Introduction
Chronic pulmonary heart disease is a serious disease
that poses a risk to human life and health, which is associated
with the pathology of chronic obstructive pulmonary disease (COPD)
(1). Hypoxic pulmonary
hypertension (HPH) is associated with various types of lung/heart
diseases, and is also prevalent in COPD (2). Pulmonary vascular proliferation and
remodeling are considered to be central to the pathogenesis of HPH,
during which, the proliferation of pulmonary artery smooth muscle
cells (PASMCs) has an important role (3).
Previous studies have demonstrated that following
exposure to low oxygen levels (hypoxia), PASMCs exhibit enhanced
proliferative ability, as compared with cells exposed to normal
oxygen levels (normoxia) (4–6). In
our previous studies (7,8), hypoxia-inducible factor (HIF)-1α was
shown to transcriptionally activate vascular endothelial growth
factor (VEGF) in PASMCs under hypoxic conditions. HIF-1α, which is
a subunit of the transcription factor hypoxia-inducible factor-1 (a
heterodimer comprised of an alpha and a beta subunit), functions as
a master regulator of the cellular and systemic homeostatic
response to hypoxia via activation of the transcription of several
genes, including those involved in energy metabolism, angiogenesis
and apoptosis (9,10).
Small ubiquitin-like modifier (SUMO) is a
post-translational modification system, which functions in a manner
similar to ubiquitin and regulates various cellular processes,
including nuclear transport, transcriptional regulation, apoptosis
and protein stability (11). In
addition, SUMOylation has an important role in the regulation of
HIF-1α under hypoxic conditions (12,13).
Conversely, SUMO can be removed from proteins by SUMO-specific
protease (SENP) (14,15), this process is known as protein
deSUMOylation. SENP-1, which is a member of the SENP family, has
been reported as essential for the stability and activity of HIF-1α
(16). Furthermore, SENP-1 levels
have been shown to increase in response to oxygen deprivation
(14). Consequently, the present
study aimed to investigate whether SENP-1 was able to regulate
HIF-1α via deSUMOylation, thus resulting in hypoxia-induced
proliferation of PASMCs.
Materials and methods
Isolation and culture of rat PASMCs
Primary PASMCs were isolated and cultured from rat
intrapulmonary arteries, according to previous methods (17). Briefly, two adult male
Sprague-Dawley rats (weight, 150–180 g), obtained from the Shanghai
Laboratory Animal Center (Shanghai, China), were maintained under a
12-h light/dark cycle at 25°C and 45% humidity, with ad
libitum access to food and water. The animal protocols of the
present study were approved by the Ethical Committee of the Hunan
Province Geriatric Hospital (Changsha, China).
The rats were sacrificed by ethyl carbamate overdose
(1 g/kg; intraperitoneal injection). Distal intrapulmonary arteries
were carefully dissected from the lungs of the rats in a biological
safety cabinet. Adventitia and endothelium were removed from the
isolated pulmonary arteries using a scalpel blade, after which the
remaining smooth muscle was cut into sections (<1
mm2), which were further digested with 2.0 mg/ml
collagenase, 0.5 mg/ml elastase (both Sigma-Aldrich, St. Louis, MO,
USA) and 1.0 mg/ml bovine serum albumin (Dingguo Changsheng
Biotechnology, Co., Ltd., Beijing, China) at 37°C in order to
produce a single cell suspension of PASMCs. Subsequently, the
PASMCs were cultured for 3–4 days in smooth muscle growth media
[Dulbecco's modified Eagle's medium (DMEM)/F12; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA], supplemented with 15%
fetal bovine serum (FBS; Gibco), 1% streptomycin and 1% penicillin
(Dingguo Changsheng Biotechnology, Co., Ltd.). PASMCs were cultured
in a hypoxic chamber (1% O2) or a normoxic chamber (21%
O2) for 2, 6, 12, 24 or 48 h. The cellular purity of the culture
was evaluated by observing the morphological appearance under a
phase-contrast microscope, and the cells underwent
immunofluorescence staining with mouse anti-α-smooth muscle actin
monoclonal antibody (AA132; Beyotime Institute of Biotechnology,
Haimen, China), which was observed under a fluorescence microscope
(IX73-A22FL/PH; Olympus Corporation, Tokyo, Japan). The cells used
for subsequent experiments were obtained from passages 3–5.
RNA extraction and semi-quantitative
polymerase chain reaction (PCR)
Total RNA was isolated from the cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). cDNA was synthesized from the total RNA using a Reverse
Transcription system (Thermo Fisher Scientific, Inc., Pittsburgh,
PA, USA), according to the manufacturer's protocol. Briefly, 1
μg RNA was mixed with 1 μl Oligo (dT)18 primer and
nuclease-free water to a total volume of 12 μl, then
centrifuged at 5000 × g for 30 sec at 4°C, prior to incubation at
65°C for 5 min and chilling on ice. The following components were
then added to the reaction mixture to a final volume of 20
μl: 4 μl 5X Reaction Buffer, 1 μl RiboLock
RNase Inhibitor, 2 μl 10 mM dNTP Mix and 1 μl
RevertAid M-MuLV Reverse Transcriptase. The reaction mixture was
incubated for 60 min at 42°C, after which the reaction was
terminated by incubation at 70°C for 5 min. Subsequently, 2
μl cDNA was amplified by PCR in a reaction mixture
containing 2 μl cDNA, 2 μl each of sense and
antisense primers, 25 μl 2X Es Tap Master Mix (CWBio,
Beijing, China) and 19 μl RNase-Free Water (Dingguo
Changsheng Biotechnology Co., Ltd.,), in a final volume of 50
μl. PCR was performed using the Mastercycler pro S
(Eppendorf, Hamburg, Germany) and the following cycling conditions:
Initialization step (94°C for 2 min); denaturation step (94°C for
30 sec); annealing step (30 sec); elongation step (72°C for 30
sec); and final elongation step (72°C for 2 min). β-actin was
amplified in parallel as an internal control. The primers were
purchased from Sangon Biotech Co., Ltd. (Shanghai, China). The
primer sequences, annealing temperatures and cycle numbers are
presented in Table I. The PCR
products were separated by 1% agarose gel electrophoresis (100 V;
30 min) and images of the gels were captured using the ChemiDoc
XRS+ system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Densitometric analyses were performed using ImageJ 2x software
(National Institutes of Health, Bethesda, MD, USA).
 | Table IPrimers used to amplify rat SENP-1,
HIF-1α, VEGF and β-actin mRNA, and the associated polymerase chain
reaction procedures. |
Table I
Primers used to amplify rat SENP-1,
HIF-1α, VEGF and β-actin mRNA, and the associated polymerase chain
reaction procedures.
| Target gene | Oligonucleotide
primers (5′-3′) | Annealing temperature
(°C) | Cycles | Produc size (bp) |
|---|
| HIF-1α | Forward
TGTGGATAGCGATATGGTCAA | 53.1 | 31 | 218 |
| Reverse
CTCTTTCCTGCTCTGTCTGGT | | | |
| SENP-1 | Forward
TGCAGTGCTTGATTCCGTAG | 63.1 | 32 | 405 |
| Reverse
TTCTTTGTCCAGCGTTTCAC | | | |
| VEGF | Forward
CATCCACCATGCACTTGCTGT | 55.8 | 28 | 178 |
| Reverse
GGCTGCTCCAAACTCCTTCCA | | | |
| β-actin | Forward
AGCCATGTACGTAGCCATCC | 65 | 28 | 228 |
| Reverse
CTCTCAGCTGTGGTGGTGAA | | | |
Western blot analysis
Total protein was extracted from PASMCs using
radioimmunoprecipitation assay lysis buffer supplemented with
phosphatase inhibitors (Roche Diagnostics GmbH, Mannheim, Germany).
Nucleoprotein was obtained using the Nucleoprotein Protein
Extraction kit (Beyotime Institute of Biotechnology). The protein
concentration of the cell lysates was quantified using
bicinchoninic acid protein assay (Beyotime Institute of
Biotechnology). Approximately 40 μg protein was separated by
8% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
was transferred to polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked in 5%
nonfat milk with 0.2% Tween-20, and were then incubated with the
following primary antibodies: Anti-α-smooth muscle actin (α-SMA;
1:500), anti-histone 3 (1:500; AH433; Beyotime Institute of
Biotechnology); anti-SENP-1 (1:300; sc-67074), anti-VEGF (1:500;
sc-507; Santa Cruz Biotechnology, Inc., Dallas, TX, USA);
anti-β-actin (1:500; BM0627; Wuhan Boster Biological Technology,
Ltd., Wuhan, China); and anti-HIF-1α (1:1,000; NB100–479; Novus
International, St Charles, MO, USA). The membranes were then
blotted with horseradish peroxidase-conjugated goat anti-rabbit
(1:4,000; sc-2004) and goat anti-mouse (1:4,000; sc-2005)
immunoglobulin G secondary antibodies (Santa Cruz Biotechnology,
Inc.). Subsequently, the membranes were extensively washed with
phosphate-buffered saline (PBS) containing 0.1% Tween and were
exposed to an enhanced chemiluminescent reagent (Beyotime Institute
of Biotechnology). After scanning the X-ray film, the optical
density of the immunoblots was calculated using ImageJ 2x software.
Histone 3 was employed as the loading control for the nucleoprotein
and β-actin was used as the loading control for total protein.
Immunofluorescence assay
PASMCs were fixed in medium containing 4%
paraformaldehyde for 30 min. The cells were permeabilized using
0.2% Triton X-100 for 2 min, and were blocked with 5% bovine serum
albumin (Dingguo Changsheng Biotechnology, Co., Ltd.) for 30 min.
The cells were then incubated with α-SMA antibody (Beyotime
Institute of Biotechnology) at 4°C overnight, were washed with PBS,
and were incubated with streptavidin biotin complex-fluorescein
isothiocyanate (Wuhan Boster Biological Technology, Ltd.) for 30
min at 37°C. After counterstaining with
4′,6-diamidino-2-phenylindole (DAPI), immunofluorescence was
observed and images were captured under a fluorescence microscope
(IX73-A22FL/PH).
Cell Counting kit (CCK)-8 assay
A 100 μl PASMCs suspension (1×104
cells/well) was seeded into 96-well plates. Once confluence reached
70%, the cells were synchronized in DMEM/F12 without FBS for 24 h.
Subsequently, the medium was replaced with DMEM containing FBS and
the plates were incubated in a hypoxic or normoxic chamber. The
plates were incubated for an appropriate duration (2, 6, 12, 24 or
48 h) in the incubator (5 duplicate wells for each time point).
Subsequently, 10 μl CCK-8 solution (CCK-8 Assay kit; Wuhan
Boster Biological Technology, Ltd.) was added to each well and the
plates were incubated for 4 h at 37°C. The absorbance was then
measured at a wavelength of 460 nm using a microplate autoreader
(EL309; BioTek Instruments, Inc., Winooski, VT, USA). All of the
experiments were repeated at least 3 times.
5-ethynyl-2′-deoxyuridine (EdU) cell
proliferation assay
PASMCs were seeded into 96 well plates
(1×104 cells/well). Following synchronization in
DMEM/F12 without FBS for 24 h, the cells were separately incubated
in a normoxic or hypoxic chamber for an appropriate length of time
(2, 6, 12, 24 or 48 h). Subsequently, 100 μl EdU (50
μM; Guangzhou Ribobio Co., Ltd., Guangzhou, China) was added
to the wells and the plates were incubated in the appropriate
chamber for 2 h, according to the manufacturer's protocol. After
EdU staining, the cells were counterstained with DAPI. EdU-positive
cells were imaged randomly by fluorescence microscopy, and the
EdU-positive cells were randomly counted in 5 fields. The ratio of
positive cells was calculated.
SENP-1 short hairpin (sh)RNA lentiviral
synthesis
SENP-1 shRNA and scrambled shRNAs were designed and
synthesized by Shanghai Genechem Co., Ltd. (Shanghai, China). The
lentiviral particles were also produced by Shanghai Genechem Co.,
Ltd. Primary cultured PASMCs in DMEM/F12 medium were infected with
lentiviral particles for 24 h. Cells were then cultured with
puromycin (4 μM; Shanghai Genechem Co., Ltd.) for 48 h, in
order to kill the uninfected cells and to ensure >95% infection
of cells. Four types of shRNA were used, as follows:
SENP-1/GV118-RNAi-LV#1, SENP-1/GV118-RNAi-LV#2,
SENP-1/GV118-RNAi-LV#3, SENP-1/GV118-RNAi-LV#4 (Table II). The knockdown efficiency of
each clone was determined using western blotting. The clones with
the maximum knockdown efficiency were selected for subsequent
experiments. As a control, cells were infected with lentiviral
particles containing scrambled shRNAs. Uninfected cells were used
as a blank group.
| Table IISequence of SENP-1 shRNA. |
Table II
Sequence of SENP-1 shRNA.
| SENP-1 shRNA | shRNA sequence
(5′-3′) |
|---|
|
SENP-1/GV118-RNAi-LV#1 |
GACCTCAAGTGGATTGTCAAACTCGAGTTTGACAATCCACTTGAGGTC |
|
SENP-1/GV118-RNAi-LV#2 |
GACCATCACACGCAAAGACATCTCGAGATGTCTTTGCGTGTGATGGTC |
|
SENP-1/GV118-RNAi-LV#3 |
AACCTTTGGTCAAAGTGCAAACTCGAGTTTGCACTTTGACCAAAGGTT |
|
SENP-1/GV118-RNAi-LV#4 |
AACCATCTAAACTGGCTCAATCTCGAGATTGAGCCAGTTTAGATGGTT |
Statistical analysis
All data were analyzed using SPSS 10.0 software
(SPSS Inc., Chicago, IL, USA). The statistical significance of the
differences between the mean values was determined using one-way
analysis of variance. The data were examined for equality of
variance, followed by either Fisher test (if variances were equal)
or approximate variance F test/Welch method (if variances were
unequal). To compare pairwise difference, Fisher's least
significant difference test was used. In the case of homogeneity of
variance, Dunnett's T3 was used. Data are presented as the mean ±
standard deviation. P<0.05 was considered to indicate a
statistically significant difference.
Results
Morphology and identification of
PASMCs
Following isolation and culture for 3–4 days, PASMCs
dissociated from the tissue exhibited numerous shapes, an abundant
cytoplasm and oval-like nuclei (Fig.
1A). After 7–10 days, PASMCs were spindle-shaped, arranged in
bundles, and grew in a typical 'hill-and-valley' pattern at high
confluence (Fig. 1B). To further
characterize these cells, immunofluorescence was used to detect
α-SMA. A strong green fluorescence was detected in the cytoplasm,
and >95% of cells were positively stained (Fig. 1C). These results indicate that the
separated cells were PASMCs.
 | Figure 1Morphology and identification of
PASMCs. (A) Isolated cells exhibited numerous shapes, a large
cytoplasm and oval-like nuclei at low confluence (magnification:
left, 100×; right, 200×). (B) These cells grew in a typical
'hill-and-valley' pattern at high confluence (>70%;
magnification: left, 40×; right, 100×). (C) Immunofluorescence
staining of α-smooth muscle actin revealed that >95% of cells
were positively stained (magnification: top, 100×; bottom, 200×).
PASMCs, pulmonary artery smooth muscle cells; FITC, fluorescein
isothiocyanate; DAPI, 4′,6-diamidino-2-phenylindole. |
Hypoxia promotes the proliferative
ability of PASMCs
To determine whether hypoxia was able to influence
the proliferative ability of PASMCs, a CCK-8 assay was used to
measure the proliferation of cells cultured in hypoxic and normoxic
conditions. As shown in Fig. 2,
the optical density (OD) value of each group increased in a
time-dependent manner. In the hypoxic group, the OD value at 12, 24
and 48 h was markedly higher than at 2 and 6 h (P<0.05). After
reaching a peak at 24 h, proliferative speed tended to slow,
resulting in a flat growth curve. PASMCs cultured in normoxic
conditions exhibited a slower multiplication speed, as compared
with the hypoxic group. There were statistical differences between
hypoxic and normoxic groups at 12, 24 and 48 h (P<0.05).
Similar results were obtained from the EdU staining
assay (Fig. 3A). Under normoxic
conditions, a small proportion of PASMCs were labeled with EdU.
Although no obvious distinction was observed in the hypoxic group
at 2 or 6 h compared with the normoxic group, fluorescence was
markedly increased in hypoxic PASMCs at 12, 24 and 48 h. There was
a statistical difference in EdU positive staining between 24, 48
and 12 h (P<0.05), whereas no significant difference was
detected between 24 and 48 h (Fig.
3B).
 | Figure 3(A) Proliferative ability of pulmonary
artery smooth muscle cells in the hypoxic and normoxic groups, as
detected by EdU assay (magnification, ×100). The number of
EdU-positive cells increased as hypoxic exposure time was
prolonged. (B) The positive rates were as follows: Normoxia,
6.86±0.25%; hypoxia 2 h, 7.08±0.38%; hypoxia 6 h, 9.11±0.24%;
hypoxia 12 h, 29.99±1.48%; hypoxia 24 h, 52.12±0.89%; hypoxia 48 h,
53.69±0.64%. Data are expressed as the mean ± standard deviation,
n=5. *P<0.05. EdU, 5-ethynyl-2′-deoxyuridine; DAPI,
4′,6-diamidino-2-phenylindole. |
Hypoxia-induced proliferation of PASMCs
is mediated by SENP-1 via the HIF-1-VEGF-dependent pathway
To investigate whether the effects on proliferation
were mediated by deSUMOylation of HIF-1α, semi-quantitative PCR was
performed to analyze the mRNA expression levels of SENP-1, HIF-1α
and VEGF in PASMCs (Fig. 4). The
mRNA expression levels of SENP-1 at 2 h in the hypoxic group were
increased, as compared with in the normoxic group (P<0.05). In
addition, the mRNA expression levels of SENP-1 were upregulated at
12 and 24 h compared with at 2 and 6 h (P<0.05). The mRNA
expression levels of HIF-1α in the hypoxic group at 2, 6 and 24 h
were increased, as compared with in the normoxic group. The mRNA
expression levels of VEGF in the hypoxic group were examined at 2,
6, 12 and 24 h. After PASMCs were exposed to hypoxia for 2 h, a
marked increased in VEGF mRNA expression was detected. In addition,
a further increase in VEGF mRNA expression was observed as the
hypoxic treatment time was prolonged.
Western blotting was used to detect protein
expression levels in the cells under normoxic and hypoxic
conditions (Fig. 5). Following 2 h
of hypoxic exposure, the expression levels of SENP-1 were elevated,
as compared with the normoxic group. Furthermore, SENP-1 exhibited
a time-dependent increase, and the difference between 12 and 24 h,
and 2 and 6 h was markedly different. Similar to SENP-1, hypoxic
treatment led to increased expression levels of HIF-1α and VEGF. As
treatment time was prolonged, the expression levels reached a peak
at 12 or 24 h, which was markedly higher, as compared with 2 or 6
h.
To demonstrate the effects of downregulation of
SENP-1 in hypoxic conditions, shRNA was used to study the effects
of SENP-1 knockdown on PASMCs. Since the shRNA lentivirus was
labeled with green fluorescent protein, PASMCs infected with the
lentivirus could easily by identified by fluorescence microscopy
(Fig. 6A). Subsequently, western
blotting was conducted to confirm the efficiency of SENP-1 shRNA
targeting. As shown in Fig. 6B,
SENP-1/GV118-RNAi-LV#2 exhibited the maximum reduction of SENP-1
expression, and was therefore selected for subsequent
experiments.
Following knockdown of SENP-1 (Fig. 7A), PASMCs were incubated in a
hypoxic chamber for 12 h. Subsequently, total RNA was extracted
from the cells and semi-quantitative PCR was performed to detect
the mRNA expression levels of HIF-1α and VEGF. The mRNA expression
levels of HIF-1α and VEGF were significantly downregulated in the
SENP-1 shRNA group, as compared with the scramble and blank groups
(Fig. 7B and C; P<0.05). In
addition, no difference was observed between the scrambled and
blank groups. As presented in Fig.
7D, the protein expression levels of HIF-1α and VEGF were also
downregulated in the SENP-1 shRNA group, as compared with in the
scrambled and blank groups. These results indicate that knockdown
of SENP-1 leads to a marked decrease in the mRNA and protein
expression levels of HIF-1α and VEGF.
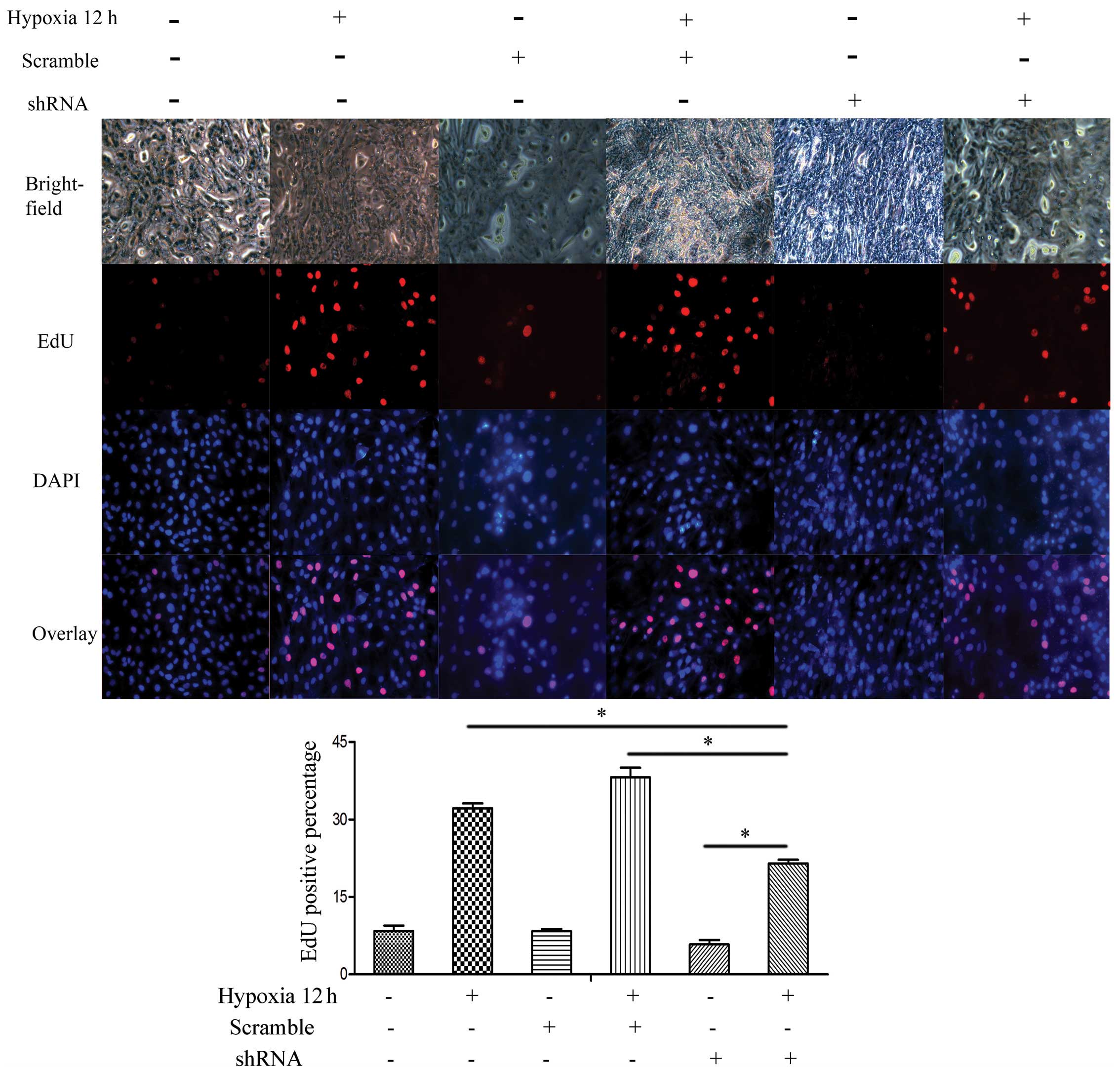
To further evaluate the effects of SENP-1 knockdown
on PASMCs, an EdU assay was conducted to compare the proliferative
ability among the SENP-1 shRNA, scrambled and blank groups. As
shown in Fig. 8, compared with the
scrambled and blank groups, there was a declining ratio of
EdU-positive cells in the SENP-1 shRNA group (P<0.05).
Furthermore, the number of EdU-positive cells in the SENP-1 shRNA
group was significantly higher at 12 h (hypoxia) than at 0 h
(normoxia) (P<0.05). Consequently, these results suggest that
knockdown of SENP-1 may exhibit an inhibitory effect on the
proliferation of PASMCs in a hypoxic environment.
Discussion
HIF-1 is a master regulator of oxygen homeostasis,
which is associated with numerous biological processes, starting in
early embryonic development and extending into adult life (18). In response to hypoxia, HIF-1α, a
subunit of HIF-1, dimerizes with HIF-1β, resulting in the formation
of activated HIF-1 protein, which can bind to enhancer sequences in
target genes that contain the core RCGTG binding motif (19,20).
VEGF is a critical downstream target gene of HIF-1, which is
essential for HIF-1-mediated neovascularization (21,22).
Therefore, the HIF-1-VEGF-dependent pathway is considered to have
an important role in the proliferation of PASMCs.
Protein SUMOylation is regarded as an important
mechanism of post-translational modification (12). Hypoxia is able to induce the
SUMOylation of HIF-1α, thus mediating its activity and stability
(13). However, the role of
SUMOylation in the regulation of HIF-1α stability remains
controversial (12). Bae et
al (23) demonstrated that the
protein level and transcriptional activity of HIF-1α were
upregulated by SUMO-1. In addition, RSUME has been reported to
participate in HIF-1α stabilization by enhancing HIF-1α SUMOylation
(24). However, a previous study
indicated that SUMO modification of HIF-1α was able to repress its
transcriptional activity (25). In
addition, SUMOylation of HIF-1α, which binds to von Hippel-Lindau
protein in a hydroxyl proline-independent manner, has been shown to
lead to ubiquitination and proteasomal degradation of HIF-1α. The
opposite process of SUMOylation, namely deSUMOylation, has also
been suggested to exert huge influence in stabilizing and
activating HIF-1α (26,27). Consequently, it is of great
significance to determine how HIF-1α is post-transcriptionally
modified by SENP-1 in PASMCs under hypoxic conditions.
The present study demonstrated that upon exposure to
hypoxia, the proliferation of PASMCs increased in a time-dependent
manner. Simultaneously, the mRNA and protein expression levels of
SENP-1, HIF-1α and VEGF were upregulated. Following knockdown of
SENP-1, hypoxia-induced proliferation of PASMCs was markedly
inhibited, and HIF-1α and VEGF expression was decreased at both the
mRNA and protein level. In conclusion, hypoxia may significantly
promote the proliferation of rat PASMCs, which is regulated by
SENP-1, via the HIF-1-VEGF-dependent pathway. Therefore, targeting
SENP-1-mediated deSUMOylation may be considered an effective method
for the suppression of the pathogenesis of HPH.
Acknowledgments
The authors acknowledge the work of all
investigators involved in the present study. This study was
supported by a grant from the National Natural Science Foundation
of China (grant no. 30971329).
References
|
1
|
Vestbo J, Hurd SS, Agusti AG, Jones PW,
Vogelmeier C, Anzueto A, Barnes PJ, Fabbri LM, Martinez FJ,
Nishimura M, et al: Global strategy for the diagnosis, management,
and prevention of chronic obstructive pulmonary disease: GOLD
executive summary. Am J Respir Crit Care Med. 187:347–365. 2013.
View Article : Google Scholar
|
|
2
|
Wright JL, Levy RD and Churg A: Pulmonary
hypertension in chronic obstructive pulmonary disease: Current
theories of pathogenesis and their implications for treatment.
Thorax. 60:605–609. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jin H, Wang Y, Zhou L, Liu L, Zhang P,
Deng W and Yuan Y: Melatonin attenuates hypoxic pulmonary
hypertension by inhibiting the inflammation and the proliferation
of pulmonary arterial smooth muscle cells. J Pineal Res.
57:442–450. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sarkar J, Gou D, Turaka P, Viktorova E,
Ramchandran R and Raj JU: MicroRNA-21 plays a role in
hypoxia-mediated pulmonary artery smooth muscle cell proliferation
and migration. Am J Physiol Lung Cell Mol Physiol. 299:L861–L871.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen B, Calvert AE, Cui H and Nelin LD:
Hypoxia promotes human pulmonary artery smooth muscle cell
proliferation through induction of arginase. Am J Physiol Lung Cell
Mol Physiol. 297:L1151–L1159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Growcott EJ, Banner KH and Wharton J:
Hypoxia amplifies the proliferative capacity of distal human
pulmonary artery smooth-muscle cells. Chest. 128(6 Suppl):
600S–601S. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li QF and Dai AG: Hypoxia-inducible
factor-1 alpha regulates the role of vascular endothelial growth
factor on pulmonary arteries of rats with hypoxia-induced pulmonary
hypertension. Chin Med J (Engl). 117:1023–1028. 2004.
|
|
8
|
Li QF and Dai AG: Hypoxia inducible
factor-1 alpha correlates the expression of heme oxygenase 1 gene
in pulmonary arteries of rat with hypoxia-induced pulmonary
hypertension. Acta Biochim Biophys Sin (Shanghai). 36:133–140.
2004. View Article : Google Scholar
|
|
9
|
Zhang R, Wu Y, Zhao M, Liu C, Zhou L, Shen
S, Liao S, Yang K, Li Q and Wan H: Role of HIF-1alpha in the
regulation ACE and ACE2 expression in hypoxic human pulmonary
artery smooth muscle cells. Am J Physiol Lung Cell Mol Physiol.
297:L631–L640. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rey S and Semenza GL: Hypoxia-inducible
factor-1-dependent mechanisms of vascularization and vascular
remodelling. Cardiovasc Res. 86:236–242. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saitoh H and Hinchey J: Functional
heterogeneity of small ubiquitin-related protein modifiers SUMO-1
versus SUMO-2/3. J Biol Chem. 275:6252–6258. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu B and Shuai K: Regulation of the
sumoylation system in gene expression. Curr Opin Cell Biol.
20:288–293. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yee Koh M, Spivak-Kroizman TR and Powis G:
HIF-1 regulation: Not so easy come, easy go. Trends Biochem Sci.
33:526–534. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu Y, Zuo Y, Zhang H, Kang X, Yue F, Yi Z,
Liu M, Yeh ET, Chen G and Cheng J: Induction of SENP1 in
endothelial cells contributes to hypoxia-driven VEGF expression and
angiogenesis. J Biol Chem. 285:36682–36688. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hickey CM, Wilson NR and Hochstrasser M:
Function and regulation of SUMO proteases. Nat Rev Mol Cell Biol.
13:755–766. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gu J, Fan Y, Liu X, Zhou L, Cheng J, Cai R
and Xue S: SENP1 protects against myocardial ischaemia/reperfusion
injury via a HIF1α-dependent pathway. Cardiovasc Res. 104:83–92.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Shimoda LA and Sylvester JT:
Capacitative calcium entry and TRPC channel proteins are expressed
in rat distal pulmonary arterial smooth muscle. Am J Physiol Lung
Cell Mol Physiol. 286:L848–L858. 2004. View Article : Google Scholar
|
|
18
|
Semenza GL: Hypoxia-inducible factor 1:
Control of oxygen homeostasis in health and disease. Pediatr Res.
49:614–617. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ortiz-Barahona A, Villar D, Pescador N,
Amigo J and del Peso L: Genome-wide identification of
hypoxia-inducible factor binding sites and target genes by a
probabilistic model integrating transcription-profiling data and in
silico binding site prediction. Nucleic Acids Res. 38:2332–2345.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mole DR, Blancher C, Copley RR, Pollard
PJ, Gleadle JM, Ragoussis J and Ratcliffe PJ: Genome-wide
association of hypoxia-inducible factor (HIF)-1alpha and HIF-2alpha
DNA binding with expression profiling of hypoxia-inducible
transcripts. J Biol Chem. 284:16767–16775. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Oladipupo S, Hu S, Kovalski J, Yao J,
Santeford A, Sohn RE, Shohet R, Maslov K, Wang LV and Arbeit JM:
VEGF is essential for hypoxia-inducible factor-mediated
neovascularization but dispensable for endothelial sprouting. Proc
Natl Acad Sci USA. 108:13264–13269. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cucina A, Borrelli V, Randone B, Coluccia
P, Sapienza P and Cavallaro A: Vascular endothelial growth factor
increases the migration and proliferation of smooth muscle cells
through the mediation of growth factors released by endothelial
cells. J Surg Res. 109:16–23. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bae SH, Jeong JW, Park JA, Kim SH, Bae MK,
Choi SJ and Kim KW: Sumoylation increases HIF-1alpha stability and
its transcriptional activity. Biochem Biophys Res Commun.
324:394–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carbia-Nagashima A, Gerez J, Perez-Castro
C, Paez-Pereda M, Silberstein S, Stalla GK, Holsboer F and Arzt E:
RSUME, a small RWD-containing protein, enhances SUMO conjugation
and stabilizes HIF-1alpha during hypoxia. Cell. 131:309–323. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Berta MA, Mazure N, Hattab M, Pouysségur J
and Brahimi-Horn MC: SUMOylation of hypoxia-inducible factor-1alpha
reduces its transcriptional activity. Biochem Biophys Res Commun.
360:646–652. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cheng J, Kang X, Zhang S and Yeh ET:
SUMO-specific protease 1 is essential for stabilization of
HIF1alpha during hypoxia. Cell. 131:584–595. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yeh ET: SUMOylation and De-SUMOylation:
Wrestling with life's processes. J Biol Chem. 284:8223–8227. 2009.
View Article : Google Scholar :
|