Introduction
Resveratrol (Fig.
1) is a chemopreventive molecule which inhibits the
proliferation of tumor cells of various etiologies. The biological
properties of resveratrol have been described previously (1–3) and
its beneficial effects have promoted the investigation of novel,
more effective analogues (4,5).
Resveratrol has been proposed to be a good anti-carcinogenic agent
due to its low toxicity and capacity to modulate numerous molecular
pathways involved in cancer progression (6). However, the direct molecular target
of resveratrol remains elusive (7). Colon cancer is one of the types of
cancer with the highest mortality rate in the United States and has
been ranked the third most common cause of cancer mortality
(8). Moreover, an increase in the
colon cancer incidence in adults under 50 years of age has been
reported (9). Cancer
chemoprevention has been reported to be a promising strategy to
prevent cancer death, particularly in colon cancer, due to the
relatively slow progression of the colorectal adenomatous polyps in
colon cancer, which enhances the opportunity for chemoprevention
treatment. Results from epidemiological studies have associated
fruit and vegetable consumption with a reduced risk of colon
cancer, and various phytochemicals from commonly consumed fruit and
vegetables have been identified as potential anticancer agents
(10). The present study aimed to
investigate the anti-cancer effect of resveratrol in HCT116 and
Caco-2 human colon cancer cells. To the best of our knowledge, the
present study is the first to demonstrate that caspase and
cyclin-cyclin-dependent kinase (CDK) proteins are involved in
resveratrol-induced apoptosis and cell cycle arrest.
Materials and methods
Drugs and reagents
Resveratrol was purchased from Sigma-Aldrich (St.
Louis, MO, USA). Resveratrol was dissolved to a concentration of 50
mM in 100% dimethyl sulfoxide (DMSO) as a stock solution and stored
at −20°C. The final DMSO concentrations used in the present study
were ≤0.1%. Antibodies against cleaved-caspase-9,
cleaved-caspase-7, cleaved-PARP, CDK2, CDK4, Cyclin D1, PCNA and
GAPDH were all purchased from Cell Signaling Technology, Inc.
(Beverly, MA, USA) and goat anti-rabbit immunoglobulin G
(IgG)-horseradish peroxidase (HRP; EarthOx, LLC, San Francisco, CA,
USA) was used as a secondary antibody.
Cell culture and resveratrol
treatment
HCT116 and Caco-2 human colon cancer cells were
provided by the Affiliated Hospital of Guangdong Medical College
(Zhanjiang, China). The HCT116 and Caco-2 cells were cultured in
RPMI-1640 (Gibco-BRL, Grand Island, NY, USA) and McCoy’s 5A medium
(Gibco-BRL), respectively, supplemented with 10% (v/v) fetal bovine
serum (Gibco-BRL), penicillin 100 U/ml and streptomycin 100 U/ml
and maintained in a humidified atmosphere of 95% air and 5%
CO2 at 37°C. When the proliferation of the cells was
60–70%, the cells were treated with various concentrations of
resveratrol (10, 50, 100 or 150 μM) for 24 h.
Annexin V/propidium iodide (PI) double
staining
Apoptotic cells were quantified using an Annexin
V-fluorescein isothiocyanate (FITC)/PI kit (BD Biosciences, San
Jose, CA, USA) and detected using flow cytometry using a
FACSCalibur™ flow cytometer (Becton, Dickinson and Company,
Franklin Lakes, NJ, USA) and analyzed using Modfit and CellQuest™
software (Becton, Dickinson and Company). In brief, cells were
pretreated with 10, 50, 100 or 150 μM resveratrol for 24 h and
washed with phosphate-buffered saline (PBS). Cells were then
collected and resuspended in binding buffer [10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.5), 2.5 mM
CaCl2 and 140 mM NaCl). Cells were incubated with
Annexin V-fluorescein isothiocyanate and PI for 15 min in the dark,
prior to flow cytometric analysis. Annexin V-positive cells were
considered to be in the early stage of apoptosis, whereas Annexin V
and PI-positive cells were considered to be in the late stage of
apoptosis.
Cell cycle analysis
Cells were quantified using a Cell Cycle Analysis
kit (Beyotime Institute of Biotechnology, Shanghai, China),
detected using a FACSCalibur flow cytometer (Becton, Dickinson and
Company) and analyzed using Modfit and CellQuest software 6.1
(Becton, Dickinson and Company). In brief, cells were pretreated
with 10, 50, 100 or 150 μM resveratrol for 24 h, washed with PBS,
then fixed with 70% ethanol for 24 h. Cells were incubated with
propyl iodide organism dye for 30 min at 37°C, followed by flow
cytometric analysis.
MTT assay
HCT116 and Caco-2 cell densities were adjusted to
2×104 cells per 100 μl. Cells were seeded onto 96-well
plates, which were placed in an incubator overnight to allow for
attachment and recovery. In brief, cells were pretreated with 10,
50, 100 or 150 μM resveratrol for 24 h and MTT was then dissolved
to a concentration of 5 mg/ml in warm assay medium. A total of 20
μl MTT solution was transferred to each well to yield a final
volume of 120 μl/well. Plates were incubated for 4 h at 37°C in 5%
CO2. Following incubation, supernatants were removed and
150 μl DMSO was added. Plates were then placed on an orbital shaker
for 5 min and the absorbance was recorded using the EnSpire™ 2300
Multilabel Plate Reader (PerkinElmer, Inc., Waltham, MA, USA) at
595 nm.
Western blot analysis of
resveratrol-regulated apoptotic proteins and cell cycle
proteins
HCT116 and Caco-2 cells were collected following
treatment, then lysed in lysis buffer [100 mM Tris-HCl (pH 6.8), 4%
(m/v) sodium dodecylsulfonate (SDS), 20% (v/v) glycerol, 200 mM
2-mercaptoethanol, 1 mM phenylmethyl sulfonylfluoride and 1 g/ml
aprotinin] for 30 min on ice. The lysates were separated using
centrifugation at 4°C for 15 min at 3,913 × g. The total protein
concentration in the supernatants was detected by bicinchoninic
acid (BCA) assay using a BCA Protein Assay kit (Beyotime Institute
of Biotechnology). SDS-PAGE was performed using an 8–15% gradient
or standard polyacrylamide gels. Proteins were subsequently
transferred to nitrocellulose membranes, which were saturated with
5% milk in TBST (Tris-buffered saline and 1% Tween-20) and
incubated with primary antibodies in a diluent overnight at 4°C.
Membranes were washed three times with TBST and incubated with goat
anti-rabbit IgG-HRP for 1 h, followed by washing four times with
TBST. Detection was performed using an Odyssey® Infrared
Imaging System (Li-Cor Biosciences, Lincoln, NE, USA).
Statistical analysis
Data were analyzed using GraphPad Prism 5 (GraphPad
Software, Inc., San Diego, CA, USA). Data are presented as the mean
± standard deviation from triplicate experiments performed in a
parallel manner unless otherwise stated. Statistical differences
were assessed using the Student’s t-test and P<0.05 was
considered to indicate a statistically significant difference. Data
are representative of at least three independent experiments.
Results
Resveratrol inhibits cell proliferation
and promotes cell apoptosis
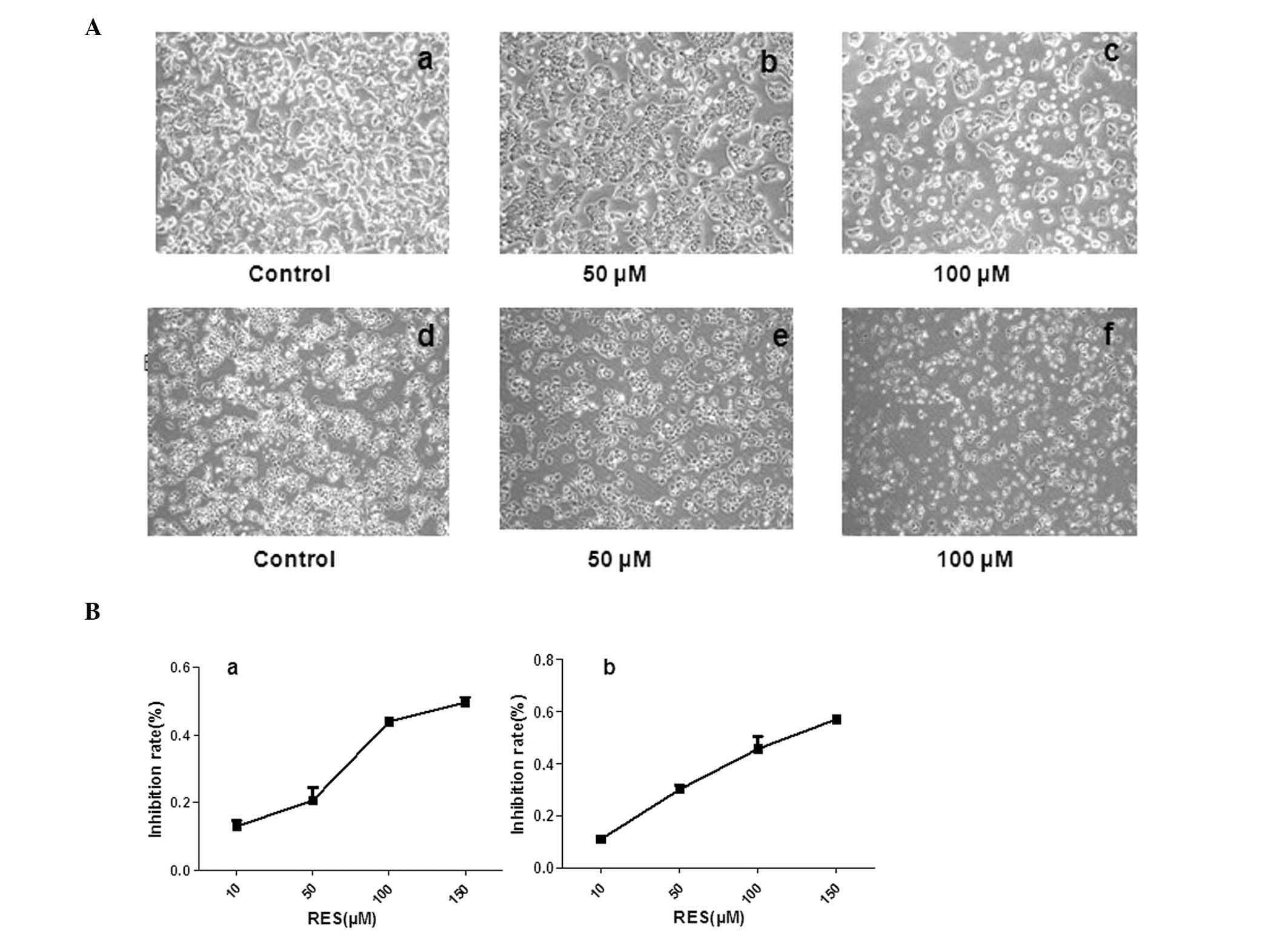
The untreated HCT116 and Caco-2 cells were observed
to be healthy with clear skeletons, whereas the cells treated with
resveratrol were distorted with certain cells becoming round.
Furthermore, the number of sloughed cells increased with increasing
drug concentration (Fig. 2A). MTT
assay was used to assess the inhibitory effect of resveratrol on
the HCT116 and Caco-2 cells and revealed a significant
dose-dependent inhibition of cell growth after 24 h of treatment
(Fig. 2B). The IC50s
for resveratrol on HCT116 and Caco-2 cells were 170 and 120 μM,
respectively, which were calculated using GRAFIT-Erithacus
IC50 software (11).
Resveratrol was found to exert a strong inhibitory effect on the
viability of HCT116 and Caco-2 cells, which may contribute to its
antitumor potency. Cells treated with 50 and 150 μM resveratrol
became round and floating, with inhibited cell growth. Furthermore,
the majority of the HCT116 and Caco-2 cells underwent severe
apoptosis with increasing resveratrol concentration.
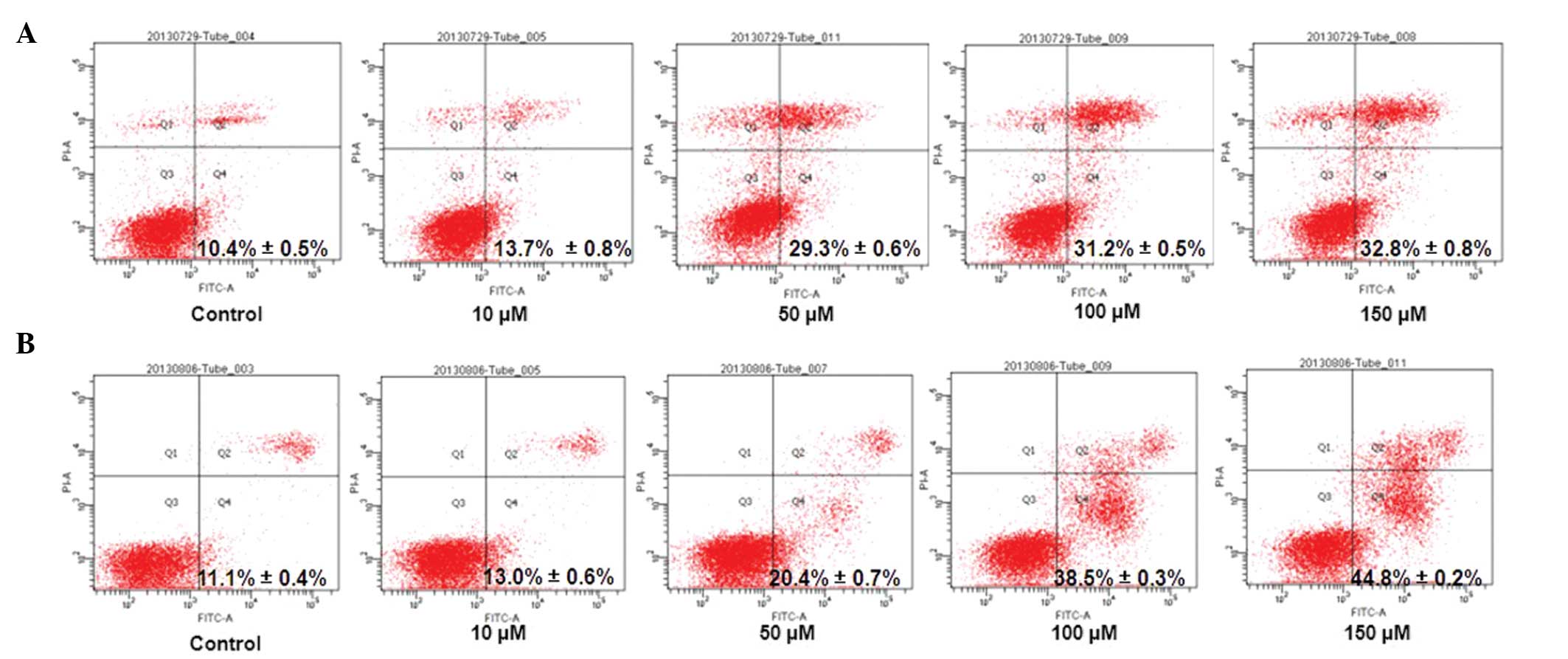
Annexin V/PI double staining was used to detect
apoptosis in the HCT116 and Caco-2 cells (Fig. 3). With increasing drug
concentration, the apoptosis rates of the cells were found to
increase. Resveratrol inhibited proliferation and promoted
apoptosis in HCT116 and Caco-2 cells concentration-dependently.
Resveratrol promotes cell cycle
arrest
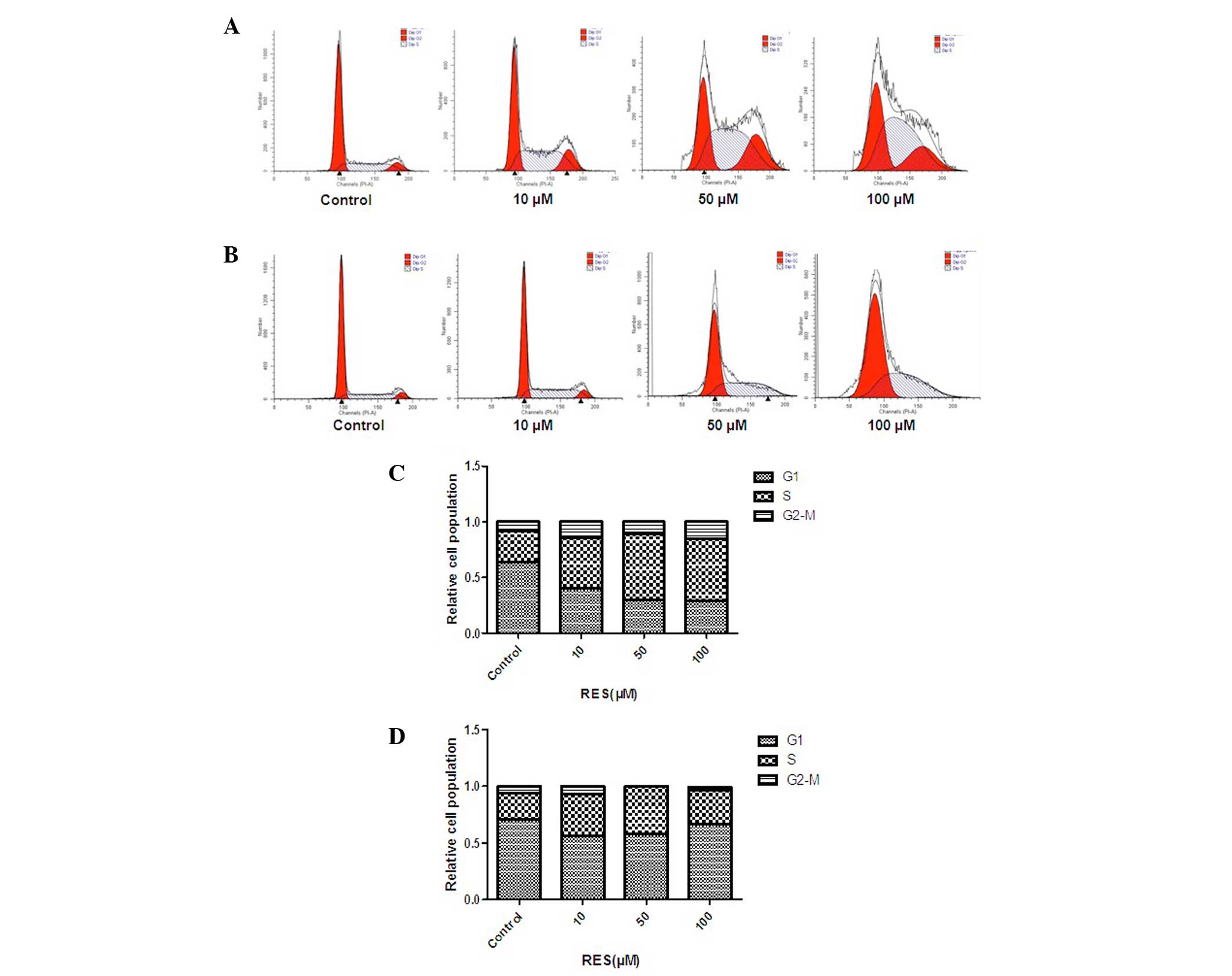
In order to determine whether resveratrol causes
cell cycle arrest in human colon cancer cells, HCT116 and Caco-2
cells treated with DMSO or resveratrol for 24 h were subjected to
flow cytometric analysis following DNA staining (Fig. 4). With increasing resveratrol
concentration, the proportion of cells in G1/S-phase was
found to increase after 24 h of resveratrol treatment. These
results demonstrated that resveratrol inhibited proliferation and
promoted apoptosis in HCT116 and Caco-2 cells in a
concentration-dependent manner.
Resveratrol activates apoptotic proteins
and induces cell cycle arrest
Cells were treated with 10, 50 or 100 μM resveratrol
for 24 h. The apoptotic protein expression of cleaved-caspase-9,
cleaved-caspase-7 and cleaved-PARP was found to increase in a
concentration-dependent manner in the resveratrol-treated HCT116
(Fig. 5A) and Caco-2 cells
(Fig. 5B) compared with the
control cells. Furthermore, the protein expression of the cycle
arrest proteins CDK2, CDK4, cyclin D1, PCNA and P21 were observed
to decrease in a concentration-dependent manner in the
resveratrol-treated HCT116 (Fig.
6A) and Caco-2 (Fig. 6B) cells
compared with the control cells.
Discussion
As a stilbene compound, resveratrol has been widely
studied due to its health promoting potential (3,12–15).
The present study aimed to investigate the effect of resveratrol in
HCT116 and Caco-2 human colon cancer cells. In the present study,
the inhibitory and apoptosis-promoting effects of resveratrol on
the growth of two human colon cancer cell lines with different
genetic aberrations and aggressiveness, were assessed. Caco-2
cancer cells were found to be more sensitive to resveratrol
treatment than HCT116 cancer cells (Fig. 4). Furthermore, the expression of
the apoptosis-associated proteins cleaved caspase-7, cleaved
caspase-9 and cleaved PARP (Fig.
5) were found to increase in a dose-dependent manner. The
activation of caspase-9 and -7 are crucial steps in apoptotic cell
death (16–19), which consequently induce PARP
cleavage. PARP has been proposed to be important for controlling
numerous cellular processes, including DNA repair, cell death,
chromatin function and genome stability (20–22).
PARP activation through cleavage is an early DNA damage response.
The present study found that resveratrol increased
cleaved-caspase-9 and -7 protein expression in HCT-116 and Caco-2
cells, while cleaved PARP, which is a marker of cell apoptosis, was
increased.
Furthermore, the present study demonstrated that
human colon cancer cells are susceptible to resveratrol-induced
inhibition of proliferation, predominantly through cell cycle
arrest. Of note, the Caco-2 cells were observed to be more
sensitive to resveratrol-induced cell cycle arrest than the HCT116
cells.
Resveratrol has been shown to induce cell cycle
arrest in a variety of other human cancer cell lines, including
breast cancer, leukemia, prostate and colon cancer (12,24,25).
Cell cycle progression, including genome duplication, is controlled
by CDKs and CDK inhibitors. Among the various roles reported for
p21, the most studied role is associated with its CDK and PCNA
binding domains (26,27). CDK genes are often constitutively
expressed and CDKs are relatively stable, whereas cyclin genes
exhibit periodic patterns of expression and are subject to
regulated degradation. During the transition between G1-
and S-phase, cyclin D1 and CDK4 are the first to be expressed
during early G1-phase (28). Cell cycle arrest occurs due to the
loss of cyclin expression and CDK activity. In order to investigate
the mechanism underlying resveratrol-induced G1/S-phase
cell cycle arrest, cell cycle regulatory proteins and CDKs were
analyzed using western blot analysis. In the present study,
resveratrol was found to induce G1/S-phase cell cycle
arrest (Fig. 3). Furthermore, the
expression of cyclin D1, CDK4, P21, PCNA and CDK2 were observed to
be reduced in a concentration-dependent manner in the HCT-116 and
Caco-2 cells (Fig. 6). These
findings therefore suggested that the resveratrol-induced
G1/S-phase cell cycle arrest may be mediated through the
cyclin-CDK checkpoint.
In conclusion, the present study identified that
resveratrol induced apoptosis and cell arrest in HCT116 and Caco-2
human colon cancer cells through caspase-dependent and cyclin-CDK
mechanisms. Furthermore, the Caco-2 cells were observed to be more
sensitive to resveratrol treatment than the HCT116 cells. These
findings suggested that resveratrol may be a novel candidate for
colon cancer therapy.
Acknowledgements
The present study was supported by grants from the
National Natural Science Fund (grant no. 81041099) and the
Guangdong Province Natural Science Fund (grant no. S2011010003750).
The present study was also supported by the Institute of Neurology,
Affiliated Hospital of Guangdong Medical College.
References
|
1
|
Fulda S: Resveratrol and derivatives for
the prevention and treatment of cancer. Drug Discov Today.
15:757–765. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Calabrese EJ, Mattson MP and Calabrese V:
Dose response biology: the case of resveratrol. Hum Exp Toxicol.
29:1034–1037. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Park SJ, Ahmad F, Philp A, et al:
Resveratrol ameliorates aging-related metabolic phenotypes by
inhibiting cAMP phosphodiesterases. Cell. 148:421–433. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Szekeres T, Fritzer-Szekeres M, Saiko P
and Jäger W: Resveratrol and resveratrol analogues -
structure-activity relationship. Pharm Res. 27:1042–1048. 2010.
View Article : Google Scholar
|
|
5
|
Szekeres T, Saiko P, Fritzer-Szekeres M,
Djavan B and Jäger W: Chemopreventive effects of resveratrol and
resveratrol derivatives. Ann NY Acad Sci. 1215:89–95. 2011.
View Article : Google Scholar
|
|
6
|
Athar M, Back JH, Kopelovich L, Bickers DR
and Kim AL: Multiple molecular targets of resveratrol:
Anti-carcinogenic mechanisms. Arch Biochem Biophys. 486:95–102.
2009. View Article : Google Scholar
|
|
7
|
Tennen RI, Michishita-Kioi E and Chua KF:
Finding a target for resveratrol. Cell. 148:387–389. 2012.
View Article : Google Scholar
|
|
8
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
9
|
Siegel RL, Jemal A and Ward EM: Increase
in incidence of colorectal cancer among young men and women in the
United States. Cancer Epidemiol Biomarkers Prev. 18:1695–1698.
2009. View Article : Google Scholar
|
|
10
|
De la Rosa L, Alvarez-Parrilla E and
González-Aguilar GA: Fruit and vegetable phytochemicals: Chemistry,
nutritional value and stability Phenolic Compounds: Chemistry and
Occurrence in Fruits and Vegetables. Wiley-Blackwell, EUA; pp.
53–88. 2010
|
|
11
|
Agarwal S, Arya V and Zhang L: Review of
P-gp Inhibition data in recently approved new drug applications:
utility of the proposed [I(1) ]/IC(50) and [I(2) ]/IC(50) criteria
in the P-gp decision tree. J Clin Pharmacol. 53:228–233. 2013.
|
|
12
|
Nutakul W, Sobers HS, Qiu P, et al:
Inhibitory effects of resveratrol and pterostilbene on human colon
cancer cells: a side-by-side comparison. J Agric Food Chem.
59:10964–10970. 2011. View Article : Google Scholar
|
|
13
|
Piotrowska H, Kucinska M and Murias M:
Biological activity of piceatannol: leaving the shadow of
resveratrol. Mutat Res. 750:60–82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
de Athayde Moncorvo Collado A, Corbalán N,
Homolya L, Morero R and Minahk C: Resveratrol modulates ATPase
activity of liposome-reconstituted ABCG1. FEBS Lett. 587:2359–2363.
2013.PubMed/NCBI
|
|
15
|
Um JH, Park SJ, Kang H, et al:
AMP-activated protein kinase-deficient mice are resistant to the
metabolic effects of resveratrol. Diabetes. 59:554–563. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yasuda Y, Saito M, Yamamura T, Yaguchi T
and Nishizaki T: Extracellular adenosine induces apoptosis in
Caco-2 human colonic cancer cells by activating caspase-9/−3 via
A(2a) adenosine receptors. J Gastroenterol. 44:56–65. 2009.
|
|
17
|
Allan LA and Clarke PR: Apoptosis and
autophagy: Regulation of caspase-9 by phosphorylation. FEBS J.
276:6063–6073. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boucher D, Blais V and Denault JB:
Caspase-7 uses an exosite to promote poly(ADP ribose) polymerase 1
proteolysis. Proc Natl Acad Sci USA. 109:5669–5674. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rodríguez-Berriguete G, Galvis L, Fraile
B, et al: Immunoreactivity to caspase-3, caspase-7, caspase-8, and
caspase-9 forms is frequently lost in human prostate tumors. Hum
Pathol. 43:229–237. 2012.PubMed/NCBI
|
|
20
|
Peralta-Leal A, Rodríguez-Vargas JM,
Aguilar-Quesada R, et al: PARP inhibitors: new partners in the
therapy of cancer and inflammatory diseases. Free Radic Biol Med.
47:13–26. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Galluzzi L, Vitale I, Abrams JM, et al:
Molecular definitions of cell death subroutines: recommendations of
the Nomenclature Committee on Cell Death 2012. Cell Death Differ.
19:107–120. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Langelier M-F and Pascal JM: PARP-1
mechanism for coupling DNA damage detection to poly(ADP-ribose)
synthesis. Curr Opin Struct Biol. 23:134–143. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang L, Zhang HW, Hu R, et al: Wogonin
induces G1 phase arrest through inhibiting Cdk4 and cyclin D1
concomitant with an elevation in p21Cip1 in human cervical
carcinoma HeLa cells. Biochem Cell Biol. 87:933–942. 2009.
View Article : Google Scholar
|
|
24
|
Kim MY: Nitric oxide triggers apoptosis in
A375 human melanoma cells treated with capsaicin and resveratrol.
Mol Med Rep. 5:585–591. 2012.PubMed/NCBI
|
|
25
|
Majumdar AP, Banerjee S, Nautiyal J, et
al: Curcumin synergizes with resveratrol to inhibit colon cancer.
Nutr Cancer. 61:544–553. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Soria G and Gottifredi V: PCNA-coupled p21
degradation after DNA damage: The exception that confirms the rule?
DNA Repair (Amst). 9:358–364. 2010. View Article : Google Scholar
|
|
27
|
Beckerman R, Donner AJ, Mattia M, et al: A
role for Chk1 in blocking transcriptional elongation of p21 RNA
during the S-phase checkpoint. Genes Dev. 23:1364–1377. 2009.
View Article : Google Scholar
|
|
28
|
Adon AM, Zeng X, Harrison MK, et al: Cdk2
and Cdk4 regulate the centrosome cycle and are critical mediators
of centrosome amplification in p53-null cells. Mol Cell Biol.
30:694–710. 2010. View Article : Google Scholar : PubMed/NCBI
|