Introduction
Breast cancer is reported to be the most common type
of cancer diagnosed among women (1). Despite advances in medicine, the
disease remains a substantial health problem worldwide. According
to statistics, 1,500,000 cases of breast cancer were diagnosed in
2010, representing almost one quarter of all the cases of cancer
diagnosed in women (2). Primary
liver cancer, particularly hepatocellular carcinoma (HCC),
continues to be a growing global health problem, and has become the
third most common cause of cancer-associated mortality worldwide,
accounting for a mortality rate of >800,000/year (2,3). The
5-year survival rate of HCC is <10% (4). The results of standard chemotherapy
and radiotherapy for the treatment of patients with breast cancer
and HCC have remained unsatisfactory (5,6).
Despite efforts, no drugs have been manufactured for the
satisfactory treatment of breast cancer and HCC in the past decade.
Therefore, more effective alternative therapies or drugs with low
side effect are required for the treatment of breast cancer and
HCC.
Natural products have gradually became one of the
most productive strategies in the development of antitumor agents
(6), which possess potent
cytotoxic abilities with fewer adverse effects. Neem leaf aqueous
extract induces granulosa cell apoptosis via the
mitochondria-caspase-mediated pathway (7). Parmotrema reticulatum, a
tropical lichen, increases the B cell lymphoma-2 (Bcl-2)-associated
X protein (Bax)/Bcl-2 ratio and activates the caspase family
leading to apoptosis in MCF-7 cells (8). Cordyceps militaris (CM), an
entomopathogenic fungus belonging to the class ascomycetes, is
usually used as a traditional tonic in China and East Asia
(9). A complex composition has
been observed in the CM fruiting body, which is responsible for its
various pharmacological activities (10). CM aqueous extract induces apoptotic
MDA-MB-231 cell death via regulation of the phosphoinositide
3-kinase/AKT-associated mitochondrial pathway (11). CM inhibits B16-F10-xenograft tumor
growth in C57BL/6 mice, associated with its angiogenic property
(12). Due to activation of the
caspase-associated pathway, CM aqueous extract also suppresses
human premyelocytic leukemia cell growth (13). Although the cytotoxicity of CM
towards MCF-7 and HepG2 cells has been reported (14,15),
the underlying mechanisms remain to be elucidated.
As an energy-dependent process, during apoptosis,
living cells are involved in their own death in an organized
manner, which is associated with various signaling pathways
(16). Mitochondrial apoptosis,
one of three death signaling pathways (17), is accompanied by mitochondrial
depolarization, abnormal expression of members of the Bcl-2 family,
the over-release of cytochrome c, and caspase-3 activation
(18,19). Caspase-3 is generally considered to
be an important effector protease during apoptosis (18), the proteolytic maturation of which
is catalyzed by initiator caspases (caspase-8, -9 and -10)
(20). The auto-catalytic
activation of procasapase-8 in extrinsic apoptosis leads to the
decrease of mitochondrial membrane permeability (Δψm) (21,22).
The purpose of the present study was to investigate
the in vitro and in vivo pro-apoptotic effects of CM
on HCC and breast cancer. The results revealed that CM induced
MCF-7 and HepG2 cell apoptosis, predominantly through the
caspase-dependent mitochondrial pathway. The results of the present
study indicated the potential for the addition of CM to the list of
possible agents for the treatment of HCC and breast cancer.
Materials and methods
CM extract preparation
Extraction of the CM fruiting body (purchased from
Qianxiang Co., Ltd., Shenyang, China) was performed at 45°C for 3
h, followed by extraction at 80°C for another 3.5 h in double
distilled water (D.D. water). The merging supernatant was
concentrated in an R1002B evaporator (Shanghai Shensheng Technology
Co., Ltd., Shanghai, China) under reduced pressure (<10 kPa),
and was further freeze-dried to produce a solid aqueous extract.
Analysis of the data revealed that the CM fruiting body water
extract contained 29.04% polysaccharides, 20.45% total proteins,
6.01% cordycepic acid, 0.204% adenosine and 0.347% cordycepin. The
polysaccharides were measured via phenol-sulfuric acid method
(23), the total proteins were
analysed via Kjeldahl method (24), and the cordycepic acid, adenosine
and cordycepin were analysed via high performance liquid
chromatography (25).
Cell culture
HepG2 cells (human HCC line; HB-8065) and MCF-7
cells (human breast carcinoma cell line; HTB-22) were cultured in
Dulbecco's modified Eagle's medium (DMEM) supplemented with 10%
fetal bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin
(all obtained from Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) under a humidified atmosphere containing 5%/95%
CO2/air at 37°C. The cultured medium was refreshed every
3 days. The cells were ready for treatment when the confluence of
cells in the plates reached. All reagents used for cell culture
were obtained from Invitrogen; Thermo Fisher Scientific, Inc.
Cell viability analysis
The cell viability was measured using a 3-(4,5)-dimethylthiahiazo(-z-y1)-3,5-di-pheny-tetrazolium-romide
(MTT; Sigma-Aldrich, St. Louis, MO, USA) assay (26). The MCF-7 and HepG2 cells were
seeded into 96-well plates at a density of 5,000 per well for 24 h.
Following treatment with CM at doses of 0.1, 0.2, 0.4, 1.0 and 2.0
mg/ml for 24 h, the cells were incubated with 0.5 mg/ml MTT for 4 h
at 37°C in the dark. Purple formazan crystals were solubilized by
adding 100 µl dimethyl sulfoxide, and the absorbance was measured
using an iMark microplate reader at a wavelength of 490 nm (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Lactate dehydrogenase (LDH) release
analysis
The MCF-7 and HepG2 cells were seeded into 24-well
plates at a density of 5×104 per well. Following
treatment with CM at doses of 0.1, 0.2, 0.4, 1.0 and 2.0 mg/ml for
24 h, the cultured medium was collected and centrifuged at 1,000 ×
g for 5 min at 4°C. The LDH released into the culture medium was
measured using an LDH assay kit (cat. no. 20141205; Nanjing
Jiancheng Bioengineering Institute, Nanjing, China).
Colony formation assays
Crystal violet staining was performed to examine the
colony formation. Briefly, the MCF-7 and HepG2 cells were seeded
into 6-well plates at a density of 5×104 cells/well. The
cells were treated with CM at 0.1, 0.2, 0.4, 1.0 and 2.0 mg/ml for
7 days. The medium was replaced, either with DMEM only or the CM in
complete DMEM, every 2 days. Following treatment, the cells were
fixed in 75% methanol for 10 min at 4°C and stained with 0.1%
crystal violet (Sigma-Aldrich) for 30 min. Images were subsequently
captured.
Apoptosis analysis via Hoechst
staining
The MCF-7 and HepG2 cells were seeded into 6-well
plates at a density of 2×105 cells/well. The cells were
treated with CM at 0.5, 1.0 and 2.0 mg/ml for 24 h. The treated
cells were then incubated with Hoechst 33342 (5 µg/ml; BD
Biosciences, San Jose, CA, USA) for 30 min at 37°C in the dark.
Following three washes with phosphate-buffered saline (PBS), images
of the altered fluorescent color in the mitochondria were captured
using a fluorescent microscope (magnification, x20; charge-coupled
device camera; TE2000; Nikon Corporation, Tokyo, Japan). The
percentage of apoptotic cells was analyzed by measuring the
fluorescence intensity using Image J software (rsb.info.nih.gov/ij/download.html) and
expressed as the ratio of red to green fluorescence intensity.
Migration assay
The cells were plated in 6-well plates
(4×104 cells/well) and cultured to >90% confluence,
following which the cell layer was scraped with a p200 pipette tip
(Shanghai Jingke Scientific Instrument Co., Ltd., Shanghai, China).
The cells were treated with CM at 0.5, 1.0 and 2.0 mg/ml for 24 h,
following which the distances of the migrating cells were used to
evaluate the migratory ability of the cells. The width of the wound
was expressed as a percentage of the control group.
Assessment of Δψm
To determine the alterations in Δψm in the cells,
5,5′,6,6′-Tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolylcarbocyanine
iodide (JC-1; Sigma-Aldrich), which selectively enters
mitochondria, was used. The MCF-7 and HepG2 cells were seeded into
6-well plates at a density of 2×105 cells/well. The
cells were treated with CM at doses of 0.5, 1.0 and 2.0 mg/ml for
12 h. The treated cells were then incubated with 2 µM JC-1 at 37°C
for 20 min in the dark. Following three washes with PBS,
alterations in the fluorescent color in the mitochondria were
analyzed using fluorescent microscopy (magnification, ×20;
charge-coupled device camera; TE2000; Nikon Corporation). Red
fluorescence indicated healthy cells with a high Δψm, whereas green
fluorescence indicated apoptotic or unhealthy cells with a low
Δψm.
MCF-7- and HepG2-xenograft tumor
models
The present study was approved by the ethics
committee of Changchun University (Changchun, China). Male
6-week-old BALB/c athymic nude mice, purchased from Weitonglihua
Laboratory Animal Technology Co., Ltd. (Beijing China; SCXK
2012-0001), were used for the in vivo experiments in the
present study. The experimental animal protocol was approved by the
Animal Ethics Committee of Jilin University (Changchun, China). The
mice were housed in groups of two in clear plastic cages, and were
maintained on a 12 h light/dark cycle at 23±1°C with water and food
available ad libitum.
The tumors were generated by harvesting MCF-7 and
HepG2 cells from mid-log phase cultures. A volume of 0.1 ml
(1×108 cells/ml) of MCF-7 or HepG2 cell suspension was
subcutaneously injected into the right side of the waist of each
mouse. After 4 days, when the largest diameter of the tumors
measured 2–3 mm, the mice were randomly divided into two groups
(n=3 each). The mice were administered with 1 g/kg CM (treated
group) or D.D. water (vehicle group) orally every other day
continuously for 2 weeks. The tumor dimensions and body weights
were measured every other day. Tumor volume (mm3) was
estimated using the following equation: Tumor volume
(mm3) = Length × (width)2 × 0.5. At the end
of the experiment, the mice were sacrificed by administration of
200 mg/kg pentobarbital (Sigma-Aldrich). The tumor tissues were
carefully dissected from each mouse and stored at −130°C prior to
western blot analysis.
Western blot analysis
The cells were plated into 6-well plates at a
density of 2×105 cells per well. The following day, the
cells were treated with CM (0.5, 1.0 and 2.0 mg/ml) for 24 h. The
cells or tumor tissues were lysed using radioimmunoprecipitation
assay buffer (Sigma-Aldrich) containing 1% protease inhibitor
cocktail (Sigma-Aldrich) and 2% phenylmethanesulfonyl fluoride
(Sigma-Aldrich). Tumor tissues were homogenized using a ZW-A trace
vibrator (Ronghua Instrument Manufacturing Co., Ltd.). Lysates were
centrifuged at 10,000 × g for 5 mins at 4°C and the concentration
of protein was determined by the Coomassie brilliant blue method
(Nanjing Jiancheng Biotechnology Co., Ltd., Nanjing, China) The
proteins (30 µg) were separated on a 12% SDS-PAGE gel [materials
obtained from Sinopharm Chemical Reagent Co., Ltd. (Shanghai,
China)] and transferred electrophoretically onto nitrocellulose
membranes (0.45 µm; Bio Basic, Inc., Markham, ON, Canada). The
transferred membranes were then blocked with 5% bovine serum
albumin (Sigma-Aldrich) for 4 h at 4°C prior to blotting with the
following primary antibodies at 4°C overnight, at a dilution of
1:1,000: Monoclonal rabbit anti-cleaved poly (ADP-ribose)
polymerase (PARP; cat. no. ab32064; Abcam, Cambridge, UK),
polyclonal rabbit anti-cleaved caspase-3 (cat. no. ab13847; Abcam),
poly-clonal rabbit anti-cleaved caspase-8 (cat. no. ab25901;
Abcam), monoclonal rabbit anti-Bax (cat. no. ab32503; Abcam) and
polyclonal rabbit anti-glyceraldehyde-3-phosphate dehydrogenase
(cat. no. ABS16; EMD Millipore, Billerica, MA,USA). The membranes
were subsequently incubated with horseradish peroxidase-conjugated
mouse anti-rabbit secondary antibody (cat. no. sc-2357; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA). Chemiluminescence was
analyzed using Amersham ECL Western Blotting Detection reagent
(cat. no. RPN2106; GE Healthcare, Buckinghamshire, UK). The
intensities of the bands were quantified by scanning densitometry
using Image J software.
Statistical analysis
All data are expressed as the mean ± standard
deviation and were analyzed using one-way analysis of variance,
followed by post-hoc multiple comparison (Dunn's test). SPSS
software version 16.0 (SPSS, Inc. Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
CM exhibits cytotoxic effects in MCF-7
and HepG2 cells
Dose- and time-dependent reductions in cell
viability were observed in the MCF-7 and HepG2 cells following
incubation with CM. The 24-h half maximal inhibitory concentrations
of CM in the MCF-7 and HepG2 cells were ~1.096 and 0.791 mg/ml,
respectively (P<0.01; Fig. 1A and
B). Following exposure to CM at doses between 0.2 and 2.0 mg/ml
for 24 h, 8.45–16.80% increases in LDH release were observed in the
MCF-7 cells (P<0.05; Fig. 1C).
Similarly, treatment with CM for 24 h (0.2–2.0 mg/ml) enhanced LDH
release in the HepG2 cells by 4.6–7.0% (P<0.05; Fig. 1D). The effects of CM on the
abilities of the MCF-7 and HepG2 cells to form colonies were
monitored for 7 days, the marked inhibitory effect of CM on MCF-7
cell colony formation was apparent at 0.2 mg/ml, and the clonogenic
ability of the MCF-7 cells was completely inhibited following
treatment with 2.0 mg/ml CM (Fig.
1E). In addition, 0.4–2.0 mg/ml CM exerted significant
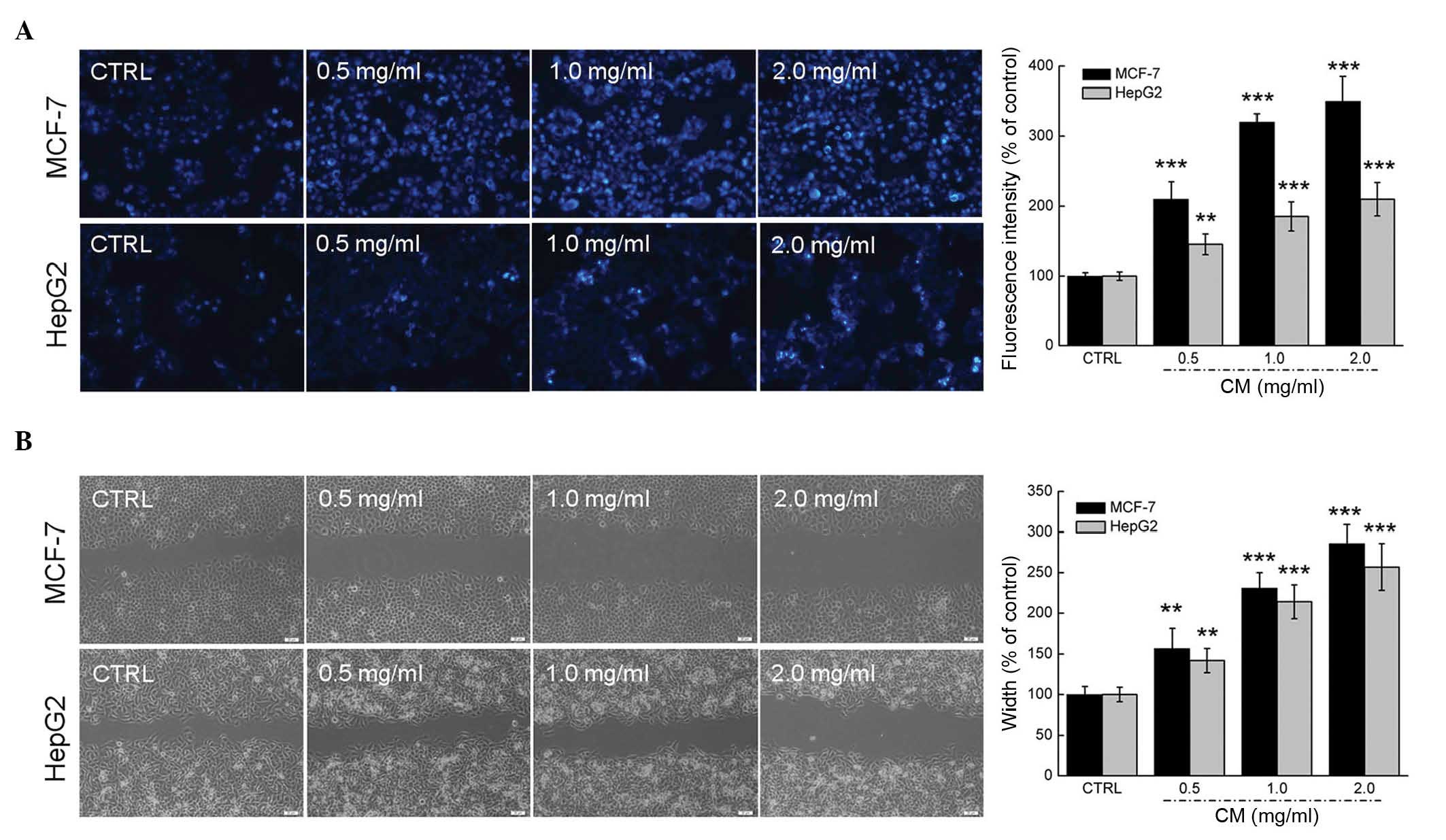
inhibitory effects on HepG2 cell colony formation (Fig. 1F). Hoechst 33342 staining
corroborated that CM at doses between 0.5 and 2.0 mg/ml markedly
induced nuclear apoptosis in the MCF-7 and HepG2 cells, indicated
by the enhanced intensity of blue fluorescence (Fig. 2A). Additionally, a wound healing
assay was performed to observe the inhibitory effect of CM on the
migratory abilities of the MCF-7 and HepG2 cell. Following
incubation for 24 h, the wound areas in the untreated cells were
almost healed. By contrast, the migratory abilities of the MCF-7
and HepG2 cells were significantly inhibited following CM treatment
at specific doses (Fig. 2B).
Collectively, these data confirmed the cytotoxic properties of CM
in MCF-7 and HepG2 cells.
CM causes alterations in apoptosis,
mitochondrial function and the expression levels of pro-apoptotic
proteins
Mitochondrial function is one of the factors
responsible for cell apoptosis. JC-1 staining was applied in the
present study to analyze the alterations of Δψm. Incubation with CM
for 24 h CM caused significant losses of Δψm in the MCF-7 and HepG2
cells, evidenced by enhanced green fluorescence intensity (Fig. 3A).
As one of the factors involved in the Bcl-2 family,
Bax contributes to cell apoptosis and mitochondrial function
(27). Treatment with CM at doses
between 0.5 and 2.0 mg/ml markedly enhanced the expression levels
of Bax in the MCF-7 and HepG2 cells (Fig. 3B). The activation of caspase-3 and
PARP is considered as to be a hallmark of apoptosis, and this was
enhanced following incubation with CM for 24 h in the present study
(Fig. 3B). In addition, increases
in the activation of caspase-8 were observed in the CM-treated
MCF-7 and HepG2 cells (Fig. 3B).
Collectively, CM induced intracellular toxicity, which was
associated with its regulation of mitochondrial function and the
expression of pro-apoptotic proteins.
CM inhibits MCF-7- and HepG2- xenograft
tumor growth in nude mice
In the MCF-7- and HepG2-xenograft tumor nude mice
models, tumor growth inhibition was most marked in the mice treated
with 1 g/kg CM for 2 weeks (every other day), where tumor sizes
reduced by almost 188.2%, compared with the vehicle-treated mice in
the MCF-7 cells (P<0.05; Figs. 4A
and B). Similar to the results obtained from the in
vitro experiment, the administration of CM for 14 days enhanced
the expression levels of cleaved PARP, cleaved caspase-3, cleaved
caspase-8 and Bax in the MCF-7- and HepG2-xenograft tumor tissues,
compared with the vehicle-treated mice (P<0.05; Fig. 4C). Similar results were observed in
the HepG2 cells, with tumor growth inhibited by almost 82.7%,
compared with the vehicle-treated mice (Fig. 5A and B), and administration of CM
for 14 days enhancing the expression levels of cleaved PARP,
cleaved caspase-3, cleaved caspase-8 and Bax (P<0.05; Fig. 5C). These in vivo data
provided further evidence of CM-mediated antitumor activities in
HCC and breast cancer.
 | Figure 5CM has a suppressive effect in the
HepG2-xenograft nude mice model. (A) Examples of tumor growth
between CM- and vehicle-treated male BALB/c athymic nude mice
bearing HepG2 tumors. (B) Growth curves of HepG2-xenograft tumors
in CM- and vehicle-treated nude mice. Tumor sizes were measured
every 2 days. Data are expressed as the mean ± standard deviation
(n=4) and were analyzed using one-way analysis of variance.
*P<0.05, vs. vehicle group. (C) Activation of PARP,
caspase-3 and caspase-8, and the levels of Bax in tumor tissues
were detected using Western blot analysis. The average fold changes
in band intensity, compared with the vehicle group, were marked.
**P<0.01 and ***P<0.001 vs. control.
CM, Cordyceps militaris; CTRL, control; PARP, poly (ADP
ribose) polymerase; Bax, B cell lymphoma-2-associated X protein;
GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
Discussion
The inhibitory activities of CM on MCF-7 and HepG2
cell growth have been reported in previous studies (28–31);
however, their associated mechanisms remain to be elucidated. The
present study investigated the potential antitumor effects of CM on
MCF-7 and HepG2 cells, and examined the possible underlying
mechanisms. Compared with other reported bioactive extracts
(32,33), the crude drug nature of CM suggests
it may have multi-effective components, which may target various
molecules. Due to the systemic targeting, less adverse side effects
are expected. CM has been used for thousands of years as a crude
drug and a traditional medicine in East Asia, further emphasizing
its safety and minimal side effects.
Apoptosis is a physiological suicide mechanism,
which occurs during normal tissue turnover. Mitochondria, triggered
by diverse apoptotic stimuli, commonly exist as the predominant
functional organelle for anticancer drugs (34,35).
Mitochondria control the intrinsic pathway of apoptosis, in which
Δψm induces the activation of caspases and other catabolic enzymes
(36,37). The functional loss of mitochondria
is accompanied with the dissipation of Δψm (38), which was confirmed in the present
study. As reported previously, caspase-8 is located predominantly
in the mitochondria, and active caspase-8 forms a complex with the
BH3-interacting domain death agonist protein on the mitochondria,
which leads to decreased Δψm (39–41).
During this process, caspase-8 undergoes dimerization and cleaves
itself to become fully activated (39). Large quantities of active caspase-8
can lead to the direct cleavage of effector caspase in the cytosol
(42). Bcl-2 family members, which
are located in the outer mitochondrial membrane, are essential
mediators in the regulation of Δψm (27). Bax, a pro-apoptotic protein of the
Bcl-2 family, acts as an important regulatory factor in
mitochondria-mediated apoptosis (27). It has been reported that
reactivation of the Bax gene induces mitochondrial apoptosis in
cholangiocarcinoma cells (43).
Collectively, the data obtained in the present study confirmed that
the mitochondrial apoptotic pathway was involved in CM-meditated
antitumor effects.
In addition, a reduction of Δψm promotes the release
of cytochrome c from the mitochondria (44), which leads to the activation of
caspase-3 and other apoptosis-inducing molecules (45). Caspase-3 is central in the
induction of the apoptotic program (46), and is important for cell death in a
tissue-, cell type- or death stimulus-specific manner (46). As reported, caspase-3 is essential
for the characteristic changes in cell morphology and certain
biochemical events associated with the execution and completion of
apoptosis (46–48). Of note, caspase-3-defective
embryonic stem cells exhibit marked resistance to apoptosis induced
by ultraviolet irradiation and osmotic shock (49). In the present study, the activation
of caspase-3 was observed, and the enhanced expression of PARP was
also found. PARP is considered to be one of the most important
downstream substrates of caspase-3 (50). The activity of PARP is completely
dependent on the number of DNA breaks, and it is totally inactive
in the absence of DNA breaks (51–53).
The over-activation of PARP triggers a cell death cascade, which
results in tissue damage (52). It
has been reported that z-DEVD-fmk can significantly inhibit
apoptosis mediated by the caspase-3/PARP apoptotic signaling
pathway (50). Together,
CM-mediated MCF-7 and HepG2 cell apoptosis is associated with the
caspase-dependent mitochondrial pathway.
In conclusion, the present study confirmed the
anti-HCC and anti-breast cancer effects of CM aqueous extract in
in vitro and in vivo experiments. CM leads to the
over-release of LDH, dissipation of Δψm and abnormal expression of
pro-apoptotic proteins. The caspase-dependent mitochondrial pathway
contributed to CM-induced cytotoxicity in the MCF-7 and HepG2
cells. These findings provide pharmacological evidence that CM
possesses antitumor effects in HCC and breast cancer, offering
potential as a chemotherapeutic agent.
Acknowledgments
This study was supported by the Science and
Technology Key Project of Jilin Province in China (grant no.
20130201006ZY), the Natural Science foundation of P.R. China (grant
no. 81402955) and the 'Twelfth Five-Year' Science and Technology
Planning Project of Jilin Province in China (grant no.
2014B033).
References
|
1
|
Hosseini BA, Pasdaran A, Kazemi T,
Shanehbandi D, Karami H, Orangi M and Baradaran B: Dichloromethane
fractions of Scrophularia oxysepala extract induce apoptosis in
MCF-7 human breast cancer cells. Bosn J Basic Med Sci. 15:26–32.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global Cancer Statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Forner A and Bruix J: Hepatocellular
carcinoma-Authors' reply. Lancet. 380:470–471. 2012. View Article : Google Scholar
|
|
4
|
Johnson PJ: Hepatocellular carcinoma: Is
current therapy really altering outcome? Gut. 51:459–462. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arii S, Yamaoka Y, Futagawa S, Inoue K,
Kobayashi K, Kojiro M, Makuuchi M, Nakamura Y, Okita K and Yamada
R: Results of surgical and nonsurgical treatment for small-sized
hepatocellular carcinomas: A retrospective and nationwide survey in
Japan. The liver cancer study group of Japan. Hepatology.
32:1224–1229. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chang CH, Chen SJ and Liu CY: Adjuvant
treatments of breast cancer increase the risk of depressive
disorders: A population-based study. J Affect Disord. 182:44–49.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chaube SK, Shrivastav TG, Tiwari M, Prasad
S, Tripathi A and Pandey AK: Neem (Azadirachta indica L) leaf
extract deteriorates oocyte quality by inducing ROS-mediated
apoptosis in mammals. Springerplus. 3:4642014. View Article : Google Scholar
|
|
8
|
Ghate NB, Chaudhuri D, Sarkar R, Sajem AL,
Panja S, Rout J and Mandal N: An antioxidant extract of tropical
lichen, Parmotrema reticulatum, induces cell cycle arrest and
apoptosis in breast carcinoma cell line MCF-7. PLoS One.
8:e822932013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Das SK, Masuda M, Sakurai A and Sakakibara
M: Medicinal uses of the mushroom Cordyceps militaris: Current
state and prospects. Fitoterapia. 81:961–968. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ng TB and Wang HX: Pharmacological actions
of Cordyceps, a prized folk medicine. J Pharm Pharmacol.
57:1509–1519. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jin CY, Kim GY and Choi YH: Induction of
apoptosis by aqueous extract of Cordyceps militaris through
activation of caspases and inactivation of Akt in human breast
cancer MDA-MB-231 cells. J Microbiol Biotechnol. 18:1997–2003.
2008.
|
|
12
|
Yoo HS, Shin JW, Cho JH, Son CG, Lee YW,
Park SY and Cho CK: Effects of Cordyceps militaris extract on
angiogenesis and tumor growth. Acta Pharmacol Sin. 25:657–665.
2004.PubMed/NCBI
|
|
13
|
Lee H, Kim YJ, Kim HW, Lee DH, Sung MK and
Park T: Induction of apoptosis by Cordyceps militaris through
activation of caspase-3 in leukemia HL-60 cells. Biol Pharm Bull.
29:670–674. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Reis FS, Barros L, Calhelha RC, Cirić A,
van Griensven LJ, Soković M and Ferreira IC: The methanolic extract
of Cordyceps militaris (L.) Link fruiting body shows antioxidant,
antibacterial, antifungal and antihuman tumor cell lines
properties. Food Chem Toxicol. 62:91–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jing Y, Cui X, Chen Z, Huang L, Song L,
Liu T, Lv W and Yu R: Elucidation and biological activities of a
new polysaccharide from cultured Cordyceps militaris. Carbohydr
Polym. 102:288–296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakagawa S, Shiraishi T, Kihara S and
Tabuchi K: Detection of DNA strand breaks associated with apoptosis
in human brain tumors. Virchows Arch. 427:175–179. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pintus F, Floris G and Rufini A: Nutrient
availability links mitochondria, apoptosis and obesity. Aging
(Albany NY). 4:734–741. 2012. View Article : Google Scholar
|
|
18
|
Chen R, Liu S, Piao F, Wang Z, Qi Y, Li S,
Zhang D and Shen J: 2,5-Hexanedione induced apoptosis in
mesenchymal stem cells from rat bone marrow via
mitochondria-dependent caspase-3 pathway. Ind Health. 53:222–235.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Wu Y, Luo K, Liu Y, Zhou M, Yan S,
Shi H and Cai Y: The protective effects of selenium on
cadmium-induced oxidative stress and apoptosis via mitochondria
pathway in mice kidney. Food Chem Toxicol. 58:61–67. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu Q, Wu D, Chen W, Yan Z and Shi Y:
Proteolytic processing of the caspase-9 zymogen is required for
apoptosome-mediated activation of caspase-9. J Biol Chem.
288:15142–15147. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boatright KM, Renatus M, Scott FL,
Sperandio S, Shin H, Pedersen IM, Ricci JE, Edris WA, Sutherlin DP,
Green DR and Salvesen GS: A unified model for apical caspase
activation. Mol Cell. 11:529–541. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kroemer G, Dallaporta B and Resche-Rigon
M: The mitochondrial death/life regulator in apoptosis and
necrosis. Annu Rev Physiol. 60:619–642. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baharara J and Amini E: The potential of
brittle star extracted polysaccharide in promoting apoptosis via
intrinsic signaling pathway. Avicenna J Med Biotechnol. 7:151–158.
2015.PubMed/NCBI
|
|
24
|
Kato M, Yamazaki T, Kato H, Eyama S, Goto
M, Yoshioka M and Takatsu A: Development of high-purity certified
reference materials for 17 proteinogenic amino acids by traceable
titration methods. Anal Sci. 31:805–814. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li SP, Yang FQ and Tsim KW: Quality
control of Cordyceps sinensis, a valued traditional Chinese
medicine. J Pharm Biomed Anal. 41:1571–1584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: Application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chan SL and Yu VC: Proteins of the bcl-2
family in apoptosis signalling: From mechanistic insights to
therapeutic opportunities. Clin Exp Pharmacol Physiol. 31:119–128.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Reis FS, Barros L, Calhelha RC, Cirić A,
van Griensven LJ, Soković M and Ferreira IC: The methanolic extract
of Cordyceps militaris (L.) Link fruiting body shows antioxidant,
antibacterial, antifungal and antihuman tumor cell lines
properties. Food Chem Toxicol. 62:91–98. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rao YK, Fang SH, Wu WS and Tzeng YM:
Constituents isolated from Cordyceps militaris suppress enhanced
inflammatory mediator's production and human cancer cell
proliferation. J Ethnopharmacol. 131:363–367. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jing Y, Cui X, Chen Z, Huang L, Song L,
Liu T, Lv W and Yu R: Elucidation and biological activities of a
new polysaccharide from cultured Cordyceps militaris. Carbohydr
Polym. 102:288–296. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wong JH, Wang H and Ng TB: A
haemagglutinin from the medicinal fungus Cordyceps militaris.
Biosci Rep. 29:321–327. 2009. View Article : Google Scholar
|
|
32
|
Nourazarian SM, Nourazarian A, Majidinia M
and Roshaniasl E: Effect of root extracts of medicinal herb
Glycyrrhiza glabra on HSP90 gene rxpression and apoptosis in the
HT-29 colon cancer cell line. Asian Pac J Cancer Prev.
16:8563–8566. 2015. View Article : Google Scholar
|
|
33
|
Hu B, An HM, Wang SS, Chen JJ and Xu L:
Preventive and therapeutic effects of Chinese herbal compounds
against hepatocellular carcinoma. Molecules. 21:1422016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fulda S and Debatin KM: Extrinsic versus
intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene.
25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Y, Xie RF, Xiao QG, Li R, Shen XL
and Zhu XG: Hedyotis diffusa Willd extract inhibits the growth of
human glioblastoma cells by inducing mitochondrial apoptosis via
AKT/ERK pathways. J Ethnopharmaco. 158:404–411. 2014. View Article : Google Scholar
|
|
36
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Galluzzi L, Vitale I, Kepp O, Séror C,
Hangen E, Perfettini JL, Modjtahedi N and Kroemer G: Methods to
dissect mitochondrial membrane permeabilization in the course of
apoptosis. Methods Enzymol. 442:355–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hisatomi T, Ishibashi T, Miller JW and
Kroemer G: Pharmacological inhibition of mitochondrial membrane
permeabilization for neuroprotection. Exp Neurol. 218:347–352.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schug ZT, Gonzalvez F, Houtkooper RH, Vaz
FM and Gottlieb E: BID is cleaved by caspase-8 within a native
complex on the mitochondrial membrane. Cell Death Differ.
18:538–548. 2011. View Article : Google Scholar :
|
|
40
|
Hyun HB, Lee WS, Go SI, Nagappan A, Park
C, Han MH, Hong SH, Kim G, Kim GY, Cheong J, et al: The flavonoid
morin from Moraceae induces apoptosis by modulation of Bcl-2 family
members and Fas receptor in HCT 116 cells. Int J Oncol.
46:2670–2678. 2015.PubMed/NCBI
|
|
41
|
Lee JW, Park C, Han MH, Hong SH, Lee TK,
Lee SH, Kim GY and Choi YH: Induction of human leukemia U937 cell
apoptosis by an ethanol extract of Dendropanax morbifera Lev.
Through the caspase-dependent pathway. Oncol Rep. 30:1231–1238.
2013.PubMed/NCBI
|
|
42
|
Lee KH, Feig C, Tchikov V, Schickel R,
Hallas C, Schütze S, Peter ME and Chan AC: The role of receptor
internalization in CD95 signaling. EMBO J. 25:1009–1023. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu XF, Jiang H, Zhang CS, Yu SP, Wang ZQ
and Su HL: Targeted drug regulation on methylation of p53-BAX
mitochondrial apoptosis pathway affects the growth of
cholangiocarcinoma cells. J Int Med Res. 40:67–75. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kroemer G, Galluzzi L and Brenner C:
Mitochondrial membrane permeabilization in cell death. Physiol Rev.
87:99–163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bao Q and Shi Y: Apoptosome: A platform
for the activation of initiator caspases. Cell Death Differ.
14:56–65. 2007. View Article : Google Scholar
|
|
46
|
Porter AG and Jänicke RU: Emerging roles
of caspase-3 in apoptosis. Cell Death Differ. 6:99–104. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Visagie M, Theron A, Mqoco T, Vieira W,
Prudent R, Martinez A, Lafanechère L and Joubert A: Sulphamoylated
2-methoxyestradiol analogues induce apoptosis in adenocarcinoma
cell lines. PLoS One. 8:e719352013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tor YS, Yazan LS, Foo JB, Armania N, Cheah
YK, Abdullah R, Imam MU, Ismail N and Ismail M: Induction of
apoptosis through oxidative stress-related pathways in MCF-7, human
breast cancer cells, by ethyl acetate extract of Dillenia
suffruticosa. BMC Complement Altern Med. 14:552014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Woo M, Hakem R, Soengas MS, Duncan GS,
Shahinian A, Kägi D, Hakem A, McCurrach M, Khoo W, Kaufman SA, et
al: Essential contribution of caspase 3/CPP32 to apoptosis and its
associated nuclear changes. Genes Dev. 12:806–819. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Liu J, Wu Y, Wang B, Yuan X and Fang B:
High levels of glucose induced the caspase-3/PARP signaling
pathway, leading to apoptosis in human periodontal ligament
fibroblasts. Cell Biochem Biophys. 66:229–237. 2013. View Article : Google Scholar
|
|
51
|
Benjamin RC and Gill DM: Poly (ADP-ribose)
synthesis in vitro programmed by damaged DNA. A comparison of DNA
molecules containing different types of strand breaks. J Biol Chem.
255:10502–10508. 1980.PubMed/NCBI
|
|
52
|
Wang H, Shimoji M, Yu SW, Dawson TM and
Dawson VL: Apoptosis inducing factor and PARP-mediated injury in
the MPTP mouse model of Parkinson's disease. Ann N Y Acad Sci.
991:132–139. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shi Y, Zhou F, Jiang F, Lu H, Wang J and
Cheng C: PARP inhibitor reduces proliferation and increases
apoptosis in breast cancer cells. Chin J Cancer Res. 26:142–147.
2014.PubMed/NCBI
|