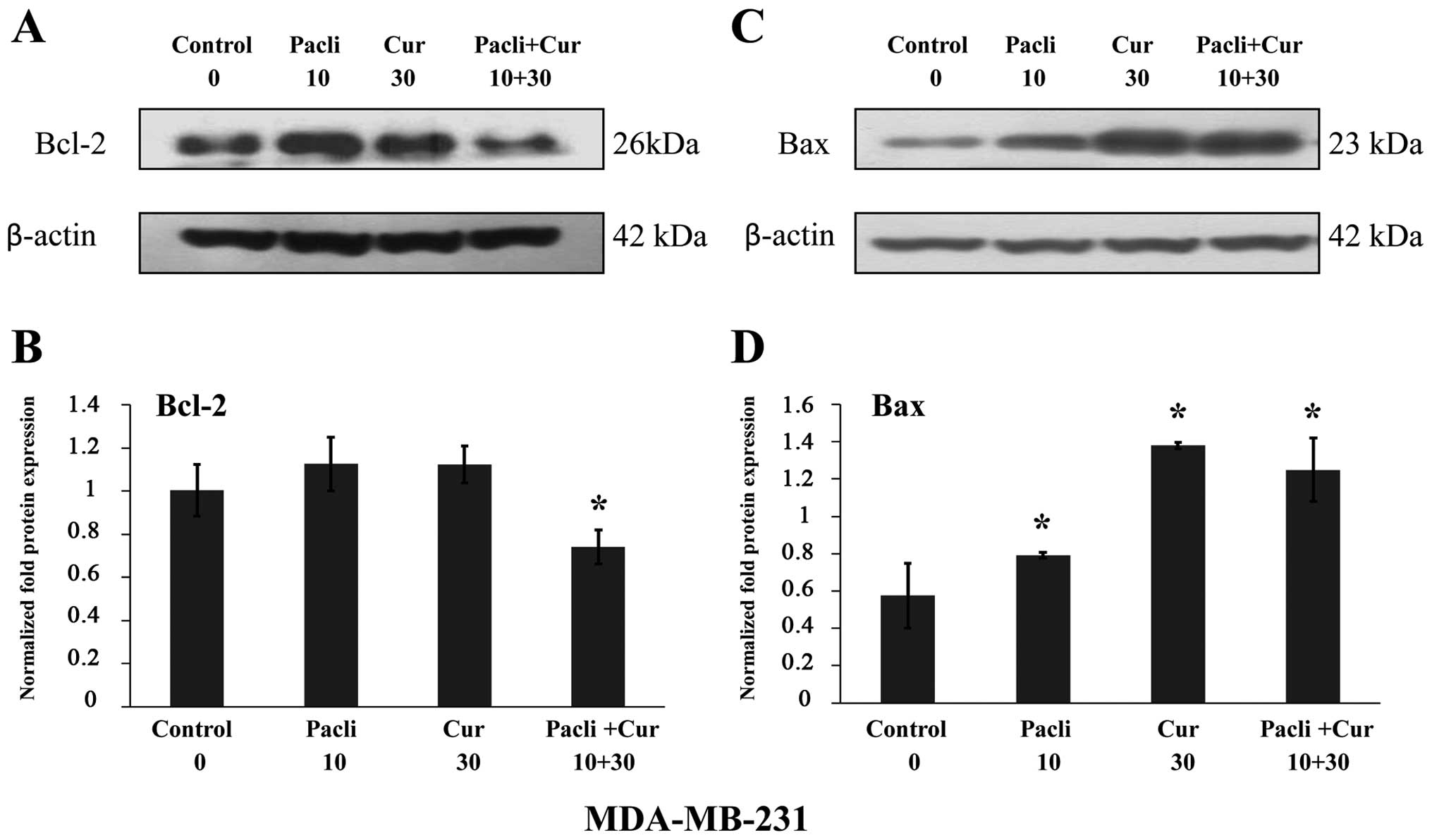
Effect of curcumin and paclitaxel on
protein expression by western blot analysis in MDA-MB-231 breast
cancer cell line
Results indicated that paclitaxel at 10 nM and
curcumin at 30 μM combined significantly (P<0.05) decreased
Bcl-2 protein expression in comparison to control and either
substance alone. However, either substance alone and combined
increased Bax protein expression in MDA-MB-231 corroborating the
apoptotic effect of these substances (Fig. 6A and C). Normalized fold protein
expression is presented in graphs of Fig. 6B and D.
Discussion
Chemotherapy in breast cancer fails due to
resistance to drugs. Chemotherapeutic strategies are now making use
of combined active compounds where they are believed to be more
active as compared to the single agent itself. Therefore, the
efficacy of treatment would increase and the possibility of toxic
effect might be lowered due to the extremely low usage of drug
(42). A phytochemical such as
curcumin can be used in a therapeutic modality as it elicits
antitumor effects. Curcumin is a natural compound derived from
turmeric (Curcuma longa) and exhibits an antitumorigenic
effect on various cancers. Curcumin and paclitaxel effect was
evaluated with the luminal MCF-7 and the basal-like MDA-MB-231
cells, respectively, these cell lines are positive and negative for
hormonal receptors (ER+ PR+ and
HER2+), respectively.
MCF-7 cells had longer DT when paclitaxel was
combined with curcumin at low doses than control, indicating that a
therapeutic modality like this would benefit the response of
patients to treatment. Similar results were observed with
MDA-MB-231 cells with low and high doses of paclitaxel in
combination with low and high doses of curcumin. All the doses used
significantly decreased the rate of growth of cells above its
controls as indicated after several passages. Since growth
inhibitory effect from combining paclitaxel with curcumin was more
effective than either agent alone it seems that curcumin
potentiates the apoptotic effects of paclitaxel in MCF-7 and
MDA-MB-231 cell lines.
Curcumin combined with paclitaxel decreased
c-Ha-Ras, Rho-A and p53 gene expression in comparison
to control and substances alone in MCF-7 cell line. This is the
first report that demonstrated that an antioxidant as curcumin and
a chemo-therapeutic drug reduces c-Ha-Ras, Rho-A and
p53 gene expression. Gene expression levels supported the
idea that paclitaxel induces changes in important signaling
pathways.
Studies have indicated that curcumin is a
pharmacologically safe compound, it has a therapeutic potential in
preventing breast cancer metastasis possibly through suppression of
NF-κB (43). Since curcumin
suppresses NF-κB activation and most chemotherapeutic agents
activate genes that mediates cell survival, proliferation,
invasion, and metastasis (43), we
analyzed whether curcumin would potentiate the effect of
chemotherapy in breast cancer cell lines. Thus, curcumin decreased
expression of NF-κB supporting the conclusion of other reports
(44) that the mechanism of growth
inhibition induced by combined effect of paclitaxel and curcumin is
through NF-κB inhibition. It was demonstrated that curcumin
suppressed the paclitaxel-induced NF-κB pathway in breast cancer
cells by inhibiting lung metastasis of human breast cancer in nude
mice (43).
Curcumin combined with paclitaxel decreased Bcl-xL
gene expression in comparison to control and either substance alone
in MCF-7 cell line. Other authors reported on the capability of
inducing apoptosis in MCF-7 by several drugs (45). On the other hand, either substance
alone increased Bax expression in comparison to control and either
substance alone in MCF-7 cells. Curcumin, paclitaxel and
combination of both decreased CCND1, whereas either substance alone
decreased IκBα and Stat-3 gene expression in MCF-7 cells in
comparison to control. Results indicated that either substance
alone as curcumin and paclitaxel as well as combined decreased
Bcl-2 and increased Bax protein expression in comparison to control
in MCF-7 cells, corroborating the apoptotic effect of these
substances. It has also been suggested that inhibition of activity
of proteins may improve the efficacy of chemotherapeutic agents
(44).
It has been shown that curcumin induced apoptosis in
breast cancer cells and delayed the outgrowth of mammary tumors in
neu transgenic mice (46). The
combination of docosahexaenoic acid and curcumin inhibited
7,12-dimethylbenz(a)anthracene (DMBA)-induced mammary tumorigenesis
in mice (47). In addition,
curcumin can reverse multidrug resistance in human colon carcinomas
and lung cancer cells in vitro and in vivo (48,49).
Authors have shown that curcumin can inhibit epithelial mesenchymal
transition by affecting transcription factors as E-cadherin and
Vimentin genes and invasive capabilities in breast cancer cell
lines (50,51).
When MDA-MB-231 cell line was analyzed it was found
that curcumin alone and combined with paclitaxel increased p53,
Bid, caspase-3, caspase-8 and Bax gene expression,
whereas Bcl-xL decreased such expression in comparison to
control. Results indicated that paclitaxel and curcumin combined
decreased Bcl-2 protein expression in comparison to control and
either substance alone. However, either substance alone and
combined increased Bax protein expression in MDA-MB-231
corroborating the apoptotic effect of these substances. Curcumin
decreased expression of apoptotic genes such as caspase-3,
caspase-8, and other genes such as cyclin D1.
We demonstrated that curcumin increased the
sensitivity of MCF-7 and MDA-MB-231 cells to chemotherapeutic drugs
as paclitaxel. It has been proposed that combined effect of natural
products may improve the effectiveness of treatment in the process
of proliferation of cancer cells (42).
Several studies have aimed to delineate the complex
molecular background of breast cancer, which has advanced
personalized treatment approaches and enabled the development of
several agents that target specific molecular aberrations
associated with breast cancer. Nevertheless, tumor heterogeneity
poses obvious impediments to the successful clinical development of
targeted agents. The combined anticancer activities of a compound
of natural origin such as curcumin has been examined in combination
with a chemotherapy drug such as paclitaxel and it was observed
that paclitaxel-curcumin compound exhibited synergistic growth
inhibition and induced significant apoptosis in MCF-7 and
MDA-MB-231 cell lines. In conclusion, curcumin may be of
considerable value in synergistic therapy of cancer since it may
reduce the associated toxicity and it has a therapeutic potential
with a drug such as paclitaxel in breast cancer.
Acknowledgements
The technical support of Guiliana Rojas, Georgina
Vargas and secretarial assistance and suggestions of Leodán A.
Crispin and Richard Ponce-Cusi are greatly appreciated. This study
was supported by grant support FONDECYT no. 1120006 (G.M.C.) and
Ministerio de Educación de Chile - Convenio de Desempeño
(UTA-GMC).
References
|
1
|
Ferlay J, Steliarova-Foucher E,
Lortet-Tieulent J, Rosso S, Coebergh JW, Comber H, Forman D and
Bray F: Cancer incidence and mortality patterns in Europe:
Estimates for 40 countries in 2012. Eur J Cancer. 49:1374–1403.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Calaf GM, Echiburú-Chau C, Roy D, Chai Y,
Wen G and Balajee AS: Protective role of curcumin in oxidative
stress of breast cells. Oncol Rep. 26:1029–1035. 2011.PubMed/NCBI
|
|
3
|
Calaf GM, Echiburú-Chau C, Wen G, Balajee
AS and Roy D: Effect of curcumin on irradiated and
estrogen-transformed human breast cell lines. Int J Oncol.
40:436–442. 2012.
|
|
4
|
Joe B, Vijaykumar M and Lokesh BR:
Biological properties of curcumin-cellular and molecular mechanisms
of action. Crit Rev Food Sci Nutr. 44:97–111. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gabizon A, Price DC, Huberty J, Bresalier
RS and Papahadjopoulos D: Effect of liposome composition and other
factors on the targeting of liposomes to experimental tumors:
Biodistribution and imaging studies. Cancer Res. 50:6371–6378.
1990.PubMed/NCBI
|
|
6
|
Fu Y, Li S, Zu Y, Yang G, Yang Z, Luo M,
Jiang S, Wink M and Efferth T: Medicinal chemistry of paclitaxel
and its analogues. Curr Med Chem. 16:3966–3985. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okano J and Rustgi AK: Paclitaxel induces
prolonged activation of the Ras/MEK/ERK pathway independently of
activating the programmed cell death machinery. J Biol Chem.
276:19555–19564. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
McGrogan BT, Gilmartin B, Carney DN and
McCann A: Taxanes, microtubules and chemoresistant breast cancer.
Biochim Biophys Acta. 1785:96–132. 2008.
|
|
9
|
Jin C, Wu H, Liu J, Bai L and Guo G: The
effect of paclitaxel-loaded nanoparticles with radiation on hypoxic
MCF-7 cells. J Clin Pharm Ther. 32:41–47. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barbacid M: ras genes. Annu Rev Biochem.
56:779–827. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kasid A, Lippman ME, Papageorge AG, Lowy
DR and Gelmann EP: Transfection of v-rasH DNA into MCF-7 human
breast cancer cells bypasses dependence on estrogen for
tumorigenicity. Science. 228:725–728. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu LLG and Gu JY: Advances in the role of
Rho sub-family in tumor invasion. Fudan Univ J Med Sci. 37:617–619.
2010.
|
|
13
|
Hall A: Rho GTPases and the actin
cytoskeleton. Science. 279:509–514. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
DerMardirossian C and Bokoch GM: GDIs:
Central regulatory molecules in Rho GTPase activation. Trends Cell
Biol. 15:356–363. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Olofsson B: Rho guanine dissociation
inhibitors: Pivotal molecules in cellular signalling. Cell Signal.
11:545–554. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schmitt CA, Fridman JS, Yang M, Lee S,
Baranov E, Hoffman RM and Lowe SW: A senescence program controlled
by p53 and p16INK4a contributes to the outcome of cancer therapy.
Cell. 109:335–346. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vogelstein B, Lane D and Levine AJ:
Surfing the p53 network. Nature. 408:307–310. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yonish-Rouach E, Grunwald D, Wilder S,
Kimchi A, May E, Lawrence JJ, May P and Oren M: p53-mediated cell
death: Relationship to cell cycle control. Mol Cell Biol.
13:1415–1423. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miyashita T, Krajewski S, Krajewska M,
Wang HG, Lin HK, Liebermann DA, Hoffman B and Reed JC: Tumor
suppressor p53 is a regulator of bcl-2 and bax gene expression in
vitro and in vivo. Oncogene. 9:1799–1805. 1994.PubMed/NCBI
|
|
21
|
Miyashita T and Reed JC: Tumor suppressor
p53 is a direct transcriptional activator of the human bax gene.
Cell. 80:293–299. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
García-Sáez AJ: The secrets of the Bcl-2
family. Cell Death Differ. 19:1733–1740. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
González de Aguilar JL, Gordon JW, René F,
de Tapia M, Lutz-Bucher B, Gaiddon C and Loeffler JP: Alteration of
the Bcl-x/Bax ratio in a transgenic mouse model of amyotrophic
lateral sclerosis: Evidence for the implication of the p53
signaling pathway. Neurobiol Dis. 7:406–415. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Delft MF and Huang DC: How the Bcl-2
family of proteins interact to regulate apoptosis. Cell Res.
16:203–213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jürgensmeier JM, Xie Z, Deveraux Q,
Ellerby L, Bredesen D and Reed JC: Bax directly induces release of
cytochrome c from isolated mitochondria. Proc Natl Acad Sci USA.
95:4997–5002. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Placzek WJ, Wei J, Kitada S, Zhai D, Reed
JC and Pellecchia M: A survey of the anti-apoptotic Bcl-2 subfamily
expression in cancer types provides a platform to predict the
efficacy of Bcl-2 antagonists in cancer therapy. Cell Death Dis.
1:e402010. View Article : Google Scholar
|
|
28
|
Davids MS and Letai A: Targeting the
B-cell lymphoma/leukemia 2 family in cancer. J Clin Oncol.
30:3127–3135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Walensky LD: From mitochondrial biology to
magic bullet: Navitoclax disarms BCL-2 in chronic lymphocytic
leukemia. J Clin Oncol. 30:554–557. 2012. View Article : Google Scholar
|
|
30
|
Dai Y, Lawrence TS and Xu L: Overcoming
cancer therapy resistance by targeting inhibitors of apoptosis
proteins and nuclear factor-kappa B. Am J Transl Res. 1:1–15.
2009.PubMed/NCBI
|
|
31
|
Han J, Sun M, Cui Y, Wang T, Zhang W, Guo
M, Zhou Y, Liu W, Zhang M, Duan J, et al: Kushen flavonoids induce
apoptosis in tumor cells by inhibition of NF-kappaB activation and
multiple receptor tyrosine kinase activities. Phytother Res.
21:262–268. 2007. View
Article : Google Scholar
|
|
32
|
Wulczyn FG, Naumann M and Scheidereit C:
Candidate proto-oncogene bcl-3 encodes a subunit-specific inhibitor
of transcription factor NF-kappa B. Nature. 358:597–599. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bours V, Franzoso G, Azarenko V, Park S,
Kanno T, Brown K and Siebenlist U: The oncoprotein Bcl-3 directly
transactivates through kappa B motifs via association with
DNA-binding p50B homodimers. Cell. 72:729–739. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dobrzanski P, Ryseck RP and Bravo R:
Differential interactions of Rel-NF-kappa B complexes with I kappa
B alpha determine pools of constitutive and inducible NF-kappa B
activity. EMBO J. 13:4608–4616. 1994.PubMed/NCBI
|
|
35
|
Baldwin AS Jr: The NF-kappa B and I kappa
B proteins: New discoveries and insights. Annu Rev Immunol.
14:649–683. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cheng JD, Ryseck RP, Attar RM, Dambach D
and Bravo R: Functional redundancy of the nuclear factor kappa B
inhibitors I kappa B alpha and I kappa B beta. J Exp Med.
188:1055–1062. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huxford T, Huang DB, Malek S and Ghosh G:
The crystal structure of the IkappaBalpha/NF-kappaB complex reveals
mechanisms of NF-kappaB inactivation. Cell. 95:759–770. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tam WF and Sen R: IkappaB family members
function by different mechanisms. J Biol Chem. 276:7701–7704. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hoffmann A, Levchenko A, Scott ML and
Baltimore D: The IkappaB-NF-kappaB signaling module: Temporal
control and selective gene activation. Science. 298:1241–1245.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Anest V, Hanson JL, Cogswell PC,
Steinbrecher KA, Strahl BD and Baldwin AS: A nucleosomal function
for IkappaB kinase-alpha in NF-kappaB-dependent gene expression.
Nature. 423:659–663. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Calaf G and Russo J: Transformation of
human breast epithelial cells by chemical carcinogens.
Carcinogenesis. 14:483–492. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhan Y, Chen Y, Liu R, Zhang H and Zhang
Y: Potentiation of paclitaxel activity by curcumin in human breast
cancer cell by modulating apoptosis and inhibiting EGFR signaling.
Arch Pharm Res. 37:1086–1095. 2014. View Article : Google Scholar
|
|
43
|
Aggarwal BB, Shishodia S, Takada Y,
Banerjee S, Newman RA, Bueso-Ramos CE and Price JE: Curcumin
suppresses the paclitaxel-induced nuclear factor-kappaB pathway in
breast cancer cells and inhibits lung metastasis of human breast
cancer in nude mice. Clin Cancer Res. 11:7490–7498. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kang HJ, Lee SH, Price JE and Kim LS:
Curcumin suppresses the paclitaxel-induced nuclear factor-kappaB in
breast cancer cells and potentiates the growth inhibitory effect of
paclitaxel in a breast cancer nude mice model. Breast J.
15:223–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Choudhuri T, Pal S, Agwarwal ML, Das T and
Sa G: Curcumin induces apoptosis in human breast cancer cells
through p53-dependent Bax induction. FEBS Lett. 512:334–340. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Masuelli L, Benvenuto M, Fantini M,
Marzocchella L, Sacchetti P, Di Stefano E, Tresoldi I, Izzi V,
Bernardini R, Palumbo C, et al: Curcumin induces apoptosis in
breast cancer cell lines and delays the growth of mammary tumors in
neu transgenic mice. J Biol Regul Homeost Agents. 27:105–119.
2013.PubMed/NCBI
|
|
47
|
Siddiqui RA, Harvey KA, Walker C,
Altenburg J, Xu Z, Terry C, Camarillo I, Jones-Hall Y and Mariash
C: Characterization of synergistic anti-cancer effects of
docosahexaenoic acid and curcumin on DMBA-induced mammary
tumorigenesis in mice. BMC Cancer. 13:4182013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lu WD, Qin Y, Yang C, Li L and Fu ZX:
Effect of curcumin on human colon cancer multidrug resistance in
vitro and in vivo. Clinics (Sao Paulo). 68:694–701. 2013.
View Article : Google Scholar
|
|
49
|
Thiyagarajan S, Thirumalai K, Nirmala S,
Biswas J and Krishnakumar S: Effect of curcumin on lung
resistance-related protein (LRP) in retinoblastoma cells. Curr Eye
Res. 34:845–851. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gallardo M and Calaf GM: Curcumin and
epithelial-mesenchymal transition in breast cancer cells
transformed by low doses of radiation and estrogen. Int J Oncol.
48:2534–2542. 2016.PubMed/NCBI
|
|
51
|
Gallardo M and Calaf GM: Curcumin inhibits
invasive capabilities through epithelial mesenchymal transition in
breast cancer cell lines. Int J Oncol. 49:1019–1027.
2016.PubMed/NCBI
|