Introduction
Ischemia/reperfusion (I/R)-induced acute kidney
injury is a major challenge during circulatory arrest, ischemic
stroke, renal and cardiovascular surgery, and kidney
transplantation, and may lead to delayed graft function or acute
renal failure (1). A previous
study attributed renal I/R injury to an inflammatory process
characterized by neutrophil infiltration into the ischemic kidney
and the generation of proinflammatory cytokines and reactive oxygen
species (ROS), which are produced during reperfusion (2). ROS may induce the production of heme
oxygenase-1 (HO-1), an important component of the cellular defense
mechanism, which acts against ROS-induced I/R tissue injury,
including renal I/R injury (3,4). The
renoprotective potency of HO-1 may be attributed to the byproducts
of the HO-1 enzymatic reaction, including carbon monoxide and
bilirubin (3,4). A HO-1 agonist may reduce oxidative
stress and inducible nitrogen oxide synthase (iNOS) activity in
renal I/R injury (5). The severity
of tubular injury following I/R exposure may also be attenuated by
inhibiting the cytotoxicity of inflammatory infiltrates (3,4).
Furthermore, peroxisome proliferator-activated receptor-γ (PPARγ)
may provide renoprotection during I/R injury. It has previously
been demonstrated that PPARγ agonists may provide a potential
beneficial effect on renal function (6). PPARγ agonists attenuated renal I/R
injury through antioxidant and anti-inflammatory effects (7), inhibited renal oxidative stress in
diabetic rabbits and rats (6), and
diminished podocyte injury triggered by oxygen/glucose
deprivation-reoxygenation (8).
Furthermore, the renoprotective mechanisms of several drugs on
renal I/R injury are associated with upregulation of PPARγ
expression (9–11).
Lipoxin A4 (LXA4) is an
eicosanoid, which acts as a ‘breaking signal’ in the inflammatory
process, by promoting the reduction of inflammation via inhibition
of neutrophil infiltration and activation, reducing the response of
various cells stimulated by pathogens and proinflammatory
cytokines, and the production of proinflammatory cytokines and
toxic compounds, including ROS (12,13).
Lipoxin analogs may induce renoprotection following I/R injury via
regulation of chemokine and cytokine production, and neutrophil
recruitment (14). In a previous
study, treatment of murine renal I/R injury with a lipoxin analog
altered the induction of various pathogenic mediators, including
growth factors, cytokines, proteases and adhesion molecules, thus
suggesting that a lipoxin analog may have a renoprotective role in
the pathophysiology of renal I/R injury (15). At present, it is unclear whether
PPARγ or HO-1 contribute to lipoxin-induced renoprotection against
I/R injury. In addition to its anti-inflammatory effects,
lipoxin-induced production of PPARγ and HO-1 may also contribute to
renoprotection following I/R injury. Previous studies have revealed
that LXA4 and aspirin-triggered LXA4
upregulated the expression of HO-1 in lung tissues, and endothelial
and corneal epithelial cells (16–18).
Our previous studies also revealed that LXA4-triggered
HO-1 inhibited hypoxia/reoxygenation (H/R) injury-induced
cardiomyocyte lesions (19,20).
In addition, LXA4 may contribute to neuroprotection as a
PPARγ agonist in cerebral ischemia (21). Treatment of adult neutrophils with
LXA4 led to increased PPARγ expression levels (22). It remains to be elucidated whether
LXA4 treatment increases PPARγ or HO-1 expression in
renal tubular epithelial cells, and whether LXA4-induced
PPARγ or HO-1 expression may participate in renoprotection
following I/R injury.
Previous studies have revealed that the signaling
pathways associated with HO-1 induction include
phosphatidyinositol-3-kinase (PI3K)/Akt, mitogen-activated protein
kinase (MAPK) pathways, nuclear factor-E2-related factor 2 (Nrf2)
and antioxidant responsive element (ARE) in the HO-1 gene promoter.
The transcription factor Nrf2 interacts with ARE, and is crucial
for HO-1 transcriptional activation (19,23–26).
The signaling molecules that trigger HO-1 gene expression are
activated in a cell-specific and inducer-specific manner. Statins
may stimulate protein kinase G to increase the expression of
extracellular signal-regulated kinase (ERK) and p38 MAPK pathways,
subsequently activating HO-1 gene induction (24). Tyrosine kinase inhibitors, but not
inhibitors of the ERK and p38 MAPK pathways, may reduce the
induction of HO-1 by cadmium chloride, hemin and sodium arsenite in
human HeLa cells (25).
Furthermore, nitric oxide may increase HO-1 expression through the
formation of the Nrf2/ARE complex, independent of the MAPK or
PI3K/Akt pathways in smooth muscle cells (26). Our previous study demonstrated that
the protective effects of LXA4-induced HO-1 expression
against H/R injury in cardiomyocytes were mediated by activation of
the p38 MAPK pathway, nuclear translocation of Nrf2, and Nrf2
binding to the HO-1/ARE complex and the E1 enhancer (19). However, the signaling pathways that
contribute to LXA4-induced HO-1 expression in renal
cells remain to be elucidated. In a previous study, LXA4
was revealed to stimulate activation of p38 MAPK and ERK; however,
not PI3K in human renal mesangial cells (27). In our previous studies,
LXA4 promoted the phosphorylation of ERK, but not
PI3K/Akt, in renal mesangial cells and lung fibroblasts (28,29).
In addition, the phosphorylation of ERK and p38 MAPK, but not PI3K,
was increased in endothelial cells (30). Our previous study did not detect
LXA4-induced activation of ERK and PI3K/Akt in renal
tubular epithelial cells (31). In
addition, it has previously been revealed that LXA4 may
inhibit lipopolysaccharide-triggered ROS generation via the Nrf2
pathway in human umbilical vein cells (32). PPARγ has also been suggested to
regulate Nrf2 (33) and HO-1
(34) expression, and nitric oxide
activates PPARγ via the p38 MAPK signaling pathway (35). Therefore, the present study aimed
to determine whether p38 MAPK, ERK, PPARγ and Nrf2/ARE contribute
to LXA4-induced HO-1 expression in renal tubular
epithelial cells. Human renal tubular epithelial cells were used
and cellular injury was triggered by incubation in low-glucose
medium and H/R injury, which mimicked in vivo renal I/R
injury (8).
Materials and methods
Reagents
Fetal calf serum (FCS) and Dulbecco's modified
Eagle's medium (DMEM) were obtained from Gibco; Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). Enzyme-linked immunosorbent
assay (ELISA) kits for N-acetyl-β-glucosaminidase (NAG;
CSB-E09450), γ-glutamyl transpeptidase (γ-GT; CSB-EL009394) and
leucine aminopeptidase (LAP; CSB-E13840) levels were purchased from
Cusabio Biotech Co., Ltd. (Wuhan, China). HO-1 ELISA kit (EKS-800)
was obtained from Assays Designs; Enzo Life Science (Farmingdale,
NY, USA). The chromatin immunoprecipitation (ChIP) assay kit
(17–295) was purchased from EMD Millipore (Billerica, MD, USA).
Superoxide dismutase (SOD) activity assay kit (A001-3), iNOS assay
kit (A014-1) and malondialdehyde (MDA) assay kit (A003-4) were
obtained from Nanjing Jiancheng Bioengineering Institute (Nanjing,
China). Prime Script™ RT reagent kit and SYBR Premix Ex Taq™ were
purchased from Takara Bio, Inc. (Otsu, Japan). TRIzol reagents were
obtained from Thermo Fisher Scientific, Inc. PPARγ transcription
factor assay kit was obtained from Abcam (Cambridge, UK).
LXA4, SB203580 (an inhibitor of p38 MAPK
phosphorylation) and LY294002 (an inhibitor of the
phosphotransferase activity of PI3K), were purchased from
Calbiochem (San Diego, CA, USA). Rabbit anti-human HO-1 (sc-10789),
PPARγ (sc-7196), Nrf2 (sc-13032), total Akt 1/2/3 (sc-8312) and
serine 473-phosphorylated Akt 1/2/3 (p-Akt; sc-7985) antibodies,
PPARγ-specific small interfering RNA (siRNA;
5′-GAACAUCGAGUGUCGAAUATT−3′), Nrf2-specific siRNA
(5′-CGCUCAGAACUGUAGGAAAAGGAAGAG-3′) and control siRNA (non-specific
siRNA) were obtained from Santa Cruz Biotechnology, Inc. (Dallas,
TX, USA). Rabbit anti-human total ERK1/2 (BS3628) and threonine
202/tyrosine 204-diphosphorylated ERK1/2 (p-ERK1/2; BS5016)
antibodies were purchased from Bioworld Technology, Inc. (St. Louis
Park, MN, USA). Rabbit anti-human total p38 MAPK (2307), threonine
180/tyrosine 204-diphosphorylated p38 MAPK (p-p38 MAPK; 4511),
α/β-tubulin (2148), β-actin (4967), GAPDH (5174) antibodies,
biotin-conjugated anti-rabbit immunoglobulin G (IgG) (14708) and
horseradish peroxidase-conjugated goat anti-rabbit IgG (7074) were
obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA).
HO-1 activity assay kit was purchased from GenMed Scientifics, Inc.
(Arlington, MA, USA). A gel shift assay kit, and total and nuclear
protein extraction kit were obtained from Active Motif (Carlsbad,
CA, USA). PD98059, an inhibitor of ERK1/2 phosphorylation, zinc
protoporphyrin-IX (ZnPP-IX; a specific inhibitor of HO-1 activity),
L-glutamine, insulin, sodium pyruvate, CdCl2, trypsin,
EDTA, Triton X-100, bovine serum albumin and pioglitazone (PPARγ
agonist) were purchased from Sigma-Aldrich; Merck Millipore
(Darmstadt, Germany). Lipofectamine 2000 reagents were obtained
from Invitrogen; Thermo Fisher Scientific, Inc. Chemiluminescent
horseradish peroxidase substrate was purchased from Merck
Millipore. Protein extraction kit and bicinchoninic acid (BCA)
protein assay kit were obtained from KeyGen Biotech Co., Ltd.
(Nanjing, China). Cell Counting Kit-8 (CCK-8) was purchased from
Dojindo Molecular Technologies, Inc. (Kumamoto, Japan). Enhanced
chemiluminescence (ECL) reagent system was obtained from Amersham;
GE Healthcare (Little Chalfont, UK). Hoechst staining kit was
purchased from Beyotime Institute of Biotechnology (Shanghai,
China).
Cell culture
HK-2 human proximal tubular epithelial cells were
obtained from Type Culture Collection of the Chinese Academy of
Sciences (Wuhan, China), originated from American Type Culture
Collection (Manassas, VA, USA; no. CRL-2190). The cell monolayers
were incubated in DMEM containing 1,000 mg/l glucose, 5 µg/ml
insulin, 10% FCS, 4 mmol/l L-glutamine, 110 mg/l sodium pyruvate,
100 µg/ml streptomycin and 100 U/ml penicillin in a 5%
CO2 incubator at 37°C. Following digestion with 0.01%
EDTA and 0.25% trypsin, 8×105 cells were seeded in 50 ml plastic
culture bottles and allowed to reach 60–70% sub-confluence.
Cellular H/R injury was induced by H/R treatment. Briefly, the
cells were incubated in low-glucose DMEM in a modular incubator
chamber (BioSpherix, Parish, NY, USA) with 1% O2, 5%
CO2 and 94% N2 for 24 h (hypoxia for 24 h),
then cultured in an atmosphere containing 21% O2, 5%
CO2 and 74% N2 for 6 h (reoxygenation for 6
h). The H/R injury was induced following pretreatment with 10 nM
LXA4 for 12 h with or without co-incubation with 30 µM
SB203580, 10 µM LY294002 or 40 µM PD98059 for 30 min, 10 µM
CdCl2 for 1 h, or 10 µM pioglitazone or 20 µM ZnPP-IX
for 12 h.
siRNA transfection
The cells were cultured in 6-well plates (1×105
cells/well) for 24 h and subsequently transfected with PPARγ-siRNA,
Nrf2-siRNA or nonspecific siRNA (control siRNA) using Lipofectamine
2000 reagents according to the manufacturer's protocol. The cells
were preincubated with or without LXA4 for 12 h, then
subjected to H/R injury. The mRNA and protein expression levels of
PPARγ and HO-1 in the transfected cells were quantified using
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) and western blotting. Nrf2 protein expression was
determined using western blotting. The efficiency of Nrf2 and
PPARγ-specific siRNA transfection was determined using non-specific
siRNA.
Cell viability assay and iNOS, SOD,
MDA, LAP, NAG and γ-GT quantification
Cell viability was assessed using the CCK-8 assay.
HK-2 cells were cultured in 96-well plates and allowed to reach
40–50% sub-confluence. Subsequently, the cells were treated with
various concentrations of LXA4 (0, 0.1, 1 and 10 nM) for
6, 12 and 24 h; subsequently, the cells were exposed to H/R injury.
CCK-8 solution (10 µl) was added to each well and the incubation at
37°C was continued for an additional 1 h. Cell viability was
quantified at A450 nm using a spectrophotometer in three
wells of each group. The iNOS and SOD activity, and MDA levels in
whole cell lysates were determined using the assay kits, according
to the manufacturer's protocols. The LAP, NAG and γ-GT levels in
cellular supernatants were determined using ELISA kits according to
the manufacturer's protocols.
RT-qPCR analysis
RT-qPCR was conducted as previously described
(30). Briefly, total RNA was
isolated using TRIzol. The RNA was reverse transcribed to cDNA
using the PrimeScrpt™ RT reagent kit according to the
manufacturer's protocol. qPCR was performed using heat-activated
SYBR Premix EX Taq DNA polymerase in a TaqMan ABI 5700 Sequence
Detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). GAPDH served as internal control. The following sets of
primers were selected using Primer Premier version 5.0 software
analysis: HO-1, sense 5′-CAACCCTGCTTGCGTCCTA-3′, antisense
5′-ACCGTTCCTCCCTCCAACTA-3′; PPARγ, sense
5′-GGTCTCGATGTTGGCGCTAT-3′, antisense 5′-CCCCTCACGAAGCAGACTTT-3′;
and GAPDH, sense 5′-ACCACAGTCCATGCCATCAC-3′ and antisense
5′-TCCACCACCTGTTGCTGTA-3′. Identical amplification conditions were
applied for all reactions: 95°C for 2 min for hot start and
template denaturation prior to PCR cycling, which consisted of
three stages: 30 sec at 95°C for denaturation, 30 sec at 59°C for
annealing, 30 sec at 72°C for extension, and an additional 20 sec
at 72°C for fluorescent signal acquisition. A total of 30 cycles
were conducted. Subsequently, the Cq values were calculated for
target genes in the samples and the results were analyzed (36).
Western blot analysis
The cellular total and nuclear proteins of the
lysates were extracted using protein extraction kits according to
the manufacturer's protocol. Protein levels were determined using
the BCA protein assay kit according to the manufacturer's protocol.
Subsequently, 30 µg protein was separated by 10% SDS-polyacrylamide
gel electrophoresis, and transferred onto polyvinylidene difluoride
membranes with an electroblotting apparatus. The membranes were
incubated with the primary antibodies against HO-1, PPARγ, Nrf2,
p38 MAPK, p-p38 MAPK, p-EKR, ERK, Akt or p-Akt at 1:200 dilution,
and tubulin or β-actin at 1:1,000 dilution overnight at 4°C prior
to washing with TBS containing 0.1% Tween-20. Subsequently, the
membranes were exposed to the horseradish peroxidase-conjugated
secondary antibodies at 1:2,000 dilution for 1 h at 37°C. Finally,
the membranes were incubated with an ECL reagent system and exposed
to Kodak Biomax films (Eastman Kodak, Rochester, NY, USA).
Semi-quantitative analysis was performed using GelDoc-It2 imaging
system (UVP, LLC, Upland, CA, USA).
Quantification of HO-1 activity and
levels
HO-1 activity was assessed using the HO-1 activity
kit according to the manufacturer's protocol. The cellular HO-1
activity of the lysates was presented as pmol/mg/h. The cellular
HO-1 concentration (ng/mg) in the lysates was determined using the
ELISA kit according to the manufacturer's protocol.
Immunofluorescence assay
Intracellular Nrf2 localizations were quantified
using an immunofluorescence assay. The cells (1×105/ml) were
incubated for 30 min at 37°C on glass coverslips, and subsequently
fixed with 4% paraformaldehyde, washed and permeabilized with 0.1%
Triton X-100, and blocked with 2% bovine serum albumin in PBS at
room temperature for 30 min. Subsequently, the cells were exposed
to the Nrf2 antibodies (1:100 dilution) at room temperature for 1
h, and were washed and incubated for 30 min at 37°C with
biotin-conjugated anti-rabbit IgG at 1:500 dilution. The cells were
then incubated with fluorescein isothiocyanate-conjugated
streptavidin at room temperature for 1 h. Hoechst stain for DNA was
performed using the Hoechst staining kit, according to the
manufacturer's protocol. Coverslips were mounted on the slides, and
images of the labeled cells were observed under a fluorescence
microscope (Axiovert 200 M; Carl Zeiss AG, Oberkochen,
Germany).
HO-1 promoter analysis
The sequences for dominant-negative Nrf2 mutant
(dnNrf2) that has had its transactivation domain deleted, for
mutant E1 enhancer (M739) that had its three ARE core sequences
mutated, and for the wild-type E1 enhancer coupled to a minimum
HO-1 promoter (E1), were synthesized as previously described
(20). The plasmids expressing
dnNrf2 (pEF-F2/Nrf2), 1 µg/ml pCMVβ-galactosidase, an empty vector
and the 1 µg/ml promoter/luciferase constructs were co-transfected
into the HK-2 cells using Lipofectamine 2000 reagent according to
the manufacturer's protocol. The cells were incubated for an
additional 24 h and subsequently treated with 10 µM
CdCl2 for 1 h or 10 nM LXA4 for 12 h. The
luciferase activity of the reporter enzyme was assessed using a
TD-20/20 Turner Designs luminometer and quantified at 560 nm using
a spectrophotometer.
PPARγ transcriptional activity
assay
The transcriptional activity of PPARγ was determined
using the PPARγ transcription factor assay kit according to the
manufacturer's protocol. The nuclear extracts were extracted from
the cells using the aforementioned kit and were incubated for 15
min at 37°C in wells coated with specific PPAR response element
oligonucleotide sequences, then exposed to the primary anti-PPARγ
polyclonal antibody. Subsequently, the horseradish
peroxidase-conjugated secondary antibody and the
3,39,5,59-tetramethylbenzidine substrate were added and the
absorbance was quantified at 450 nm using a spectrophotometer.
Electrophoretic mobility shift assay
(EMSA)
The cellular nuclear protein was extracted using a
nuclear protein extraction kit according to the manufacturer's
protocol. EMSA was performed using a gel shift assay kit according
to the manufacturer's protocol. Briefly, the nuclear extracts
containing 30 µg protein were pretreated with gel shift binding
buffer for 10 min, and then exposed to the double-stranded,
biotin-labeled oligonucleotide probe of ARE (3 µg) for 20 min. The
oligonucleotide pairs of ARE were 5′-TTTATGCTGTGTCATGGTT-3′ and
5′-AACCATGACACAGCATAAA-3′. The resulting nuclear protein-DNA
complexes were merged in 4% non-denaturing polyacrylamide gels and
electrophoresis was performed at 220 V for 2 h. Subsequently, the
active bands in the gels were visualized on X-ray films. The
antibody supershift assay was performed using 1 µg Nrf2 antibody
added to the reaction mixture and incubated for 3 h at 4°C prior to
addition of the probe. To determine the reaction specificity,
competition assays were conducted with 100-fold excess of unlabeled
consensus oligonucleotide pairs of ARE, which were added to the
binding reaction mixture 10 min prior to the addition of the
labeled probes.
ChIP assay
ChIP assays were conducted using the ChIP assay kit
according to the manufacturer's protocol. Briefly, the cells were
lysed in SDS-lysis buffer and subsequently sonicated. The DNA and
proteins were cross-linked with formaldehyde. Sheared chromatin was
immunocleared using protein agarose-A. The portions of the
precleared chromatin were stored and labeled as ‘input DNA’. The
remaining chromatin was immunoprecipated with Nrf2 antibodies, IgG
was used as a control. The protein-DNA complexes were eluted using
elution buffer from the antibodies. The formaldehyde cross-links
were reversed by exposure to NaCl and heating at 65°C for 4 h. The
DNA was purified and PCR was performed under aforementioned
thermocycling conditions using a primer pair that spanned the mouse
HO-1 E1 enhancer. The following primers were used: E1 forward,
5′-AAGAGCTCCACCCCCACCCA-3′ and reverse, 5′-GGGCTAGCATGCGAAGTGAG-3′.
The PCR products were separated and examined using electrophoresis
with 1.5% agarose gel and ethidium bromide.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Significant differences were analyzed using one-way analysis of
variance followed by least significant difference test using SPSS
version 14.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
LXA4 reduces H/R
injury
The treatment duration required for
LXA4-induced protection against H/R injury was assessed.
Pretreatment with LXA4 for 12 h at 10 nM provided
maximum protection during H/R injury as indicated by CCK-8
assessment (data not shown). As indicated in Fig. 1A, LXA4 pretreatment led
to significantly increased cell viability during H/R injury
(P<0.05). Similarly, the H/R injury-induced changes in oxidative
and nitrosative stress parameters, such as MDA and iNOS, were
significantly reduced by LXA4 pretreatment (Fig. 1B; P<0.05). H/R injury
significantly reduced SOD levels, which in turn may increase MDA
levels, this was also significantly reversed in the group that
received LXA4 pretreatment (P<0.05; Fig. 1C). As presented in Fig. 1C and D the levels of γ-GT, LAP and
NAG were significantly higher in the group undergoing H/R injury
compared with the control group (P<0.05). LXA4
pretreatment significantly reduced the levels of γ-GT, LAP and NAG
during H/R injury compared with the H/R only treatment group
(P<0.05). In addition, inhibition of PPARγ expression with PPARγ
siRNA and suppression of HO-1 with ZnPP-IX, significantly reduced
the cytoprotection of LXA4 on cell viability (P<0.05;
Fig. 1A) and the changes of MDA,
iNOS, SOD, γ-GT, LAP and NAG levels (Fig. 1B-D).
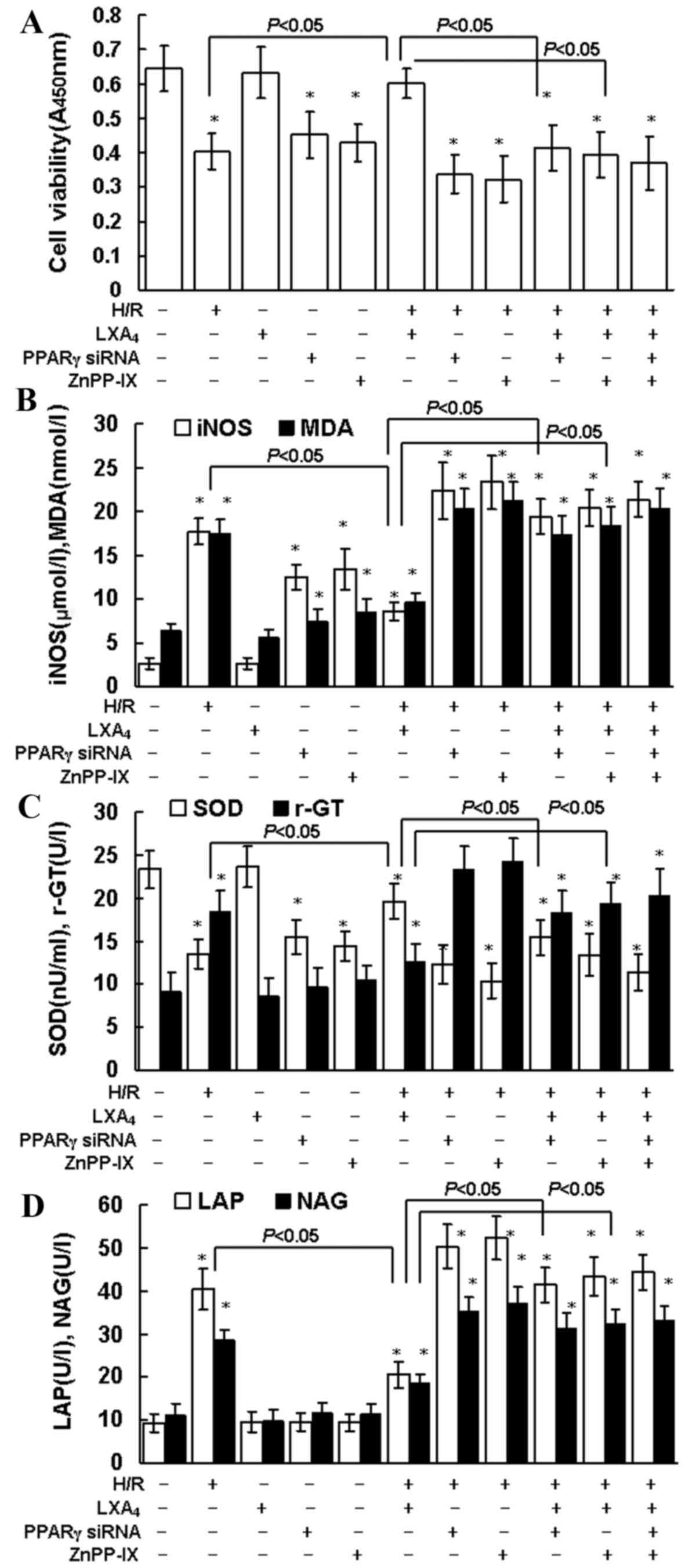 | Figure 1.Cell viability, iNOS, MDA, SOD, γ-GT,
LAP and NAG production. (A) HK-2 cell viability was determined
using Cell Counting Kit-8 assay. (B) iNOS (µmol/l) and MDA (nmol/l)
concentration, and (C) SOD and γ-GT activity in whole cell lysates
were determined using the assay kits. (D) LAP and NAG
concentrations in cellular supernatants were assessed using ELISA
kits. The cells were pretreated with 10 nM LXA4 for 12
h, 20 µM ZnPP-IX or PPARγ siRNA transfection for 12 h, and then
exposed to H/R. Untreated cells were used as the control group.
Data are presented as the mean ± standard deviation of 5
independent experiments. *P<0.05 vs. control group. iNOS, nitric
oxide synthase; MDA, malondialdehyde; SOD, superoxide dismutase;
γ-GT, γ-glutamyl transpeptidase; LAP, leucine aminopeptidase; NAG,
N-acetyl-β-glucosaminidase; LXA4, lipoxin A4;
PPARγ, peroxisome proliferator-activated receptor-γ; siRNA, small
interfering RNA; H/R, hypoxia/reoxygenation. |
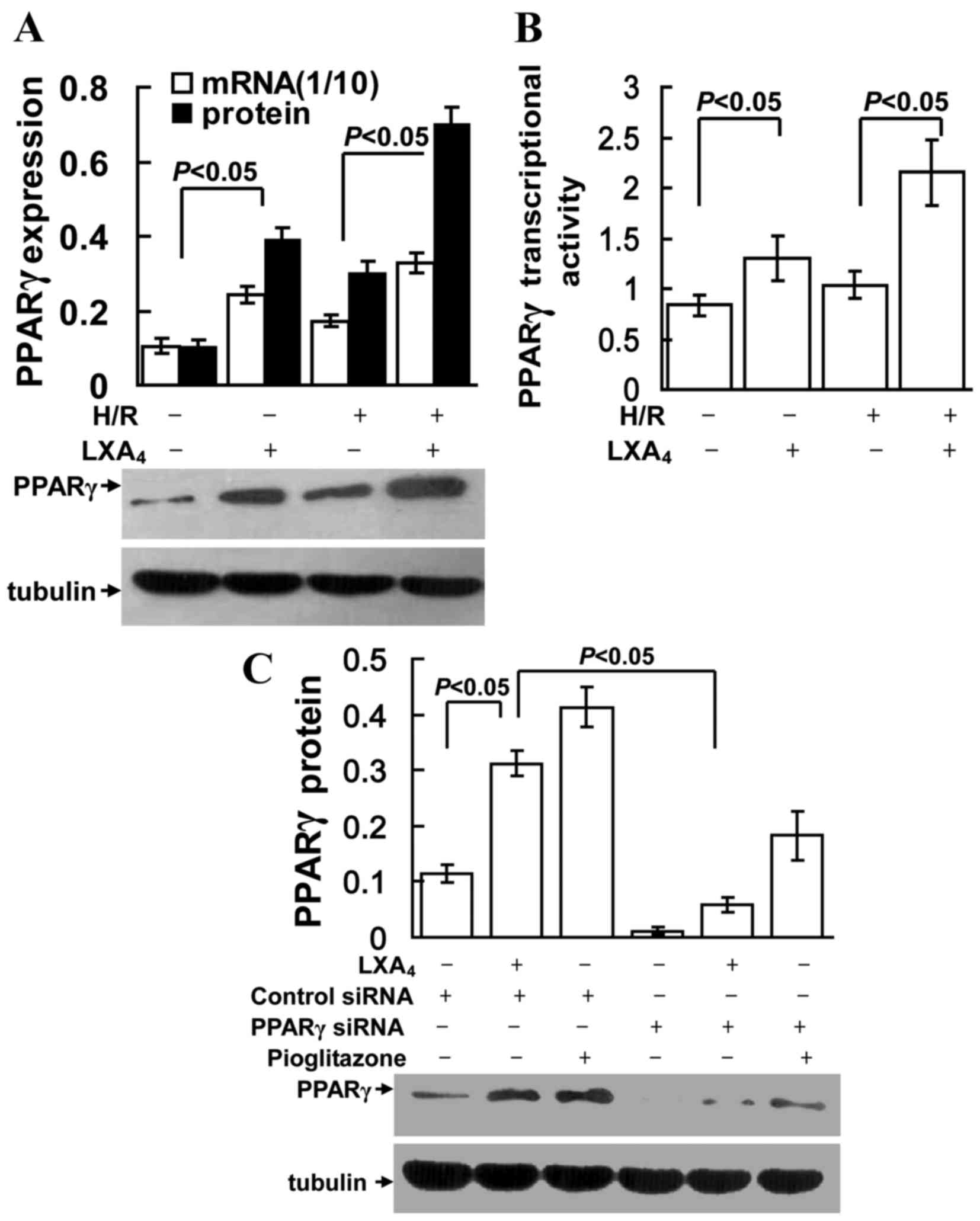
LXA4 induces PPARγ
expression
As presented in Fig. 2A
and B, LXA4 treatment significantly increased PPARγ
mRNA and protein expression, and transcriptional activity in groups
with or without H/R injury (P<0.05). However, as shown in
Fig. 2C, the LXA4- and
pioglitazone (PPARγ agonist)-induced PPARγ expression was
specifically suppressed by transfection of the cells with PPARγ
siRNA.
LXA4 induces PPARγ
expression via p38 MAPK
The role of ERK, Akt and p38 MAPK inhibition on the
induction of PPARγ is presented in Fig. 3. SB203580 (a p38 MAPK pathway
inhibitor) significantly reduced LXA4-induced, H/R
injury-triggered, and LXA4 pretreatment followed by H/R
injury-induced PPARγ expression, conversely the ERK inhibitor
PD98059 or PI3K inhibitor LY294002 did not affect PPARγ expression.
As indicated in Fig. 3B,
LXA4 pretreatment slightly increased p-p38 MAPK
expression compared with the control group. However, no significant
difference was observed in p-Akt or p-ERK1/2 expression between the
control and LXA4 groups (Fig. 3C and D). H/R exposure significantly
upregulated p-ERK1/2, p-p38 MAPK and p-Akt expression levels in
cells without LXA4 pretreatment (P<0.05; Fig. 3). In addition, LXA4
pretreatment significantly upregulated H/R injury-induced p-p38
MAPK expression levels (P<0.05; Fig. 3B); however, no significant
difference was identified for p-ERK1/2 or p-Akt expression levels
(Fig. 3C and D).
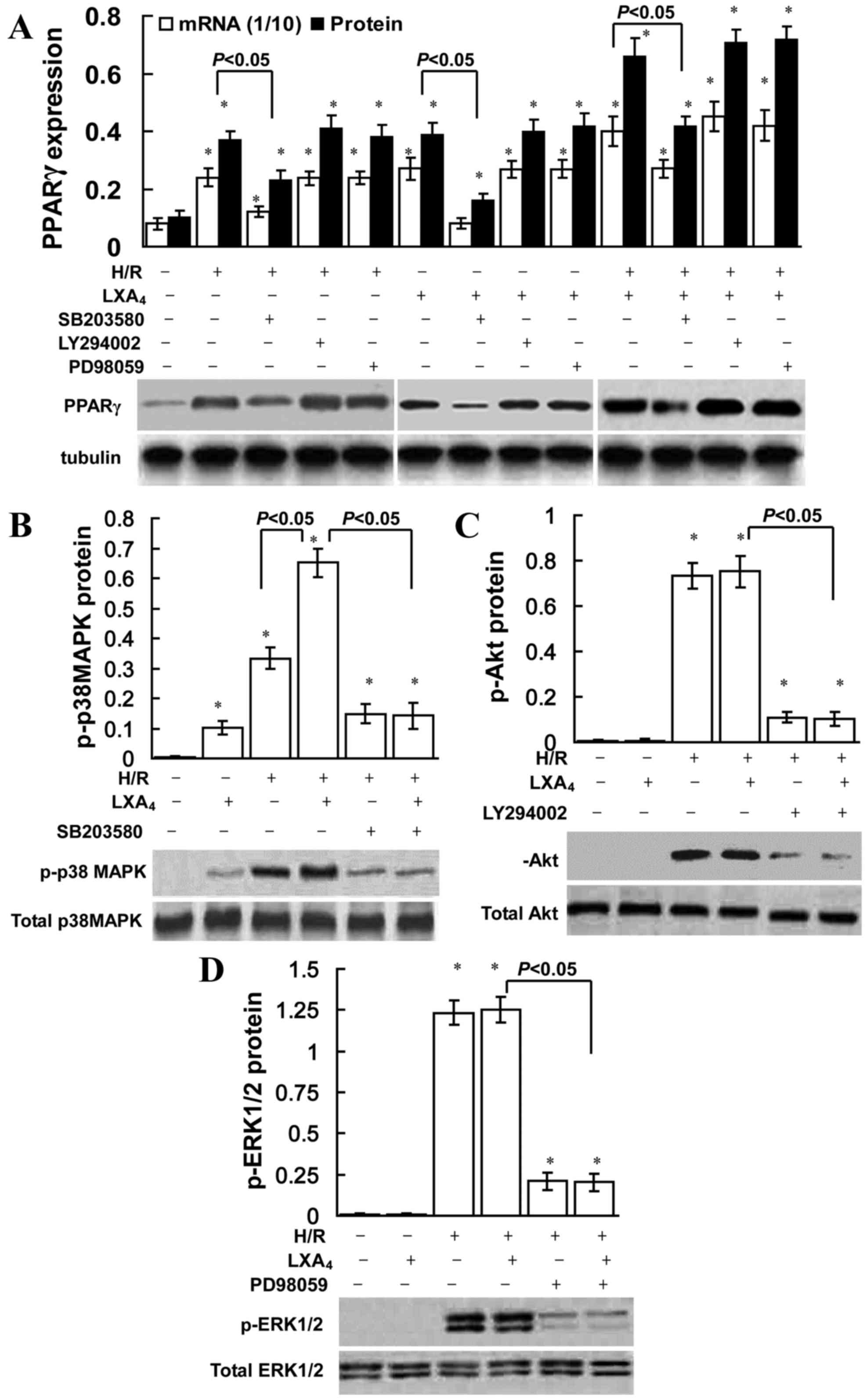 | Figure 3.Role of p38 MAPK in
LXA4-triggered PPARγ expression. HK-2 cells were
pretreated with 10 nM LXA4 for 12 h, 40 µM PD98059, 10
µM LY294002 and 30 µM SB203580 for 30 min, and then exposed to H/R
injury. (A) mRNA and nuclear protein expression levels of PPARγ
were determined using quantitative PCR and western blotting. The
amount of PCR products was normalized with GAPDH to determine the
relative expression ratio (mRNA expression ratio ×1/10) for each
mRNA. PPARγ protein expression was presented as PPARγ/tubulin ratio
for each sample. Expression levels of (B) total and p-p38 MAPK, (C)
total and p-Akt, (D) and total and p-ERK1/2 protein were assessed
using western blotting. Data are presented as the mean ± standard
deviation of 5 independent experiments. *P<0.05 vs. control
group. PPARγ, peroxisome proliferator-activated receptor-γ; H/R,
hypoxia/reoxygenation; LXA4, lipoxin A4;
p-p38MAPK, phosphorylated-p38 mitogen-activated protein kinase;
p-Akt, phosphorylated-Akt; p-ERK1/2, phosphorylated
extracellular-signal regulated kinase; PCR, polymerase chain
reaction. |
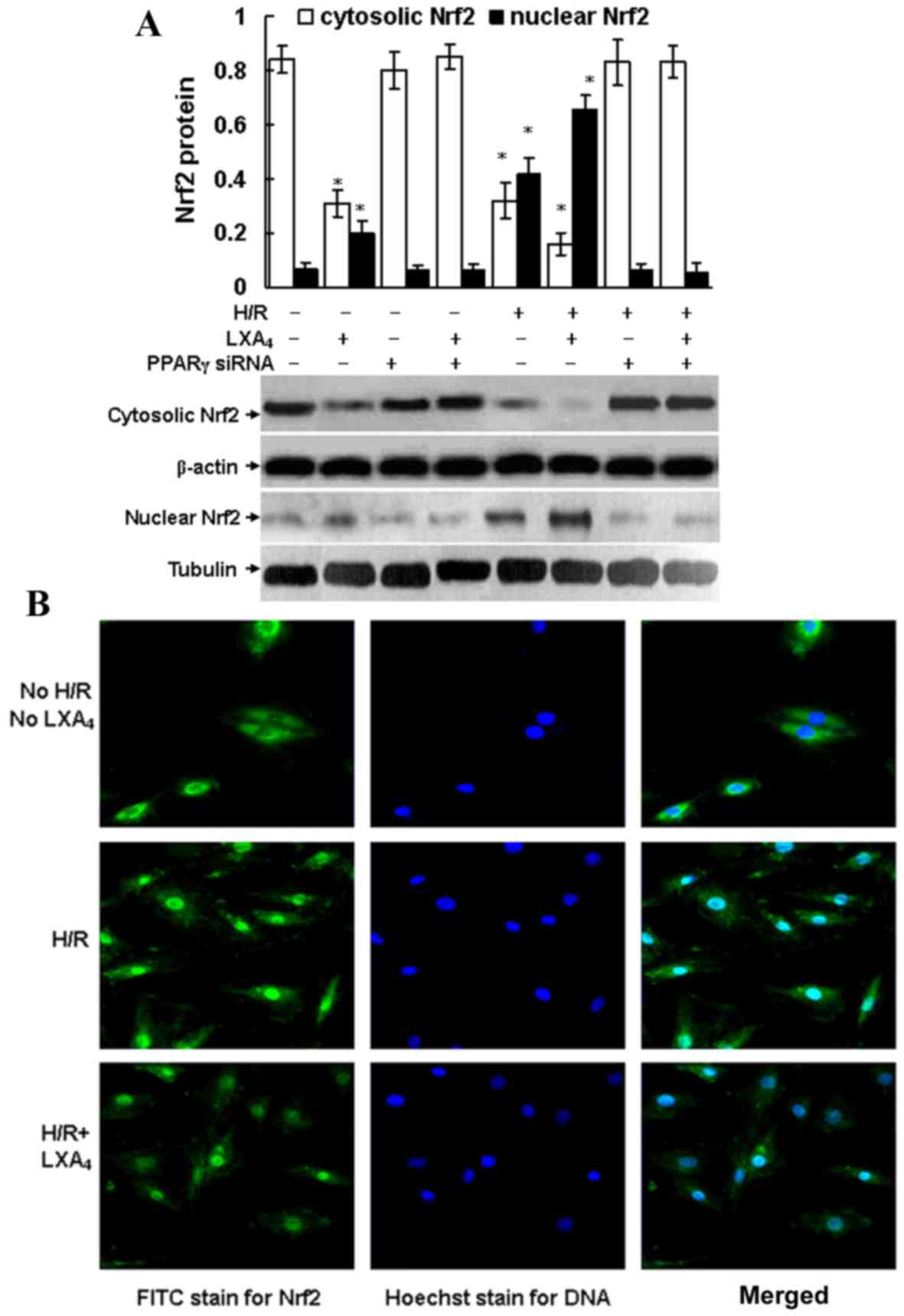
LXA4-triggered Nrf2
translocation is dependent on PPARγ activation
As shown in Fig.
4A, intracellular Nrf2 translocation from cytoplasm to nuclei
was triggered by H/R injury and LXA4 pretreatment
followed by H/R injury, since Nrf2 protein expression was
suppressed in the cytoplasm and enhanced in the nuclei of the cells
after H/R exposure and LXA4 pretreatment followed by H/R
injury. Pretreatment with LXA4 increased the H/R
injury-induced Nrf2 translocation from the cytoplasm to the nucleus
(Fig. 4). Fluorescence microscopy
confirmed the translocation of Nrf2. As presented in Fig. 4B, Nrf2 protein was only detected in
the cytoplasm of cells that were not exposed to H/R and
LXA4. In addition, LXA4 pretreatment of
H/R-injured cells led to a significantly increased expression of
Nrf2 in the nucleus compared with the control group (P<0.05;
Fig. 4A). No immunofluorescence
was identified in the negative controls using secondary antibodies
alone (data not shown). Furthermore, it was determined that Nrf2
translocation was dependent on PPARγ activation since transfection
of the cells with PPARγ siRNA completely abolished the Nrf2
translocation induced by H/R injury and LXA4
pretreatment followed by H/R injury. The cytosolic and nuclear Nrf2
expressions in the cells transfected with PPARγ siRNA were the same
as the expressions in the control cells without any treatment
(Fig. 4A).
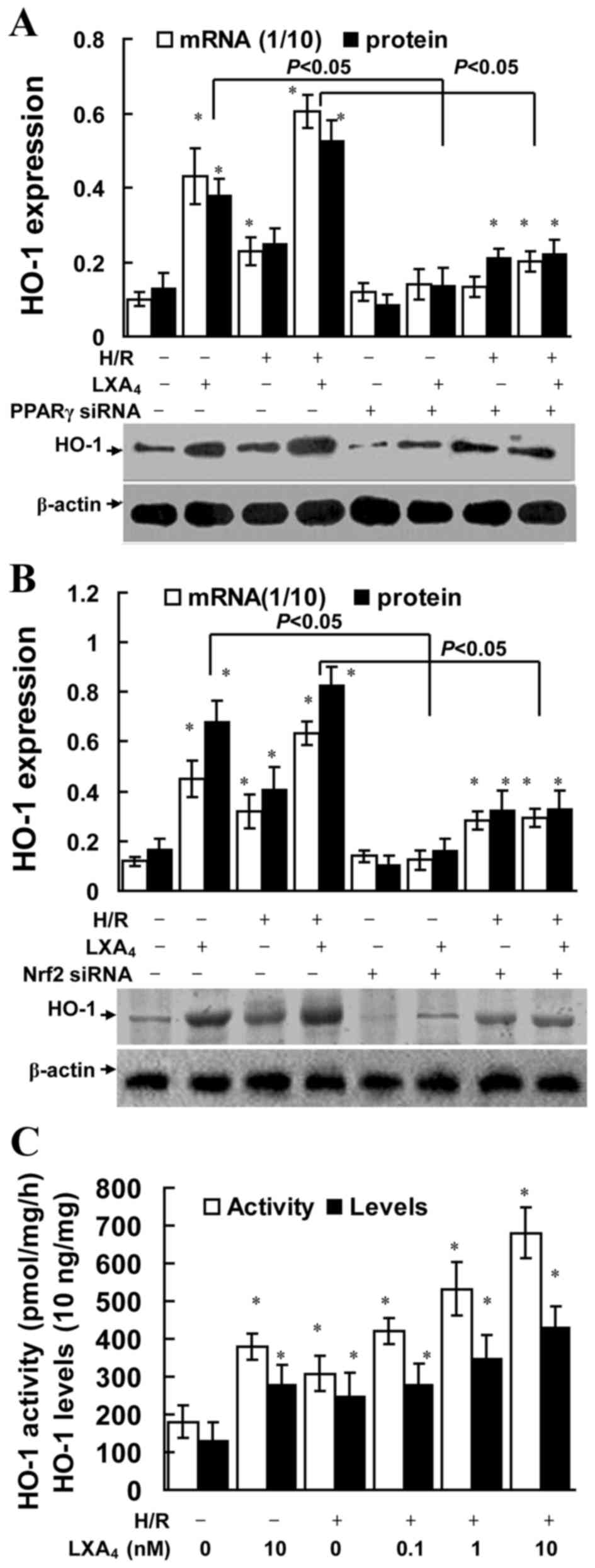
LXA4 pretreatment induces
HO-1 expression
As illustrated in Fig.
5A and B, LXA4 significantly increased HO-1 mRNA and
protein expression in cells with or without H/R injury (P<0.05).
LXA4 pretreatment significantly upregulated HO-1
activity levels in the cells without H/R injury (P<0.05;
Fig. 5C). In addition,
LXA4 pretreatment significantly upregulated the levels
and enzyme activity of HO-1 in a dose-dependent manner in the cells
enduring H/R injury (P<0.05; Fig.
5C). Furthermore, it was revealed that the
LXA4-induced HO-1 expression levels were dependent of
PPARγ and Nrf2 expression, since cells transfected with PPARγ siRNA
or Nrf2 siRNA exhibited significantly reduced HO-1 expression
levels (P<0.05; Fig. 5A and
B).
LXA4 stimulates HO-1 gene
transcription through formation of the Nrf2/ARE complex
As presented in Fig.
6A, treatment with CdCl2, which is an established
activator of the HO-1 promoter, led to a 4.5-fold increase in HO-1
promoter activity. Pretreatment with LXA4 or exposure to
H/R injury let to a 4.8 fold increase in HO-1 promoter activity.
Pretreatment with LXA4 and exposure to H/R injury may
induce a 6.3 fold upregulation in HO-1 promoter activity.
Transfection with M739, which induced an ARE mutation and served as
a negative control for E1 enhancer, reduced the cellular HO-1
promoter response to LXA4 pretreatment, H/R exposure,
and LXA4 pretreatment followed by H/R injury. In
addition, transfection with dnNrf2, which blocked formation of the
Nrf2/ARE complex, limited the cellular HO-1 promoter response to
LXA4 pretreatment, H/R exposure and LXA4
pretreatment followed by H/R injury. EMSA demonstrated the Nrf2/ARE
complex formation. As depicted in Fig.
6B, the Nrf2 binding activity was upregulated following the
pretreatment with LXA4, H/R exposure and LXA4
pretreatment followed by H/R injury. Nrf2 antibody pretreatment
reduced Nrf2 binding activity triggered by LXA4
pretreatment, H/R exposure and LXA4 pretreatment
followed by H/R injury and promoted the formation of the
Nrf2-anti-Nrf2 complex. Competition assay was performed using the
unlabeled oligonucleotide probes in order to determine the
specificity of ARE. As indicated in Fig. 6C, the ChIP assay confirmed the
findings elucidated from EMSA. The ChIP assay used the Nrf2
antibody and demonstrated the binding of Nrf2 to the E1 enhancer
and that this binding was upregulated following pretreatment with
LXA4, H/R exposure and LXA4 pretreatment
followed by H/R injury.
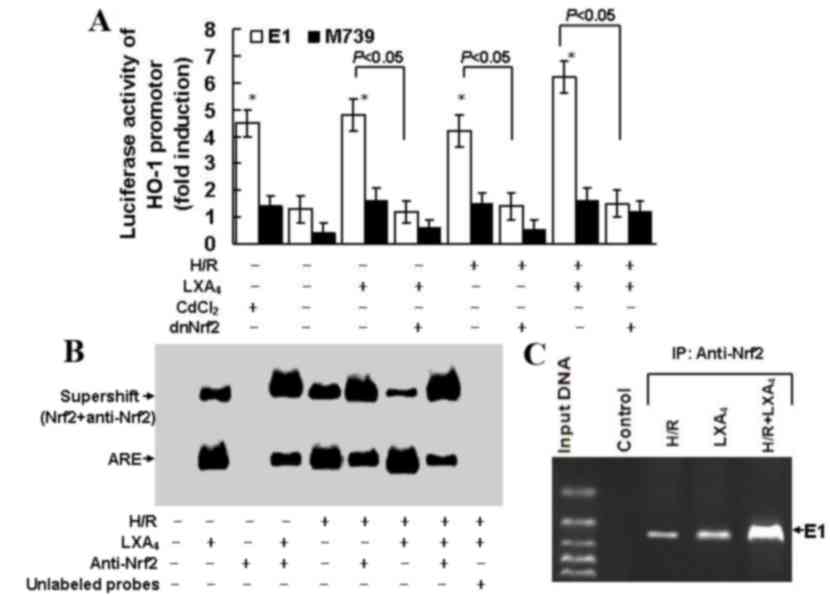 | Figure 6.LXA4-induced HO-1
expression was dependent on formation of the Nrf2/ARE complex. (A)
HK-2 cells were transfected with M739, E1, dnNrf2, then treated
with LXA4 followed by exposure to H/R injury. Fold
induction of luciferase activity of the HO-1 promoter was
determined using reporter gene transfection assays. Data are
presented as the mean ± standard deviation of 5 independent
experiments. *P<0.05 vs. untreated control group. (B) Nuclear
extracts of the cells were subjected to electrophoretic mobility
shift assay with biotin-labeled double-stranded oligonucleotide
probe of ARE. Supershift assay was conducted using the Nrf2
antibody. (C) Binding activity of Nrf2 to E1 was assessed using a
chromatin immunoprecipitation assay in cells subjected to H/R
injury, LXA4 or H/R injury and LXA4. HO-1,
heme oxygenase-1; E1, mouse HO-1 promoter construct; M739, mutated
mouse HO-1 promoter construct; H/R, hypoxia/reoxygenation;
LXA4, lipoxin A4; dnNrf, dominant negative
nuclear factor E2-related factor 2; ARE, antioxidant responsive
element. |
Discussion
A previous study suggested that there is therapeutic
potential for the use of lipoxin and its analogs in animal models
of renal disease (37). In a
previous study, a lipoxin analog reduced neutrophil recruitment in
anti-glomerular basement membrane cells in a murine nephritis model
(38), whereas LXA4 and
benzo-LXA4 have been reported to attenuate experimental
renal fibrosis (39). Furthermore,
the LXA4-triggered increase of let-7c has been reported
to lead to suppression of renal fibrosis (40) and LXA4 protected renal
function against rhabdomyolysis-induced acute kidney injury in rats
(41). Our previous study also
revealed that the LXA4 analog inhibited mesangial
proliferation and inflammation in mesangioproliferative nephritis
in rats (42). As aforementioned,
LXA4 analogs may lead to protection against renal I/R
injury (15) and ischemic acute
renal failure (14). The present
study revealed that LXA4 protected renal tubular
epithelial cells against H/R injury. The CCK-8 assay demonstrated
that LXA4 provided protection for cells exposed to H/R
injury, with regards to cellular viability (Fig. 1A). In addition, H/R injury-induced
changes in oxidative and nitrosative stress parameters were
reversed by LXA4 pretreatment (Fig. 1B). LXA4 pretreatment
also reduced the release of γ-GT, LAP and NAG from tubular
epithelial cells subjected to H/R injury (Fig. 1C).
Since in vitro H/R injury mimics in
vivo renal I/R injury (8), the
present study was conducted in HK-2 cells in order to determine the
intracellular mechanism by which LXA4 induced protection
against H/R injury. Previous studies have reported that
LXA4-induced protection against renal I/R injury may be
attributed to the inhibition of neutrophil recruitment, and to the
suppression of proinflammatory cytokines and chemokines (14,15).
To the best of our knowledge, the present study was the first to
reveal that LXA4-induced PPARγ and HO-1 expression was
involved in LXA4-induced renoprotection. Initially,
LXA4 pretreatment alone or LXA4 pretreatment
followed by H/R injury increased the mRNA and protein expression
levels, and activities, of PPARγ and HO-1 in HK-2 cells (Figs. 2A and B; 5A and C). In addition, ZnPP-IX treatment
and PPARγ siRNA transfection inhibited the LXA4-induced
protection on cell viability (Fig.
1A), and reversed the LXA4-modulated oxidative and
nitrosative stress parameters (Fig.
1B), and release of γ-GT, LAP and NAG (Fig. 1C and D) in cells exposed to H/R
injury. These findings demonstrated that LXA4-induced
renoprotection was mediated by PPARγ and HO-1 upregulation in HK-2
cells subjected to H/R injury. These findings are consistent with a
previous study, which indicated that LXA4 induced
neuroprotection by acting as a PPARγ agonist in cerebral ischemia
(21). Furthermore, treatment of
adult neutrophils with LXA4 previously resulted in
increased PPARγ expression (22),
and topical application of LXA4 exerted an
anti-inflammatory effect on corneal wound healing through HO-1
upregulation in murine and human corneal epithelial cells (17). In addition, an LXA4
analog ameliorated lipopolysaccharide-evoked acute lung injury in
mice via upregulation of HO-1 in lung tissues (18), LXA4-triggered HO-1 has
also been revealed to reduce cardiomyocyte injury induced by
exposure to H/R (19). The present
study demonstrated that LXA4-induced HO-1 expression was
dependent on PPARγ expression, since transfection with PPAR-γ siRNA
reduced LXA4-induced HO-1 upregulation (Fig. 5A). Similarly, previous studies
indicated that HO-1 induction was upregulated by PPARγ in human
smooth muscle cells and vascular endothelial cells (34), and cilostazol, an inhibitor of
phosphodiesterase type III, protected endothelial cells against
tumor necrosis factor-α-induced cytotoxicity through HO-1 induction
via a PPARγ-dependent pathway (43).
As aforementioned, signaling transduction pathways
that participate in PPARγ and HO-1 production, act in an
inducer-specific and cell-specific manner (24–26,35).
In the present study, LXA4 alone and LXA4
pretreatment followed by H/R injury induced PPARγ expression, which
was partially dependent on p38 MAPK activation, since suppression
of p-p38MAPK with SB203580, but not p-ERK or PI3K/Akt suppression,
reduced PPARγ production triggered by LXA4 pretreatment,
H/R exposure and LXA4 pretreatment followed by H/R
injury (Fig. 3A). In addition,
LXA4 increased p-p38 MAPK expression; however, no
difference in p-Akt or p-ERK1/2 expression was detected in the
cells with or without LXA4 treatment (Fig. 3B-D). These findings, to the best of
our knowledge, were the first to indicate that p38 MAPK
phosphorylation may contribute to PPARγ/HO-1 production triggered
by LXA4 pretreatment in renal cells. It has previously
been demonstrated that nitric oxide-induced PPARγ activation may
occur in a p38 MAPK signaling pathway-dependent manner in human
umbilical vein endothelial cells (35). Furthermore, LXA4-induced
HO-1 was able to inhibit cardiomyocyte injury following exposure to
H/R, via p38 MAPK activation (19).
Previous studies have suggested that Nrf2 acts as
regulator of the transcriptional upregulation of HO-1 and of the
adaptive response to oxidative stress (44,45).
Previous studies have also indicated that Nrf2 may contribute to
ARE-triggered transcriptional upregulation of HO-1 and antioxidant
gene expression (44,45). LXA4 reduced the
permeability of endothelial cells via Nrf2 activation in HO-1
upregulation (32). Therefore, the
role of Nrf2/ARE in LXA4-induced renoprotection required
further investigation. The present study was, to the best of our
knowledge, the first to clarify that Nrf2 translocation and
Nrf2/ARE activation were required for LXA4-induced HO-1
expression in renal cells. Initially, LXA4 pretreatment
and LXA4 pretreatment followed by H/R injury induced
upregulation of HO-1 mRNA and protein levels, which were reduced
following transfection with Nrf2-siRNA (Fig. 5B). In addition, LXA4
pretreatment, H/R exposure or LXA4 pretreatment followed
by H/R injury all promoted Nrf2 translocation from the cytoplasm to
the nucleus (Fig. 4).
Immunofluorescence also confirmed that LXA4 resulted in
Nrf2 translocation from the cytoplasm into the nuclei following
exposure to H/R, since LXA4 treatment led to an increase
in nuclear Nrf2 staining (Fig.
4B).
The present study aimed to determine whether
LXA4-induced HO-1 expression was associated with
transcriptional activation of ARE in the HO-1 promoter by
transfecting cells with a HO-1 promoter construct E1 and mutant E1
(M739). Subsequently, LXA4 pretreatment and
LXA4 pretreatment followed by H/R injury upregulated
HO-1 basal transcription, whereas transfection with M739 reduced
the upregulation of HO-1 promoter activity (Fig. 6A). Therefore, it is possible that
LXA4 activated HO-1 gene transcription via
transcriptional activation of ARE. Furthermore, the Nrf2/ARE
binding in the cells following LXA4 pretreatment was
assessed using EMSA, which demonstrated that the DNA binding
activity of Nrf2 was enhanced by LXA4 pretreatment, H/R
exposure and LXA4 pretreatment followed by H/R injury.
From the supershift EMSA reactions, it was evident that treatment
with anti-Nrf2 reduced migration of the ARE complex, thus
suggesting Nrf2 was present in the ARE-nuclear protein complex
(Fig. 6B). In addition, ChIP
assays indicated that Nrf2 binding to the HO-1 E1 enhancer may be
triggered by LXA4 pretreatment, H/R exposure and
LXA4 pretreatment followed by H/R injury (Fig. 6C). The present study also
demonstrated that LXA4 pretreatment and LXA4
pretreatment followed by H/R injury-induced translocation of Nrf2
were dependent on PPARγ expression levels, since cells transfected
with PPARγ siRNA exhibited reduced Nrf2 translocation (Fig. 4A). A previous study demonstrated
that PPARγ was able to regulate the expression of Nrf2 in the
pathogenesis of intracerebral hemorrhage (33).
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that LXA4
protected tubular epithelial cells against H/R injury via
activation of the p38 MAPK/PPARγ/Nrf2-ARE/HO-1 pathway; however,
the PI3K/Akt or ERK pathways were not involved. These findings
elucidated the underlying mechanisms by which LXA4
induced renoprotection during H/R injury. In conjunction with
previous studies on the efficacy of LXA4 and
LXA4 analogs in treatment of renal diseases (14,15,38–42),
the present study determined that LXA4 may be a novel
promising therapeutic agent for ischemic renal diseases.
Acknowledgements
The present study was supported by the National
Natural Scientific Grant (grant nos. 81270821 and 81300521) from
the Government of China and by the Priority Academic Program
Development of Jiangsu Higher Education Institutions (grant no.
JX10231801).
References
|
1
|
Wen X, Murugan R, Peng Z and Kellum JA:
Pathophysiology of acute kidney injury: A new perspective. Contrib
Nephrol. 165:39–45. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Eltzschig HK and Eckle T: Ischemia and
reperfusion-from mechanism to translation. Nat Med. 17:1391–1401.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chok MK, Ferlicot S, Conti M, Almolki A,
Dürrbach A, Loric S, Benoît G, Droupy S and Eschwège P:
Renoprotective potency of heme oxygenase-1 induction in rat renal
ischemia-reperfusion. Inflamm Allergy Drug Targets. 8:252–259.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nath KA: Heme oxygenase-1: A provenance
for cytoprotective pathways in the kidney and other tissues. Kidney
Int. 70:432–443. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li Volti G, Sorrenti V, Murabito P,
Galvano F, Veroux M, Gullo A, Acquaviva R, Stacchiotti A, Bonomini
F, Vanella L and Di Giacomo C: Pharmacological induction of heme
oxygenase-1 inhibits iNOS and oxidative stress in renal
ischemia-reperfusion injury. Transplant Proc. 39:2986–2991. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Iglesias P and Diez JJ: Peroxisome
proliferator-activated receptor gamma agonists in renal disease.
Eur J Endocrinol. 154:613–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Reel B, Guzeloglu M, Bagriyanik A, Atmaca
S, Aykut K, Albayrak G and Hazan E: The effects of PPAR-γ agonist
pioglitazone on renal ischemia/reperfusion injury in rats. J Surg
Res. 182:176–184. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miglio G, Rosa AC, Rattazzi L, Grange C,
Collino M, Camussi G and Fantozzi R: The subtypes of peroxisome
proliferator-activated receptors expressed by human podocytes and
their role in decreasing podocyte injury. Br J Pharmacol.
162:111–125. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ragab D, Abdallah DM and El-Abhar HS:
Cilostazol renoprotective effect: Modulation of PPAR-γ, NGAL, KIM-1
and IL-18 underlies its novel effect in a model of
ischemia-reperfusion. PLoS One. 9:e953132014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Singh JP, Singh AP and Bhatti R: Explicit
role of peroxisome proliferator-activated receptor gamma in gallic
acid-mediated protection against ischemia-reperfusion-induced acute
kidney injury in rats. J Surg Res. 187:631–639. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu QQ, Wang Y, Senitko M, Meyer C, Wigley
WC, Ferguson DA, Grossman E, Chen J, Zhou XJ, Hartono J, et al:
Bardoxolone methyl (BARD) ameliorates ischemic AKI and increases
expression of protective genes Nrf2, PPARγ and HO-1. Am J Physiol
Renal Physiol. 300:F1180–F1192. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Serhan CN and Chiang N: Endogenous
pro-resolving and anti-inflammatory lipid mediators: A new
pharmacologic genus. Br J Pharmacol. 153 Suppl 1:S200–S215. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nascimento-Silva V, Arruda MA,
Barja-Fidalgo C and Fierro IM: Aspirin-triggered lipoxin A4 blocks
reactive oxygen species generation in endothelial cells: A novel
antioxidative mechanism. Thromb Haemost. 97:88–98. 2007.PubMed/NCBI
|
|
14
|
Leonard MO, Nannan K, Burne MJ, Lappin DW,
Doran P, Coleman P, Stenson C, Taylor C, Daniels F, Godson C, et
al: 15-Epi-15-(para-fluorophenoxy)-lipoxin A(4)-methyl ester, a
synthetic analogue of 15-epi-lipoxin A(4), is protective in
experimental ischemic acute renal failure. J Am Soc Nephrol.
13:1657–1662. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kieran NE, Doran PP, Connolly SB, Greenan
MC, Higgins DF, Leonard M, Godson C, Taylor CT, Henger A, Kretzler
M, et al: Modification of the transcriptomic response to renal
ischemia/reperfusion injury by lipoxin analog. Kidney Int.
64:480–492. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nascimento-Silva V, Arruda MA,
Barja-Fidalgo C, Villela CG and Fierro IM: Novel lipid mediator
aspirin-triggered lipoxin A4 induces heme oxygenase-1 in
endothelial cells. Am J Physiol Cell Physiol. 289:C557–C563. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Biteman B, Hassan IR, Walker E, Leedom AJ,
Dunn M, Seta F, Laniado-Schwartzman M and Gronert K:
Interdependence of lipoxin A4 and heme-oxygenase in
counter-regulating inflammation during corneal wound healing. FASEB
J. 21:2257–2266. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jin SW, Zhang L, Lian QQ, Liu D, Wu P, Yao
SL and Ye DY: Posttreatment with aspirin-triggered lipoxin A4
analog attenuates lipopolysaccharide-induced acute lung injury in
mice: The role of heme oxygenase-1. Anesth Analg. 104:369–377.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen XQ, Wu SH, Zhou Y and Tang YR:
Lipoxin A4-induced heme oxygenase-1 protects cardiomyocytes against
hypoxia/reoxygenation injury via p38 MAPK activation and Nrf2/ARE
complex. PLoS One. 8:e671202013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen XQ, Wu SH, Zhou Y and Tang YR:
Involvement of K+ channel-dependent pathways in lipoxin A4-induced
protective effects on hypoxia/reoxygenation injury of
cardiomyocytes. Prostaglandins Leukot Essent Fatty Acids.
88:391–397. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sobrado M, Pereira MP, Ballesteros I,
Hurtado O, Fernández-López D, Pradillo JM, Caso JR, Vivancos J,
Nombela F, Serena J, et al: Synthesis of lipoxin A4 by
5-lipoxygenase mediates PPARgamma-dependent, neuroprotective
effects of rosiglitazone in experimental stroke. J Neurosci.
29:3875–3884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weinberger B, Quizon C, Vetrano AM, Archer
F, Laskin JD and Laskin DL: Mechanisms mediating reduced
responsiveness of neonatal neutrophils to lipoxin A4. Pediatr Res.
64:393–398. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alam J and Cook JL: Transcriptional
regulation of the heme oxygenase-1 gene via the stress response
element pathway. Curr Pharm Des. 9:2499–2511. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen JC, Huang KC and Lin WW: HMG-CoA
reductase inhibitors upregulate heme oxygenase-1 expression in
murine RAW264.7 macrophages via ERK, p38 MAPK and protein kinase G
pathways. Cell Signal. 18:32–39. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Masuya Y, Hioki K, Tokunaga R and Taketani
S: Involvement of the tyrosine phosphorylation pathway in induction
of human heme oxygenase-1 by hemin, sodium arsenite, and cadmium
chloride. J Biochem. 124:628–633. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu XM, Peyton KJ, Ensenat D, Wang H,
Hannink M, Alam J and Durante W: Nitric oxide stimulates heme
oxygenase-1 gene transcription via the Nrf2/ARE complex to promote
vascular smooth muscle cell survival. Cardiovasc Res. 75:381–389.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McMahon B, Stenson C, McPhillips F,
Fanning A, Brady HR and Godson C: Lipoxin A4 antagonizes the
mitogenic effects of leukotriene D4 in human renal mesangial cells.
Differential activation of MAP kinases through distinct receptors.
J Biol Chem. 275:27566–27575. 2000.PubMed/NCBI
|
|
28
|
Wu SH, Wu XH, Lu C, Dong L and Chen ZQ:
Lipoxin A4 inhibits proliferation of human lung fibroblasts induced
by connective tissue growth factor. Am J Respir Cell Mol Biol.
34:65–72. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu SH, Wu XH, Lu C, Dong L, Zhou GP and
Chen ZQ: Lipoxin A4 inhibits connective tissue growth
factor-induced production of chemokines in rat mesangial cells.
Kidney Int. 69:248–256. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu SH, Liao PY, Dong L and Chen ZQ: Signal
pathway involved in inhibition by lipoxin A(4) of production of
interleukins in endothelial cells by lipopolysaccharide. Inflamm
Res. 57:430–437. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu SH, Zhang YM, Tao HX and Dong L:
Lipoxin A(4) inhibits transition of epithelial to mesenchymal cells
in proximal tubules. Am J Nephrol. 32:122–136. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pang H, Yi P, Wu P, Liu Z, Liu Z, Gong J,
Hao H, Cai L, Ye D and Huang Y: Effect of lipoxin A4 on
lipopolysaccharide-induced endothelial hyperpermeability. Sci World
Journal. 11:1056–1067. 2011. View Article : Google Scholar
|
|
33
|
Zhao X, Gonzales N and Aronowski J:
Pleiotropic role of PPARγ in intracerebral hemorrhage: An intricate
system involving Nrf2, RXR, and NF-κB. CNS Neurosci Ther.
21:357–366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kronke G, Kadl A, Ikonomu E, Bluml S,
Fürnkranz A, Sarembock IJ, Bochkov VN, Exner M, Binder BR and
Leitinger N: Expression of heme oxygenase-1 in human vascular cells
is regulated by peroxisome proliferator-activated receptors.
Arterioscler Thromb Vasc Biol. 27:1276–1282. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ptasinska A, Wang S, Zhang J, Wesley RA
and Danner RL: Nitric oxide activation of peroxisome
proliferator-activated receptor gamma through a p38 MAPK signaling
pathway. FASEB J. 21:950–961. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta deltaC(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kieran NE, Maderna P and Godson C:
Lipoxins: Potential anti-inflammatory, proresolution and
antifibrotic mediators in renal disease. Kidney Int. 65:1145–1154.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ohse T, Ota T, Kieran N, Godson C, Yamada
K, Tanaka T, Fujita T and Nangaku M: Modulation of
interferon-induced genes by lipoxin analogue in anti-glomerular
membrane nephritis. J Am Soc Nephrol. 15:919–927. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Börgeson E, Docherty NG, Murphy M, Rodgers
K, Ryan A, O'Sullivan TP, Guiry PJ, Goldschmeding R, Higgins DF and
Godson C: Lipoxin A4 and benzo-lipoxin A4 attenuate experimental
renal fibrosis. FASEB J. 25:2967–2979. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Brennan EP, Nolan KA, Börgeson E, Gough
OS, McEvoy CM, Docherty NG, Higgins DF, Murphy M, Sadlier DM,
Ali-Shah ST, et al: Lipoxins attenuate renal fibrosis by inducing
let-7c and suppressing TGFβR1. J Am Soc Nephrol. 24:627–637. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Deng LL, Zhong L, Lei JR, Tang L, Liu L,
Xie SQ and Liao XH: Protective effect of lipoxin A4 against
rhabdomyolysis-induced acute kidney injury in rats. Xi Bao Yu Fen
Zi Mian Yi Xue Za Zhi. 28:907–910. 2012.(In Chinese). PubMed/NCBI
|
|
42
|
Wu SH, Wu XH, Liao PY and Dong L: Signal
transduction involved in protective effects of
15(R/S)-methyl-lipoxin A(4) on mesangioproliferative nephritis in
rats. Prostaglandins Leukot Essent Fatty Acids. 76:173–180. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park SY, Bae JU, Hong KW and Kim CD: HO-1
induced by cilostazol protects against TNF-α-associated
cytotoxicity via a PPAR-γ-dependent pathway in human endothelial
cells. Korean J Physiol Pharmacol. 15:83–88. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mann GE, Niehueser-Saran J, Watson A, Gao
L, Ishii T, de Winter P and Siow RC: Nrf2/ARE regulated antioxidant
gene expression in endothelial and smooth muscle cells in oxidative
stress: Implications for atherosclerosis and preeclampsia. Sheng Li
Xue Bao. 59:117–127. 2007.PubMed/NCBI
|
|
45
|
Kensler TW, Wakabayashi N and Biswal S:
Cell survival responses to environmental stresses via the
Keap1-Nrf2-ARE pathway. Annu Rev Pharmacol Toxicol. 47:89–116.
2007. View Article : Google Scholar : PubMed/NCBI
|