MicroRNAs (miRNAs) are an extensive family of small
(18–24 nucleotides), single-stranded non-coding RNAs, which
regulate gene expression in mammalian cells through binding to the
seed sequences of the 3′-untranslated region (UTR) of target mRNA
sequences and mediate the degradation of mRNA in the RNA-induced
silencing complex (1,2). miRNAs repress translation by
decreasing stability through targeting specific mRNA targets. Each
miRNA regulates numerous mRNAs, and miRNAs are involved in various
cellular processes, including proliferation, differentiation, cell
cycle, invasion and apoptosis (3–6). The
alterations in their expression levels may lead to human diseases,
including cardiovascular disease and cancer (7–10).
The formation of bone by osteoblast cells and their primary
functional activities involve a series of multiple signals,
including bone morphogenic protein (BMP), Wnt ligands, Notch
ligands, hormones and growth factors, including transforming growth
factor (TGF) and tumor necrosis factor (TNF) and cytokines. In
addition to these factors, tissue-specific transcription factors
and co-factors mediate the expression of genes for the biosynthesis
and mineralization of bone matrix, and the remodeling and formation
of bone (11–15). Mesenchymal stem cells (MSCs) are
induced to differentiate into preosteoblasts, which are regulated
by the signaling cascades initiated by the various signals,
including cytokine stimuli. Pre-osteoblasts differentiate into
osteoblasts, which is controlled by gene expression affected by
chromatin modifiers, transcription factors or miRNA alterations
(16). Osteoblasts produce bone
matrix, which mineralizes into bone tissue, and osteoblasts are
engulfed in the matrix during bone formation. The entire process is
complex and well organized by the signaling network.
Experimental evidence shows that miRNAs are critical
for stem cell activities, particularly in the stemness maintenance
or direct differentiation into lineage cells (7,8). To
date, several studies have examined the functions of miRNAs in MSCs
(17,18). Certain miRNAs have been shown to
regulate the process of differentiation of MSCs into different cell
lineages using high-through gene expression microprofiling assays.
This has led to the specific targets of these miRNAs being
determined, and the roles of miRNAs are gradually being elucidated
(Fig. 1).
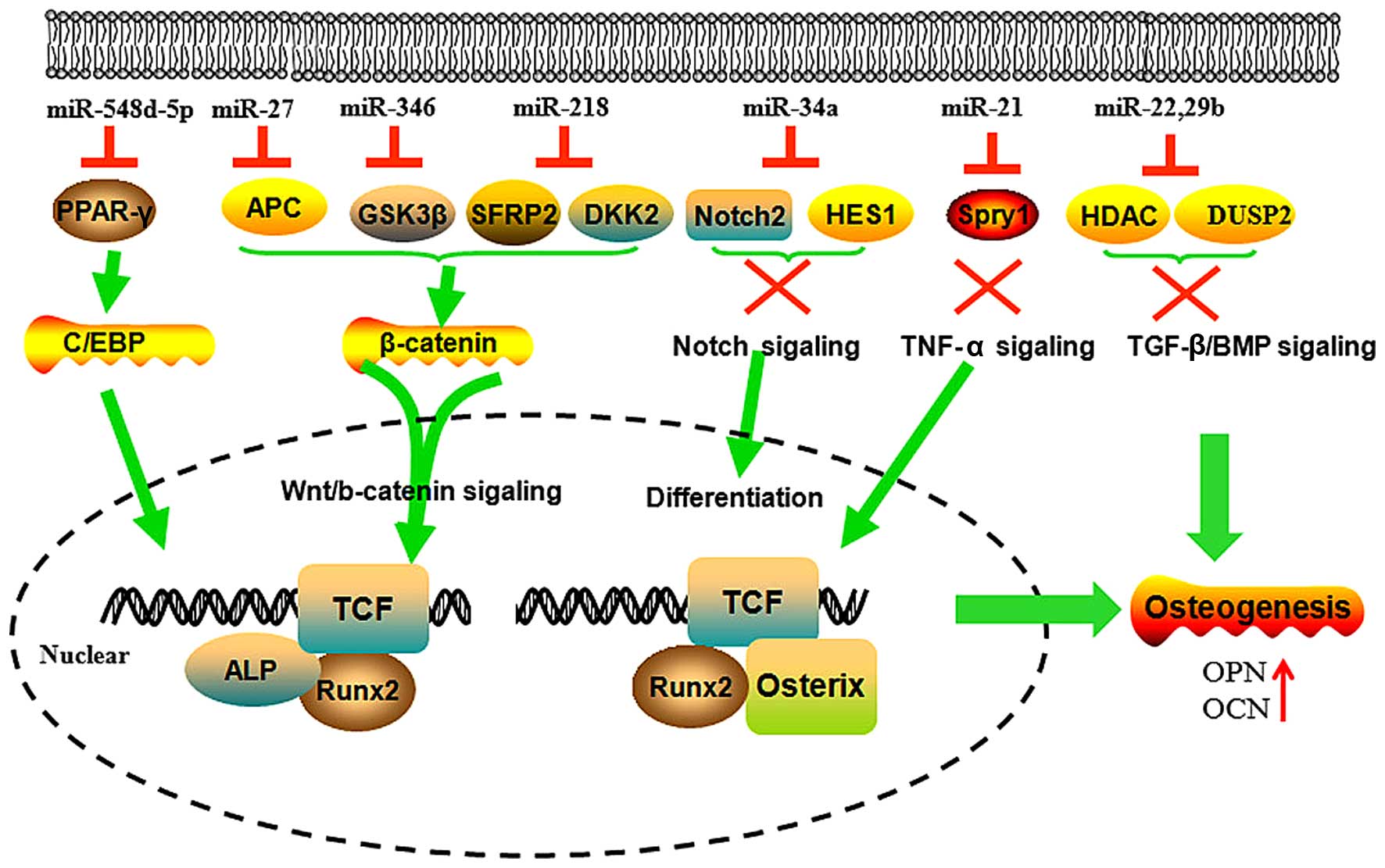
Wnt/β-catenin signaling has been well defined in the
osteogenic differentiation of MSCs (24–27).
miR-346 promotes the osteogenic differentiation of human bone
marrow MSCs (hBMSCs) by targeting glycogen synthase kinase-3β
(GSK-3β) through binding to the 3′-UTR of its mRNA. The decreased
GSK-3β results in an increase of β-catenin, which is translocated
into the nucleus and activates the downstream genes of the
Wnt/β-catenin pathway. β-catenin knockdown almost completely
inhibits the positive effect of miR-346 on osteogenic
differentiation. Therefore, miR-346 positively regulates the
osteogenic differentiation of hBMSCs through the Wnt/β-catenin
pathway (28–31). The expression of miR-27 is
increased during hFOB1.19 cell (human SV40 transfected osteoblast)
differentiation. The ectopic expression of miR-27 promotes the
differentiation of hFOB1.19 cells by directly targeting and
inhibiting the gene expression of adenomatous polyposis coli (APC).
This inhibition of the expression of APC leads to the accumulation
of β-catenin, which is a key protein activating Wnt signaling. This
suggests that miR-27 is an important promoter of osteogenic
differentiation (32). miR-218
positively regulates the osteogenesis of hADSC, directly targeting
secreted frizzled-related protein 2 and dickkopf WNT signaling
pathway inhibitor 2, thus enhancing Wnt/β-catenin signaling
progression. Mimics of the Wnt/β-catenin signal increase the
expression of miR-218, which forms a positive feedback loop to
promote osteogenesis (33).
The upregulation of miR-10a results in increased
osteogenic differentiation. miR-10a mimics significantly repress
luciferase activity by the direct binding to the 3′-UTR of
Kruppel-like factor 4 (KLF4). The inhibition of KLF4 in hBMSCs
increases cell differentiation. Therefore, miR-10a enhances the
differentiation capability of hBMSCs through the repression of KLF4
(40).
MiR-21 has been confirmed to promote the osteogenic
differentiation of mouse bone marrow cells by targeting Sprouty
homolog 1 (Spry1), negatively regulating the osteogenic
differentiation of MSCs. The expression of miR-21 partially rescues
TNF-α-impaired osteogenesis of MSCs. Inhibition of the TNF-α
signaling pathway evidently improves bone formation and
down-regulates Spry1 expression, which suggests that miR-21
contributes towards bone formation through targeting Spry1 in MSCs
(41).
miR-96 and miR-199a were both up-regulated during
osteogenic induction of human bone marrow derived MSCs. They may
function through transcription factor SRY-box 9, and fatty acid
binding protein 4, however, the detailed molecular mechanism
remains poorly understood (42).
The expression of miR-22 is increased during the
process of osteogenic differentiation of hADSCs. miR-22 inhibits
the expression of histone deacetylase 6 (HDAC6) by binding to the
similar sequence of the 3′-UTR (43). HDAC6 deficiency results in a minor
increase in trabecular bone density. Thus, miR-22 increases
osteogenesis through targeting HDAC6.
miR-29b is understood to promote osteogenesis by
supressing the negative regulators, including histone deacetylase
4, TGF-β3, activin A receptor (Type IIA), catenin beta interacting
protein 1 and dual specificity phosphatase 2, during the induction
of osteogenic differentiation. These anti-osteogenic factors
negatively modulate extracellular matrix excretion by
differentiated osteoblasts and bone formation (44,45).
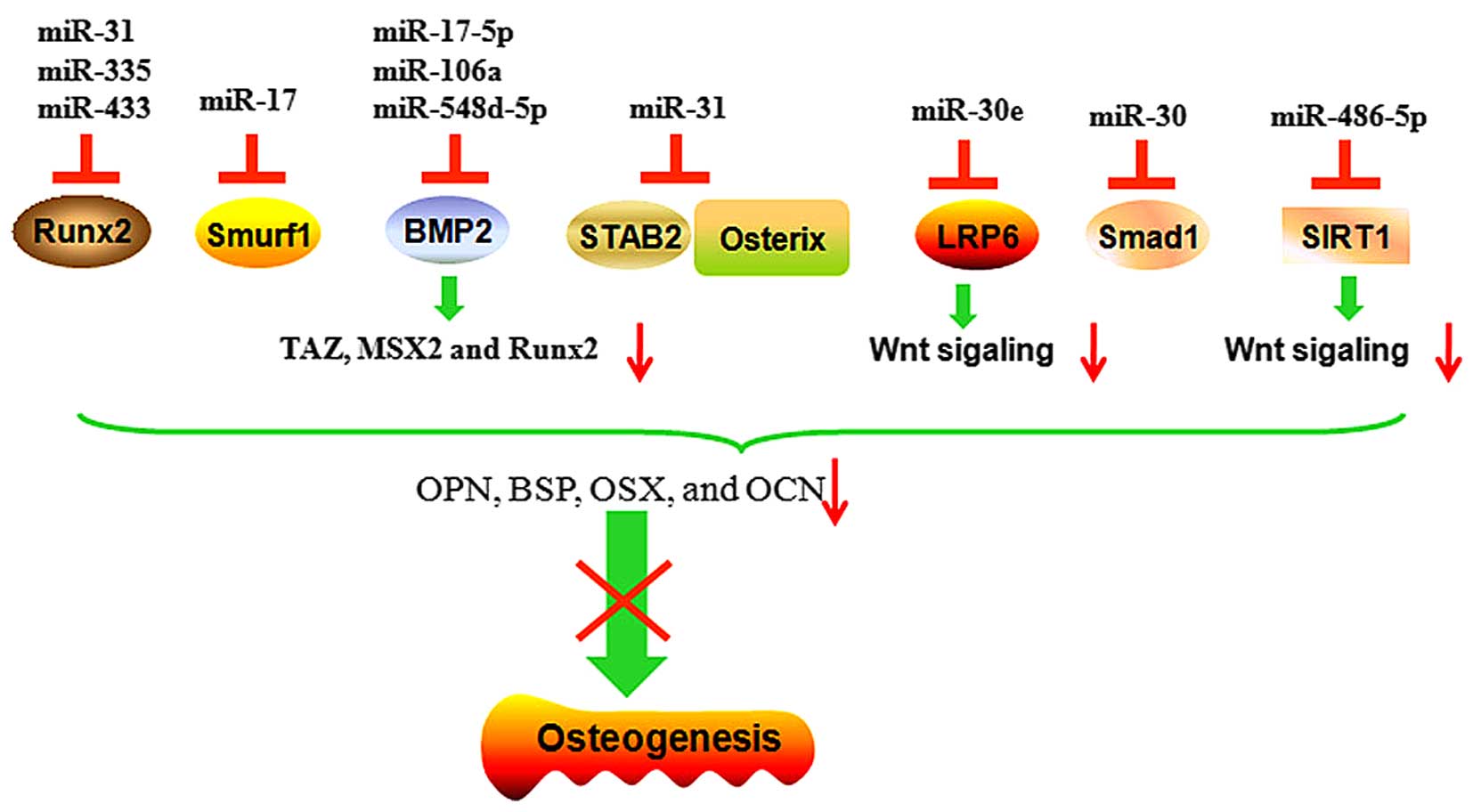
Several miRNAs directly target transcription
factors, including osteopotin (OPN), OCN and RUNX2, or signaling
molecules, which promote the osteogenesis of MSCs and inhibit
osteogenic differentiation (Fig. 3
and Table I). miR-140-5p inhibits
the osteogenic differentiation in hMSCs by directly blocking BMP2,
and consequently blocking BMP signaling components and critical
regulators (46).
The overexpression of miR-31 represses the
osteogenesis of hMSCs by directly targeting special AT-rich
sequence-binding protein 2 (SATB2), the knockdown of SATB2 by
specific siRNA against SATB2 inhibits osteogenic differentiation
(47). Baglio et al found
an inverse trend in miRNA-target expression during osteogenic
differentiation between the levels of miR-31 and OSX. The
inhibition of miR-31 leads to an increase in the endogenous
expression of OSX (48). Deng
et al showed that the expression of miR-31 decreased
progressively in BMSC cultures during differentiation. The
upregulation of miR-31 significantly reduces the expression levels
of osteogenic transcription factors, OPN, bone sialoprotein, OSX
and OCN. The inhibition of miR-31 markedly decreases the activity
of ALP and inhibits osteogenesis. These results suggest that an
miR-31/SATB2 axis is involved in the osteogenic differentiation of
BMSCs (49).
miR-30 family members are also important regulators
during the biomineralization process. The overexpression of miR-30e
stimulates adipocyte formation and inhibits osteoblast
differentiation from marrow stromal cells. Low-density lipoprotein
receptor-related protein 6 (LRP6) is one of the critical
co-receptors for Wnts. Blocking LRP6 in 3T3-L1 cells downregulates
β-catenin/T-cell factor transcriptional activity and enhances
osteogenic differentiation. miR-30 has been reported to target the
LRP6 directly and inhibit the expression of LRP6 expression. Thus,
miR-30e also controls osteogenesis in periodontal ligament stem
cell (PDLSCs) by targeting LRP6 and affecting canonical
Wnt/β-catenin signaling (50).
There is also evidence that miR-30 family members negatively
regulate BMP-2-induced osteoblast differentiation by targeting
Small mothers against decapentaplegic (Smad)1 and RUNX2. As Smad1
and RUNX2 are key positive transcription factors in osteogenic
differentiation, miR-30 family members negatively regulate the
osteogenic differentiation through Smad1 and RUNX2 (51).
During osteoblastic differentiation, the
overexpression of Estrogen-related receptor γ (ERRγ) or miR-433
inhibits the expression levels of osteogenic marker genes including
RUNX2 and ALP. miR-433 directly targets three binding sites on the
3′-UTR of RUNX2 mRNA, and decreases the levels of the RUNX2
transcript in C3H10T1/2 cells. Anti-miR-433 recovers the
ERRγ-suppressed expression of RUNX2 and activity of ALP. ERRγ is
able to upregulate the levels of miR-433 expression and further
enhance the inhibitory role in osteogenic differentiation. This
evidence demonstrates that miR-433 suppresses BMP2-induced
osteogenic differentiation by targeting RUNX2 C3H10T1/2 cells
(54).
The overexpression of miR-486-5p inhibits osteogenic
differentiation of hADSCs. miR-486-5p regulates the expression of
silent information regulator 1 (SIRT1), a major regulator of
longevity and metabolic disorders. SIRT1 also has an important role
in the osteogenic process by targeting FOXO3A and then upregulating
the levels of RUNX2 promoter activity. miR-486-5p inhibits the
expression of SIRT1 through binding to the 3′-UTR region of SIRT1
mRNA (55). The overexpression of
miR-335 in hMSCs derived from bone marrow, adipose tissue and
articular cartilage inhibits their osteogenic and adipogenic
potential. The expression of miR-335 in hMSCs is upregulated by the
activated canonical Wnt signaling pathway. It has also been
confirmed that RUNX2 is a direct target of miR-335. These results
suggest that the downregulation of miR-335 is critical for the
acquisition of MSC phenotype differentiation into osteoblasts
(56). The overexpression of
miR-135b downregulates the osteogenic differentiation of
unrestricted somatic stem cells through targeting the key
osteogenic factors, integrin-binding sialoprotein and OSX,
inhibiting the process of osteogenesis (57).
There is substantial evidence and experimental data
confirming that miRNAs have multi-dimensional roles in the
induction of MSCs into osteoblasts. miRNAs function at all stages
of osteoblast differentiation by inhibiting the negative regulators
of signaling pathways operating in these cells (16). They also have direct and indirect
effects on phenotype development through the promotion or
inhibition of positive or negative transcription factors in
signaling pathways, which are involved in complex regulatory
networks. This indicates an important technique, by which the
network of osteogenic differentiation can be regulated through
miRNAs. Nicotine can alter the expression of miRNA and reduce human
adult stem cell regenerative potential, demonstrating that miRNAs
function in the network (58,59).
This study was supported by funds from The Natural
Science Foundation of China (grant nos. 81572577 and 81428018), the
Program for New Century Excellent Talents in University (grant no.
NCET-12-0544), the Open-End Fund for the Valuable and Precision
Instruments of Central South University, Hunan Provincial Natural
Science Foundation of China (grant no. 14JJ1006) and the Project of
Innovation-driven Plan of Central South University (grant no.
2016CX023).
|
1
|
Brennecke J, Hipfner DR, Stark A, Russell
RB and Cohen SM: Bantam encodes a developmentally regulated
microRNA that controls cell proliferation and regulates the
proapoptotic gene hid in drosophila. Cell. 113:25–36. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP and Chen CZ: Micromanagers of
gene expression: The potentially widespread influence of metazoan
micrornas. Nat Rev Genet. 5:396–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hyun J, Wang S, Kim J, Rao KM, Park SY,
Chung I, Ha CS, Kim SW, Yun YH and Jung Y: MicroRNA-378 limits
activation of hepatic stellate cells and liver fibrosis by
suppressing Gli3 expression. Nat Commun. 7:109932016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cui Y, Han J, Xiao Z, Chen T, Wang B, Chen
B, Liu S, Han S, Fang Y, Wei J, et al: The miR-20-Rest-Wnt
signaling axis regulates neural progenitor cell differentiation.
Sci Rep. 6:233002016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gao Y, Chen L, Song H, Chen Y, Wang R and
Feng B: A double-negative feedback loop between E2F3b and miR-200b
regulates docetaxel chemosensitivity of human lung adenocarcinoma
cells. Oncotarget. Mar 25–2016.Epub ahead of print.
|
|
6
|
Ma L, Ma S, Zhao G, Yang L, Zhang P, Yi Q
and Cheng S: miR-708/LSD1 axis regulates the proliferation and
invasion of breast cancer cells. Cancer Med. 5:684–692. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim VN: Microrna biogenesis: Coordinated
cropping and dicing. Nat Rev Mol Cell Biol. 6:376–385. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: Microrna expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thum T and Mayr M: Review focus on the
role of microRNA in cardiovascular biology and disease. Cardiovasc
Res. 93:543–544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Van Rooij E and Olson EN: Microrna
therapeutics for cardiovascular disease: Opportunities and
obstacles. Nat Rev Drug Discov. 11:860–872. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Oryan A, Alidadi S, Moshiri A and
Bigham-Sadegh A: Bone morphogenetic proteins: A powerful
osteoinductive compound with non-negligible side effects and
limitations. Biofactors. 40:459–481. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yuan Z, Li Q, Luo S, Liu Z, Luo D, Zhang
B, Zhang D, Rao P and Xiao J: PPARγ and Wnt signaling in adipogenic
and osteogenic differentiation of mesenchymal stem cells. Curr Stem
Cell Res Ther. 11:216–225. 2016. View Article : Google Scholar
|
|
13
|
Ongaro A, Pellati A, Bagheri L, Rizzo P,
Caliceti C, Massari L and De Mattei M: Characterization of notch
signaling during osteogenic differentiation in human osteosarcoma
cell line MG63. J Cell Physiol. Mar 4–2016.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen YJ, Yao CC, Huang CH, Chang HH and
Young TH: Hexosamine-induced TGF-β signaling and osteogenic
differentiation of dental pulp stem cells are dependent on
N-acetylglucosaminyltransferase V. Biomed Res Int.
2015:9243972015.
|
|
15
|
Marupanthorn K, Tantrawatpan C,
Tantikanlayaporn D, Kheolamai P and Manochantr S: The effects of
TNF-α on osteogenic differentiation of umbilical cord derived
mesenchymal stem cells. J Med Assoc Thai. 98(Suppl 3): S34–S40.
2015.
|
|
16
|
Lian JB, Stein GS, van Wijnen AJ, Stein
JL, Hassan MQ, Gaur T and Zhang Y: microRNA control of bone
formation and homeostasis. Nat Rev Endocrinol. 8:212–227. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Van Wijnen AJ, van de Peppel J, van
Leeuwen JP, Lian JB, Stein GS, Westendorf JJ, Oursler MJ, Im HJ,
Taipaleenmäki H, Hesse E, et al: Microrna functions in osteogenesis
and dysfunctions in osteoporosis. Curr Osteoporos Rep. 11:72–82.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dong S, Yang B, Guo H and Kang F:
microRNAs regulate osteogenesis and chondrogenesis. Biochem Biophys
Res Commun. 418:587–591. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lamplot JD, Qin J, Nan G, Wang J, Liu X,
Yin L, Tomal J, Li R, Shui W, Zhang H, et al: Bmp9 signaling in
stem cell differentiation and osteogenesis. Am J Stem Cells.
2:1–21. 2013.PubMed/NCBI
|
|
20
|
Peng Y, Kang Q, Cheng H, Li X, Sun MH,
Jiang W, Luu HH, Park JY, Haydon RC and He TC: Transcriptional
characterization of bone morphogenetic proteins (BMPs)-mediated
osteogenic signaling. J Cell Biochem. 90:1149–1165. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hupkes M, Sotoca AM, Hendriks JM, van
Zoelen EJ and Dechering KJ: Microrna mir-378 promotes BMP2-induced
osteogenic differentiation of mesenchymal progenitor cells. BMC Mol
Biol. 15:12014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wagner ER, Zhu G, Zhang BQ, Luo Q, Shi Q,
Huang E, Gao Y, Gao JL, Kim SH, Rastegar F, et al: The therapeutic
potential of the Wnt signaling pathway in bone disorders. Curr Mol
Pharmacol. 4:14–25. 2011. View Article : Google Scholar
|
|
23
|
Kim JH, Liu X, Wang J, Chen X, Zhang H,
Kim SH, Cui J, Li R, Zhang W, Kong Y, et al: Wnt signaling in bone
on and its therapeutic potential for bone diseases. Ther Adv
Musculoskelet Dis. 5:13–31. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Marcellini S, Henriquez JP and Bertin A:
Control of osteogenesis by the canonical Wnt and Bmp pathways in
vivo: Cooperation and antagonism between the canonical Wnt and BMP
pathways as cells differentiate from osteochondroprogenitors to
osteoblasts and osteocytes. Bioessays. 34:953–962. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Krause U and Gregory CA: Potential of
modulating Wnt signaling pathway toward the development of bone
anabolic agent. Curr Mol Pharmacol. 5:164–173. 2012. View Article : Google Scholar
|
|
26
|
Wang Q, Cai J, Cai XH and Chen L: Mir-346
regulates osteogenic differentiation of human bone marrow-derived
mesenchymal stem cells by targeting the Wnt/β-catenin pathway. PLoS
One. 8:e722662013. View Article : Google Scholar
|
|
27
|
Westendorf JJ, Kahler RA and Schroeder TM:
Wnt signaling in osteoblasts and bone diseases. Gene. 341:19–39.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gaur T, Lengner CJ, Hovhannisyan H, Bhat
RA, Bodine PV, Komm BS, Javed A, van Wijnen AJ, Stein JL, Stein GS
and Lian JB: Canonical WNT signaling promotes osteogenesis by
directly stimulating Runx2 gene expression. J Biol Chem.
280:33132–33140. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang T and Xu Z: Mir-27 promotes
osteoblast differentiation by modulating Wnt signaling.
BiochemBiophys Res Commun. 402:186–189. 2010. View Article : Google Scholar
|
|
31
|
Zhang WB, Zhong WJ and Wang L: A
signal-amplification circuit between miR-218 and Wnt/β-catenin
signal promotes human adipose tissue-derived stem cells osteogenic
differentiation. Bone. 58:59–66. 2014. View Article : Google Scholar
|
|
32
|
Li Y, Li SQ, Gao YM, Li J and Zhang B:
Crucial role of Notch signaling in osteogenic differentiation of
periodontal ligament stem cells in osteoporotic rats. Cell Biol
Int. 38:729–736. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shimizu T, Tanaka T, Iso T, Doi H, Sato H,
Kawai-Kowase K, Arai M and Kurabayashi M: Notch signaling induces
osteogenic differentiation and mineralization of vascular smooth
muscle cells role of Msx2 gene induction via Notch-RBP-Jk
signaling. Arterioscler Thromb Vasc Biol. 29:1104–1111. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shimizu T, Tanaka T, Iso T, Matsui H,
Ooyama Y, Kawai-Kowase K, Arai M and Kurabayashi M: Notch signaling
pathway enhances bone morphogenetic protein 2 (BMP2) responsiveness
of Msx2 gene to induce osteogenic differentiation and
mineralization of vascular smooth muscle cells. J Biol Chem.
286:19138–19148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shindo K, Kawashima N, Sakamoto K,
Yamaguchi A, Umezawa A, Takagi M, Katsube K and Suda H: Osteogenic
differentiation of the mesenchymal progenitor cells, Kusa is
suppressed by Notch signaling. Exp Cell Res. 290:370–380. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ugarte F, Ryser M, Thieme S, Fierro FA,
Navratiel K, Bornhäuser M and Brenner S: Notch signaling enhances
osteogenic differentiation while inhibiting adipogenesis in primary
human bone marrow stromal cells. Exp Hematol. 37:867–875. 2009.
View Article : Google Scholar
|
|
37
|
Sun F, Wan M, Xu X, Gao B, Zhou Y, Sun J,
Cheng L, Klein OD, Zhou X and Zheng L: Crosstalk between miR-34a
and notch signaling promotes differentiation in apical papilla stem
cells (SCAPs). J Dent Res. 93:589–595. 2014.Epub ahead of print.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li J, Dong J, Zhang ZH, Zhang DC, You XY,
Zhong Y, Chen MS and Liu SM: Mir-10a restores human mesenchymal
stem cell differentiation by repressing KLF4. J Cell Physiol.
228:2324–2336. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gamez B, Rodriguez-Carballo E, Bartrons R,
Rosa JL and Ventura F: microRNA-322 (miR-322) and its target
protein Tob2 modulate osterix (osx) mrna stability. J Biol Chem.
288:14264–14275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang N, Wang G, Hu C, Shi Y, Liao L, Shi
S, Cai Y, Cheng S, Wang X, Liu Y, et al: Tumor necrosis factor α
suppresses the mesenchymal stem cell osteogenesis promoter miR-21
in estrogen deficiency-induced osteoporosis. J Bone Miner Res.
28:559–573. 2013. View Article : Google Scholar
|
|
41
|
Laine SK, Alm JJ, Virtanen SP, Aro HT and
Laitala-Leinonen TK: microRNAs miR-96, miR-124 and miR-199a
regulate gene expression in human bone marrow-derived mesenchymal
stem cells. J Cell Biochem. 113:2687–2695. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang S, Wang S, Bian C, Yang Z, Zhou H,
Zeng Y, Li H, Han Q and Zhao RC: Upregulation of miR-22 promotes
osteogenic differentiation and inhibits adipogenic differentiation
of human adipose tissue-derived mesenchymal stem cells by
repressing HDAC6 protein expression. Stem Cells Dev. 21:2531–2540.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li Z, Hassan MQ, Jafferji M, Aqeilan RI,
Garzon R, Croce CM, van Wijnen AJ, Stein JL, Stein GS and Lian JB:
Biological functions of miR-29b contribute to positive regulation
of osteoblast differentiation. J Biol Chem. 284:15676–15684. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Crane JL and Cao X: Bone marrow
mesenchymal stem cells and TGF-β signaling in bone remodeling. J
Clin Invest. 124:466–472. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kim YJ, Bae SW, Yu SS, Bae YC and Jung JS:
Mir-196a regulates proliferation and osteogenic differentiation in
mesenchymal stem cells derived from human adipose tissue. J Bone
Miner Res. 24:816–825. 2009. View Article : Google Scholar
|
|
46
|
Hwang S, Park SK, Lee HY, Kim SW, Lee JS,
Choi EK, You D, Kim CS and Suh N: Mir-140-5p suppresses
BMP2-mediated osteogenesis in undifferentiated human mesenchymal
stem cells. FEBS Lett. 588:2957–2963. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xie Q, Wang Z, Bi X, Zhou H, Wang Y, Gu P
and Fan X: Effects of miR-31 on the osteogenesis of human
mesenchymal stem cells. Biochem Biophys Res Commun. 446:98–104.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Baglio SR, Devescovi V, Granchi D and
Baldini N: microRNA expression profiling of human bone marrow
mesenchymal stem cells during osteogenic differentiation reveals
osterix regulation by miR-31. Gene. 527:321–331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Deng Y, Wu S, Zhou H, Bi X, Wang Y, Hu Y,
Gu P and Fan X: Effects of a miR-31, Runx2 and Satb2 regulatory
loop on the osteogenic differentiation of bone mesenchymal stem
cells. Stem Cells Dev. 22:2278–2286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang J, Guan X, Guo F, Zhou J, Chang A,
Sun B, Cai Y, Ma Z, Dai C, Li X and Wang B: Mir-30e reciprocally
regulates the differentiation of adipocytes and osteoblasts by
directly targeting low-density lipoprotein receptor-related protein
6. Cell Death Dis. 4:e8452013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wu T, Zhou H, Hong Y, Li J, Jiang X and
Huang H: miR-30 family members negatively regulate osteoblast
differentiation. J Biol Chem. 287:7503–7511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu Y, Liu W, Hu C, Xue Z, Wang G, Ding B,
Luo H, Tang L, Kong X, Chen X, et al: miR-17 modulates osteogenic
differentiation through a coherent feed-forward loop in mesenchymal
stem cells isolated from periodontal ligaments of patients with
periodontitis. Stem Cells. 29:1804–1816. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li H, Li T, Wang S, Wei J, Fan J, Li J,
Han Q, Liao L, Shao C and Zhao RC: miR-17-5p and miR-106a are
involved in the balance between osteogenic and adipogenic
differentiation of adipose-derived mesenchymal stem cells. Stem
Cell Res. 10:313–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kim EJ, Kang IH, Lee JW, Jang WG and Koh
JT: miR-433 mediates ERRγ-suppressed osteoblast differentiation via
direct targeting to Runx2 mRNA in C3H10T1/2 cells. Life Sci.
92:562–568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kim YJ, Hwang SH, Lee SY, Shin KK, Cho HH,
Bae YC and Jung JS: miR-486-5p induces replicative senescence of
human adipose tissue-derived mesenchymal stem cells and its
expression is controlled by high glucose. Stem Cells Dev.
21:1749–1760. 2012. View Article : Google Scholar
|
|
56
|
Tome M, López-Romero P, Albo C, Sepúlveda
JC, Fernández-Gutiérrez B, Dopazo A, Bernad A and González MA:
miR-335 orchestrates cell proliferation, migration and
differentiation in human mesenchymal stem cells. Cell Death Differ.
18:985–995. 2011. View Article : Google Scholar :
|
|
57
|
Schaap-Oziemlak AM, Raymakers RA,
Bergevoet SM, Gilissen C, Jansen BJ, Adema GJ, Kögler G, le Sage C,
Agami R, van der Reijden BA and Jansen JH: microRNA hsa-miR-135b
regulates mineralization in osteogenic differentiation of human
unrestricted somatic stem cells. Stem Cells Dev. 19:877–885. 2010.
View Article : Google Scholar
|
|
58
|
Ng TK, Carballosa CM, Pelaez D, Wong HK,
Choy KW, Pang CP and Cheung HS: Nicotine alters microRNA expression
and hinders human adult stem cell regenerative potential. Stem
Cells Dev. 22:781–790. 2013. View Article : Google Scholar
|
|
59
|
Chen HC, Lee YS, Sieber M, Lu HT, Wei PC,
Wang CN, Peng HH, Chao AS, Cheng PJ, Chang SD, et al: microRNA and
messenger RNA analyses of mesenchymal stem cells derived from teeth
and the Wharton jelly of umbilical cord. Stem Cells Dev.
21:911–922. 2012. View Article : Google Scholar
|
|
60
|
Dong R, Du J, Wang L, Wang J, Ding G, Wang
S and Fan Z: Comparison of long noncoding RNA and mRNA expression
profiles in mesenchymal stem cells derived from human periodontal
ligament and bone marrow. Biomed Res Int. 2014:3178532014.
View Article : Google Scholar : PubMed/NCBI
|