Introduction
Streptococcus pneumoniae (S.
pneumoniae) is part of the commensal flora of the human
respiratory tract, however, virulent strains of S.
pneumoniae (1,2) or co-infection with other microbes
(3), may transform this commensal
bacterium into a pathogen. Pathogenic S. pneumoniae can
spread to sterile mucosal surfaces and cause otitis media and
pneumonia, or may lead to sepsis or meningitis through invasion of
the host (4). Despite the
currently available antibiotic treatments and vaccines for S.
pneumoniae infections, the annual worldwide morbidity in
children under five years of age reached approximately one million
children, whilst mortality reached around 200,000 children
(5). These rates clearly highlight
the requirement for the development of alternative therapeutic
approaches and reliable assays for determining treatment
effectiveness.
Antibiotic treatments against S. pneumoniae
are currently less effective than they were three decades ago, as a
result of the persistent emergence of antibiotic-resistant strains
(6,7). Anti-S. pneumoniae vaccines in
current use, are based on the capsular polysaccharides of the
bacterium. An example of this includes the Pneumovax vaccine, which
has exhibited 60% effectiveness in preventing invasive pneumococcal
diseases in the elderly (8). The
incomplete coverage occurs as a result of incomplete coverage
against all 97 currently known serotypes or a poor immune response
against certain serotypes included in the vaccine, including
serotype 3. However, Pneumovax does not elicit long-term immune
memory or protective immune responses in infants <2 years of age
(8). To overcome this caveat,
pneumococcal capsular polysaccharides have been conjugated to
various carrier proteins to produce conjugate vaccines. These
currently include between 10 and 15 serotypes (9,10).
However, to date, 98 S. pneumoniae serotypes have been
identified (11,12), and pneumococcal carriage and
disease caused by serotypes that are not included in the conjugate
vaccines are constantly emerging (13,14).
This further emphasizes the need for new therapeutic approaches and
improved vaccines. The development of new preventative and
therapeutic interventions is hampered due to an incomplete
understanding of pneumococcal pathogenesis.
S. pneumoniae colonizes the nasopharynx by
adhering to mucosal cells of the upper respiratory tract, which is
a prerequisite for disease development (15). Therefore, the nature of S.
pneumoniae adhesins has been investigated over the past two
decades (16,17). Among the molecules known to
initiate the S. pneumoniae-host interaction that leads to
nasopharyngeal colonization, include two types of pili. The type I
pilus is an oligomeric appendage, which is encoded by the
rlrA operon (18,19). Its adhesin, the RrgA protein, binds
to the toll-like receptor (TLR) 2 (20). The type II pilus is encoded by a
pathogenicity islet (PI)-II (21).
Following the initial attachment to mucosal cells, the bacteria
partially shed their polysaccharide capsule at the site of adhesion
to provide access to the respiratory mucosa and facilitate the
exposure of adhesive molecules embedded within the bacterial cell
wall or cytoplasmic membrane (22). Among the adhesins present on the
S. pneumoniae membrane and cell wall are the proteins; the
lipoprotein PsaA (23), which
binds to the E-cadherin receptor (24), and the PavA protein (25), which binds to the extracellular
matrix (ECM) protein fibronectin and to the cell adhesion molecule
integrin. Other adhesins include fructose bisphosphate aldolase,
which binds to the flamingo cadherin receptor (26), NADH oxidase, which binds to the ECM
protein laminin α5 (among other putative receptors) (27), and phosphoenolpyruvate protein
phosphotransferase, which binds to the BMP binding endothelial
regulator and Eps 1 proteins, among other putative receptors
(28).
After attachment of the bacteria to the mucosal
cells of the host has been established, S. pneumoniae may
invade the cells either by binding phosphorylcholine or
choline-binding protein A (CbpA), also known as SpsA or PspC to the
platelet-activating factor receptor (PAF-R) (29), which is present in epithelial and
endothelial cells (30–32). This binding may initiate the PAF-R
recycling pathway, which transports bacteria to the basal membrane
of the host and leads to the development of an invasive disease. In
addition, the pneumococcal CbpA may bind to the polymeric
immunoglobulin receptor (pIgR) or to secretory IgA (33–35).
Following attachment to the pIgR, the pneumococci exploit the
recycling pathway to traverse the epithelium from the apical to the
basement membrane. Notably, many adhesins and invasins, including,
PspA, CbpA, PavA, PavB and PhtD, are known to be immunogenic and
elicit a protective immune response in mouse model systems
(36–39).
In the search for effective therapeutic targets,
tissue cultures of cancer-derived or immortalized cell lines are
most often used to identify new molecules that may be involved in
bacterial virulence (22–29). In the current study, these tissue
cultures were examined to investigate whether they are appropriate
models for studying the interactions between S. pneumoniae
and its host. In addition, although a significant amount of
information has been gathered regarding the interaction between
S. pneumoniae and its host cells (22–29),
the extent and specificity of the interaction with primary upper
respiratory cells and secondary lower respiratory cells has not
been fully described. Thus, the present study compared the
abilities of S. pneumoniae to adhere to and invade
nasal-derived epithelial cells (nasal cells), oral-derived
epithelial cells (oral cells) from primary cell lines, as well as
bronchial-virus transformed cells (BEAS-2B) and type II
adenocarcinoma derived epithelial (A549) cells.
The present study investigated the interaction
between non-encapsulated pneumococcal strains and the respiratory
tissue derived epithelial cells, as they have been recently
demonstrated to be involved in S. pneumoniae diseases to a
larger extent than previously assumed (40). Non-encapsulated S.
pneumoniae are subdivided into the following two groups: Group
I, which possess a mutation or deletion in the cps locus,
thereby preventing capsular synthesis; and group II, where the
cps locus is replaced by a gene or genes that code for
proteins, such as two aliB homologues (aliC and
aliD), which are peptide-binding molecules associated with
an ATP-binding cassette transporter (41,42)
or pspK (40). pspK
encodes a protein with a long alpha-helical region containing an
LPxTG motif and a YPT motif known to bind human pIgR (43), necessary for nasopharyngeal
carriage in mice.
Respiratory epithelial cells derived from cancers of
the respiratory system and virus immortalized cells are widely used
as in vitro models for S. pneumoniae infections
(22–29). However, the validity of these cell
lines to reproduce the functions of primary respiratory cells
remains to be fully determined. The present study aimed to compare
the S. pneumoniae adhesion to and invasion of primary
respiratory cells with their interaction with the cancer-derived
cell lines and virus-immortalized cells. The results of the present
study suggest that the adhesion and invasion to the different types
of cells is concomitant with the natural course of infection and
disease development. In addition, the exposure of surface proteins,
which occurs in unencapsulated strains, alters the target cell
preferences of S. pneumoniae. The epithelial cell cultures
described in the current study may be used as a platform for the
future identification of molecules that define the specificity of
S. pneumoniae infection and its affinity to various
tissues.
Materials and methods
Reagents
All chemicals and biochemical reagents were of the
highest purity available and, unless otherwise stated, were
purchased from Sigma-Aldrich; Merck Millipore (Darmstadt,
Germany).
Bacterial strains
The bacterial strains used in the current study
included the encapsulated S. pneumoniae serotype 3 WU2
strain and its unencapsulated variant 3.8-DW (kindly provided by
Professor David Watson, University of Texas, Galveston, TX, USA)
(44), the encapsulated serotype 2
strain D39-LM (provided by Professor Larry S. McDaniel, University
of Mississippi Medical Center, Jackson, MS, USA) and its
unencapsulated variant R6 [American Type Cell Collection (ATCC)]
(45). The encapsulated strains
23F-RD (serotype 23F) and 6B-RD (serotype 6B) were clinical strains
obtained from the collection of the Pediatric Infectious Disease
Unit (Soroka University Medical Center, Beer-Sheva, Israel). S.
pneumoniae were plated onto tryptic soy agar supplemented with
5% sheep erythrocytes (Biological Industries, Beit Haemek, Israel)
and incubated for 17–18 h at 37°C under anaerobic conditions. The
bacteria were then transferred to a Todd-Hewitt broth supplemented
with 0.5% yeast extract (THY), incubated at 37°C until
mid-logarithmic phase (as determined by a growth curve), and
isolates were harvested when the optical density (OD) was equal to
~0.55 at absorbance (A)620. The colony-forming unit
(CFU) content of the preparations was verified in each experiment
by using agar plates supplemented with 5% sheep erythrocytes
(Biological Industries).
Cell culture
Mucosal epithelial cells were isolated from
discarded segments of healthy oral or nasal mucosa, using the
method described by Ueda et al (46) with minor modifications. These
segments were provided by Professor Lipa Bodner (Oral and
Maxillofacial Surgery Unit, Soroka University Medical Center) and
obtained from one patient who underwent oral surgery and one
patient that underwent nasal surgery. The samples were obtained
from healthy areas in the oral and nasal cavities. All human
studies, protocol revisions, and consent procedures were approved
by the Helsinki Ethics Committee of the Soroka University Medical
Center (permit no. 4995). Written informed consent was obtained
from each individual.
Ten mucosal segments (5 mm in diameter) were
immersed for 30 min in a phosphate-buffered saline (PBS) solution
containing penicillin (100 U/ml), streptomycin (100 µg/ml),
gentamycin (50 µg/ml), amphotericin B (2.5 µg/ml) and neomycin
(0.4%). The mucosal segments were then incubated for 16 h at 4°C in
Dulbecco's Modified Eagle's Medium (DMEM) containing the
aforementioned antibiotics, along with dispase (2 U/ml; Roche
Diagnostics GmbH, Mannheim, Germany). The epidermis was peeled off
and incubated in 0.25% trypsin with antibiotics (aforementioned
mixture of penicillin, streptomycin, gentamycin, amphotericin and
neomycin) at 37°C for 60 min, before being transferred to a 10 ml
volume of keratinocyte seeding medium, according to the procedures
described by Rheinwald and Green (47). The keratinocyte seeding medium
contained DMEM: F12 (3:1), fetal calf serum (FCS; 10%), glutamine
(2 mM), penicillin (100 U/ml), streptomycin (100 µg/ml), gentamycin
(50 µg/ml), amphotericin B (2.5 µg/ml), adenine (0.18 mM),
hydrocortisone (0.4 µg/ml), insulin (5 µg/ml), transferrin (5
µg/ml) and tri-iodothyronine (2 nM). In addition to the above, the
keratinocyte growth medium contained cholera toxin (0.1 nM) and
epidermal growth factor (EGF; 5 ng/ml). A single-cell suspension,
obtained following a 2-min vortex, was then plated on a mitomycin
C-treated feeder layer of 3T3 cells (ATCC) in 35-mm diameter tissue
culture plates (Corning Inc., Corning, NY, USA) at a seeding
density of 20,000 cells/cm2. Cultures were incubated in
an environment of 37°C, 8% CO2 and 95% humidity, and the
keratinocyte growth medium was replenished every other day.
Keratinocytes were routinely sub-cultured onto fresh feeder cells
after reaching 80% confluence, following the selective removal of
the feeders with EDTA (0.02% in PBS) and trypsinization (48). Experiments were performed with
nasal and oral mucosal cells at passage four or five. At this
point, cells (2.5×104/well) were seeded in 96-well
plates (Corning Inc.) with a keratinocyte seeding medium in the
absence of feeder cells. The cells were incubated for 24 h without
antibiotics prior to conducting the adhesion assay, at which time
they had reached a density of ~5×104 cells/well.
The virus-transformed bronchial BEAS-2B cell line
(ATCC) was cultured in an LHC-9 medium (Biological Industries)
containing 0.5 ng/ml recombinant EGF, 500 ng/ml hydrocortisone,
0.005 mg/ml insulin, 0.035 mg/ml bovine pituitary extract, 500 nM
ethanolamine, 500 nM phosphoethanolamine, 0.01 mg/ml transferrin,
6.5 ng/ml 3,3′,5-triiodothyronine, 500 ng/ml epinephrine, 0.1 ng/ml
retinoic acid.
The A549 lung adenocarcinoma cell line (ATCC) was
used as it retains the morphological, biochemical, and
immunological characteristics of type II lung epithelial cells
(49–52). The cells were cultured in DMEM
(Biological Industries) supplemented with 10% FCS (Biological
Industries), penicillin and streptomycin (100 µg/ml each). At 24 h
prior to experiments, the cells were transferred to 96-well plates
without antibiotics, and the cultures were then blocked with DMEM
supplemented with 0.5% gelatin and incubated for 4 h at 37°C. The
cell count was, on average, 50,000 cells/well.
Cell adhesion assay
S. pneumoniae from fresh overnight blood agar
cultures were inoculated into the THY broth and cultivated at 37°C
until they reached mid-logarithmic phase (A620,
OD=~0.55). The bacteria were diluted in PBS to a final
concentration of 2×107 CFU/ml. A total of 200 µl diluted
bacteria (~4×106 CFU/200 µl) was added to the
5×104 cultured cells/well (multiplicity of infection
~80:1). Bacteria and cells were incubated for 15, 30, 60 or 90 min
at 37°C. These time points were selected as preliminary experiments
revealed no alteration in the survival of the cells in the presence
of bacteria during this time period. As a negative control, the
same volume of PBS containing the bacteria was added to wells that
did not contain mammalian cells. At each of the denoted time
points, the wells were extensively washed with PBS five times, and
the cells were removed from the wells by incubating the culture
with 0.25% trypsin and 0.02% EDTA for 5 min. Cultured cells were
counted at the end of the experiment and the cell number did not
differ significantly from their number prior to the addition of the
bacteria. The suspension was serially diluted and plated on 5%
sheep blood agar plates, before they were incubated at 37°C under
anaerobic conditions for 17–18 h for CFU determination. For each
experiment, 4–6 replicates were performed at least three times.
Representative experiments are presented in the figures.
Cell invasion assay
All of the initial steps were the same as those used
in the adhesion assay, except that penicillin (100 µg/ml) and
gentamicin (50 µg/ml) were added to the culture media for 45 min at
the same aforementioned time points. The antibiotics in the
supernatant were removed and the cells were removed from the wells
by using 0.25% trypsin and 0.02% EDTA for 5 min. The cells were
then lysed using 0.02% saponin, and the suspension was
serially-diluted and plated on sheep blood agar plates, which were
then incubated under anaerobic conditions for 17–18 h for CFU
determination. For each experiment, six replicates were performed
at least three times.
Statistical analyses
The present study used non-parametric Pearson
analysis to determine alterations in adhesion efficiency over time
(at 15, 30, 60, 90 and 120 min of incubation). One-way analysis of
variance was employed to compare the adhesion and invasion curves
of the different epithelial cells. For multiple comparisons at all
time points the Tukey post-hoc test was used. Statistical analyses
were performed in GraphPad Prism software version 7 (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Adhesion of S. pneumoniae serotype 3
strain WU2 and the unencapsulated derivative 3.8 strain to primary
and secondary respiratory epithelial cells
The adhesion of the WU2 strain to the epithelial
cells increased in a time-dependent manner in all cell types tested
(Fig. 1A; nasal cells, r=0.9995,
P=0.0005; oral cells, r=0.9461, P=0.0149; A549 cells, r=0.948,
P<0.02; BEAS-2B cells, r=0.9707, P=0.006). However, analysis of
all curves (Fig. 1A) revealed that
WU2 adhesion was significantly more efficient to nasal cells
compared with BEAS-2B cells (P=0.0193), A549 cells (P=0.0003) and
oral cells (P=0.0001). Analysis performed at each time point
revealed significantly greater efficiency of WU2 adhesion to nasal
cells than to any of the other types of epithelial cell assessed
(nasal vs. A549, P=0.0003; nasal vs. oral, P=0.0001; nasal vs.
BEAS-2B, P=0.0193). The adhesion of WU2 to BEAS-2B cells was
significantly more efficient than to A549 cells at the 90 and 120
min time points (P=0.0024 and P=0.0295, respectively).
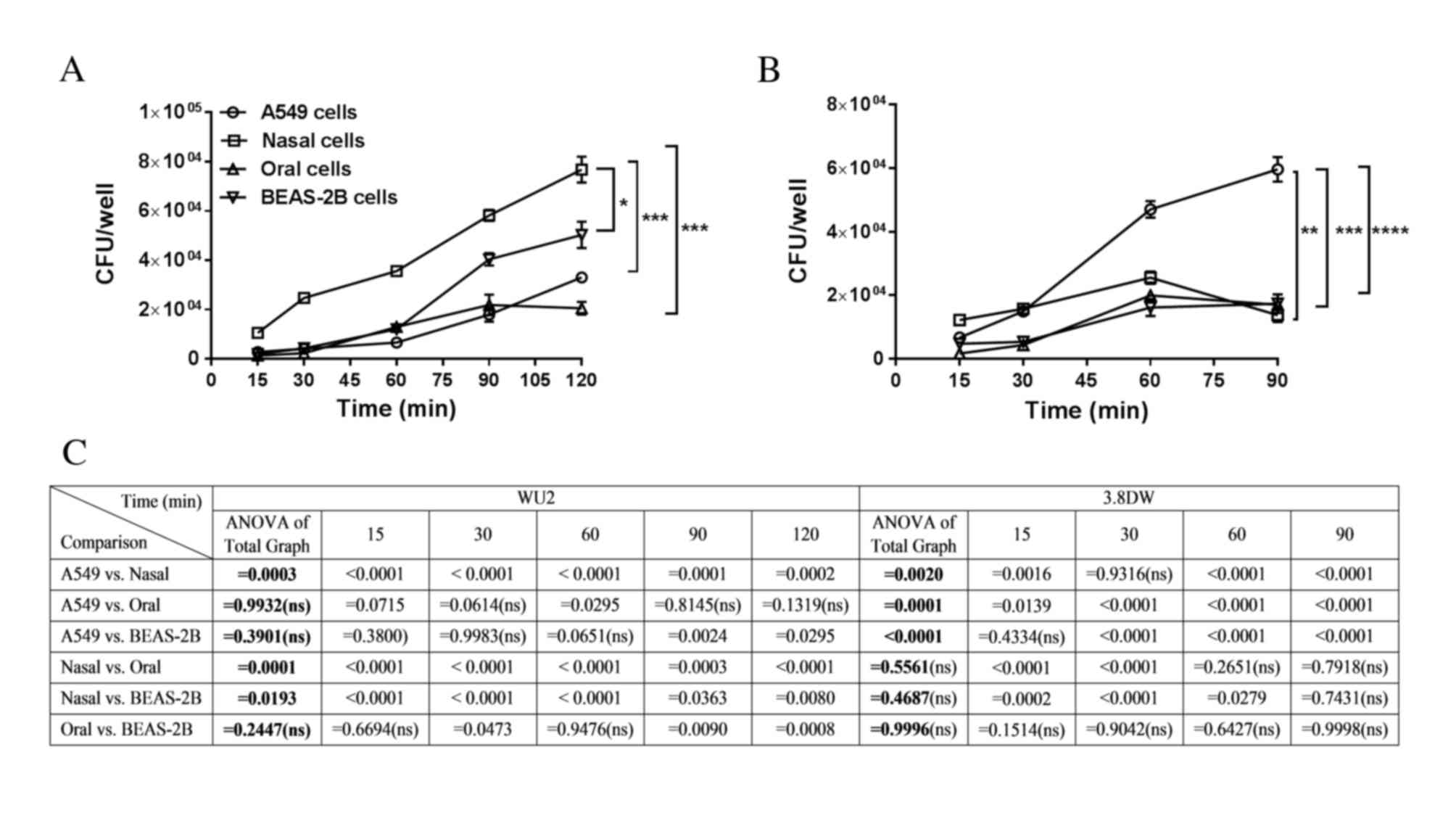 | Figure 1.Adhesion of the encapsulated S.
pneumoniae strain WU2 and its unencapsulated derivative strain
3.8-DW to A549, oral, nasal and BEAS-2B respiratory epithelial
cells. (A) Adhesion of WU2 increased in a time-dependent manner in
all cultured epithelial cells examined (nasal cells, r=0.9995,
P=0.0005; oral cells, r=0.9461, P=0.0149; A549 cells, r=0.948,
P<0.02; BEAS-2B cells, r=0.9707, P=0.006). *P<0.05 and
***P<0.001. (B) The adhesion of the unencapsulated S.
pneumoniae strain 3.8-DW to A549 cells was time-dependent
(r=0.9835, P=0.0165), and adhesion to BEAS-2B cells demonstrated a
tendency to be time-dependent (r=0.941, P=0.059). **P<0.01;
***P<0.001; and ****P<0.0001. (C) Statistical analysis for
WU2 and 3.8DW adhesion for whole curve analysis comparisons (ANOVA
of total graph) and at each time point. CFU, colony-forming units;
S. pneumoniae; Streptococcus pneumonia; ANOVA, analysis of
variance. |
Next, the adherence experiments were repeated using
the unencapsulated S. pneumoniae strain 3.8-DW (Fig. 1B). Adhesion to the A549 cells was
time-dependent (r=0.9835, P=0.0165) and adhesion to BEAS-2B cells
demonstrated a tendency toward time-dependency (r=0.941, P=0.059;
Fig. 1B). Analysis of all curves
revealed that 3.8-DW adhesion to A549 cells was significantly
greater compared with the adhesion to the other cell types (A549
vs. nasal, P=0.0020; A549 vs. oral, P=0.0001; A549 vs. BEAS-2B,
P<0.0001). In addition, analysis of the individual time points
revealed that adhesion of 3.8-DW to nasal cells was significantly
greater than to BEAS-2B cells at the 15, 30 and 60 min time points
(P=0.0002, P<0.0001 and P=0.0279, respectively; Fig. 1B and C).
Invasion of the encapsulated S.
pneumoniae WU2 strain and its unencapsulated derivative strain
3.8-DW to primary and secondary respiratory epithelial cells
In order to determine the extent of invasion of the
WU2 strain to the cultured cells, antibiotics were added to the
cell cultures at the denoted time points for 45 min prior to
determination of bacterial count. Notably, the invasion of nasal
cells compared with all other cells types increased within the
first 60 min reaching the significance of P<0.0001 at 30 min,
and decreased thereafter (Fig.
2A), possibly mimicking the transcytosis process that occurs in
nasal mucosal cells. However, after 60 min the invasion of nasal
cells remained significantly greater in comparison to the other
cells types (P<0.0001). Cell invasion of A549 and BEAS-2B cells
was time-dependent (r=0.9728, P=0.0054; and r=0.9787, P=0.0037,
respectively); however, the invasion of oral cells was not.
Consistently, WU2 invasion of BEAS-2B and A549 cells was
significantly greater compared to the invasion of oral cells
(P=0.002 and P=0.0008, respectively; Fig. 2A). The invasion of WU2 into BEAS-2B
cells was not significantly different from the invasion of A549
cells at any of the examined time points.
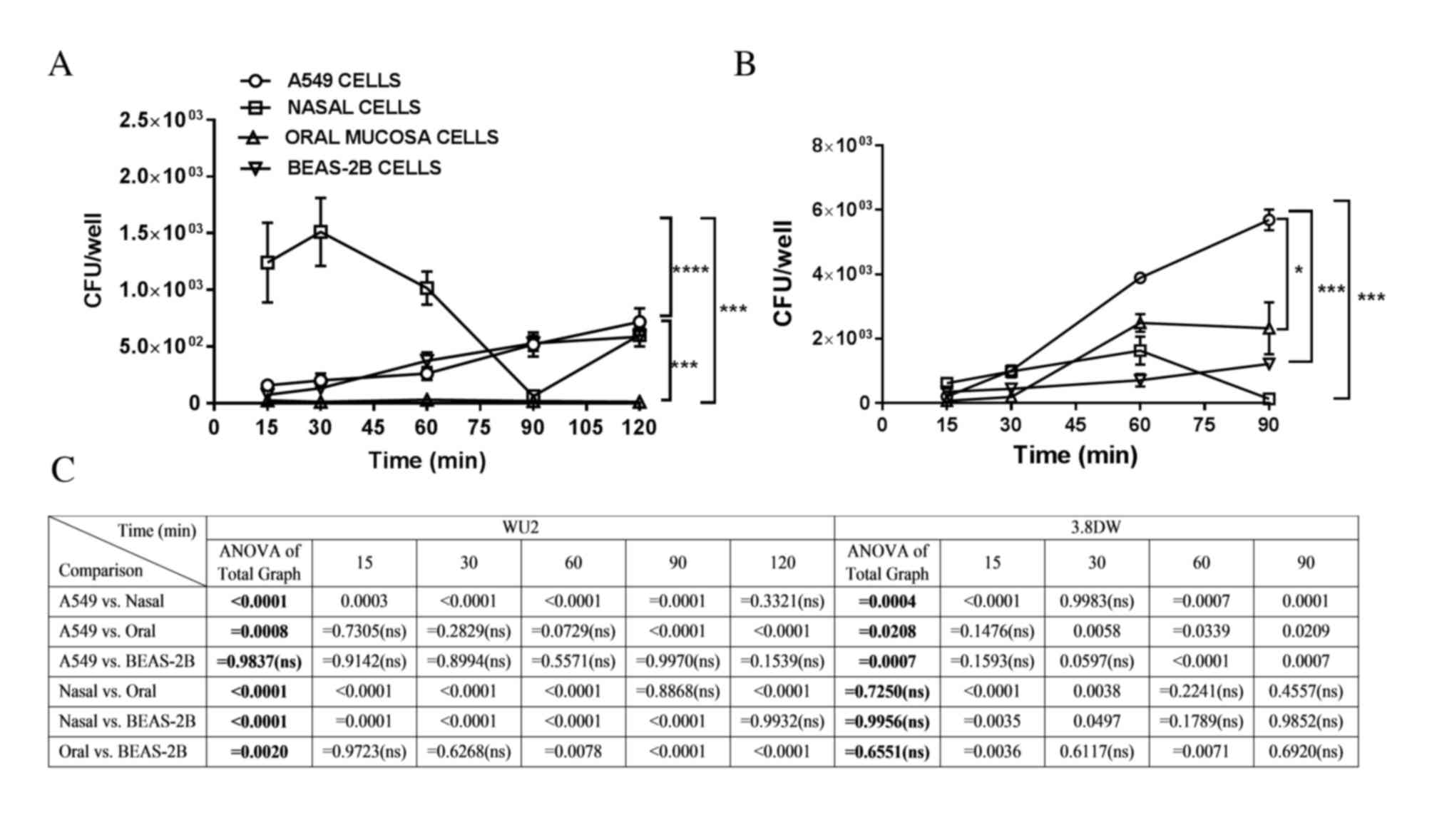 | Figure 2.Invasion of the encapsulated S.
pneumoniae strain WU2 and its unencapsulated derivative strain
3.8-DW to A549, nasal, oral and BEAS-2B respiratory epithelial
cells. In order to determine invasion capabilities, extracellular
bacteria were eliminated by adding antibiotics to the culture at
the denoted time points, for 45 min prior to determination of
bacterial counts. (A) Invasion of WU2 strain to cells at the
indicated time points. The invasion of nasal cells vs. all other
cells types was significantly greater within the first 60 min,
reaching the significance of P<0.0001 at 30 min. Cell invasion
increased continuously in A549 and in BEAS-2B cells (r=0.9728,
P=0.0054; and r=0.9787, P=0.0037, respectively). (B) Invasion of
the unencapsulated strain 3.8DW to A549 and BEAS-2B cells was
time-dependent (r=0.9946, P=0.0054; and r=0.9798 P=0.0202,
respectively). Analysis of all curves indicated that invasion of
A549 cells was significantly more efficient than to nasal, oral,
and BEAS-2B cells (P=0.0004, P=0.0208 and P=0.0007, respectively).
No significant differences in invasion efficiency were observed
among the nasal, oral and BEAS-2B cell types. *P<0.05;
***P<0.001; and ****P<0.0001. (C) Statistical analysis for
WU2 and 3.8DW invasion for whole curve analysis comparisons (ANOVA
of total graph) and at each time point. CFU, colony-forming units;
S. pneumoniae; Streptococcus pneumoniae; ANOVA, analysis of
variance; ns, non significant. |
The invasion of the 3.8-DW unencapsulated strain of
A549 and to BEAS-2B cells was time-dependent (r=0.9946, P=0.0054;
and r=0.9798, P=0.0202, respectively; Fig. 2B). Analysis of all curves revealed
that invasion of A549 cells was significantly higher when compared
with nasal, oral and BEAS-2B cells (P=0.0004, P=0.0208 and
P=0.0007, respectively; Fig. 2B and
C). 3.8-DW invasion of A549 cells at each time point was
significantly greater compared with oral cells at 30, 60 and 90 min
(P=0.0058, P=0.0339 and P=0.0209, respectively). The invasion of
3.8-DW into A549 cells was significantly greater than into nasal
cells at 60 and 90 min time points (P=0.0007 and P=0.0001,
respectively). The invasion into A549 cells was significantly
greater than into BEAS-2B cells at the 60 and 90 min time point
(P<0.0001 and P=0.0007, respectively). A total of 90 min
following inoculation, invasion of nasal cells decreased (Fig. 2B and C), similar to the decrease
observed for the WU2 strain invasion of nasal cells (Fig. 2A). Invasion of nasal cells was not
significantly different when compared with oral cells.
Adhesion and invasion of encapsulated
and unencapsulated S. pneumoniae strains to A549 cells
The next aim of the study was to compare the
adhesion to and invasion of A549 cells of selected encapsulated and
unencapsulated S. pneumoniae strains. The encapsulated
strains included the following: i) The WU2 serotype 3 strain, which
has the thickest capsule among all strains tested in the current
study, and is considered to be a highly invasive strain (53–55);
ii) the D39-LM serotype, which is widely used for studying S.
pneumoniae pathogenesis (28);
iii) the 6B-RD serotype, which was isolated from the nasopharynx of
a healthy child and was previously demonstrated to colonize the
nasopharynx but not disseminate to the lungs or any other tissue
(56,57); and iv) the 23F-RD serotype, which
is considered to be pathogenic, as its capsule was included in the
7 valent conjugate vaccine (58).
The unencapsulated strains included the 3.8-DW, which is a derived
from WU2 (44) and the R6 strain
(45), which was derived from the
D39 strain. Although the results presented so far indicated that
the encapsulated WU2 strain adhered and invaded most efficiently to
nasal epithelial cells, these cells are very difficult to obtain
and maintain. Therefore, the adhesion to and invasion of A549 cells
by the various encapsulated and unencapsulated strains were
assessed.
All strains indicated a time-dependent adhesion to
A549 cells (WU2, r=0.9487, P=0.0139; D39-LM, r=0.9866, P=0.0019;
6B-RD, r=0.9809, P=0.0032; 23F-RD, r=0.9424, P=0.0165; 3.8-DW,
r=8955, P=0.0399; R6, r=0.9936, P=0.0006; Fig. 3A). A whole-curve comparison
revealed that the adhesion of the unencapsulated strain 3.8-DW was
significantly higher when compared to that of the encapsulated
strains (6B-RD, P=0.004; 23F-RD P<0.0001; D39-LM, P<0.0001;
WU2, P<0.0001), but was not significantly different from the
adhesion of the unencapsulated strain R6 (Fig. 3A). In addition, whole-curve
analysis indicated that the adhesion of the R6 strain was
significantly higher than the adhesion of any of the encapsulated
strains (23F-RD, P=0.004; D39-LM, P<0.0001; WU2, P=0.0007)
except for 6B-RD (Fig 3A).
Significant differences in adherence among the unencapsulated
3.8-DW and encapsulated strains were observed at each of the
individual time points examined except for 3.8-DW vs. 6B-RD at the
30 min time point. The strain 3.8-DW adhered more efficiently to
the A549 cells up to 60 min of incubation. The adherence of the
unencapsulated R6-RD was significantly greater than the
encapsulated strains in all time points after 15 min of incubation.
The adhesion of 6B-RD was significantly greater than that of the
23F-RD between 30 and 60 min of incubation, D39 after 15 min of
incubation and WU2 strains at the 30 min time point.
 | Figure 3.Adhesion and invasion of encapsulated
and unencapsulated clinical and laboratory S. pneumoniae
strains to A549 cells. (A) The S. pneumoniae unencapsulated
strains 3.8-DW and R6, and the encapsulated strains 23F-RD, 6B-RD,
WU2, and D39-LM, were incubated with A549 cells for 15, 30, 60, 90
or 120 min. All strains demonstrated time-dependent adhesion to the
A549 cells (WU2, r=0.9487, P=0.0139; D39-LM, r=0.9866, P=0.0019;
6B-RD, r=0.9809, P=0.0032; 23F-RD, r=0.9424, P=0.0165; 3.8-DW,
r=8955, P=0.0399; R6, r=0.9936, P=0.0006). (B) Invasion of the
6B-RD, R6, D39-LM, and WU2 strains to A549 cells was time-dependent
(6B-RD, r=0.9416, P=0.0168; R6, r=0.9628, P<0.0086; D39-LM,
r=0.9588, P=0.01; WU2, r=0.9728, P<0.0054). The unencapsulated
strain 3.8-DW demonstrated the highest invasion capabilities out of
all strains examined in the full-curve analysis. **P<0.01;
***P<0.001; and ****P<0.0001. (C) Statistical analysis for
adhesion and invasion of the full curve comparisons (ANOVA of total
graph) and at each time point. CFU, colony-forming units; S.
pneumoniae; Streptococcus pneumoniae; ANOVA, analysis of
variance; ns, non significant. |
The invasion of the 6B-RD, R6, D39-LM, and WU2
strains to the A549 cells was time-dependent (6B-RD, r=0.9416,
P=0.0168; R6, r=0.9628, P<0.0086; D39-LM, r=0.9588, P=0.01; WU2,
r=0.9728, P<0.0054; Fig. 3B).
The invasion of the unencapsulated 3.8-DW strain was significantly
greater compared with all the other strains assessed, by
whole-curve analysis and at each of the examined time points,
except in comparison to R6-DW at the 30 min time point (Fig. 3B and C).
Discussion
Cell lines are frequently used to understand the
interactions between S. pneumoniae and its host (26,27,52,59–64).
Notably, the lung-derived adenocarcinoma A549 cell line and the
virus-transformed BEAS-2B bronchial cell line have been used for
studying the function of virulence factors and the alterations that
take place in the bacterium and the host cell upon bacteria-host
interactions (3,65–68).
However, the suitability of these cell lines as in vitro
models for investigating the interaction between S.
pneumoniae and nasal or lung mucosal cells has not been
verified directly. To the best of our knowledge, the present study
is the first to demonstrate that the extent of adhesion and
invasion of a highly encapsulated pathogenic S. pneumoniae
strain (WU2) into nasal cells is superior in comparison to the
extent of its adhesion and invasion into primary oral cells and
into the cancer-derived or virus-transformed respiratory cell
lines.
The adhesion of S. pneumoniae to the upper
respiratory tract is a prerequisite for its colonization and is
therefore considered to be a major risk factor for the development
of pneumococcal disease (3,69,70).
The respiratory epithelial cells are the first line of defense of
the innate immune system, and the interaction between pneumococci
and the respiratory epithelial cells activates signal transduction
cascades, in which p38 mitogen-activated protein kinase-dependent
nuclear factor-κB activation upregulates the expression of
interleukin-8 (71). The
activation of this cascade may be the result of the bacteria
binding to innate immune system-associated pattern recognition
receptors on the surface of mammalian cells (e.g. TLRs) (72), or to intracellular pattern
recognition receptors, such as the nucleotide-binding
oligomerization domain proteins (73). In addition, S. pneumoniae
may bind to additional human membrane receptors, which are capable
of activating different signal transduction pathways (74), which may increase cytokine
expression. As a result, PAF-R expression is increased, which
increases the binding of S. pneumoniae to the cells
(29).
In the current study, the WU2 strain adhered to all
cell types in a time-dependent manner. The adhesion of WU2 to nasal
cells was more efficient than the adhesion to other cells tested
throughout the incubation period. In addition, WU2 invasion of
nasal cells was significantly more efficient when compared with the
other cell types examined. Invasion of the WU2 strain to the A549
and BEAS-2B cell lines was significantly higher than its invasion
of oral cells. At 30 min following inoculation of nasal cells with
the WU2 strain, a decline in the number of bacteria in these cells
was observed, which decreased further at 60 and 90 min time points.
Following adherence, S. pneumoniae is able to invade nasal
cells by binding of bacterial phosphorylcholine or CbpA to the
PAF-R of the host cell (29); the
latter also binds to the pIgR (33), which allows subsequent exploitation
of the recycling pathways. This facilitates transcytosis through
the mucosal epithelial cell layer, and leads to the development of
invasive disease. PAF-R is present in epithelial and endothelial
cells (29,75,76).
The authors of the current study hypothesized that adhesion and
invasion of S. pneumoniae strains to nasal cells may occur
through these receptors or through other, as yet unknown receptors,
which recycle and expel the WU2 strain more efficiently from the
nasal cells than from the other types of cells examined. It has
been demonstrated that S. pneumoniae adhere to and penetrate
the nasopharynx to gain access to the middle ear via the Eustachian
tube (77,78). Thus, bacterial strains capable of
invading nasopharyngeal cells may be those that disseminate and
cause invasive disease (79). Out
of all cell types examined in the present study, the adhesion and
invasion of the WU2 strain to oral cells was the least efficient.
The oral mucosa is not a primary site for S. pneumoniae
colonization, although adhesion of S. pneumoniae to primary
oral cells in vitro has been reported (80). Ultimately, differences in capsule
size or the differential expression of virulence factors involved
in the invasion process, may explain the ability of pathogenic
pneumococci to adhere and invade different types of respiratory
epithelial cells in different manners.
The adhesion of the unencapsulated strain 3.8-DW (a
derivative of the encapsulated WU2 strain) (81) to A549 cells was time-dependent.
However, unlike the WU2 strain, the 3.8-DW strain most efficiently
adhered to and invaded A549 cells. This alteration in adhesion and
invasion preference may be due to the exposure of surface proteins
that are otherwise masked by the capsule, which is a major
virulence factor that protects the bacterium from phagocytosis and
prolongs its survival in phagocytes (81,82).
The authors attribute these differences in adhesion and invasion to
the absence of the capsule, which allows the cell wall proteins to
be exposed and available for interaction with the host cells. It
has not yet been determined whether there are any genetic
differences between the encapsulated and their unencapsulated
derivatives. In addition, a different pattern of mRNA and protein
expression in the unencapsulated strain compared with the parental
strain may exist.
Previous studies have suggested that the capsule
prevents bacterial adhesion to epithelial cells (62,83,84).
In addition, partial capsule shedding at the site of adhesion is a
prerequisite for S. pneumoniae adhesion and invasion, and
variations in the capsule size of different strains or variants
affects the extent of adhesion (22,85–88).
In general, unencapsulated S. pneumoniae strains are less
virulent than encapsulated strains; a phenomenon primarily
associated to the lack of the capsular polysaccharide shield
(89). However, infection with
unencapsulated strains leads to streptococcal ocular keratitis in a
rabbit model system (90) and
conjunctivitis in humans (91).
The unencapsulated strains 3.8-DW and R6
demonstrated similar adhesion efficiencies in the A549 cells, and
were significantly higher than that of the other encapsulated
strains tested. These results are consistent with a previous study
(22). In contrast to their
similar adhesion capabilities, invasion of A549 cells by the R6
strain was as inefficient as that of the encapsulated strains when
compared to the 3.8-DW strain. An increasing number of reports have
identified non-encapsulated S. pneumoniae as a
disease-causing pathogen (40).
This phenomenon may originate from the introduction of the
pneumococcal conjugate vaccine in 2000, or from an improved
identification of the smaller colonies of non-encapsulated S.
pneumoniae in microbiology laboratories (40). Keller et al (40) described the different
characteristics of non-encapsulated S. pneumoniae obtained
from patients with nasopharyngeal colonization or invasive
pneumococcal diseases. In addition, a recent report demonstrated
the involvement of non-encapsulated S. pneumoniae as a cause
of chronic adenoiditis (92).
In conclusion, the results of the present study
provide evidence of an association between the ability of
pathogenic encapsulated S. pneumoniae to adhere to and
invade specific respiratory epithelial cells in vitro and
the natural course of S. pneumoniae-induced diseases. In
addition, the results demonstrate that alternative cell cultures
may be used to further contribute to research concerning the
interaction between S. pneumoniae and its host. Whereas the
cancer-derived cell lines are informative, primary cultures may
assist in deciphering specific interactions and the identification
of molecules responsible for the affinity of S. pneumoniae
to specific tissues. Furthermore, cell origin and type may cause
differences that should be taken into consideration when drawing
conclusions and translating to in vivo cell biology.
Acknowledgements
The present study was supported by The Israel
Ministry of Health (grant nos. 4476, 5540 and 3000003867), BG Negev
Technologies, Ben-Gurion University of the Negev (grant no.
80,904,101), The Center of Emerging Diseases (grant no. 2506), and
the Israel Academy of Science (grant no. 613/04). This manuscript
is written in the memory of Professor Nili Grossman.
References
|
1
|
Weinberger DM, Harboe ZB, Sanders EA,
Ndiritu M, Klugman KP, Rückinger S, Dagan R, Adegbola R, Cutts F,
Johnson HL, et al: Association of serotype with risk of death due
to pneumococcal pneumonia: A meta-analysis. Clin Infect Dis.
51:692–699. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Harvey RM, Trappetti C, Mahdi LK, Wang H,
McAllister LJ, Scalvini A, Paton AW and Paton JC: The variable
region of pneumococcal pathogenicity island 1 is responsible for
unusually high virulence of a serotype 1 isolate. Infect Immun.
84:822–832. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vernatter J and Pirofski LA: Current
concepts in host-microbe interaction leading to pneumococcal
pneumonia. Curr Opin Infect Dis. 26:277–283. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simell B, Auranen K, Kayhty H, Goldblatt
D, Dagan R and O'Brien KL: Pneumococcal Carriage Group: The
fundamental link between pneumococcal carriage and disease. Expert
Rev Vaccines. 11:841–855. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
van der Poll T and Opal SM: Pathogenesis,
treatment, and prevention of pneumococcal pneumonia. Lancet.
374:1543–1556. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim L, McGee L, Tomczyk S and Beall B:
Biological and epidemiological features of antibiotic-resistant
Streptococcus pneumoniae in pre- and post-conjugate vaccine eras: A
United States perspective. Clin Microbiol Rev. 29:525–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brauner A, Fridman O, Gefen O and Balaban
NQ: Distinguishing between resistance, tolerance and persistence to
antibiotic treatment. Nat Rev Microbiol. 14:320–330. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hammitt LL, Bulkow LR, Singleton RJ,
Nuorti JP, Hummel KB, Miernyk KM, Miernyk KM, Zanis C, Whaley M and
Romero-Steiner S: Repeat revaccination with 23-valent pneumococcal
polysaccharide vaccine among adults aged 55–74 years living in
Alaska: No evidence of hyporesponsiveness. Vaccine. 29:2287–2295.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Käyhty H and Eskola J: New vaccines for
the prevention of pneumococcal infections. Emerg Infect Dis.
2:289–298. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Golden AR, Adam HJ and Zhanel GG: Canadian
Antimicrobial Resistance A: Invasive Streptococcus pneumoniae in
Canada, 2011–2014: Characterization of new candidate 15-valent
pneumococcal conjugate vaccine serotypes 22F and 33F. Vaccine.
34:2527–2530. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jauneikaite E, Tocheva AS, Jefferies JM,
Gladstone RA, Faust SN, Christodoulides M, Hibberd ML and Clarke
SC: Current methods for capsular typing of Streptococcus
pneumoniae. J Microbiol Methods. 113:41–49. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Geno KA, Gilbert GL, Song JY, Skovsted IC,
Klugman KP, Jones C, Konradsen HB and Nahm MH: Pneumococcal
capsules and their types: Past, present, and future. Clin Microbiol
Rev. 28:871–899. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hicks LA, Harrison LH, Flannery B, Hadler
JL, Schaffner W, Craig AS, Jackson D, Thomas A, Beall B, Lynfield
R, et al: Incidence of pneumococcal disease due to non-pneumococcal
conjugate vaccine (PCV7) serotypes in the United States during the
era of widespread PCV7 vaccination, 1998–2004. J Infect Dis.
196:1346–1354. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dagan R: Serotype replacement in
perspective. Vaccine. 27:(Suppl 3). C22–C24. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shak JR, Vidal JE and Klugman KP:
Influence of bacterial interactions on pneumococcal colonization of
the nasopharynx. Trends in Microbiol. 21:129–135. 2013. View Article : Google Scholar
|
|
16
|
Hammerschmidt S: Adherence molecules of
pathogenic pneumococci. Curr Opin Microbiol. 9:12–20. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paterson GK and Orihuela CJ: Pneumococcal
microbial surface components recognizing adhesive matrix molecules
targeting of the extracellular matrix. Mol Microbiol. 77:1–5. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Harfouche C, Filippini S, Gianfaldoni C,
Ruggiero P, Moschioni M, Maccari S, Pancotto L, Arcidiacono L,
Galletti B, Censini S, et al: RrgB321, a fusion protein of the
three variants of the pneumococcal pilus backbone rrgb, is
protective in vivo and elicits opsonic antibodies. Infect Immun.
80:451–460. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moschioni M, Donati C, Muzzi A, Masignani
V, Censini S, Hanage WP, Bishop CJ, Reis JN, Normark S,
Henriques-Normark B, et al: Streptococcus pneumoniae contains 3
rlrA pilus variants that are clonally related. J Infect Dis.
197:888–896. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Basset A, Zhang F, Benes C, Sayeed S, Herd
M, Thompson C, Golenbock DT, Camilli A and Malley R: Toll-like
receptor (TLR) 2 mediates inflammatory responses to oligomerized
RrgA pneumococcal pilus type 1 protein. J Biol Chem. 288:2665–2675.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bagnoli F, Moschioni M, Donati C,
Dimitrovska V, Ferlenghi I, Facciotti C, Muzzi A, Giusti F, Emolo
C, Sinisi A, et al: A second pilus type in Streptococcus pneumoniae
is prevalent in emerging serotypes and mediates adhesion to host
cells. J Bacteriol. 190:5480–5492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hammerschmidt S, Wolff S, Hocke A, Rosseau
S, Muller E and Rohde M: Illustration of pneumococcal
polysaccharide capsule during adherence and invasion of epithelial
cells. Infect Immun. 73:4653–4667. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Berry AM and Paton JC: Sequence
heterogeneity of PsaA, a 37-kilodalton putative adhesin essential
for virulence of Streptococcus pneumoniae. Infect Immun.
64:5255–5262. 1996.PubMed/NCBI
|
|
24
|
Anderton JM, Rajam G, Romero-Steiner S,
Summer S, Kowalczyk AP, Carlone GM, Sampson JS and Ades EW:
E-cadherin is a receptor for the common protein pneumococcal
surface adhesin A (PsaA) of Streptococcus pneumoniae. Microb
Pathog. 42:225–236. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pracht D, Elm C, Gerber J, Bergmann S,
Rohde M, Seiler M, Kim KS, Jenkinson HF, Nau R and Hammerschmidt S:
PavA of Streptococcus pneumoniae modulates adherence, invasion, and
meningeal inflammation. Infect Immun. 73:2680–2699. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blau K, Portnoi M, Shagan M, Kaganovich A,
Rom S, Kafka D, Caspi V Chalifa, Porgador A, Givon-Lavi N, Gershoni
JM, et al: Flamingo cadherin: A putative host receptor for
Streptococcus pneumoniae. J Infect Dis. 195:1828–1837. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Muchnik L, Adawi A, Ohayon A, Dotan S,
Malka I, Azriel S, Shagan M, Portnoi M, Kafka D, Nahmani H, et al:
NADH oxidase functions as an adhesin in Streptococcus pneumoniae
and elicits a protective immune response in mice. PloS One.
8:e611282013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nebenzahl Y Mizrachi, Blau K, Kushnir T,
Shagan M, Portnoi M, Cohen A, Azriel S, Malka I, Adawi A, Kafka D,
et al: Streptococcus pneumoniae cell-wall-localized
phosphoenolpyruvate protein phosphotransferase can function as an
adhesin: Identification of its host target molecules and evaluation
of its potential as a vaccine. PloS One. 11:e01503202016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cundell DR, Gerard NP, Gerard C,
Idanpaan-Heikkila I and Tuomanen EI: Streptococcus pneumoniae
anchor to activated human cells by the receptor for
platelet-activating factor. Nature. 377:435–438. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shivshankar P, Boyd AR, Le Saux CJ, Yeh IT
and Orihuela CJ: Cellular senescence increases expression of
bacterial ligands in the lungs and is positively correlated with
increased susceptibility to pneumococcal pneumonia. Aging Cell.
10:798–806. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Nagaoka K, Yanagihara K, Morinaga Y,
Nakamura S, Harada T, Hasegawa H, Izumikawa K, Ishimatsu Y, Kakeya
H, Nishimura M and Kohno S: Prevotella intermedia induces severe
bacteremic pneumococcal pneumonia in mice with upregulated
platelet-activating factor receptor expression. Infect Immun.
82:587–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Suri R, Periselneris J, Lanone S,
Zeidler-Erdely PC, Melton G, Palmer KT, Andujar P, Antonini JM,
Cohignac V, Erdely A, et al: Exposure to welding fumes and lower
airway infection with Streptococcus pneumoniae. J Allergy Clin
Immunol. 137:527–534.e7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rosenow C, Ryan P, Weiser JN, Johnson S,
Fontan P, Ortqvist A and Masure HR: Contribution of novel
choline-binding proteins to adherence, colonization and
immunogenicity of Streptococcus pneumoniae. Mol Microbiol.
25:819–829. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hammerschmidt S, Talay SR, Brandtzaeg P
and Chhatwal GS: SpsA, a novel pneumococcal surface protein with
specific binding to secretory immunoglobulin A and secretory
component. Mol Microbiol. 25:1113–1124. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Brooks-Walter A, Briles DE and
Hollingshead SK: The pspC gene of Streptococcus pneumoniae encodes
a polymorphic protein, PspC, which elicits cross-reactive
antibodies to PspA and provides immunity to pneumococcal
bacteremia. Infect Immun. 67:6533–6542. 1999.PubMed/NCBI
|
|
36
|
Gamez G and Hammerschmidt S: Combat
pneumococcal infections: Adhesins as candidates for protein-based
vaccine development. Curr Drug Targets. 13:323–337. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Plumptre CD, Ogunniyi AD and Paton JC:
Polyhistidine triad proteins of pathogenic streptococci. Trends
Microbiol. 20:485–493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Khan MN and Pichichero ME: Vaccine
candidates PhtD and PhtE of Streptococcus pneumoniae are adhesins
that elicit functional antibodies in humans. Vaccine. 30:2900–2907.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kallio A, Sepponen K, Hermand P, Denoël P,
Godfroid F and Melin M: Role of Pht proteins in attachment of
Streptococcus pneumoniae to respiratory epithelial cells. Infect
Immun. 82:1683–1691. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Keller LE, Robinson DA and McDaniel LS:
Nonencapsulated Streptococcus pneumoniae: Emergence and
pathogenesis. MBio. 7:e017922016.PubMed/NCBI
|
|
41
|
Alloing G, de Philip P and Claverys JP:
Three highly homologous membrane-bound lipoproteins participate in
oligopeptide transport by the Ami system of the gram-positive
Streptococcus pneumoniae. J Mol Biol. 241:44–58. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Claverys JP, Grossiord B and Alloing G: Is
the Ami-AliA/B oligopeptide permease of Streptococcus pneumoniae
involved in sensing environmental conditions? Res Microbiol.
151:457–463. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park IH, Kim KH, Andrade AL, Briles DE,
McDaniel LS and Nahm MH: Nontypeable pneumococci can be divided
into multiple cps types, including one type expressing the novel
gene pspK. mBio. 3:e00035–e00112. 2012.PubMed/NCBI
|
|
44
|
Watson DA and Musher DM: Interruption of
capsule production in Streptococcus pneumonia serotype 3 by
insertion of transposon Tn916. Infect Immun. 58:3135–3138.
1990.PubMed/NCBI
|
|
45
|
Carvalho SM, Kuipers OP and Neves AR:
Environmental and nutritional factors that affect growth and
metabolism of the pneumococcal serotype 2 strain D39 and its
nonencapsulated derivative strain R6. PloS One. 8:e584922013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ueda M, Hata K, Horie K and Torii S: The
potential of oral mucosal cells for cultured epithelium: A
preliminary report. Ann Plast Surg. 35:498–504. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rheinwald JG and Green H: Serial
cultivation of strains of human epidermal keratinocytes: The
formation of keratinizing colonies from single cells. Cell.
6:331–343. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Rheinwald J: Methods for clonal growth and
serial cultivation of normal human epidermal keratinocytes and
mesothelial cellsCell Growth and Division: A Practical Approach.
Baserga R: IRL Press; Oxford: pp. 81–94. 1989
|
|
49
|
Lieber M, Smith B, Szakal A, Nelson-Rees W
and Todaro G: A continuous tumor-cell line from a human lung
carcinoma with properties of type II alveolar epithelial cells. Int
J Cancer. 17:62–70. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Balis JU, Bumgarner SD, Paciga JE,
Paterson JF and Shelley SA: Synthesis of lung surfactant-associated
glycoproteins by A549 cells: Description of an in vitro model for
human type II cell dysfunction. Exp Lung Res. 6:197–213. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Asano K, Chee CB, Gaston B, Lilly CM,
Gerard C, Drazen JM and Stamler JS: Constitutive and inducible
nitric oxide synthase gene expression, regulation, and activity in
human lung epithelial cells. Proc Natl Acad Sci USA.
91:10089–10093. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bergmann S, Schoenen H and Hammerschmidt
S: The interaction between bacterial enolase and plasminogen
promotes adherence of Streptococcus pneumoniae to epithelial and
endothelial cells. Int J Med Microbiol. 303:452–462. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Briles DE, Nahm M, Schroer K, Davie J,
Baker P, Kearney J and Barletta R: Antiphosphocholine antibodies
found in normal mouse serum are protective against intravenous
infection with type 3 Streptococcus pneumoniae. J Exp Med.
153:694–705. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Nebenzahl Y Mizrachi, Porat N, Lifshitz S,
Novick S, Levi A, Ling E, Liron O, Mordechai S, Sahu RK and Dagan
R: Virulence of Streptococcus pneumoniae may be determined
independently of capsular polysaccharide. FEMS Microbiol Lett.
233:147–152. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sahu RK, Mordechai S, Pesakhov S, Dagan R
and Porat N: Use of FTIR spectroscopy to distinguish between
capsular types and capsular quantities in Streptococcus pneumoniae.
Biopolymers. 83:434–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mizrachi-Nebenzahl Y, Lifshitz S,
Teitelbaum R, Novick S, Levi A, Benharroch D, Ling E and Dagan R:
Differential activation of the immune system by virulent
Streptococcus pneumoniae strains determines recovery or death of
the host. Clin Exp Immunol. 134:23–31. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ling E, Feldman G, Dagan R and
Mizrachi-Nebenzahl Y: Cytokine mRNA expression in pneumococcal
carriage, pneumonia, and sepsis in young mice. J Infect Dis.
188:1752–1766. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
58
|
Whitney CG, Farley MM, Hadler J, Harrison
LH, Bennett NM, Lynfield R, Reingold A, Cieslak PR, Pilishvili T,
Jackson D, et al: Decline in invasive pneumococcal disease after
the introduction of protein-polysaccharide conjugate vaccine. N
Engl J Med. 348:1737–1746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Daniely D, Portnoi M, Shagan M, Porgador
A, Givon-Lavi N, Ling E, Dagan R and Nebenzahl Y Mizrachi:
Pneumococcal 6-phosphogluconate-dehydrogenase, a putative adhesin,
induces protective immune response in mice. Clin Exp Immunol.
144:254–163. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Nebenzahl Y Mizrachi, Bernstein A, Portnoi
M, Shagan M, Rom S, Porgador A and Dagan R: Streptococcus
pneumoniae surface-exposed glutamyl tRNA synthetase, a putative
adhesin, is able to induce a partially protective immune response
in mice. J Infect Dis. 196:945–953. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zahlten J, Kim YJ, Doehn JM, Pribyl T,
Hocke AC, Garcia P, Hammerschmidt S, Suttorp N, Hippenstiel S and
Hübner RH: Streptococcus pneumoniae-induced oxidative stress in
lung epithelial cells depends on pneumococcal autolysis and is
reversible by resveratrol. J Infect Dis. 211:1822–1830. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Adamou JE, Wizemann TM, Barren P and
Langermann S: Adherence of Streptococcus pneumoniae to human
bronchial epithelial cells (BEAS-2B). Infect Immun. 66:820–822.
1998.PubMed/NCBI
|
|
63
|
Robson RL, Reed NA and Horvat RT:
Differential activation of inflammatory pathways in A549 type II
pneumocytes by Streptococcus pneumoniae strains with different
adherence properties. BMC Infect Dis. 6:712006. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Mushtaq N, Ezzati M, Hall L, Dickson I,
Kirwan M, Png KM, Mudway IS and Grigg J: Adhesion of Streptococcus
pneumoniae to human airway epithelial cells exposed to urban
particulate matter. J Allergy Clin Immunol. 127:1236–1242. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Li-Korotky HS, Lo CY, Zeng FR, Lo D and
Banks JM: Interaction of phase variation, host and pressure/gas
composition: Pneumococcal gene expression of PsaA, SpxB, Ply and
LytA in simulated middle ear environments. Int J Pediatr
Otorhinolaryngol. 73:1417–1422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Statt S, Ruan JW, Huang CT, Wu R and Kao
CY: Lipidome and transcriptome profiling of pneumolysin
intoxication identifies networks involved in statin-conferred
protection of airway epithelial cells. Sci Rep. 5:106242015.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li P, Shi J, He Q, Hu Q, Wang YY, Zhang
LJ, Chan WT and Chen WX: Streptococcus pneumoniae induces autophagy
through the inhibition of the PI3K-I/Akt/mTOR pathway and ROS
hypergeneration in A549 cells. PloS One. 10:e01227532015.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zahlten J, Herta T, Kabus C, Steinfeldt M,
Kershaw O, García P, Hocke AC, Gruber AD, Hübner RH, Steinicke R,
et al: Role of pneumococcal autolysin for KLF4 expression and
chemokine secretion in lung epithelium. Am J Respir Cell Mol Biol.
53:544–554. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Leiberman A, Dagan R, Leibovitz E,
Yagupsky P and Fliss DM: The bacteriology of the nasopharynx in
childhood. Int J Pediatr Otorhinolaryngol. 49:(Suppl). S151–S153.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Siegel SJ and Weiser JN: Mechanisms of
bacterial colonization of the respiratory tract. Annu Rev
Microbiol. 69:425–444. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Schmeck B, Zahlten J, Moog K, van Laak V,
Huber S, Hocke AC, Opitz B, Hoffmann E, Kracht M, Zerrahn J, et al:
Streptococcus pneumoniae-induced p38 MAPK-dependent phosphorylation
of RelA at the interleukin-8 promotor. J Biol Chem.
279:53241–53247. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ratner AJ, Lysenko ES, Paul MN and Weiser
JN: Synergistic proinflammatory responses induced by polymicrobial
colonization of epithelial surfaces. Proc Natl Acad Sci USA.
102:3429–3434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Opitz B, Püschel A, Schmeck B, Hocke AC,
Rosseau S, Hammerschmidt S, Schumann RR, Suttorp N and Hippenstiel
S: Nucleotide-binding oligomerization domain proteins are innate
immune receptors for internalized Streptococcus pneumoniae. J Biol
Chem. 279:36426–36432. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Iovino F, Brouwer MC, van de Beek D,
Molema G and Bijlsma JJ: Signalling or binding: The role of the
platelet-activating factor receptor in invasive pneumococcal
disease. Cell Microbiol. 15:870–881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhang JR, Mostov KE, Lamm ME, Nanno M,
Shimida S, Ohwaki M and Tuomanen E: The polymeric immunoglobulin
receptor translocates pneumococci across human nasopharyngeal
epithelial cells. Cell. 102:827–837. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Elm C, Rohde M, Vaerman JP, Chhatwal GS
and Hammerschmidt S: Characterization of the interaction of the
pneumococcal surface protein SpsA with the human polymeric
immunoglobulin receptor (hpIgR). Indian J Med Res. 119:(Suppl).
61–65. 2004.PubMed/NCBI
|
|
77
|
Stenfors LE and Räisänen S: Bacterial
adhesion to epithelial cells of the nasopharynx essential in the
development of otitis media. Nord Med. 107:278–279. 1992.(In
Swedish). PubMed/NCBI
|
|
78
|
Stenfors LE and Räisänen S: In vivo
attachment of Streptococcus pneumoniae and Haemophilus influenzae
to nasopharyngeal epithelium in children. ORL J Otorhinolaryngol
Relat Spec. 54:25–28. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Nelson AL, Roche AM, Gould JM, Chim K,
Ratner AJ and Weiser JN: Capsule enhances pneumococcal colonization
by limiting mucus-mediated clearance. Infect Immun. 75:83–90. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Mbaki N, Rikitomi N, Akiyama M and
Matsumoto K: In vitro adherence of Streptococcus pneumoniae to
oropharyngeal cells: Enhanced activity and colonization of the
upper respiratory tract in patients with recurrent respiratory
infections. Tohoku J Exp Med. 157:345–354. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
McCullers JA and Tuomanen EI: Molecular
pathogenesis of pneumococcal pneumonia. Front Biosci. 6:D877–D889.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Schembri MA, Dalsgaard D and Klemm P:
Capsule shields the function of short bacterial adhesins. J
Bacteriol. 186:1249–1257. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ring A, Weiser JN and Tuomanen EI:
Pneumococcal trafficking across the blood-brain barrier. Molecular
analysis of a novel bidirectional pathway. J Clin Invest.
102:347–360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Talbot UM, Paton AW and Paton JC: Uptake
of Streptococcus pneumoniae by respiratory epithelial cells. Infect
Immun. 64:3772–3773. 1996.PubMed/NCBI
|
|
85
|
Cundell DR, Weiser JN, Shen J, Young A and
Tuomanen EI: Relationship between colonial morphology and adherence
of Streptococcus pneumoniae. Infect Immun. 63:757–761.
1995.PubMed/NCBI
|
|
86
|
Weiser JN, Austrian R, Sreenivasan PK and
Masure HR: Phase variation in pneumococcal opacity: Relationship
between colonial morphology and nasopharyngeal colonization. Infect
Immun. 62:2582–2592. 1994.PubMed/NCBI
|
|
87
|
Kim JO and Weiser JN: Association of
intrastrain phase variation in quantity of capsular polysaccharide
and teichoic acid with the virulence of Streptococcus pneumoniae. J
Infect Dis. 177:368–377. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
88
|
Schaffner TO, Hinds J, Gould KA, Wüthrich
D, Bruggmann R, Küffer M, Mühlemann K, Hilty M and Hathaway LJ: A
point mutation in cpsE renders Streptococcus pneumoniae
nonencapsulated and enhances its growth, adherence and competence.
BMC Microbiol. 14:2102014. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Okumura CY and Nizet V: Subterfuge and
sabotage: Evasion of host innate defenses by invasive gram-positive
bacterial pathogens. Annu Rev Microbiol. 68:439–458. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Reed JM, O'Callaghan RJ, Girgis DO,
McCormick CC, Caballero AR and Marquart ME: Ocular virulence of
capsule-deficient Streptococcus pneumoniae in a rabbit keratitis
model. Invest Ophthalmol Vis Sci. 46:604–608. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Crum NF, Barrozo CP, Chapman FA, Ryan MA
and Russell KL: An outbreak of conjunctivitis due to a novel
unencapsulated Streptococcus pneumoniae among military trainees.
Clin Infect Dis. 39:1148–1154. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
92
|
Dixit C, Keller LE, Bradshaw JL, Robinson
DA, Swiatlo E and McDaniel LS: Nonencapsulated Streptococcus
pneumoniae as a cause of chronic adenoiditis. IDCases. 4:56–58.
2016. View Article : Google Scholar : PubMed/NCBI
|














