Introduction
Bone marrow mesenchymal stem cells (BMSCs) serve as
a primary source of adult stem cells for tissue maintenance and
regeneration. Bone marrow is the largest reservoir of mesenchymal
stem cells (MSCs). A previous study has indicated that BMSCs
differentiate into hepatocyte-like cells (HLCs) in vitro and
in vivo (1). Liver
cirrhosis occurs as a result of the progression of liver fibrosis,
when the initial injury is not resolved. Liver fibrosis is a
consequence of chronic liver disease characterized by the
replacement of liver tissue with an accumulation of collagen and
extracellular matrix proteins, scar tissue and regenerative
nodules, resulting in dysfunction of the liver. The key to the
reversal of liver cirrhosis is the degradation of connective tissue
with abnormal hyperplasia, and the regeneration of liver
parenchymal cells. In addition, numerous cytokines are important in
the development and reversal of liver cirrhosis (2). A previous study has demonstrated that
traditional medicines with comprehensive pharmacological effects
are beneficial for the treatment of liver cirrhosis (3). Yi Guan Jian decoction (YGD) has been
used to treat liver fibrosis and cirrhosis (4). YGD contains Radix Rehmanniae
(18 g), Radix Glehniae (9 g), Radix Angelicae
Sinensis (9 g), Fructus Lycii (9 g), Radix
Ophiopogonis (9 g) and Fructus toosendan (5 g). YGD is a
traditional Chinese medicine and has been used to treat hepatic
diseases in China for centuries. Our previous study demonstrated
that YGD promotes the expression of hepatic oval cells (HOCs) and
the differentiation of HOCs into HLCs (5). In the present study,
dimethylnitrosamine (DMN) was used to induce liver cirrhosis in
mice, and the alterations of liver function and pathology were
observed in untreated and YGD-treated groups. In addition, the
molecular mechanism underlying the differentiation of mouse BMSCs
into HLCs following YGD treatment was identified.
Materials and methods
Animals
A total of 192 male and female Kunming mice (age,
4–6 weeks; weight, 16–22 g) were purchased from the Laboratory
Animal Center of Dalian Medical University (Dalian, China). The
mice were acclimatized to the facility for a week prior to
experimentation and maintained under standard conditions
(temperature, 20–25°C; humidity, 60–80%; 12-h light/dark cycle)
with ad libitum water and food. All mice received humane
care in accordance with the institutional guidelines. The present
study was approved by the Medical Ethics Committee of The First
Affiliated Hospital of Dalian Medical University (Dalian, China;
approval no. KY2013-49).
Therapeutic agents
YGD includes Radix Rehmanniae (18 g),
Radix Glehniae (9 g), Radix Angelicae Sinensis (9 g),
Fructus Lycii (9 g), Radix Ophiopogonis (9 g) and
Fructus toosendan (5 g). The formula was decocted in
distilled water and prepared in the Pharmacy Department of Dalian
Medical University. Hepatocyte growth factor (HGF) was provided by
Weihai Sinogen Pharmaceutical Co., Ltd. (Weihai, China) and
dissolved in normal saline.
Reagents
DMN was purchased from Tokyo Kasei Kogyo Co., Ltd.
(Tokyo, Japan). The primary rabbit anti-mouse polyclonal antibodies
against α-smooth muscle actin (α-SMA; catalog no. ab66133), C-X-C
chemokine receptor type 4 (CXCR4; catalog no. ab2074),
extracellular signal-regulated kinase (ERK1/2; catalog no. ab17942)
and nuclear factor κB (NF-κB) p65 subunit (NF-κBp65; catalog no.
ab16502), a rabbit anti-mouse monoclonal antibody against β-catenin
(catalog no. ab32572), a rat anti-mouse monoclonal antibody against
cluster of differentiation (CD) 90 (catalog no. ab3105), rabbit
anti-rat polyclonal antibodies against albumin (Alb; catalog no.
ab135575) and cytokeratin (CK) 18 (catalog no. ab189444), and the
secondary antibodies goat anti-rat IgG H&L (Alexa
Fluor® 488; catalog no. ab150157) and goat anti-rabbit
IgG H&L (Alexa Fluor® 647; catalog no. ab150079),
were purchased from Abcam (Cambridge, MA, USA). The primary
antibody against β-actin (catalog no. bs-0061R) was obtained from
BIOSS (Beijing, China). The secondary antibody
peroxidase-conjugated AffiniPure goat anti-rabbit IgG (catalog no.
ZB-2301) was obtained from ZSGB-BIO (Beijing, China). The primers,
Takara Mini BEST Universal RNA Extraction kit (catalog no. 9767),
PrimeScript™ RT Master mix (catalog no. RR036A) and
SYBR® Premix Ex Taq™ (catalog no. RR420A) were obtained
from Takara Bio, Inc. (Otsu, Japan).
Liver cirrhosis model
A liver cirrhosis model was established as
previously described by Jenkins et al (6). Animals were randomly divided into two
groups. The model group (n=172) received 10 mg/kg DMN
intraperitoneally once per day for three consecutive days within a
week for a total of four weeks. The control group (n=20) mice were
injected with an identical volume of normal saline on the same
regimen. To monitor the development of liver cirrhosis, five mice
from the control group and ten mice from the model group were
randomly selected and sacrificed by cervical dislocation, following
anesthesia with an intraperitoneal injection of 10% chloral hydrate
(Qingdao Yulong Algae Co., Ltd., Qingdao, China), at weeks 1, 2, 3
and 4 following the first injection of DMN.
From the fifth week following the first injection of
DMN, DMN administration was ceased, and mice from the model group
were further divided into the following three subgroups
(n=44/group): i) Negative control (NC), ii) positive control (HGF)
and iii) YGD. YGD (10 ml/kg body weight) was administered by oral
gavage once daily for four weeks. HGF (200 µg/kg body weight) was
administered once daily by subcutaneous injection for four weeks.
Mice in the NC group received normal saline on the same regimen. A
total of 10 mice from each group were randomly selected and
sacrificed at weeks 5, 6, 7 and 8 following the first injection of
DMN.
Following anesthesia with an intraperitoneal
injection of 10% chloral hydrate, whole blood was collected from
mice and the serum was isolated and stored at −80°C for the liver
function analysis. The liver was then dissected. A portion of the
tissue was partially fixed with formalin and embedded in paraffin
for hematoxylin and eosin (HE) staining, Masson's trichrome (MT)
staining and immunohistochemistry (IHC). A further portion of the
liver tissue was embedded in optimum cutting temperature compound
for immunofluorescence, and the remaining tissue was snap-frozen in
liquid nitrogen and stored at −80°C for western blotting and
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR).
Monitoring of mice
Analyses of the overall health of the mice were
undertaken, including alterations in weight, feeding, drinking,
activity and hair loss.
Measurement of serum liver
function
The serum levels of alanine aminotransferase (ALT),
aspartate aminotransferase (AST), Alb, total bilirubin (TBil),
direct bilirubin (DBil), cholinesterase (CHE), alkaline phosphatase
(ALP) and gamma-glutamyltranspeptidase (GGT) were determined using
an automatic biochemical analyzer.
Pathology
Macroscopic alterations in the shape, size, color
and texture of livers were observed. Paraffin sections were cut
into sections (6-µm thick), deparaffinized and rehydrated for HE
and MT staining, to enable observation of the alterations to liver
cell degeneration, necrosis and the degree of hepatic fibrosis
under a light microscope. For HE staining, sections were stained
with hematoxylin for 1 min and washed in tap water, followed by
staining with eosin for 1–2 min and further washing in tap water.
Sections were subsequently dehydrated, cleared in xylene and
mounted. For MT staining, sections were stained with ponceau acid
fuchsin solution for 5 min, briefly differentiated with 0.2% acetic
acid aqueous solution, rinsed with distilled water, incubated with
5% phosphomolybdic acid solution for 8 min and directly immersed
into aniline blue dye for 5 min. Sections were incubated with 0.2%
acetic acid aqueous solution for 2 min and washed in tap water.
Sections were subsequently dehydrated, cleared in xylene and
mounted.
IHC
Liver sections were deparaffinized and rehydrated,
and 5% bovine serum albumin (BSA; Solarbio, Beijing, China) was
used to block non-specific protein binding at room temperature for
2 h. Sections were incubated overnight at 4°C with the following
primary antibodies: Anti-α-SMA (1:200), anti-CXCR4 (1:50),
anti-ERK1/2 (1:200), anti-NF-κBp65 (1:500) or anti-β-catenin
(1:500), or 5% BSA in PBS without antibody as a negative control.
The sections were washed and incubated with a peroxidase-conjugated
AffiniPure goat anti-rabbit IgG secondary antibody (1:1,000) for 30
min at 37°C. Color was developed with diaminobenzidine
tetrahydrochloride (ZSGB-BIO). Slides were counterstained with
hematoxylin, mounted and visualized under a light microscope.
Immunofluorescence
Following blocking as aforementioned, slides were
incubated with anti-CD90 (1:300) and anti-Alb (1:100) or anti-CK18
(1:100) overnight at 4°C. Goat anti-rat IgG-AlexaFluor488 (1:200)
and goat anti-rabbit IgG-AlexaFluor647 (1:200) secondary antibodies
were added and incubated in the dark for 1 h at room temperature.
Images were captured by confocal laser scanning microscopy (Leica
Microsystems, GmbH, Wetzlar, Germany). Identical exposure times and
light intensities were applied to all images. ImageJ software was
used (version, JAVA 1.6.0–24 for Windows; National Institutes of
Health, Bethesda, MD, USA).
Western blotting
The liver tissues were homogenized in
radioimmunoprecipitation assay buffer containing a protease
inhibitor cocktail (Beyotime Institute of Biotechnology, Haimen,
China). Total protein (25 µg) was loaded into each well, subjected
to 10% SDS-PAGE and transferred onto polyvinylidene difluoride
membranes. Following this, membranes were blocked in 5% non-fat
milk in PBS containing Tween 20 at 37°C for 1 h, and subsequently
incubated with antibodies against α-SMA (1:500), CXCR4 (1:1,000),
ERK1/2 (1:1,000), NF-κBp65 (1:1,000), β-catenin (1:5,000) or
β-actin (1:1,500), which served as an internal standard, overnight
at 4°C. Membranes were incubated with a peroxidase-conjugated
AffiniPure goat anti-rabbit IgG secondary antibody (dilution,
1:1,000) at 37°C for 1 h. Proteins were visualized with a
Diaminobenzidine Horseradish Peroxidase Color Development kit
(Beyotime Institute of Biotechnology) and quantified by
densitometry using the AlphaView Stand Alone Analysis software
(ProteinSimple, version 3.4.0.0; San Jose, CA, USA).
RT-qPCR
Total RNA was purified from liver tissues using a
Takara MiniBEST Universal RNA Extraction kit, and subjected to RT
using PrimeScript RT Master mix. qPCR was performed using SYBR
Premix Ex Taq and aStepOnePlus™ Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
GAPDH served as an internal control. Sequences of primers were as
follows: Forward, 5′-GACAATGGCTCTGGGCTCTGTA-3′ and reverse,
5′-TTTGGCCCATTCCAACCATTA-3′ for α-SMA; forward,
5′-CCATGGAACCGATCAGTGTG-3′ and reverse, 5′-GCCGACTATGCCAGTCAAGAA-3′
for CXCR4; forward, 5′-CACACGTTGGTACAGAGCTCCAG-3′ and reverse,
5′-TGCAGCCCACAGACCAAATATC-3′ for ERK1; forward,
5′-CTGGACCAGCTCAACCACATTC-3′ and reverse,
5′-AGGTAGTTTCGGGCCTTCATGTTA-3′ for ERK2; forward, 5′-
AGCACAGATACCACCAAGACACA-3′ and reverse,
5′-CAGGTCTCGCTTCTTCACACAC-3′ for NF-κBp65; forward,
5′-GCCACAGGATTACAAGAAGC-3′ and reverse, 5′-CCACCAGAGTGAAAAGAACG-3′
for β-catenin; and forward, 5′-AAATGGTGAAGGTCGGTGTGAAC-3′ and
reverse, 5′-CAACAATCTCCACTTTGCCACTG-3′ for GAPDH. The details of
the thermocycling conditions were as follows: 95°C for 30 sec
(initial denaturation), followed by 40 cycles at 95°C for 5 sec
(exact denaturation) and 60°C for 30 sec (primer annealing and PCR
product elongation). The relative quantity of target genes mRNA was
normalized to that of GAPDH using the 2-ΔΔCq method, as
previously described (7).
Statistical analysis
Statistical analysis was performed using SPSS
software version 19.0 (IBM SPSS, Armonk, NY, USA) and data are
presented as the mean ± standard deviation. Data were analyzed
using Shapiro-Wilk (normality) and Levene's (homogeneity) tests.
Differences between the control and treated groups were analyzed
using one-way analysis of variance, followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Physical state of mice
Immediately following administration of DMN, the
model group exhibited anxiety and were irritable, gradually
becoming inactive and calm over a period of 5 h. Behavioral
improvement was observed 4 days later, and returned to normal 7
days later, except with hair loss and a small quantity of ascites.
There were no significant differences in eating, drinking and
weight between groups of mice at any time point. The symptoms
improved following treatment with YGD or HGF.
Liver function decreases in the model
group
ALT, AST, TBil, DBil and ALP levels at 1–4 weeks and
GGT levels at 1–3 weeks increased (P<0.05), and Alb levels at
1–4 weeks and CHE levels at 2–4 weeks decreased (P<0.05), in the
model compared with the control group. These results revealed that
liver function gradually decreased in the model group (Table I).
 | Table I.Comparison of liver function in the
control and model groups. |
Table I.
Comparison of liver function in the
control and model groups.
| Group | n | ALT (IU/l) | AST (IU/l) | Alb (g/l) | TBIL (umol/l) | DBIL (umol/l) | CHE (U/l) | ALP (IU/l) | GGT (IU/l) |
|---|
| Control | 20 | 33.67±2.16 | 132.83±2.85 | 37.38±1.11 | 0.95±0.19 | 0.3±0.14 | 56.33±5.32 | 87.13±0.61 | 1.00±0.00 |
| Model week 1 | 10 |
61.40±5.32a |
148.8±3.19a |
34.82±0.26a |
1.88±0.38a |
0.6±0.16a | 50.88±0.40 |
114.32±3.06a |
1.15±0.04a |
| Model week 2 | 10 |
80.29±37.63a |
164.00±12.32a,b |
32.27±2.39a,b |
2.97±1.40a |
0.83±0.42a |
45.79±0.99a,b |
138.00±7.23a,b |
1.31±0.18a |
| Model week 3 | 10 |
104.20±4.66a |
318.00±27.06a,c |
27.12±0.29a,c |
3.20±0.26a |
1.16±0.43a |
38.74±0.32a,c |
164.72±3.15a,c |
1.61±0.04a,c |
| Model week 4 | 10 |
138.25±9.32a,d |
484.75±89.46a,d |
20.60±0.32a,d |
3.83±0.49a,d |
1.65±0.21a |
35.75±6.40a |
183.75±2.50a,d | 1.80±0.59 |
Serum liver function improves in the
HGF and YGD groups
Following HGF treatment, ALT and AST levels at 1–4
weeks decreased (P<0.05), Alb levels at 1–4 weeks and CHE levels
at 3–4 weeks increased (P<0.05), DBil levels decreased at 4
weeks (P<0.05), as did TBil levels at 3–4 weeks (P<0.05) and
ALP levels at 1, 3 and 4 weeks (P<0.05), compared with the model
group. Therefore, improvement of liver function was observed in the
HGF group. Compared with the NC group at the same time point, the
levels of ALT, AST and GGT at 1–4 weeks decreased (P<0.05), Alb
levels increased at 1–4 weeks (P<0.05), TBil and ALP levels
decreased at week 4 (P<0.05), DBil levels decreased at weeks 1
and 4 (P<0.05), and CHE levels increased at weeks 1,3 and 4
(P<0.05; Table II).
| Table II.Comparison of liver function in the
NC, YGD and HGF groups. |
Table II.
Comparison of liver function in the
NC, YGD and HGF groups.
| Group | n | ALT (IU/l) | AST (IU/l) | Alb (g/l) | TBil (umol/l) | DBil (umol/l) | CHE (U/l) | ALP (IU/l) | GGT (IU/l) |
|---|
| NC week 1 | 10 | 143.00±2.16 | 460.25±6.40 |
21.38±0.25a | 3.55±0.26 | 2.25±0.13 | 35.35±1.08 | 183.50±2.08 | 1.38±0.07 |
| NC week 2 | 10 | 142.20±1.92 |
406.80±10.96b |
22.00±0.29a,b | 3.36±0.23 |
1.78±0.28& |
40.00±2.45b | 184.40±3.58 |
1.29±0.10b |
| NC week 3 | 10 | 141.20±10.96 |
378.60±2.07a,c |
27.04±0.48a,c | 3.42±0.19 |
1.38±0.19c | 39.32±0.47 | 174.80±67.94 | 1.29±0.06 |
| NC week 4 | 14 | 137.80±3.56 |
360.40±9.81a,d |
29.08±0.37a,d | 3.42±0.26 | 1.48±0.24 | 40.46±1.55 | 175.60±17.95 |
1.08±0.06a,d |
| HGF week 1 | 10 |
80.33±28.71a,k |
196.50±3.62a,k |
29.33±4.46a,k | 3.53±0.22 |
1.65±0.19k |
42.03±0.57k |
159.17±23.34a |
1.47±0.02k |
| HGF week 2 | 10 |
57.00±4.69a,1 |
148.60±2.07a,e,l |
29.64±0.94a,l | 3.50±0.16 | 1.00±1.47 | 45.40±10.88 | 147.20±96.83 |
1.22±0.03e,l |
| HGF week 3 | 10 |
40.80±3.27a,f,m |
138.20±2.59a,f,m |
33.76±0.80a,f,m |
3.14±0.23a,f | 0.72±0.19 |
60.26±0.58a,f,m |
130.00±27.51a |
1.20±0.03m |
| HGF week 4 | 14 |
39.60±7.02a,n |
116.80±4.66a,g,n |
35.58±0.97a,g,n |
3.00±0.16a,n |
0.08±0.64a,n |
67.00±7.07a,n |
69.32±1.19a,g,n |
1.19±0.02n |
| YGD week 1 | 10 |
70.60±20.91a,k |
184.60±93.06a,k |
25.78±0.51a,k | 3.60±0.16 |
1.40±0.43k |
46.60±0.92a,k,o |
163.52±2.92a,k | 1.47±0.18 |
| YGD week 2 | 10 |
51.02±13.35a,l |
163.40±4.51a,l,p |
32.04±4.15a,h,l | 3.50±0.29 |
1.30±0.16a |
51.00±4.30a,l |
148.80±3.35a,h,l |
1.46±0.02l,p |
| YGD week 3 | 10 |
49.80±1.92a,m,q |
122.00±2.55a,i,m,q |
33.30±1.40a,m | 3.10±0.50 |
0.82±0.19a,i,m |
56.20±1.48a,i,m,q | 128.40±54.29 |
1.30±0.05i,q |
| YGD week 4 | 14 |
35.00±3.16a,j,n |
107.20±4.66a,j,n,r |
36.36±2.00a,j,n |
3.04±0.38a |
0.5±0.16a,j,n |
73.96±2.42a,j,n |
120.00±2.92a,n,r |
1.06±0.10a,j,r |
Following YGD treatment, ALT and AST levels at 1–4
weeks decreased (P<0.05), Alb and CHE levels at 1–4 weeks
increased (P<0.05), TBil and GGT levels decreased at 4 weeks
(P<0.05), as did DBil levels at 2–4 weeks (P<0.05) and ALP
levels at 1, 2 and 4 weeks (P<0.05), compared with the model
group at 4 weeks. Compared with the NC group at the same time
point, the levels of ALT and AST at 1–4 weeks decreased
(P<0.05), Alb and CHE levels increased at 1–4 weeks (P<0.05),
and the levels of TBil at week 4, DBil at weeks 1, 3 and 4, ALP at
weeks 1, 2 and 4, and GGT at week 1, increased (P<0.05). The
results suggested that liver function improved gradually in the YGD
group (Table II).
Additionally, CHE levels were increased (P<0.05)
following YGD treatment compared with the HGF group at week 1. AST
and GGT levels were decreased (P<0.05) in the YGD group compared
with the HGF group at week 2. ALT, AST and GGT levels decreased
(P<0.05), whereas CHE levels increased, in the YGD group
compared with the HGF group at week 3. AST, ALP and GGT levels
decreased (P<0.05) in the YGD group compared with the HGF group
at 4 weeks. There were no significant differences (P>0.05)
between the two groups except for those described above (Table II).
Liver tissue alterations
The control group liver presented a reddish brown
color, with a smooth and soft surface. In the model group, the
liver tissues were markedly swollen and green/yellow, with part of
the liver surface nodular, hard and with a blunt edge. In the YGD
group, the liver swelling decreased and the liver surface was
yellow/brown and smooth. In the HGF group, the liver tissue
appeared similar to the YGD group; however, alterations in this
group were less marked compared with the YGD group.
Liver HE staining
Model group liver cells at 1 week were markedly
swollen and degenerated, exhibiting a visible infiltration of
inflammatory cells in the necrotic area. At weeks 2 and 3,
inflammatory cell infiltration had increased, with destruction of
hepatic lobules and portal area structure, and disordered
arrangement of liver cells. At 4 weeks liver cells exhibited
increased degeneration, steatosis around portal areas and centric
veins, with collagenous fibers in the portal area.
In the YGD group at weeks 1–3, the degeneration
necrosis of liver cells gradually reduced, with disordered
arrangement of the regenerating hepatic cord, dual-core liver cells
were visible under a high power lens and steatosis around portal
areas and centric veins gradually reduced. At week 4, the liver
cells almost resembled those in the control group. Similar hepatic
pathology and ultrastructure alterations were observed in the HGF
group. In the NC group at weeks 1–4, the degeneration necrosis,
steatosis around portal areas and centric veins, with collagenous
fibers in the portal area of liver cells were continued. A small
quantity of liver cell regeneration could be seen at week 4
(Fig. 1A).
Liver MT staining
In the model group, fibrous septa formed in the
portal areas of livers at week 1, extended and subsequently spread
to the liver parenchyma. The fibrous septa thickened at weeks 2 and
3 and extended to the intralobular area surrounding the liver
cells, with the central vein wall thickened. The hyperplasia of
fibrous tissue and pseudolobule proliferation occurred in mice
liver tissues at week 4, in which atresia, absence, bias and double
central vein structure were visible. These results suggested that
the liver cirrhosis model had been successfully established.
In the YGD group the hyperplasia of fibrous tissue
and fibrous septa gradually reduced at weeks 1–3. The collagen
fiber levels resembled those in the control group by week 4, and a
small quantity of collagen fibers were viewed only near the portal
area and central vein. In the HGF group, there was a decrease in
fibrous septa and collagen fiber levels. In the NC group at weeks
1–4, pseudolobules, atresia, absence, bias and double central vein
structure remained visible (Fig.
1B).
Co-localization of CD90 with Alb or
CK18
To monitor BMSCs and their potential to
differentiate into hepatocytes as well as biliary epithelial cells
during the development and reversal of liver cirrhosis, double
labeling immunofluorescence was performed in liver tissue from the
model and YGD groups. The expression of the BMSC marker CD90 with
the hepatocyte marker Alb or the biliary epithelial marker CK18 was
investigated. In the model group, there was minimal co-localization
between CD90 and Alb or CK18 at weeks 1 and 2, while at weeks 3 and
4 the co-localization gradually increased. The co-localization of
CD90 with Alb or CK18 increased markedly following YGD treatment,
and reached a peak at week 4 (Fig.
2).
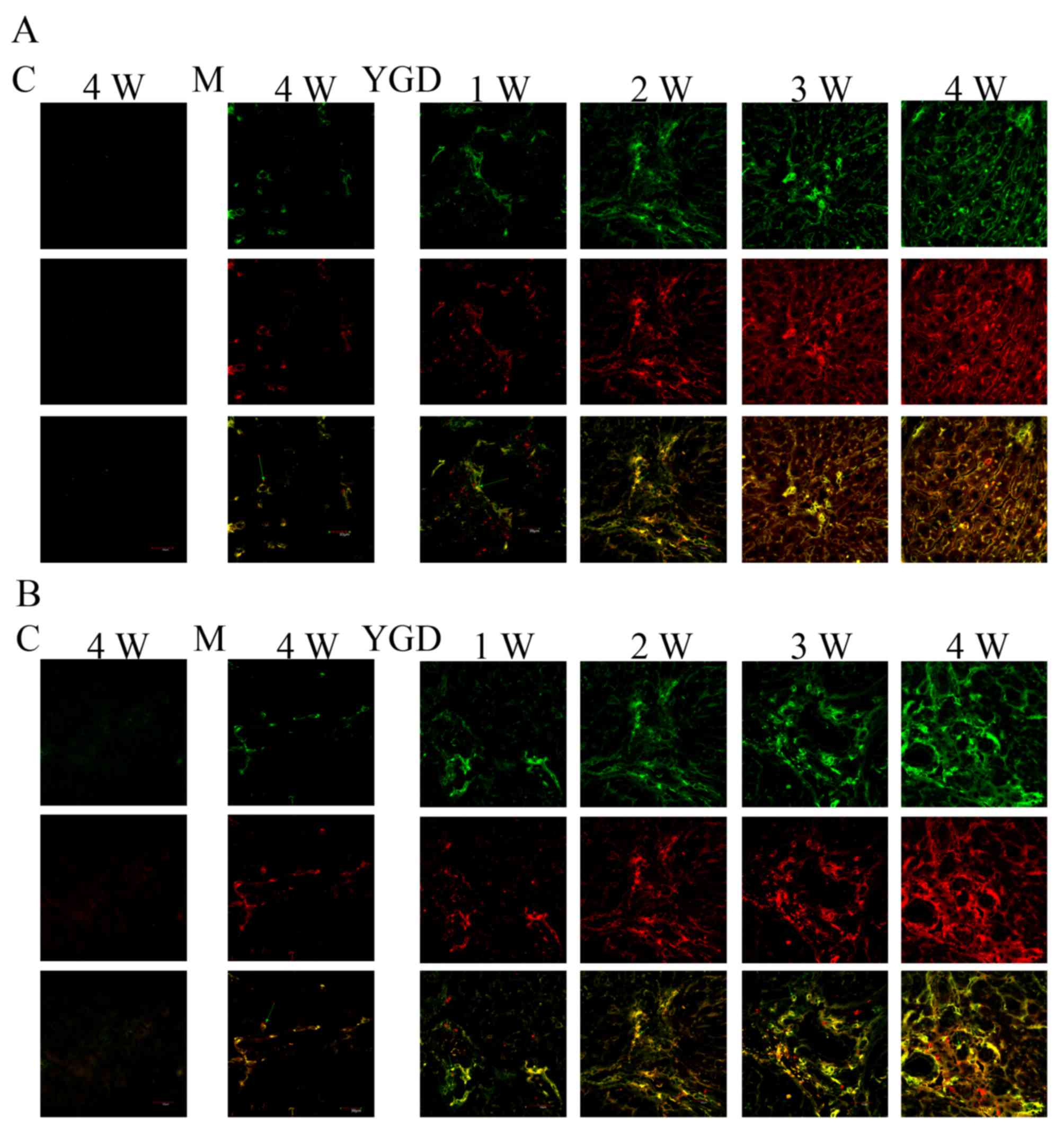 | Figure 2.DMN and YGD regulate the
differentiation of BMSCs into hepatocytes and biliary epithelial
cells. Mice were injected with DMN or normal saline for 4 weeks.
Following 4 weeks of DMN treatment, mice were treated with YGD or
HGF for a further 4 weeks. (A) The expression of CD90 (green, top
row) and Alb (red, middle row) was detected by immunofluorescence,
with the co-localization analyzed by merging these images (bottom
row). (B) The expression of CD90 (green, top row) and CK18 (red,
middle row) was detected by immunofluorescence, with the
co-localization analyzed by merging these images (bottom row).
Expression and co-localization was increased by DMN injection and
further increased by YGD treatment. Magnification, ×60. YGD, Yi
Guan Jian decoction; HGF, hepatocyte growth factor; DMN,
dimethylnitrosamine; CD, cluster of differentiation; Alb, Albumin;
CK18, cytokeratin 18; C, control; M, model; W, week. |
Protein expression of α-SMA, CXCR4,
ERK1/2, NF-κB p65 and β-catenin
IHC on the control group revealed no or very few
positively stained cells. In the model group, α-SMA, CXCR4, ERK1/2,
NF-κB p65 and β-catenin staining markedly increased and was
primarily concentrated in the portal area, near the fibrous septa
and hepatic cord. Following YGD or HGF administration, CXCR4 and
ERK1/2 staining gradually increased compared with the NC group at
the same point and the model group at week 4, peaking at 4 weeks in
the YGD and HGF groups. α-SMA, NF-κB p65 and β-catenin levels
gradually decreased the NC group at the same point and compared
with the model group at week 4; by week 4 in the YGD and HGF
groups, the expression was almost at the level of the control group
(Fig. 3). As the model group week
4 demonstrated more severe damage, according to measurements of
serum liver function, pathology and IHC, compared with the NC group
at any time point, it was selected for comparison with the YGD and
HDF groups in subsequent experiments.
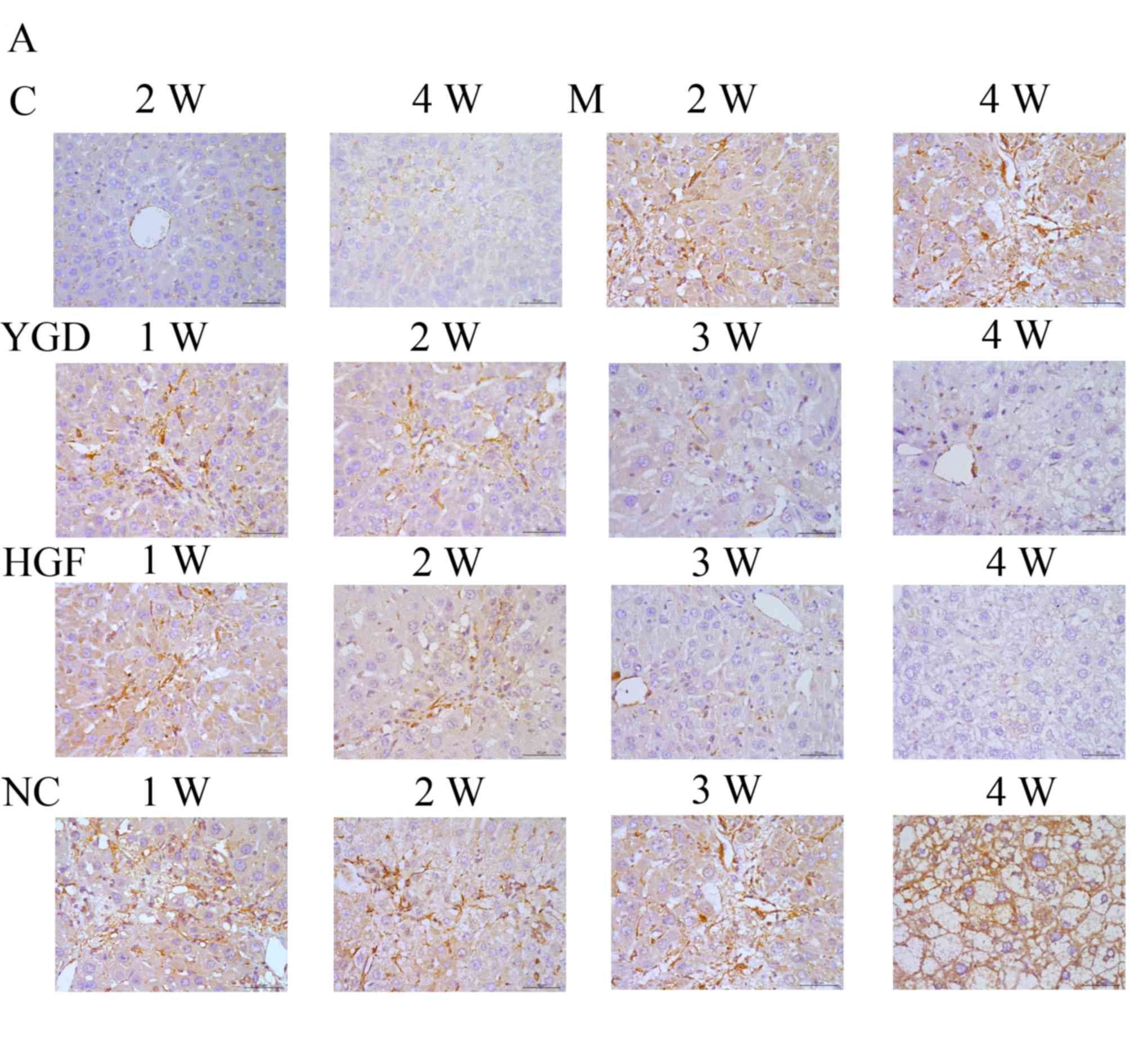 | Figure 3.DMN, YGD and HGF regulate the
expression of α-SMA, CXCR4, ERK1/2, NF-κB p65 and β-catenin. Mice
were injected with DMN or normal saline for 4 weeks. Following 4
weeks of DMN treatment, mice were treated with YGD or HGF for a
further 4 weeks. Liver sections were stained for (A) α-SMA (B)
CXCR4. (C) ERK1/2 (D) NF-κB p65. (E) β-catenin. A representative
image from each group is presented. Very little staining was
observed in the control group. YGD or HGF treatment increased CXCR4
and ERK1/2 staining, and decreased α-SMA, NF-κB p65 and β-catenin
staining, compared with the DMN-injected group. Magnification,
×400; scale bar=50 µm.YGD, Yi Guan Jian decoction; HGF, hepatocyte
growth factor; DMN, dimethylnitrosamine; α-SMA, α-smooth muscle
actin; CXCR4, C-X-C chemokine receptor type 4; ERK1/2,
extracellular signal-regulated kinase 1/2; NF-κB p65, nuclear
factor κB p65 subunit; C, control; M, model; NC, negative control;
W, week. |
Western blotting revealed that the protein
expression levels of α-SMA, ERK1/2, NF-κB p65 and β-catenin in the
model group at 4 weeks were increased compared with the control
group (P<0.05). Following YGD treatment, the CXCR4 and ERK1/2
protein expression levels significantly increased (P<0.05)
compared with the model group at week 4, with a peak at 4 weeks.
α-SMA, NF-κB p65 and β-catenin protein expression levels
significantly decreased following YGD treatment in a time-dependent
manner (P<0.05) compared with the model group at week 4, almost
reaching control group levels (P<0.05; Fig. 4A and B; Table III).
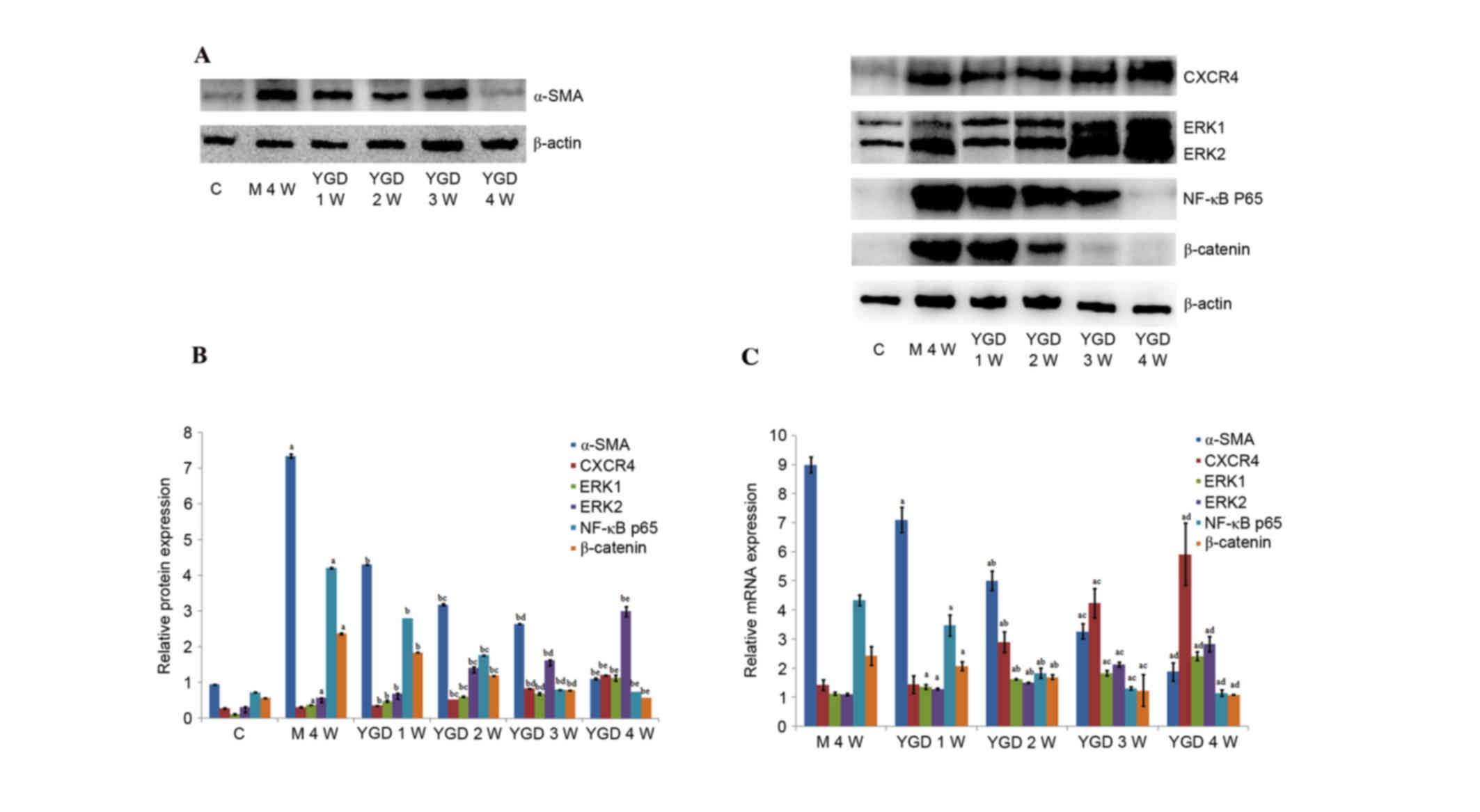 | Figure 4.DMN and YGD regulate mRNA and protein
expression levels of α-SMA, CXCR4, ERK1/2, NF-κB p65 and β-catenin.
Mice were injected with DMN or normal saline for 4 weeks. Following
4 weeks of DMN treatment, mice were treated with YGD or HGF for a
further 4 weeks. (A) The protein expression levels of α-SMA, CXCR4,
ERK1/2, NF-κB p65 and β-catenin were detected by western blotting
and (B) the western blots were quantified. aP<0.05
vs. control; bP<0.05 vs. model week 4;
cP<0.05 vs. YGD week 1; dP<0.05 vs. YGD
week 2; and eP<0.05 vs. YGD week 3. (C) Reverse
transcription-quantitative polymerase chain reaction was performed
to analyze the mRNA expression levels. aP<0.05 vs.
model week 4; bP<0.05 vs. YGD week 1;
cP<0.05 vs. YGD week 2; and dP<0.05 vs.
YGD week 3. YGD, Yi Guan Jian decoction; DMN, dimethylnitrosamine;
α-SMA, α-smooth muscle actin; CXCR4, C-X-C chemokine receptor type
4; ERK1/2, extracellular signal-regulated kinase 1/2; NF-κB p65,
nuclear factor κB p65 subunit; C, control; M, model; W, week. |
| Table III.Comparison
ofrelativeproteinexpression levels in the control, model and YGD
groups. |
Table III.
Comparison
ofrelativeproteinexpression levels in the control, model and YGD
groups.
| Group | n | a-SMA | CXCR4 | ERK1 | ERK2 | NF-κB p65 | β-catenin |
|---|
| Control | 20 | 0.945±0.013 | 0.277±0.019 | 0.115±0.011 | 0.313±0.014 | 0.728±0.003 | 0.567±0.008 |
| Model week 4 | 10 |
7.351±0.051a | 0.312±0.014 |
0.373±0.011a |
0.579±0.002a |
4.211±0.021a |
2.376±0.024a |
| YGD week 1 | 10 |
4.309±0.006a,b |
0.356±0.008a,b |
0.482±0.020a,b |
0.687±0.025a,b |
2.803±0.005a,b |
1.842±0.006a,b |
| YGD week 2 | 10 |
3.188±0.017a,b,c |
0.523±0.002a,b,c |
0.610±0.017a,b,c |
1.413±0.025a,b,c |
1.765±0.005a,b,c |
1.189±0.013a,b,c |
| YGD week 3 | 10 |
2.647±0.010a,b,d |
0.829±0.004a,b,d |
0.685±0.032a,b,d |
1.617±0.020a,b,d |
0.801±0.007a,b,d |
0.784±0.008a,b,d |
| YGD week 4 | 14 |
1.103±0.022a,b,e |
1.205±0.010a,b,e |
1.125±0.080a,b,e |
2.992±0.141a,b,e |
0.742±0.002a,b,e |
0.569±0.001b,e |
mRNA expression levels of α-SMA,
CXCR4, ERK1/2, NF-κB p65 and β-catenin
In the model group at week 4, the mRNA relative
expression levels were as follows: α-SMA, 8.998±0.267; CXCR4,
1.424±0.186; ERK1, 1.133±0.050; ERK2, 1.104±0.046; NF-κB p65,
4.338±0.176l; and β-catenin, 2.433±0.324.
These values were all greater compared with the
control group. Following YGD treatment, the relative mRNA
expression levels of CXCR4 and ERK1/2 gradually increased, with
maximum levels achieved at week 4. The relative mRNA expression
levels of CXCR4 at 2–4 weeks and ERK1/2 at 1–4 weeks all
significantly increased (P<0.05) when compared with the model
group at 4 weeks. Furthermore, among YGD groups, the expression of
CXCR4 and ERK1/2 increased and exhibited a significant difference
(P<0.05). The relative mRNA expression levels of α-SMA, NF-κB
p65 and β-catenin gradually decreased in the YGD groups in a
time-dependent manner, almost reaching those of the control group
at week 4, and revealing a significant decrease (P<0.05)
compared with the model group at 4 weeks (P<0.05; Fig. 4C; Table IV).
| Table IV.Comparison ofrelativemRNA expression
levels in the model and YGD groups. |
Table IV.
Comparison ofrelativemRNA expression
levels in the model and YGD groups.
| Group | n | a-SMA | CXCR4 | ERK1 | ERK2 | NF-κB p65 | β-catenin |
|---|
| Model week 4 | 10 | 8.998±0.267 | 1.424±0.186 | 1.133±0.050 | 1.104±0.046 | 4.338±0.176 | 2.433±0.324 |
| YGD week 1 | 10 |
7.107±0.429a | 1.441±0.302 |
1.369±0.081a |
1.291±0.039a |
3.475±0.369a |
2.080±0.147a |
| YGD week 2 | 10 |
5.016±0.333a,b |
2.903±0.350a,b |
1.620±0.026a,b |
1.505±0.021a,b |
1.837±0.175a,b |
1.699±0.083a,b |
| YGD week 3 | 10 |
3.269±0.267a,c |
4.236±0.510a,c |
1.835±0.090a,c |
2.123±0.082a,c |
1.309±0.053a,c |
1.237±0.539a,c |
| YGD week 4 | 14 |
1.883±0.311a,d |
5.925±1.067a,d |
2.408±0.151a,d |
2.831±0.256a,d |
1.147±0.115a,d |
1.088±0.012a,d |
Discussion
The mutagen DMN results in liver function disorders
and cirrhosis. It is widely used in experimental animals, including
mice, to create a low-mortality and reproducible model of liver
cirrhosis (8,9). DMN significantly induces the
deposition of collagen fibers and the formation of fibrous septa,
as reported by previous studies (10,11),
and as demonstrated by MT staining in the present study. Symptoms
of typical liver cirrhosis appeared in the model group at week 4
and the α-SMA level gradually increased, as demonstrated by IHC,
western blotting and RT-qPCR. This revealed that the degree of
liver fibrosis increased with the development of DMN-induced liver
cirrhosis. In addition, the serum levels of ALT, AST, ALP, GGT,
TBil and DBil significantly increased, whereas the serum Alb and
CHE levels decreased following DMN administration. Furthermore, the
infiltration of inflammatory cells, with destruction of the hepatic
lobule, portal area structure and disordered arrangement of liver
cells as revealed by HE staining suggested the presence of liver
cell damage and defective liver function. This data suggested that
the administration of DMN successfully induced liver cirrhosis in
the model system of the current study.
HGF is a well-known angiogenic, anti-fibrotic and
anti-inflammatory cytokine (12,13).
Certain studies (14–16) have reported that HGF induces MSCs
to differentiate into hepatocytes in vitro and attenuate
liver fibrosis in vivo. The anti-fibrotic effect of HGF may
be achieved via inhibition of the proliferation and activation of
hepatic stellate cells (HSCs), and attenuation of the expression of
the fibrogenic cytokines transforming growth factor β 1 and
platelet-derived growth factor-BB in the liver (17). The effectiveness and broad mode of
action of this cytokine prompted the selection of HGF as a positive
control to investigate the underlying mechanism of YGD in the
reversal of liver cirrhosis. YGD is a traditional Chinese medicine
in which Radix Rehmanniae is the primary active substance,
and has ‘yin-blood-enriching’ effects as well as providing
nourishment to liver and kidney (18). The formula may cure the
‘liver-kidney yin deficiency’ (19) syndrome of chronic hepatitis and
liver cirrhosis. In addition, it may induce the differentiation of
rat BMSCs into HLCs (20).
Following YGD or HGF treatment, the serum levels of ALT, AST, Alb,
CHE, ALP, GGT, TBil and DBil demonstrated a marked improvement
compared with the model group. Liver histology indicated that the
necrotic degeneration of liver cells gradually decreased,
regeneration of the hepatic cord disordered arrangement occurred
and hyperplasia of fibrous tissue and fibrous septa gradually
reduced. Elevation of α-SMA mRNA and protein expression levels,
which is a marker of HSC activation, were additionally inhibited.
This data indicated that YGD achieved a similar effect to HGF on
improvement of liver function and structure, and reversing
DMN-induced liver cirrhosis. This may be mediated via inducing the
differentiation of BMSCs into HLCs and inhibiting the activation of
HSCs to reduce the expression of α-SMA.
BMSCs are multipotent cells capable of
differentiating into multiple lineages that contribute to tissue
repair and regeneration, including adipocytes, osteoblasts,
hepatocytes, nerve cells, chondroblasts and myocardial cells
(21). A previous study has
demonstrated that BMSCs may repair liver function, promote the
regeneration of hepatocytes and reduce liver fibrosis via
inhibiting HSC activation (22).
The present study monitored the differentiation of BMSCs by
analyzing the co-localization of CD90 with the hepatocyte marker
Alb or the biliary epithelial cell marker CK18. In the DMN-induced
cirrhotic liver, the co-expression of the two pairs of markers was
significantly upregulated to a similar level, compared with the
control mice, which indicated a balanced differentiation to the two
epithelial cell subtypes. This differentiation, induced in response
to injury, may be limited, and therefore not sufficient to reverse
the damage induced by DMN. Following YGD treatment, the
co-localization of CD90 with Alb and CK18 was increased, suggesting
that YGD exerted a potent pro-differentiation effect on BMSCs,
which may have reversed the DMN-induced liver cirrhosis.
The underlying mechanism of BMSC differentiation
into hepatocytes and biliary epithelial cells remains to be
elucidated; however, previous studies (23) suggest it is associated with
multiple signaling pathways. The chemokine stromal cell-derived
factor-1 (SDF-1) binds to CXCR4. Numerous studies have reported
that SDF-1 is critical for stem/progenitor and mesenchymal cell
chemotaxis and inflammatory cell homing to injured tissue via
interaction with CXCR4 on the surface of these cells; CXCR4 maybe
highly expressed in humans and mice following chemotaxis along an
SDF-1 gradient (24,25). This indicates that the level of
CXCR4 expression may reflect the activation of the SDF-1/CXCR4
axis, which is associated with stem cells. The mitogen activated
protein kinase (MAPK)/ERK1/2 signaling pathway is one of the
downstream signaling pathways of SDF-1/CXCR4, which maybe activated
by various growth factors and cytokines via phosphorylation and is
involved in regulating the cell cycle and promoting cell
proliferation and differentiation (26). In the present study, following
administration of DMN, the mRNA and protein expression levels of
CXCR4 and ERK1/2 increased significantly; these levels increased
further during YGD treatment, indicating that YGD induces the
differentiation of BMSCs into HLCs by activating SDF-1/CXCR4 and
directly phosphorylating the downstream effectors of the
MAPK/ERK1/2 signaling pathway.
NF-κB is responsible for the rapid induction of
various cytokines and adhesion molecules involved in immune and
inflammatory responses, is the downstream substrate for numerous
cytokine signaling pathways and is important in regulating the
proliferation, survival and apoptosis of cells. Following cellular
stimulation, inhibitor of κB α is phosphorylated and promotes the
translocation of NF-κB to the nucleus where it binds to NF-κB DNA
binding sites and induces transcription. Previous studies have
verified that NF-κB may participate in adjusting the activation of
Kupffer cells and HSCs during liver fibrosis (27), which is regulated by the
MAPK/ERK1/2 and Wnt/β-catenin signaling pathways via the
differentiation of BMSCs (28,29).
In the present study, the NF-κB p65 mRNA and protein expression
levels increased following DMN administration, and gradually
decreased following YGD treatment; these results indicated that YGD
induced differentiation of BMSCs to reverse liver fibrosis by
inhibiting the activation of HSCs, leading to a decrease in NF-κB
p65 levels.
Furthermore, the present study observed that the
mRNA and protein expression levels of β-catenin increased following
DMN administration, and reduced following YGD treatment, which
indicated that YGD inhibited β-catenin expression and thus induced
the differentiation of BMSCs into HLCs. β-catenin is a
multifunctional protein, which is important in the regulation of
cell differentiation and proliferation. A previous study suggested
that Wnt/β-catenin signaling pathway downregulation may induce the
differentiation of BMSCs into HLCs (30); however, in the context of the
present study, this remains to be elucidated.
In conclusion, the results of the present study
suggested that YGD reverses DMN-induced liver cirrhosis in mice,
and improves liver function and structure. YGD may act via
upregulation of the SDF-1/CXCR4 axis, which activates the
MAPK/ERK1/2 signaling pathway, leading to NF-κB p65 downregulation,
thus inducing the differentiation of BMSCs into HLCs and inhibiting
activation of HSCs.
Finally, there are remain several shortcomings in
the present study that require further improvement. The model group
at week 4 demonstrated more severe damage according to measurements
of serum liver function, pathology and IHC, compared with the NC
group at any time point, so it was selected for comparison with the
YGD and HGF groups in Western blotting and RT-qPCR experiments, but
not the NC group. This may be a limitation of this study, and
future experiments are required in order to make the substantiate
the conclusions drawn in this work.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81273925 and
81673728) and the Scientific Research Foundation of Dalian (grant
no. 2012E15SF143). The authors thank the Central Laboratory of the
First Affiliated Hospital of Dalian Medical University for support
with experimental techniques.
References
|
1
|
Wu XB and Tao R: Hepatocyte
differentiation of mesenchymal stem cells. HepatobiliaryPancreat
Dis Int. 11:360–371. 2012. View Article : Google Scholar
|
|
2
|
Pilat N, Unger L and Berlakovich GA:
Implication for bone marrow stem cells in hepatocyte regeneration
after orthotopic liver transplantation. Int J Hepatol.
2013:3106122013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang L, Ping L, Yongping M, et al: Study
on TCM recipe and syndrome of dimethylnitrosamine-induced hepatic
fibrosis in rats. J Trad Chin Med. 47:929–932. 2006.
|
|
4
|
Min L, Zhiying C and Guobin W: Investigate
Yiguanjian treatment the liver-kidney yin deficiency syndrome of
posthepatitic cirrhosis. TCM Research. 22:3–4. 2009.
|
|
5
|
Ying Z and Ping Z: Effect of YGJ on
proliferation and differenciation of hepatic oval cells in rat
cirrhosis by DMN. Journal of Dalian Medical University. 33:11–16.
2011.
|
|
6
|
Jenkins SA, Crandison A, Baxter JN, Day
DW, Taylor I and Shields R: A dimethyl nitrosamine-induced model of
cirrhosis and portal hypertension in the rat. J Hepatol. 1:489–499.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schmittgen TD, Zakrajsek BA, Mills AG,
Gorn V, Singer MJ and Reed MW: Quantitative reverse
transcription-polymerase chain reaction to study Mrna decay:
Comparison of endpoint and real-timemethods. Anal Biochem.
285:194–204. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jézéquel AM, Mancini R, Rinaldesi ML,
Ballardini G, Fallani M, Bianchi F and Orlandi F:
Dimethylnitrosamine-induced cirrhosis. Evidence for an
immunological mechanism. J Hepatol. 8:42–52. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim KS, Yang HJ, Lee JY, Na YC, Kwon SY,
Kim YC, Lee JH and Jang HJ: Effects of β-sitosterol derived from
Artemisia capillaris on the activated human hepatic stellate cells
and dimethylnitrisamine-induced mouse liver fibrosis. BMC
Complement Altern Med. 14:3632014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nishibe Y, Kaneko H, Suzuki H, Abe T,
Matsuura Y and Takaku H: Baculovirus-mediated interferon alleviates
dimethyl nitrosamine-induced liver cirrhosis symptomsin a murine
model. Gene Ther. 15:990–997. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen P, Li J, Huo Y, Lu J, Wan L, Yang Q,
Huang J, Gan R and Guo C: Adenovirus-mediated expression of orphan
nuclear receptor NR4A2 targeting hepatic stellate cell attenuates
liver fibrosis in rats. Sci Rep. 6:335932016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bell LN, Cai L, Johnstone BH, Traktuev DO,
March KL and Considine RV: A central role for hepatocyte growth
factor in adipose tissue angiogenesis. Am J Physiol Endocrinol
Metab. 294:E336–E344. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Inagaki Y, Higashi K, Kushida M, Hong YY,
Nakao S, Higashiyama R, Moro T, Itoh J, Mikami T, Kimura T, et al:
Hepatocyte growth factor suppresses profibrogenic signal
transduction via nuclear export of Smad3 with galectin-7.
Gastroenterology. 134:1180–1190. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Oh SH, Miyazaki M, Kouchi H, Inoue Y,
Sakaguchi M, Tsuji T, Shima N, Higashio K and Namba M: Hepatocyte
growth factor induces differentiation of adult rat bone marrow
cells into a hepatocyte lineage in vitro. Biochem Biophys Res
Commun. 279:500–504. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Okumoto K, Saito T, Hattori E, Ito JI,
Adachi T, Takeda T, Sugahara K, Watanabe H, Saito K, Togashi H and
Kawata S: Diffferentiation of bone marrow cells into cells that
express liver-specific genes in vitro: Implication of the Notch
signals in differentiation. Biochem Biophys Res Commun.
304:691–695. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
van de Kamp J, Jahnen-Dechent W, Rath B,
Knuechel R and Neuss S: Hepatocyte growth factor-loaded
biomaterials for mesenchymal stem cell recruitment. Stem Cells Int.
2013:8920652013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim WH, Matsumoto K, Bessho K and Nakamura
T: Growth inhibition and apoptosis in liver myofibroblasts promoted
by hepatocyte growth factor leads to resolution from liver
cirrhosis. Am J Pathol. 166:1017–1028. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tian DD, Wang W, Wang HN, Sze SC and Zhang
ZJ: Pharmacokinetic evaluation of clozapine in concomitant use of
Radix Rehmanniae, fructus schisandrae, radix bupleuri, or fructus
gardeniae in rats. Molecules. 21:E6962016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lu YY, Zhao Y, Song YN, Dong S, Wei B,
Chen QL, Hu YY and Su SB: Serum cytokine profiling analysis for
zheng differentiation in chronic hepatitis B. Chin Med. 10:242015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ping J, Chen HY, Yang Z, Yang C and Xu LM:
Effect of yiguan decoction on differentiation of bone marrow
mesenchymal stem cells into hepatocyte-like cells: An experimental
research. Zhongguo Zhong Xi Yi Jie He Za Zhi. 34:348–354. 2014.(In
Chinese). PubMed/NCBI
|
|
21
|
Anzalone R, Lo Iacono M, Corrao S, Magno
F, Loria T, Cappello F, Zummo G, Farina F and La Rocca G: New
energing potentials for human Wharton's jelly mesenchymal stem
cells: Immunological features and hepatocyte-like differentiative
capacity. Stem Cells Dev. 19:423–438. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Haridass D, Narain N and Ott M: Hepatocyte
transplantation: Waiting for stem cells. CurrOpin Organ Transplant.
13:627–632. 2008. View Article : Google Scholar
|
|
23
|
Facciorusso A, Antonino M, Del Prete V,
Neve V, Scavo MP and Barone M: Are hematopoietic stem cells
involved in hepatocarcinogenesis? Hepatobiliary Surg Nutr.
3:199–206. 2014.PubMed/NCBI
|
|
24
|
Kucia M, Ratajczak J, Reca R,
Janowska-Wieczorek A and Ratajczak MZ: Tissue-specific muscle,
neural and liver stem/progenitor cells reside in the bone marrow,
respond to an SDF-1 gradient and are mobilized into peripheral
blood during stress and tissue injury. Blood Cells Mol Dis.
32:52–57. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun L, Fan X, Zhang L, Shi G, Aili M, Lu
X, Jiang T and Zhang Y: Bone mesenchymal cell transplantation via
four routes for the treatment of acute liver failure in rats. Int J
Mol Med. 34:987–996. 2014.PubMed/NCBI
|
|
26
|
Delgado-Martín C, Escribano C, Pablos JL,
Riol-Blanco L and Rodríguez-Fernández JL: Chemokine CXCL12 uses
CXCR4 and a signaling core formed by bifunctional AKT,
extracellular signal-regulated kinase (ERK) 1/2 and mammalian
target of rapamycin complex 1(m TORC1) proteins to control
chemotaxis and survival simultaneously in mature dendritic cells. J
Biol Chem. 286:37222–37236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luedde T and Schwabe RF: NF-κB in the
liver-linking injury, fibrosis and hepatocellular carcinoma. Nat
Rev Gastroenterol Hepatol. 8:108–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang J, Liu F, Lee M, Wu B, Ting K, Zara
JN, Soo C, Al Hezaimi K, Zou W, Chen X, et al: NF-κB inhibits
osteogenic differentiation of mesenchymal stem cells by promoting
β-catenin degradation. Proc Natl Acad Sci USA. 110:9469–9474. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pandey AC, Semon JA, Kaushal D, O'Sullivan
RP, Glowacki J, Gimble JM and Bunnell BA: MicroRNA profiling
reveals age-dependent differential expression of nuclear factor κB
and mitogen-activated protein kinase in adipose and bone
marrow-derived human mesenchymal stem cells. Stem Cell Res Ther.
2:492011. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ke Z, Zhou F, Wang L, Chen S, Liu F, Fan
X, Tang F, Liu D and Zhao G: Down-regulation of Wnt signaling could
promote bone marrow-derived mesenchymal stem cells to differentiate
into hepatocytes. Biochem Biophys Res Commun. 367:342–348. 2008.
View Article : Google Scholar : PubMed/NCBI
|















