Introduction
Osteosarcoma is the most common malignant bone tumor
that occurs primarily in children and adolescents. Despite surgery
combined with chemotherapy has been widely applied for osteosarcoma
treatment, an overwhelming tendency for invasion and early
metastasis lead to poor prognosis. The 5-year survival rate of
patients with recur rent and metastasis remains at only 20%
(1). The mechanisms for
osteosarcoma development has drawn great attention (2,3).
However, due to lack of effective drug targets, a breakthrough for
osteosarcoma treatment has not yet been achieved.
As a well-documented driver of cell cycle, E2F1
regulates the expression of genes required in the G1→S phase
transition (4). However, recent
studies showed a role of E2F1 in metastasis and invasion of several
cancer types. E2F1-induced metastasis was verified by clinical
studies showing that high expression of E2F1 and its related target
genes predicted the transformation from superficial to invasive
bladder cancer (5). In addition,
it was demonstrated that the depletion of endogenous E2F1
attenuated tumor invasion and pulmonary metastasis in
therapy-resistant metastatic melanoma xenografts (6). Recently, Liang et al reported
a novel function of E2F1 as enhancer of tumor invasion and
migration in prostate cancer via regulating the expression of CD147
(7). However, the mechanisms
underlying the metastasis promoted by E2F1 remain unclear.
DDR1 belongs to discoidin domain receptor (DDR)
family which consists of two highly homologous members, DDR1 and
DDR2. DDRs function as the unique receptor tyrosine kinases (RTKs)
which are activated by collagen, a major extracellular matrix (ECM)
component (8). Previous studies
have reported abnormal expression of DDR1 in some high-invasive
tumors, such as lung, breast, and prostate cancers (9–11).
Growing evidence suggested that DDR1 was closely associated with
lymph node metastasis and shorter survival, and overexpression of
DDR1 promoted cell mobility and invasion (12,13).
Although DDR1 has been verified to be involved in tumor
development, dysregulation of DDR1 as well as the roles of DDR1 in
tumor aggressiveness is poorly understood.
In the present study, E2F1 was found to be critical
for the migration and invasion of osteosarcoma cells through
transactivating DDR1. Furthermore, DDR1 enhances the
phosphorylation of STAT3 which drives the epithelial-mesenchymal
transition (EMT) of osteosarcoma cells. Finally, in vivo
experiments and clinical analysis confirm that E2F1 and DDR1 are
important for maintaining the malignant features of osteosarcoma.
Together, these findings suggest a novel mechanism for
E2F1-dependent migration and invasion in osteosarcoma, and provide
a new understanding of E2F1-driven tumor progression.
Materials and methods
Cell culture and transfection
Two osteosarcoma cell lines, U2OS (TP53-WT; RB1-WT)
and SaOs-2 (TP53-Mut; RB1-Mut), and an immortalized osteoblast cell
line hFOB1.19 (TP53-WT; RB1-WT) were purchased from the Institute
of Biochemistry and Cell Biology of the Chinese Academy of Sciences
(Shanghai, China) and grown in Dulbeco's modified Eagle's medium
supplemented with 10% fetal bovine serum, and the cells were
incubated at 37°C and 5% CO2. Before experiments, cells
were cultured for 3–5 passages. Cell transfection was performed
using Lipofectamine 3000 reagent (Thermo Fisher Scientific,
Waltham, MA, USA) according to the manufacturer's instructions. The
U2OS and SaOs-2 cells with stable knockdown of E2F1 or DDR1 were
established with indicated shRNA and maintained by G418 (Sigma, St.
Louis, MO, USA).
Antibodies and reagents
Antibodies against E2F1 (sc-251), DDR1 (sc-532),
E-cadherin (sc-8426), vimentin (sc-6260), MMP2 (sc-10736), MMP9
(sc-10737), and GAPDH (sc-32233) were from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). The p-AKT (S473) (#4051), AKT (#2966),
p-STAT3 (Y705) (#4113), and STAT3 (#9139) antibodies were from Cell
Signaling Technology (Beverly, MA, USA). The siRNAs targeting STAT3
were from Santa Cruz.
Western blotting
Cells were lysed in RIPA buffer supplemented with
PMSF, and the protein lysates were separated by SDS-PAGE and
transferred to nitrocellulose membranes (Whatman, Maidstone, UK).
Then the membranes were incubated overnight at 4°C by using primary
antibodies. After incubation with goat anti-rabbit (926-32211;
926-68071) or goat anti-mouse (926-32210; 926-68070) IgG secondary
antibodies (1:10,000; LI-COR, Lincoln, NE, USA) at room temperature
for 1 h, the fluorescence intensity was visualized by the Odyssey
Infrared Imaging system (LI-COR).
Cell viability assay
Cell viability was detected using the Cell Counting
Kit-8 assay (Dojindo, Kumamoto, Japan). Transfected cells were
dispensed in triplicate into 96-well plates and incubated overnight
at 37°C. After indicated time, 10 µl of CCK-8 kit solution
was added to the cells, which were then incubated for 2.5 h at
37°C. Absorbance was then measured by a microplate reader at 450
nm. Data were obtained from at least three separate experiments
done in triplicate.
Quantitative real-time PCR analysis
Total RNA was prepared using the RNAiso Plus
(Takara, Kyoto, Japan). cDNA was synthesized from total RNA using
PrimeScript™ RT-PCR kit (RR014B; Takara). Quantitative real-time
PCR (qRT-PCR) was performed by the 7500 Real-Time PCR system
(Applied Biosystems, Foster City, CA, USA) with GAPDH being used as
an internal control. The results are representative of at least
three independent experiments.
RNA-seq analysis
RNA from U2OS cells with or without E2F1 knockdown
was extracted and purified for quantification, RNA-seq library
preparation, and sequencing. The libraries were sequenced on the
illumina HiSeq 2500 platform. The reads containing adapter or
poly-N and reads of low quality were removed from raw data to
generate clean reads for further analyses. Based on the clean
reads, the Q20 (>90%), Q30 (>85%), and error rate (<0.1%)
of the clean data were required. Then mapped reads were obtained by
Tophat2 through aligning clean reads to the human genome reference
(hg19). The number of mapped clean reads for each unigene was
counted and normalized into a reads per kb per million reads (RPKM)
to calculate the expression level of the unigene.
Luciferase reporter assay
Various lengths of DDR1 promoter were amplified by
PCR and inserted into the pGL3 vector. The reporter construct with
a mutated E2F1 binding site was generated by subsequent PCR-based
cloning. These reporter plasmids were transfected into osteosarcoma
cells, and pRL-TK was co-transfected as an internal control for
transfection efficiency. At 48 h post-transfection, cells were
lysed and the luciferase activities were measured using the
Dual-Luciferase Reporter assay system (Promega, Madison, WI, USA).
The experiments were carried out in triplicate.
Chromatin immunoprecipitation
Cells were cross-linked with 1% formaldehyde, lysed,
and then sheared to an average size of 250–500 bp by sonication.
ChIP was performed by incubating chromatin with anti-E2F1 antibody.
After that, chromatin-antibody complexes were isolated using
Protein A/G Plus-Agarose (Santa Cruz). Finally, the crosslinking
was reversed and immunoprecipitated DNA was purified. Enrichment of
DNA fragments was analyzed by qRT-PCR. Three independent
experiments were performed.
Migration assay
A scratch wound healing assay was used for
determining cell migration. Briefly, cell monolayer was scratched
with a sterile pipette. The culture was then washed with medium to
remove free-floating cells and debris and cultured in serum-free
medium for an additional 24 h. The coverage of the scratched area
was measured at three different positions. All experiments were
performed in triplicates.
Invasion assay
Transfected cells in serum-free media were placed
into the upper chamber of an insert coated with Matrigel (BD, San
Diego, CA, USA). Media containing 10% FBS were added to the lower
chamber. After incubation for 48 h, non-invaded cells on the upper
surface of the filter were removed carefully with a cotton swab and
the undersurface adherent cells that had invaded through the
Matrigel were fixed in methanol and stained with 0.5% crystal
violet. The filter membrane was viewed under a microscope and 5
random fields were selected for cell counting. Experiments were
independently repeated three times.
Colony formation assay
Stable transfected cells were suspended in medium
containing DMEM medium with 10% FBS and 2.5 ml of 0.3% noble agar
(Sigma). Cells were plated onto a solidified medium containing 2.5
ml of 0.6% noble agar in a 60-mm dish. The dishes were incubated at
37°C with 5% CO2, and fresh medium was added every 3–4
days. After culture for 25 days, cells were staining with 0.05%
crystal violet, and forming colonies (>100 mm in diameter) were
counted using microscopy.
Xenografts
Fifteen female BALB/c nude mice were injected with
U2OS cells with stable knockdown of E2F1 or DDR1. Mice were
randomly chosen for the two experimental groups and one control
group, and each received a subcutaneous injection of a viable cell
suspension mixture (5×106) containing a 90% U2OS cells.
Subcutaneous tumor size was blindly measured twice a week with a
caliper. Tumor volumes were calculated using the formula: V =
W2 × L × 0.5, where W is tumor width and L is tumor
length. All of the mice were sacrificed on the fifth week after
injection, and the individual tumors were weighed. The animal
experiment was approved by the institutional review board of
Zhejiang Rongjun Hospital.
Immunohistochemistry
All procedures performed in studies involving human
participants were in accordance with the Ethics Committee of
Zhejiang Rongjun Hospital and the Second Hospital of Jiaxing and
with the 1964 Helsinki declaration and its later amendments or
comparable ethical standards. Osteosarcoma tissues were collected
by surgery from 81 patients at Zhejiang Rongjun Hospital (Jiaxing,
China) and the Second Hospital of Jiaxing (Jiaxing, China) from
2008 to 2011. All specimens were fixed in 10% neutral formalin,
embedded in paraffin and cut into 4-µm sections for
immunohistochemical staining. The EnVision™ two-step method was
used (Dako, Hamburg, Germany), as well as the following antibodies:
antibody against E2F1 and antibody against DDR1. To estimate the
score for each slide, at least 10 individual fields at ×200 were
chosen, and 100 cancer cells were counted in each field. The
immunostaining intensity was divided into four grades: 0, no
expression; 1, mildly positive; 2, moderately positive; and 3,
markedly positive. The proportion of positive-staining cells was
divided into five grades: 0, <10%; 1, 11–25%; 2, 26–50%; 3,
51–75%; and 4, >75%. The staining results were assessed and
confirmed by two independent investigators blinded to the clinical
data. The percentage of positivity of the tumor cells and the
staining intensities were then multiplied in order to generate the
IHC score, and graded as 0–6, low expression; 7–12, high
expression. Cases with a discrepancy in scores were discussed to
obtain a consensus.
Statistical analysis
All statistical analyses were carried out using the
GraphPad Prism 5.0 and SPSS 17.0 statistical software. Experimental
data are presented as mean ± SD from at least three independent
experiments. The χ2 test or Fisher's exact probability
test was used to compare clinicopathological features of the
patients with protein expression. Kaplan-Meier plots and log-rank
tests were used for survival analysis. Spearman test was used in
analyzing the correlation. A P-value of ≤0.05 was considered
statistically significant.
Results
E2F1 silencing not only reduces
osteosarcoma cell proliferation but attenuates migration and
invasion
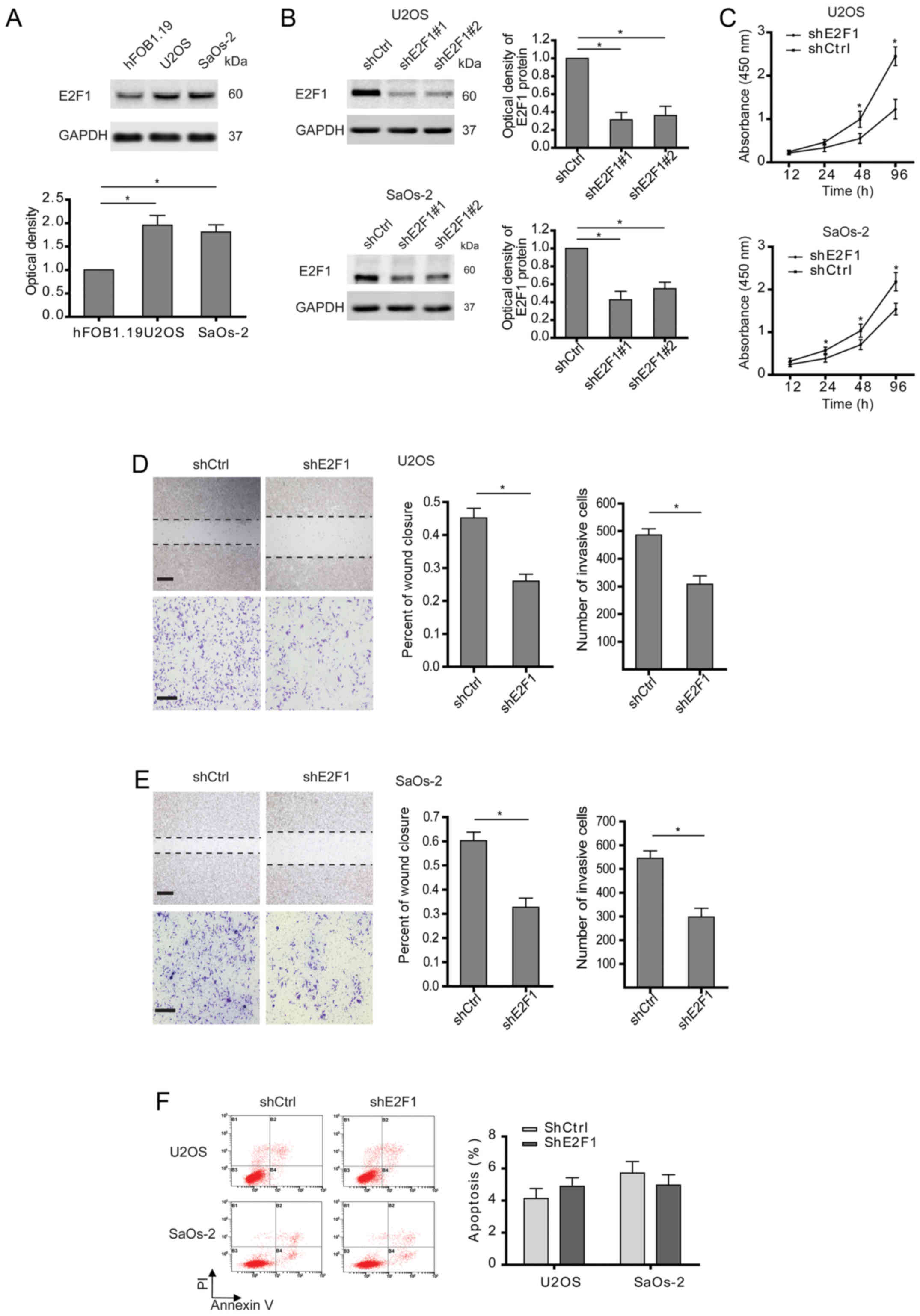
Consistent with previous observations (14), E2F1 was highly expressed in
osteosarcoma cells compared with osteoblast (Fig. 1A). To explore the roles of E2F1 in
osteosarcoma, two osteosarcoma cell lines (U2OS and SaOS-2) with
high E2F1 levels were examined after stable transfection with E2F1
shRNA (Fig. 1B). Two shRNAs
targeting E2F1 were checked for their efficacies, and as indicated
by western blotting, shE2F1#1 was slightly more effective for
inhibiting E2F1 expression than shE2F1#2. So, shE2F1#1 was applied
in the following experiments. As expected, cell growth was markedly
suppressed in the osteosarcoma cells with E2F1 silence compared to
control cells (Fig. 1C).
Interestingly, cell migration and invasion determined by wound
healing and Transwell assays were also inhibited in the
osteosarcoma cells with E2F1 knockdown (Fig. 1D and E). In consideration of the
relationship between E2F1 and apoptosis described in osteosarcoma
(15), the apoptosis of
osteosarcoma cells after abolishing E2F1 was analyzed. As shown in
Fig. 1F, knockdown of E2F1 had no
significant effect on apoptosis. Therefore, these findings indicate
that high expression of E2F1 is essential for the malignancy of
osteosarcoma.
Knockdown of E2F1 suppresses the
transcription of DDR1 in osteosarcoma cells
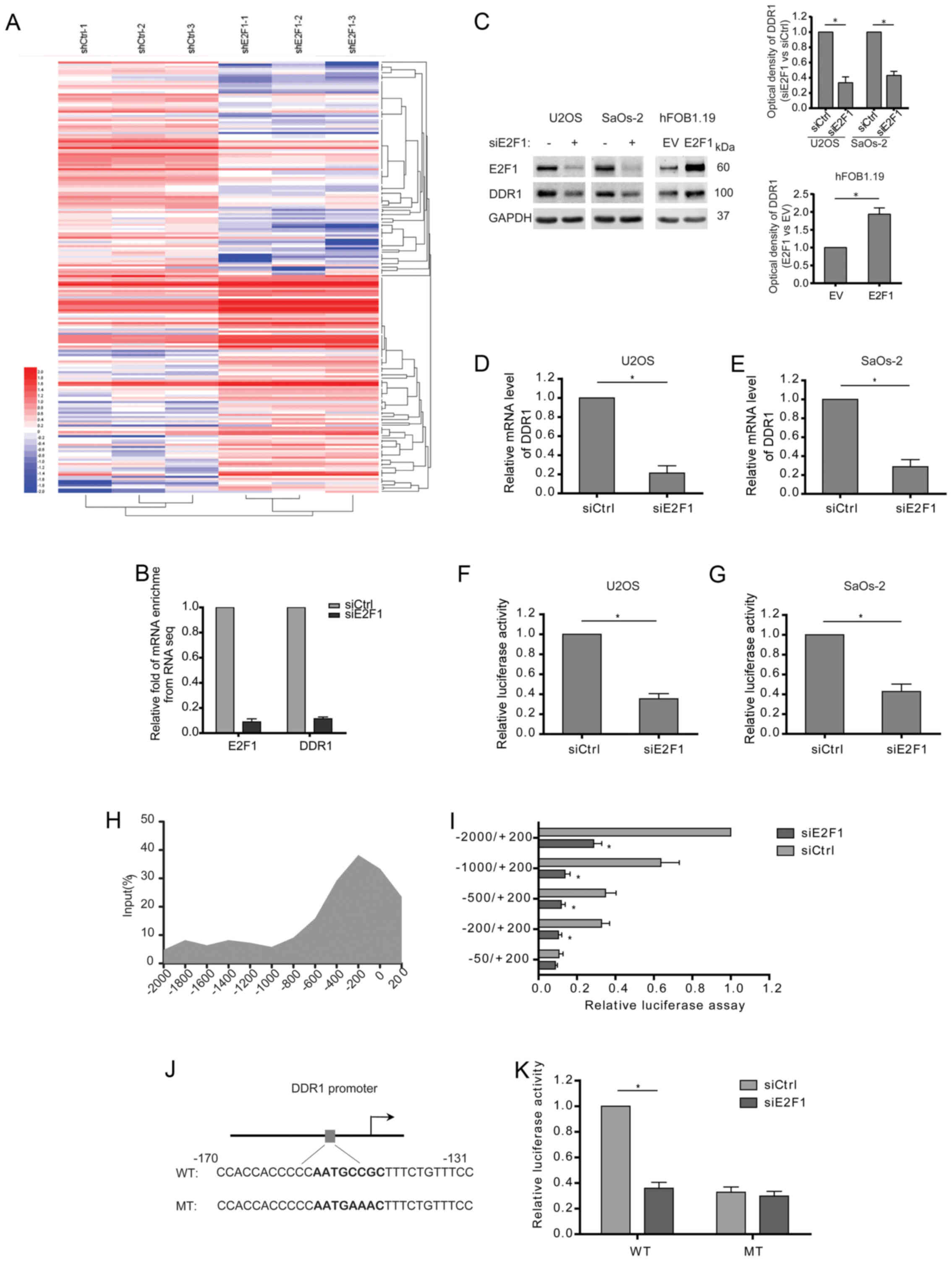
In attempt to understand the potential mechanisms
behind E2F1-induced migration and invasion, the expression of mRNA
isolated from the osteosarcoma cells with stable knockdown of E2F1
was profiled by RNA-seq analysis. Differentially expressed genes
(DEGs) were clustered and displayed in the heat map (Fig. 2A). Especially, the enrichment of
DDR1 mRNA was dramatically reduced after E2F1 knockdown (Fig. 2B). Consistently, the expression of
DDR1 was decreased in the E2F1-silenced osteosarcoma cells both at
the protein and mRNA levels (Fig.
2C–E). To determine whether E2F1 transactivates the
transcription of DDR1, 2 kb DDR1 promoter region upstream
transcription start site was cloned to a luciferase gene and then
co-transfected with E2F1 siRNA. The results showed that DDR1
promoter activity was strongly repressed in response to E2F1
knockdown in osteosarcoma cells (Fig.
2F and G). The direct regulation for DDR1 transcription by E2F1
was further confirmed by chromatin immunoprecipitation (ChIP),
which showed a peak binding of endogenous E2F1 to DDR1 promoter at
200 bp upstream the transcription start site (Fig. 2H). Moreover, the truncated DDR1
promoter without the region (−200/−50) failed to respond to E2F1
knockdown (Fig. 2I). Then, we
analyzed the promoter sequence around −200 bp by bioinformatic
software (JASPAR and Tfsearch), and a putative E2F1 binding site
(AATGCCGC) was identified (Fig.
2J). Furthermore, mutation of the site greatly diminished the
inhibitory effect of E2F1 silence on the transcription activity of
DDR1 promoter (Fig. 2K). Taken
together, E2F1 is required for DDR1 transcription by directly
binding to the promoter.
Silence of DDR1 reduces the levels of
p-AKT and p-STAT3 accompanied with an attenuated aggressiveness of
osteosarcoma cells
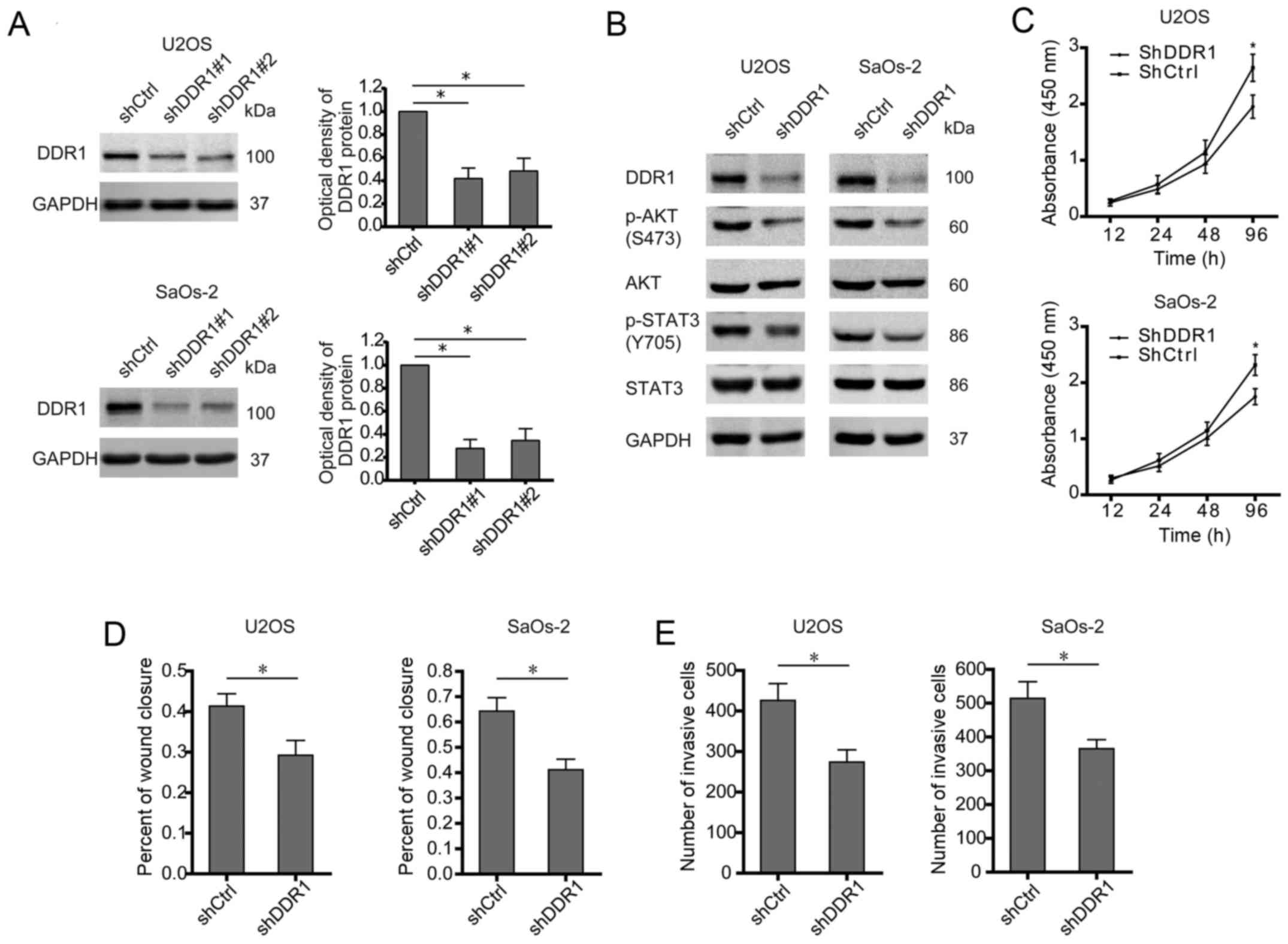
DDR1 serves as docking sites for the assembly of
downstream signaling molecules and then activates different target
mediators which are involved in several oncogenic processes
including survival, metastasis, and chemosensitivity (16,17).
In osteosarcoma cells (U2OS and SaOS-2), DDR1 was stably knocked
down by two shRNAs (Fig. 3A). Even
though no significant difference between shDDR1#1 and shDDR1#2 in
knocking down DDR1 was observed, shDDR1#1 showed a more powerful
effect than shDDR1#2, especially in SaOs-2 cells. Thus shDDR1#1 was
used for silencing DDR1 in this study. As a result, silence of DDR1
abolished the activation of both AKT and STAT3 signaling (Fig. 3B). Similarly with E2F1 knockdown,
deprivation of DDR1 undermined the malignant phenotypes of
osteosarcoma cells, as shown by decreased proliferation, migration,
and invasion (Fig. 3C–E).
Therefore, as a target gene of E2F1, DDR1 is also required for
triggering oncogenic signaling and aggressive behavior in
osteosarcoma.
DDR1 mediates E2F1-dependent malignant
phenotypes of osteosarcoma cells
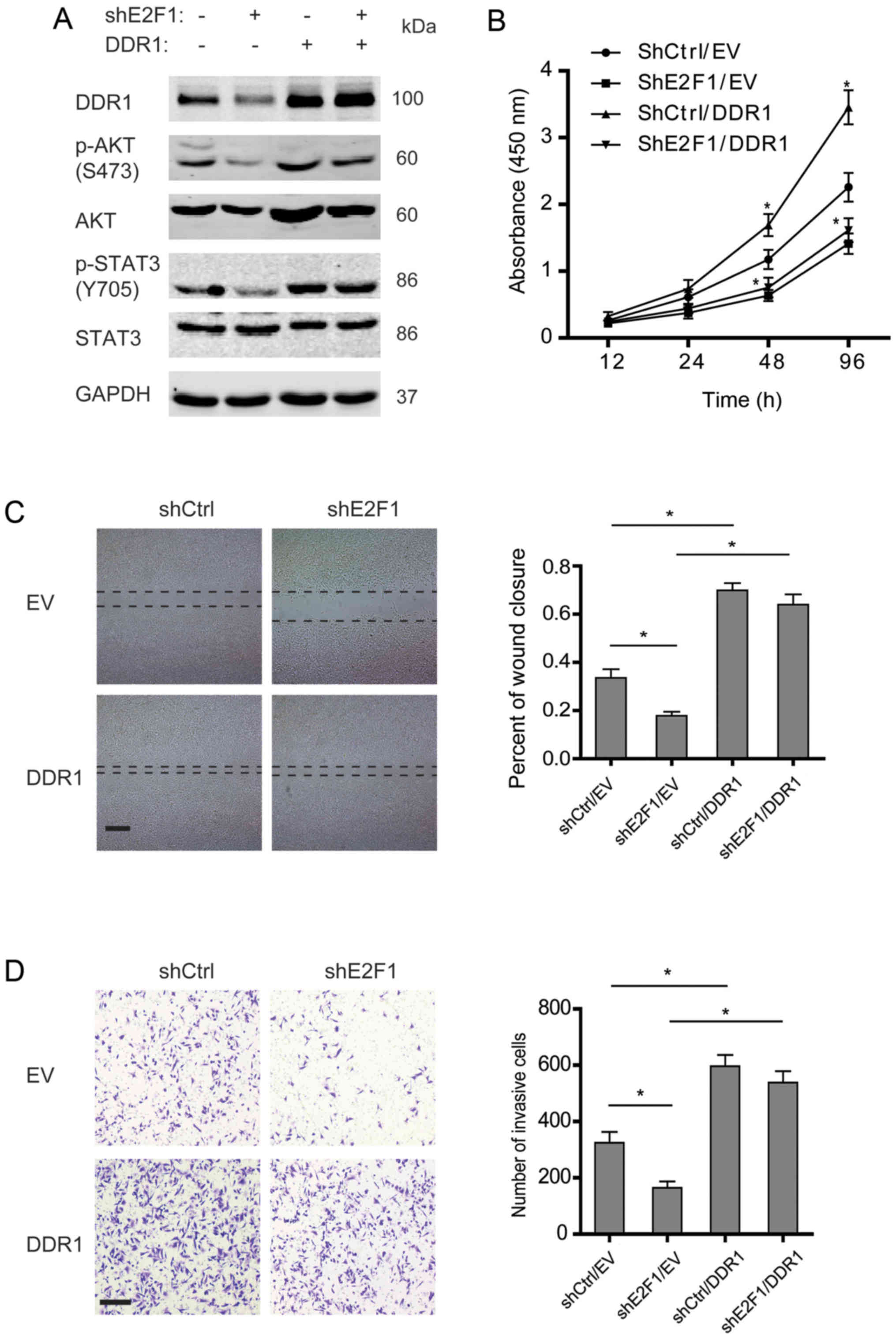
In order to determine the role of DDR1 in
E2F1-dependent malignancy of osteosarcoma, DDR1 was overexpressed
in the osteosarcoma cells with stable knockdown of E2F1. As shown
in Fig. 4A, knockdown of E2F1
decreased phosphorylation of AKT and STAT3, while DDR1
overexpression rescued the reduction. Further studies showed that
knockdown of E2F1 impaired the proliferation, migration, and
invasion of osteosarcoma cells, and overexpression of DDR1 could
enhance the aggressive phenotypes (Fig. 4B–D). Notably, DDR1 overexpression
was able to restore the incapability of migration and invasion in
the E2F1-silenced osteosarcoma cells but failed to rescue the
proliferation, which suggested that DDR1 displayed a crucial role
in E2F1-dependent migration and invasion but not proliferation in
osteosarcoma.
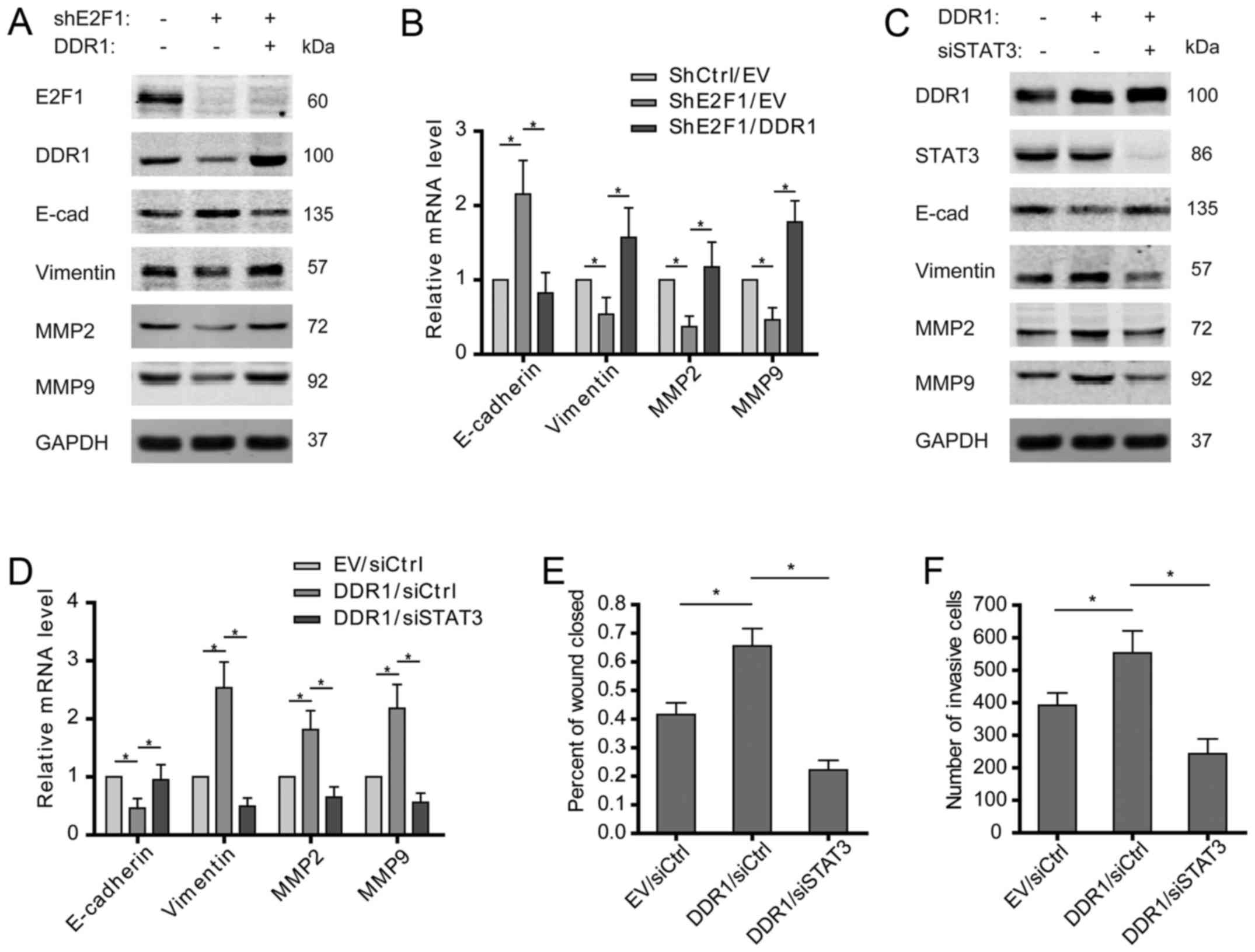
E2F1/DDR1/STAT3 axis drives the EMT of
osteosarcoma cells
Activation of epithelial-mesenchymal transition
(EMT) is important for driving cancer cell migration and invasion,
and STAT3 was reported to be involved in the EMT of various tumors.
To clarify whether E2F1/DDR1/STAT3 pathway is indispensable for
promoting osteosarcoma EMT, the expression levels of EMT markers in
osteosarcoma cells were examined. The results showed that E2F1
knockdown increased E-cadherin but decreased vimentin at both mRNA
and protein levels, which was then reversed by overexpression of
DDR1 (Fig. 5A and B). Furthermore,
overexpressed DDR1 was found to induce the EMT of osteosarcoma
cells, as shown by decreased E-cadherin and increased vimentin. As
expected, STAT3 depletion abolished the promotive effect of DDR1 on
the features of EMT (Fig. 5C and
D). Except for the canonical EMT markers expression of MMP2 and
MMP9 was also found to rely on an intact E2F1/DDR1/STAT3 signaling
in osteosarcoma cells (Fig. 5A–D).
Consistently, overexpression of DDR1 promoted the migration and
invasion of osteosarcoma cells, which was attenuated by silence of
STAT3 (Fig. 5E and F). These
findings suggest that E2F1/DDR1/STAT3 pathway may contribute to the
migration and invasion of osteosarcoma cells via driving EMT.
Knockdown of either E2F1 or DDR1 impairs
tumorigenesis of osteosarcoma cells
To further elucidate the roles of E2F1 and DDR1 in
osteosarcoma development, the osteosarcoma cells with stable
knockdown of DDR1 were also constructed (Fig. 6A). Then the colony-formation
ability of the osteosarcoma cells was determined by colony number
and size in soft-agar colony formation assays. As shown in Fig. 6B, knockdown of either E2F1 or DDR1
significantly decreased the number of colony formation compared
with control. Moreover, the constructed osteosarcoma cells were
subcutaneously injected into nude mice, and tumor growth was
monitored. It was shown that silence of E2F1 or DDR1 apparently
reduced the volumes and weights of xenografts (Fig. 6C–E), which suggested an E2F1 and
DDR1-dependent tumorigenesis in vivo. Collectively, these
data indicate a critical role of E2F1 and DDR1 in tumorigenesis of
osteosarcoma cells.
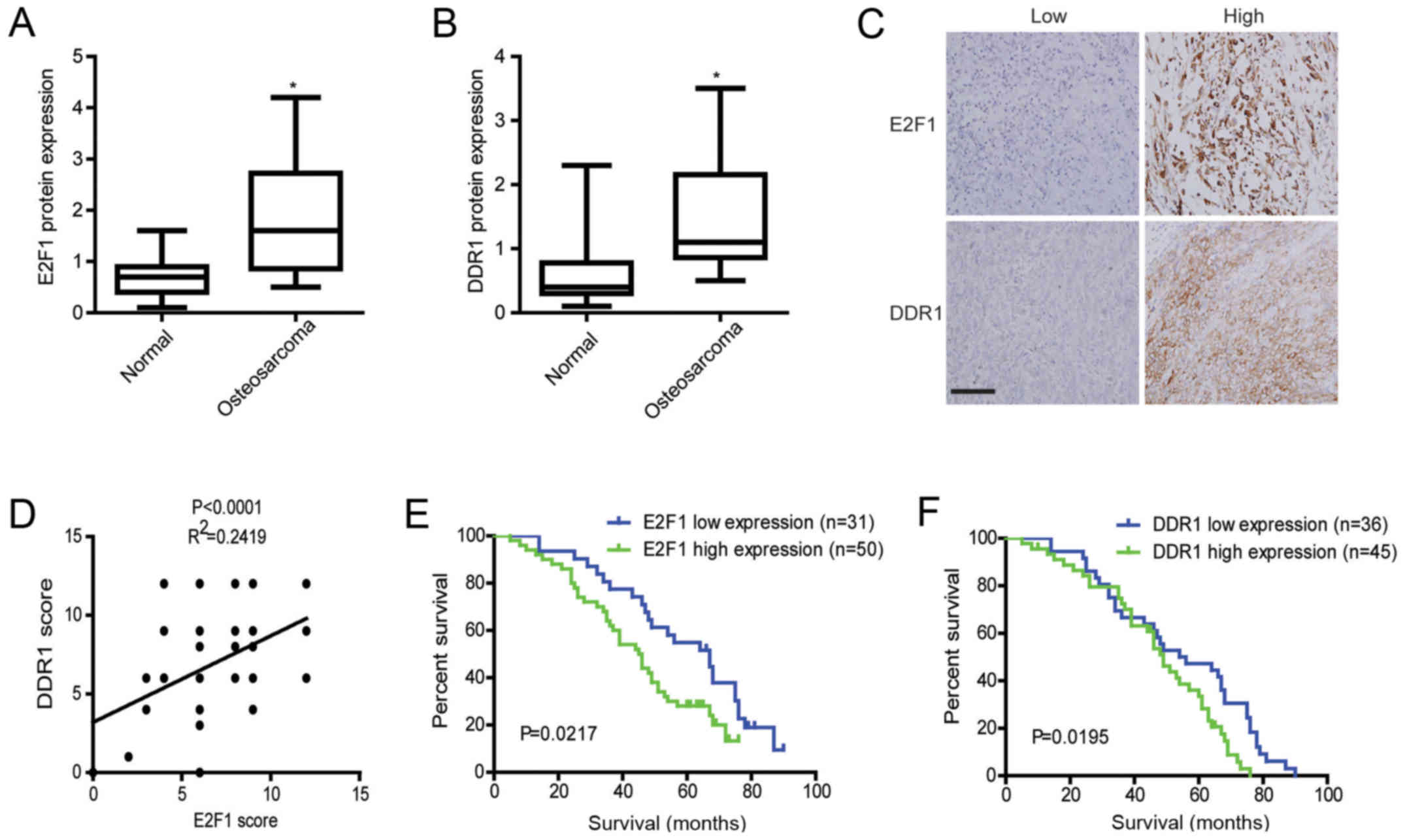
High expression of E2F1 and DDR1 predicts
worse survival in osteosarcoma
The results of western blotting showed that the
expression levels of E2F1 and DDR1 protein in osteosarcoma tissues
were significantly higher than those in adjacent non-cancerous
tissues (Fig. 7A and B).
Meanwhile, expressions of E2F1 and DDR1 were assessed in
osteosarcoma by immunohistochemistry. As demonstrated by E2F1 and
DDR1 staining, E2F1 was mainly accumulated in the nucleus and
cytoplasm, while DDR1 was diffusely expressed in the cytoplasm and
cytoplasmic membrane of osteosarcoma cells (Fig. 7C). Furthermore, E2F1 expression
levels paralleled the changes of DDR1 in the osteosarcoma cases as
shown by immunohistochemical analyses (Fig. 7D). Also, further analysis showed
that E2F1 and DDR1 staining was positively correlated with grades,
advanced TNM stages, and metastasis (Table I). Importantly, Kaplan-Meier
survival analyses showed that patients with osteosarcoma expressing
high E2F1 or DDR1 had a significantly worse overall survival than
those expressing low E2F1 or DDR1 (Kaplan-Meier, log-rank p=0.0217
(E2F1); p=0.0195 (DDR1), Fig. 7E and
F). These data suggest that abnormally expressed E2F1 and DDR1
promote osteosarcoma development and indicate a poor prognosis for
osteosarcoma patients.
 | Table ICorrelation of the expression of E2F1
and DDR1 with clinicopathological features in osteosarcoma. |
Table I
Correlation of the expression of E2F1
and DDR1 with clinicopathological features in osteosarcoma.
|
Characteristics | Cases | E2F1 expression
| DDR1 expression
|
|---|
| Low cases | High cases | P-value | Low cases | High cases | P-value |
|---|
| 81 | 45 | 36 | | 44 | 37 | |
| Sex | | | | 0.8542 | | | 0.4856 |
| Male | 40 | 22 | 18 | | 19 | 21 | |
| Female | 41 | 23 | 18 | | 25 | 16 | |
| Age (years) | | | | 0.6425 | | | 0.2489 |
| <30 | 62 | 35 | 27 | | 31 | 31 | |
| ≥30 | 19 | 10 | 9 | | 13 | 6 | |
| Tumor size
(cm) | | | | 0.1462 | | | 0.0611 |
| <5 | 53 | 30 | 23 | | 34 | 19 | |
| ≥5 | 28 | 15 | 13 | | 10 | 18 | |
| Grade | | | | 0.0157a | | | 0.0019a |
| Low | 16 | 12 | 4 | | 15 | 1 | |
| High | 65 | 33 | 32 | | 29 | 36 | |
| TNM | | | | 0.0101a | | | 0.0034a |
| I | 8 | 8 | 0 | | 8 | 0 | |
| II | 42 | 24 | 18 | | 28 | 24 | |
| III | 21 | 12 | 9 | | 8 | 13 | |
| IV | 10 | 1 | 9 | | 0 | 10 | |
| Histological
type | | | | 0.1601 | | | 0.2193 |
| Osteoblastic | 29 | 17 | 12 | | 14 | 15 | |
| Fibroblastic | 6 | 4 | 2 | | 5 | 1 | |
|
Chondroblastic | 30 | 16 | 14 | | 15 | 15 | |
| Others | 16 | 8 | 8 | | 10 | 6 | |
| Metastasis | | | | 0.0009a | | | 0.0002a |
| Yes | 10 | 1 | 9 | | 0 | 10 | |
| No | 71 | 44 | 27 | | 44 | 27 | |
Discussion
In the past, E2F1 was recognized as a crucial
transcription factor that regulated cell cycle and apoptosis.
However, the latest studies revealed that E2F1 prompted metastatic
and invasive progresses, except for its proliferative and apoptotic
functions (18). The diverse
behavior of E2F1 rely on different cell context and its various
target genes in different cancer types (19). In osteosarcoma, E2F1 was found to
be indispensable for proliferation (20), however, overexpression of E2F1 not
only exerted a growth-suppressing effect but facilitated the
chemotherapeutic drug-induced apoptosis (21,22).
Our findings revealed that knockdown of E2F1 had no effect on
apoptosis of untreated osteosarcoma cells but raised an apparent
growth retardation. More importantly, E2F1 silence significantly
suppressed the migration and invasion of osteosarcoma cells, which
suggested the necessity of E2F1 for osteosarcoma metastasis. Except
for promoting metastasis, high expression of E2F1 sensitizes
osteosarcoma to chemotherapy. More and more evidence support that
the E2F1-induced apoptosis was dependent on DNA damage which
altered the post-translational modifications of E2F1 (23), then phosphorylation and acetylation
of E2F1 predominantly strengthened its proapoptotic activity
(24).
Gene set enrichment analysis (GSEA) performed by Gao
et al indicated that the aberrant expression of E2F1 was a
key regulator of the entire metastatic process (25). Increased abundance of E2F1 triggers
invasion and metastasis by activating growth receptor signaling
pathways, which sheds light on the molecular mechanisms underlying
E2F1-induced prometastatic activity. Recent study revealed that
E2F1 enhanced metastatic behavior in melanomas by transactivating
the VEGFC receptor, VEGFR-3, and epidermal growth factor receptor
(EGFR) (6,26). In prostate cancer, E2F1 was found
to activate the transcription of androgen receptor which was
essential for the initiation and progression of prostate cancer
(27). Similarly, we reported that
DDR1, a discoidin domain receptor, was directly regulated by E2F1
which was verified as a transcription activator of DDR1 in
osteosarcoma. Then, the E2F1-dependent expression of DDR1
facilitated the proliferation, migration, and invasion of
osteosarcoma cells. However, overexpression of DDR1 partially
restored the impaired migration and invasion caused by E2F1
knockdown, but not for proliferation. So, even though DDR1 could
induce the proliferation of osteosarcoma cells, other factors or
pathways regulated by E2F1 are necessary for proliferation.
Although the roles of DDR1 in the metastasis of
osteosarcoma have rarely been reported, acquisition of a more
mesenchymal-like phenotype was associated with expression of DDR1
in other cancer types, such as gastric, colorectal, and pancreatic
cancer (13,28,29).
In the present study, we found that DDR1 was required for EMT which
might contribute to the E2F1-dependent aggressive behavior in
osteosarcoma cells. Mechanistically, DDR1 promoted EMT by
activating its downstream signaling pathways. Due to the extensive
deposition of collagen I characterized in osteosarcoma, activation
of highly expressed DDR1 by collagen is believed to activate
downstream signaling molecules that display as mediators to drive
EMT. In a recent study, a non-canonical DDR1 signaling was found to
enable the metastatic reactivation of breast cancer cells by
activating JAK2-STAT3 signaling (30). Given the important role of STAT3 as
an EMT signaling transducer, we examined the phosphorylated STAT3
(Y705) that was crucial for its dimerization and activation. Also,
evidence showed that activation of STAT3 by DDR1 mediated the EMT
and malignant phenotypes of osteosarcoma cells. Altogether, we
hypothesize that the E2F1/DDR1/STAT3 axis is critical for the
metastasis of osteosarcoma.
As another widely reported oncogene regulated by
DDR1, Matrix metalloproteinase (MMP) contributes to the matrix
components degradation that is required for tumor invasion
(31). Several studies reported
that DDR1 could play an invasive role in various tumors via
inducing MMP2 and MMP9 (32,33).
Multiple mechanisms may be employed in the induction of MMPs by
DDR1. In breast cancer, type IV collagen increased MMP2 and MMP9
secretion and invasion through activating DDR1-Src signaling
(34). Whereas, receptor signaling
activated by collagen was not requisite for DDR1 to support
invasion in oral squamous cell carcinoma (35). Except for inducing MMPs expression,
DDR1 activation by collagen could accelerate the conversion of
pro-MMP2 into its active form (36). In the present study, we found that
E2F1/DDR1/STAT3 signaling was essential for maintaining the high
expression of MMP2 and MMP9 which might enhance an invasive
behavior to osteosarcoma.
In conclusion, the present study demonstrates that
E2F1 is important for osteosarcoma proliferation, migration and
invasion. In addition, our findings identify a novel regulatory
pathway, E2F1/DDR1/STAT3 axis, which drives EMT and increases the
expressions of MMPs in osteosarcoma cells. As a consequence,
overactivation of the pathway results in the aggressive phenotypes
of osteosarcoma. Consistently, clinical analysis suggests that high
expressions of E2F1 and DDR1 indicate a relative poor prognosis in
osteosarcoma patients. Although this study provides a possible
molecular mechanism for E2F1-dependent osteosarcoma metastasis,
further studies are needed to elucidate the details of the
deregulation of E2F1 and its downstream pathways.
Acknowledgments
This study was supported by the Science Technology
Department of Zhejiang Province (2014C33122).
References
|
1
|
Anderson ME: Update on survival in
osteosarcoma. Orthop Clin North Am. 47:283–292. 2016. View Article : Google Scholar
|
|
2
|
Poletajew S, Fus L and Wasiutyński A:
Current concepts on pathogenesis and biology of metastatic
osteosarcoma tumors. Ortop Traumatol Rehabil. 13:537–545. 2011.
View Article : Google Scholar
|
|
3
|
Maximov VV and Aqeilan RI: Genetic factors
conferring metastasis in osteosarcoma. Future Oncol. 12:1623–1644.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dyson N: The regulation of E2F by
pRB-family proteins. Genes Dev. 12:2245–2262. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee JS, Leem SH, Lee SY, Kim SC, Park ES,
Kim SB, Kim SK, Kim YJ, Kim WJ and Chu IS: Expression signature of
E2F1 and its associated genes predict superficial to invasive
progression of bladder tumors. J Clin Oncol. 28:2660–2667. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Alla V, Engelmann D, Niemetz A, Pahnke J,
Schmidt A, Kunz M, Emmrich S, Steder M, Koczan D and Pützer BM:
E2F1 in melanoma progression and metastasis. J Natl Cancer Inst.
102:127–133. 2010. View Article : Google Scholar
|
|
7
|
Liang YX, Lu JM, Mo RJ, He HC, Xie J,
Jiang FN, Lin ZY, Chen YR, Wu YD, Luo HW, et al: E2F1 promotes
tumor cell invasion and migration through regulating CD147 in
prostate cancer. Int J Oncol. 48:1650–1658. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rammal H, Saby C, Magnien K, Van-Gulick L,
Garnotel R, Buache E, El Btaouri H, Jeannesson P and Morjani H:
Discoidin domain receptors: Potential actors and targets in cancer.
Front Pharmacol. 7:552016.PubMed/NCBI
|
|
9
|
Miao L, Zhu S, Wang Y, Li Y, Ding J, Dai
J, Cai H, Zhang D and Song Y: Discoidin domain receptor 1 is
associated with poor prognosis of non-small cell lung cancer and
promotes cell invasion via epithelial-to-mesenchymal transition.
Med Oncol. 30:6262013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Koh M, Woo Y, Valiathan RR, Jung HY, Park
SY, Kim YN, Kim HR, Fridman R and Moon A: Discoidin domain receptor
1 is a novel transcriptional target of ZEB1 in breast epithelial
cells undergoing H-Ras-induced epithelial to mesenchymal
transition. Int J Cancer. 136:E508–E520. 2015. View Article : Google Scholar
|
|
11
|
Shimada K, Nakamura M, Ishida E, Higuchi
T, Yamamoto H, Tsujikawa K and Konishi N: Prostate cancer antigen-1
contributes to cell survival and invasion though discoidin receptor
1 in human prostate cancer. Cancer Sci. 99:39–45. 2008.
|
|
12
|
Huo Y, Yang M, Liu W, Yang J, Fu X, Liu D,
Li J, Zhang J, Hua R and Sun Y: High expression of DDR1 is
associated with the poor prognosis in Chinese patients with
pancreatic ductal adenocarcinoma. J Exp Clin Cancer Res. 34:882015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xie R, Wang X, Qi G, Wu Z, Wei R, Li P and
Zhang D: DDR1 enhances invasion and metastasis of gastric cancer
via epithelial-mesenchymal transition. Tumour Biol. 37:12049–12059.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pagliarini V, Giglio P, Bernardoni P, De
Zio D, Fimia GM, Piacentini M and Corazzari M: Downregulation of
E2F1 during ER stress is required to induce apoptosis. J Cell Sci.
128:1166–1179. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shats I, Gatza ML, Liu B, Angus SP, You L
and Nevins JR: FOXO transcription factors control E2F1
transcriptional specificity and apoptotic function. Cancer Res.
73:6056–6067. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Summy JM and Gallick GE: Src family
kinases in tumor progression and metastasis. Cancer Metastasis Rev.
22:337–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Valencia K, Ormazabal C, Zandueta C,
Luis-Ravelo D, Antó I, Pajares MJ, Agorreta J, Montuenga LM,
Martínez-Canarias S, Leitinger B, et al: Inhibition of collagen
receptor discoidin domain receptor-1 (DDR1) reduces cell survival,
homing, and colonization in lung cancer bone metastasis. Clin
Cancer Res. 18:969–980. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Engelmann D and Pützer BM: The dark side
of E2F1: In transit beyond apoptosis. Cancer Res. 72:571–575. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meng P and Ghosh R: Transcription
addiction: Can we garner the Yin and Yang functions of E2F1 for
cancer therapy? Cell Death Dis. 5:e13602014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ahn JD, Kim CH, Magae J, Kim YH, Kim HJ,
Park KK, Hong S, Park KG, Lee IK and Chang YC: E2F decoy
oligodeoxynucleotides effectively inhibit growth of human tumor
cells. Biochem Biophys Res Commun. 310:1048–1053. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liontos M, Niforou K, Velimezi G, Vougas
K, Evangelou K, Apostolopoulou K, Vrtel R, Damalas A,
Kontovazenitis P, Kotsinas A, et al: Modulation of the E2F1-driven
cancer cell fate by the DNA damage response machinery and potential
novel E2F1 targets in osteosarcomas. Am J Pathol. 175:376–391.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Russo AJ, Magro PG, Hu Z, Li WW, Peters R,
Mandola J, Banerjee D and Bertino JR: E2F-1 overexpression in U2OS
cells increases cyclin B1 levels and cdc2 kinase activity and
sensitizes cells to antimitotic agents. Cancer Res. 66:7253–7260.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Stevens C, Smith L and La Thangue NB: Chk2
activates E2F-1 in response to DNA damage. Nat Cell Biol.
5:401–409. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Galbiati L, Mendoza-Maldonado R, Gutierrez
MI and Giacca M: Regulation of E2F-1 after DNA damage by
p300-mediated acetylation and ubiquitination. Cell Cycle.
4:930–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gao Y, Li H, Ma X, Fan Y, Ni D, Zhang Y,
Huang Q, Liu K, Li X, Wang L, et al: KLF6 suppresses metastasis of
clear cell renal cell carcinoma via transcriptional repression of
E2F1. Cancer Res. 77:330–342. 2017. View Article : Google Scholar
|
|
26
|
Engelmann D, Mayoli-Nüssle D, Mayrhofer C,
Fürst K, Alla V, Stoll A, Spitschak A, Abshagen K, Vollmar B, Ran
S, et al: E2F1 promotes angiogenesis through the VEGF-C/VEGFR-3
axis in a feedback loop for cooperative induction of PDGF-B. J Mol
Cell Biol. 5:391–403. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sharma A, Yeow WS, Ertel A, Coleman I,
Clegg N, Thangavel C, Morrissey C, Zhang X, Comstock CE, Witkiewicz
AK, et al: The retinoblastoma tumor suppressor controls androgen
signaling and human prostate cancer progression. J Clin Invest.
120:4478–4492. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hu Y, Liu J, Jiang B, Chen J, Fu Z, Bai F,
Jiang J and Tang Z: MiR-199a-5p loss up-regulated DDR1 aggravated
colorectal cancer by activating epithelial-to-mesenchymal
transition related signaling. Dig Dis Sci. 59:2163–2172. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shintani Y, Fukumoto Y, Chaika N, Svoboda
R, Wheelock MJ and Johnson KR: Collagen I-mediated up-regulation of
N-cadherin requires cooperative signals from integrins and
discoidin domain receptor 1. J Cell Biol. 180:1277–1289. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao H, Chakraborty G, Zhang Z, Akalay I,
Gadiya M, Gao Y, Sinha S, Hu J, Jiang C, Akram M, et al:
Multi-organ site metastatic reactivation mediated by non-canonical
discoidin domain receptor 1 signaling. Cell. 166:47–62. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gialeli C, Theocharis AD and Karamanos NK:
Roles of matrix metalloproteinases in cancer progression and their
pharmacological targeting. FEBS J. 278:16–27. 2011. View Article : Google Scholar
|
|
32
|
Juin A, Di Martino J, Leitinger B, Henriet
E, Gary AS, Paysan L, Bomo J, Baffet G, Gauthier-Rouvière C,
Rosenbaum J, et al: Discoidin domain receptor 1 controls linear
invadosome formation via a Cdc42-Tuba pathway. J Cell Biol.
207:517–533. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang SH, Baek HA, Lee HJ, Park HS, Jang
KY, Kang MJ, Lee DG, Lee YC, Moon WS and Chung MJ: Discoidin domain
receptor 1 is associated with poor prognosis of non-small cell lung
carcinomas. Oncol Rep. 24:311–319. 2010.PubMed/NCBI
|
|
34
|
Castro-Sanchez L, Soto-Guzman A,
Guaderrama-Diaz M, Cortes-Reynosa P and Salazar EP: Role of DDR1 in
the gelatinases secretion induced by native type IV collagen in
MDA-MB-231 breast cancer cells. Clin Exp Metastasis. 28:463–477.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hidalgo-Carcedo C, Hooper S, Chaudhry SI,
Williamson P, Harrington K, Leitinger B and Sahai E: Collective
cell migration requires suppression of actomyosin at cell-cell
contacts mediated by DDR1 and the cell polarity regulators Par3 and
Par6. Nat Cell Biol. 13:49–58. 2011. View
Article : Google Scholar :
|
|
36
|
Ram R, Lorente G, Nikolich K, Urfer R,
Foehr E and Nagavarapu U: Discoidin domain receptor-1a (DDR1a)
promotes glioma cell invasion and adhesion in association with
matrix metalloproteinase-2. J Neurooncol. 76:239–248. 2006.
View Article : Google Scholar
|