Introduction
Hepatocellular carcinoma (HCC), one of the most
common human malignancies, causes a large amount of mortalities
worldwide, and its incidence has increased rapidly in recent years
(1,2). Despite great efforts in diagnosis and
treatment, including surgical resection, radiotherapy and
chemotherapy, the prognosis of patients with HCC remains poor,
predominantly due to the recurrence and metastasis (2). Therefore, revealing the molecular
mechanism underlying HCC growth and metastasis is urgently
required, which may help to develop targeted treatment
strategies.
MicroRNAs (miRs) are a type of short non-coding RNA,
and generally function as negative regulators of gene expression
through directly binding to the 3′-untranslational region (3′-UTR)
of their target genes, resulting in mRNA degradation or translation
inhibition (3,4). In the last decade, miRs have been
demonstrated to participate in various biological processes,
including cell survival, proliferation, differentiation, apoptosis
and motility (3,5). Furthermore, various miRs have been
reported to act as oncogenes or tumor suppressors in human
malignancies including HCC (6–8). For
instance, miR-212, downregulated in HCC tissues, suppresses tumor
growth by inhibition of forkhead box A1 (9). miR-98 suppresses cell proliferation,
migration and invasion in HCC by targeting the collagen triple
helix repeat containing 1 oncogene (10). These miRs associated with the
development and progression of HCC may become important therapeutic
targets or reagents for this disease.
Recently, miR-137 was reported to function as a
tumor suppressor in some common human cancer types (11,12).
Shen et al (11) reported
that miR-137 inhibited tumor growth and sensitize chemosensitivity
to paclitaxel and cisplatin in lung cancer. Liang et al
(12) reported that downregulation
of miR-137 promoted cell proliferation in pediatric high-grade
gliomas. Furthermore, the suppressive role of miR-137 in HCC has
gradually been elucidated (13).
Liu et al (14) reported
that miR-137 suppresses tumor growth and metastasis in HCC by
targeting AKT serine/threonine kinase 2. Gao et al (15) reported that the suppressive effects
of miR-137 on HCC cell proliferation and metastasis were regulated
by cell division cycle 42. As one miR has multiple target genes,
there may also be other important targets of miR-137 in HCC
(3). The current study aimed to
investigate the molecular mechanism of miR-137 in regulating HCC
cell proliferation, migration and invasion.
Materials and methods
Tissue collection
The current study was approved by the Ethics
Committee of Central Hospital of Linyi (Linyi, China). A total of
89 primary HCC tissues and adjacent non-tumor tissues were
collected from patients at the Central Hospital of Linyi between
September 2010 and June 2012, and informed consent was obtained
from all participants. No patients received radiation therapy or
chemotherapy prior to surgical resection. Tissues were immediately
snap-frozen in liquid nitrogen following surgical resection, and
stored in liquid nitrogen prior to use. The clinical information of
patients involved in this study is summarized in Table I.
 | Table I.Association between miR-137 expression
and clinicopathological characteristics in hepatocellular
carcinoma. |
Table I.
Association between miR-137 expression
and clinicopathological characteristics in hepatocellular
carcinoma.
| Variable | High miR-137
expression (n=47) | Low miR-137
expression (n=42) | P-value |
|---|
| Age |
|
| 0.399 |
|
<55 | 18 | 20 |
|
| ≥55 | 29 | 22 |
|
| Sex |
|
| 1 |
|
Male | 32 | 28 |
|
|
Female | 15 | 14 |
|
| Tumor number |
|
| 0.211 |
|
Single | 33 | 35 |
|
|
Multiple | 14 | 7 |
|
| Histologic
grade |
|
| 0.384 |
|
Well-moderate | 28 | 29 |
|
|
Poor | 19 | 13 |
|
| Nodal
metastasis |
|
| 0.017a |
|
Present | 25 | 11 |
|
|
Absent | 22 | 31 |
|
| Vessel
invasion |
|
| 0.003b |
|
Present | 29 | 12 |
|
|
Absent | 18 | 30 |
|
| Hepatitis B
infection |
|
| 0.226 |
|
Present | 32 | 34 |
|
|
Absent | 15 | 8 |
|
| Clinical T
stage |
|
| 0.005b |
|
T1-2 | 27 | 11 |
|
|
T3-4 | 20 | 31 |
|
Cell culture and transfection
Human normal liver epithelial cell line THLE-3 and
four liver cancer cell lines including HepG2, Hep3B HCCLM3 and
SMCC7721 were purchased from Cell Bank of Chinese Academy of
Sciences (Shanghai, China). Cells were cultured in Dulbecco's
modified Eagle's medium (DMEM; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Thermo Fisher Scientific, Inc.) in a 37°C humidified atmosphere
with 5% CO2. For cell transfection, HepG2 cells were
transfected with scramble miR mimic (miR-NC) (CmiR0001-MR04),
miR-137 mimic (HmiR0011-MR04), negative control (NC) inhibitor
(CmiR-AN0001-SN), miR-137 inhibitor (HmiR-AN0175-SN-10) (all
purchased from Guangzhou Fulengen Co., Ltd., Guangzhou, China) or
co-transfected with miR-137 mimic and pcDNA3.1-enhancer of zeste 2
polycomb repressive complex 2 subunit (EZH2) expression plasmid
(Yearthbio, Changsha, China) using Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from 1×107 cells
(THLE-3 cells and the liver cancer cell lines) using TRIzol Reagent
(Thermo Fisher Scientific, Inc.). Total RNA was then converted into
cDNA using PrimeScript First Strand cDNA Synthesis kit (Takara Bio,
Inc., Otsu, Japan). For miR expression, RT-qPCR was performed using
miRNA Q-PCR Detection kit (GeneCopoeia, Inc., Rockville, MD, USA)
according to the manufacturer's instructions on an ABI 7500
thermocycler (Thermo Fisher Scientific, Inc.). U6 was used as a
reference gene internal control for miR expression. U6 (HmiRQP9001)
and miR-137 (HmiRQP0175) primers were purchased from Guangzhou
Fulengen Co., Ltd. For mRNA expression detection, RT-qPCR was
performed using SYBR-Green I Real-Time PCR kit (Biomics
Biotechnologies Co., Ltd., Nantong, China). The primer sequences
for EZH2 were as follows: Forward, 5′-AATCAGAGTACATGCGACTGAGA-3′
and reverse, 5′-GCTGTATCCTTCGCTGTTTCC-3′. The primer sequences for
GAPDH were as follows: Forward, 5′-CTGGGCTACACTGAGCACC-3′ and
reverse, 5′-AAGTGGTCGTTGAGGGCAATG-3′. The PCR reaction system was
0.33 µl cDNA solution, 10 µl SYBR-Green I mix, 2 µl primers and
7.67 µl H2O were mixed to obtain a final reaction volume
of 20 µl. The reaction condition were 95°C for 5 min, followed by
40 cycles of denaturation at 95°C for 15 sec and
annealing/elongation step at 60°C for 30 sec. The relative
expression was analyzed by the 2−ΔΔCq method (16).
Western blot analysis
Cells (1×107) were lysed with ice-cold
radioimmunoprecipitation assay lysis buffer (Beyotime Institute of
Biotechnology, Haimen, China). Proteins were collected, quantified
using a bicinchoninic acid assay and separated (50 µg/lane) by 10%
SDS-PAGE (Beyotime Institute of Biotechnology). Proteins were
transferred onto polyvinylidene difluoride (PVDF) membrane (Thermo
Fisher Scientific, Inc.). The PVDF membrane was incubated with
phosphate buffered saline (PBS; Thermo Fisher Scientific, Inc.)
containing 5% non-fat milk overnight at 4°C, and then incubated
with the polyclonal mouse anti-human EZH2 antibody (1:100; cat. no.
ab168764; Abcam, Cambridge, MA, USA) and polyclonal mouse
anti-human GAPDH antibody (1:50; cat. no. ab8245; Abcam) at room
temperature for 3 h. After washing with PBS-Tween three times, the
membrane was incubated with goat polyclonal anti-mouse
HRP-conjugated secondary antibody (1:5,000; cat. no. ab97040;
Abcam) at room temperature for 1 h. The enhanced chemiluminescence
system (Thermo Fisher Scientific, Inc.) was used to detect the
immunoreactive bands. The protein expression was measured using
Image Pro Plus software (Media Cybernetics, Inc., Rockville, MD,
USA). GAPDH was used as the internal reference.
Cell proliferation analysis
HepG2 cell suspension (5×104 cells/well)
was plated in a 96-well plate, and cultured for 12, 24, 48 or 96 h.
Subsequently, MTT (10 µl, 5 mg/ml) was added, and cells were then
incubated at 37°C for 4 h. The supernatant was removed, and 100 µl
dimethyl sulfoxide was added into each well. The absorbance at 570
nm was determined using the Model 680 Microplate Reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA).
Cell migration analysis
HepG2 cells were cultured to confluence, and a wound
was created with a plastic scriber. Then, cells were washed and
incubated in serum-free DMEM at 37°C for 24 h. Subsequently, the
medium was replaced with DMEM with 10% FBS, and then cultured at
37°C for 24 h. Cells were observed under a Nikon microscope (Nikon
Corporation, Tokyo, Japan).
Cell invasion analysis
A 24-well Transwell chamber (Chemicon; Merck KGaA,
Darmstadt, Germany) with a layer of Matrigel was used to perform
Transwell assays. A HepG2 cell suspension (5×104 cells)
was added in the upper chamber, and DMEM containing 10% FBS was
added into the lower chamber. After incubation for 24 h,
non-invading cells on the interior of the inserts were removed
using a cotton-tipped swab. Cells on the lower surface of the
membrane was stained with 0.5% gentian violet at room temperature
for 10 min, and then rinsed by water, and air-dried. Invading cells
were counted under a Nikon microscope.
Luciferase reporter assay
Targetscan 7.0 (www.targetscan.org) was used to predicate the putative
targets of miR-137, according to the manufacturer's instructions.
According to the manufacturer's instructions, the QuickChange
Site-Directed Mutagenesis kit (Stratagene; Agilent Technologies,
Inc., Santa Clara, CA, USA) was used to construct the mutant type
(MT) EZH2 3′ untranslated region (3′UTR) lacking complementarity
with the miR-137 seed sequence. The wild-type (WT) or MT of EZH2
3′UTR was cloned into the downstream of the firefly
luciferase-coding region of pMIR-GLO™ Luciferase vector (Promega
Corporation, Madison, WI, USA). The cloning procedure was performed
by Yearthbio. HepG2 cells were co-transfected using Lipofectamine
2000 with the WT- or MT-EZH2-3′UTR luciferase reporter plasmid, and
miR-NC or miR-137 mimic, respectively. The luciferase activity was
detected after transfection for 48 h using the Dual Luciferase
Reporter Assay system (Promega Corporation), according to the
manufacturer's instruction.
Statistical analysis
Data are expressed as the mean ± standard deviation
of three independent experiments. SPSS 19.0 (IBM Corp., Armonk, NY,
USA) was used to perform statistical analysis. The association
between miR-137 expression and clinical characteristics in HCC were
analyzed using χ2 test. The Kaplan-Meier estimator was
used for survival analysis. The difference between two groups was
analyzed using Student's t-test. The difference among more than two
groups was analyzed using analysis of variance and Tukey post hoc
test. Pearson correlation analysis was used to analyze the
correlation between miR-137 and EZH2 mRNA levels in HCC tissues.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Downregulation of miR-137 is
associated with malignant progression and poor prognosis in
HCC
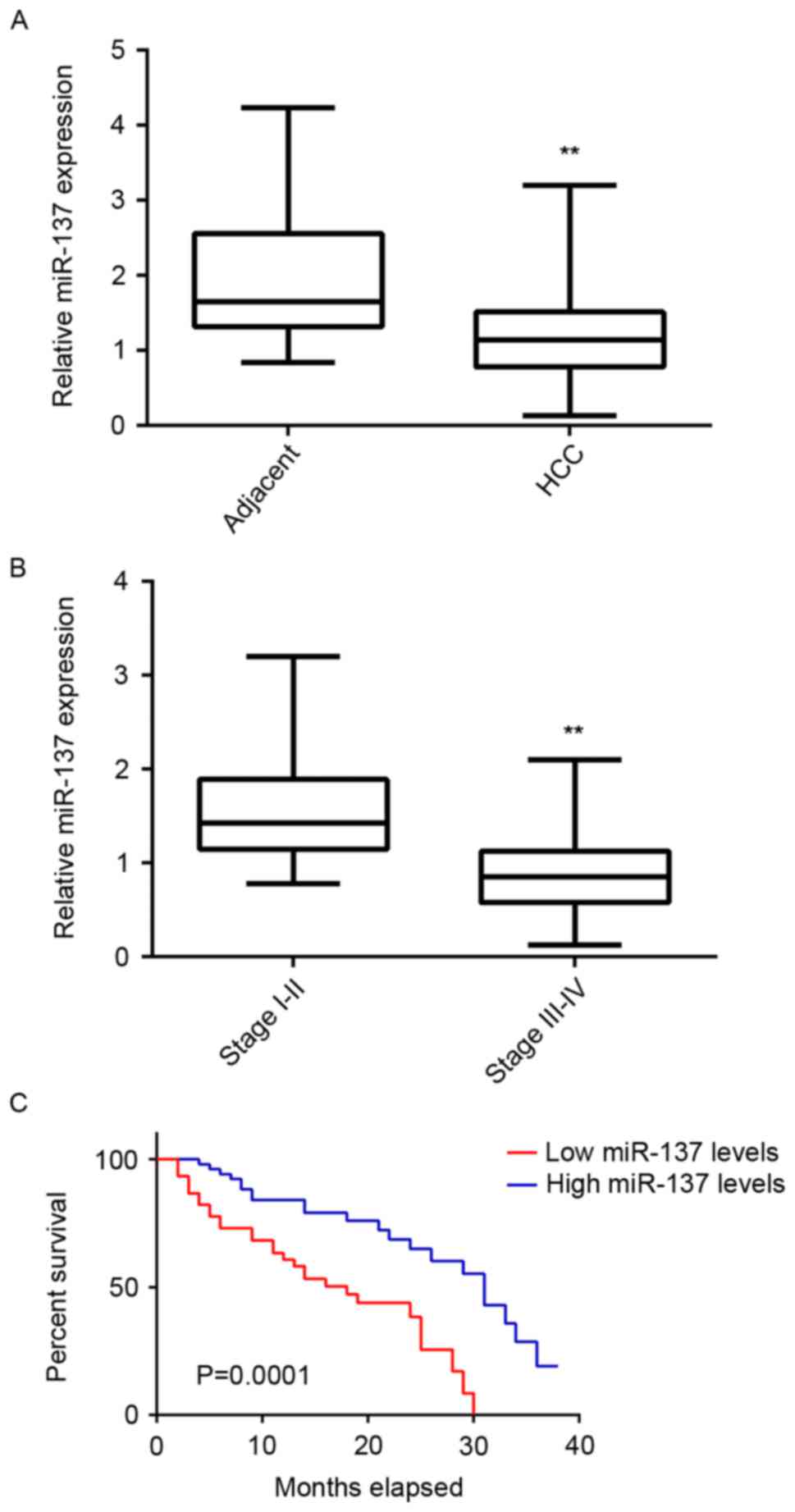
To study the regulatory mechanism of miR-137 in HCC,
its expression levels were initially examined in HCC tissues and
adjacent non-tumor tissues. RT-qPCR data demonstrated that miR-137
was significantly downregulated in HCC tissues compared with
adjacent non-tumor tissues (Fig.
1A). Additionally, the expression of miR-137 was further lower
in HCC at high stage (III–IV) compared with that in HCC at low
stage (I–II; Fig. 1B). These
findings suggest that the decreased expression of miR-137 may
contribute to the malignant progression of HCC. To further confirm
these findings, the association between the miR-137 expression and
clinical characteristics in HCC was examined. Using the mean value
of miR-137 expression as a cutoff, the HCC patients were divided
into a high miR-137 expression group and low miR-137 expression
group. Low expression of miR-137 was significantly associated with
vein invasion, lymph node metastasis and advanced clinical stage in
HCC, but not associated with age, sex, tumor number, histologic
grade, or HPV infection (Table I).
Taken these findings together, we suggest that downregulation of
miR-137 is involved in HCC progression. Subsequently, the
association between miR-137 expression and overall survival time of
patients with HCC was analyzed. As presented in Fig. 1C, the patients with HCC with low
expression of miR-137 had shorter survival time compared with those
with high miR-137 levels. These findings suggest that
downregulation of miR-137 is associated with poor prognosis in
HCC.
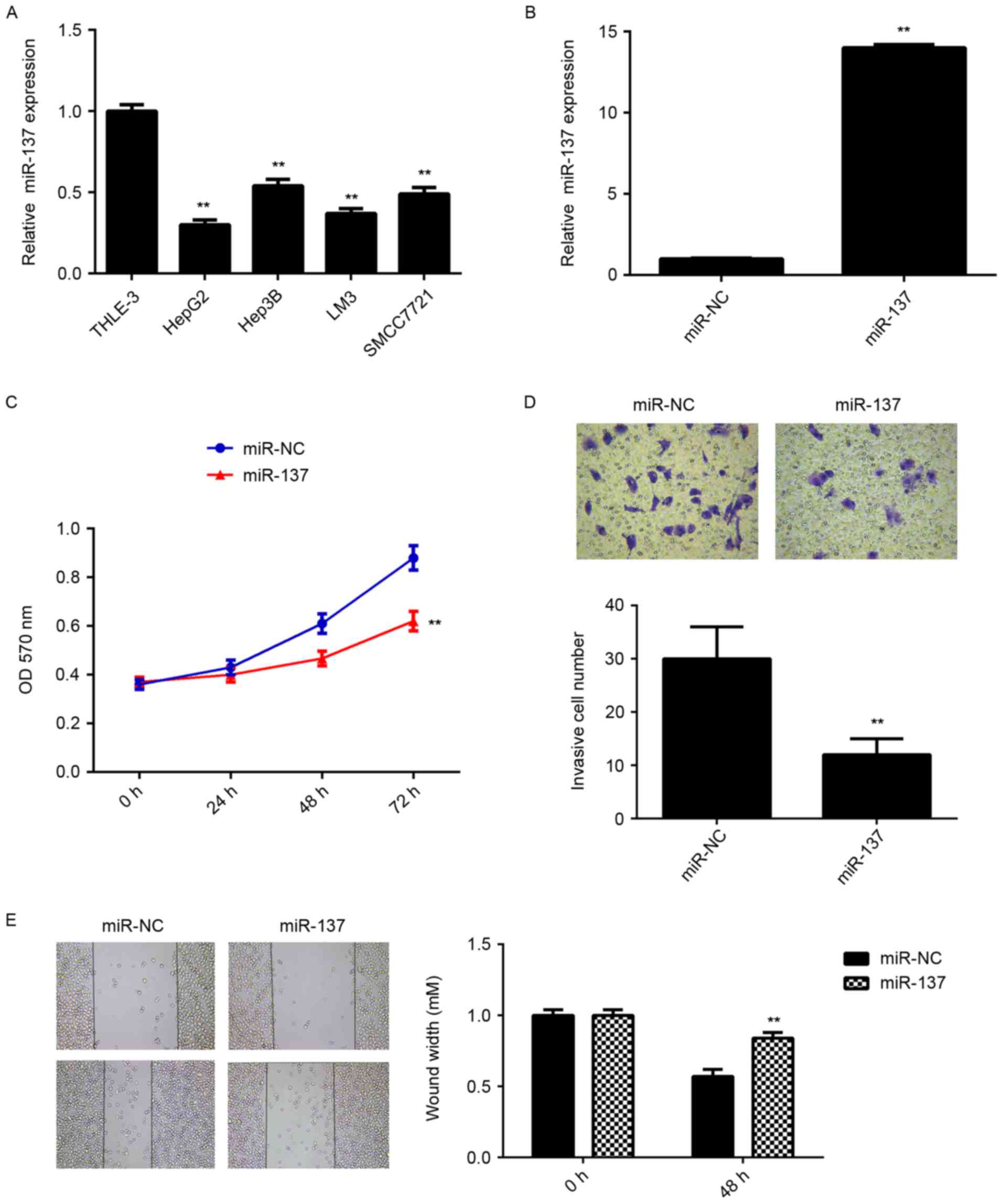
miR-137 inhibits HCC cell
proliferation and invasion
To further investigate the underlying mechanism,
in vitro experiments using liver cancer cell lines were
performed. RT-qPCR data showed that the miR-137 levels were
significantly reduced in HepG2, Hep3B, LM3 and SMCC7721 cell lines
compared with THLE-3 normal liver epithelial cells (Fig. 2A). HepG2 cells were then
transfected with miR-137 mimic or miR-NC as the control group.
Following transfection, the miR-137 levels were examined using
RT-qPCR. As presented in Fig. 2B,
the miR-137 levels were significantly increased in HepG2 cells
transfected with miR-137 mimic compared with the miR-NC group.
Further studies demonstrated that overexpression of miR-137 caused
a significant reduction in cell proliferation, migration and
invasion compared with the miR-NC group (Fig. 2C-E). These data suggest that
miR-137 has a suppressive role in HCC growth and metastasis.
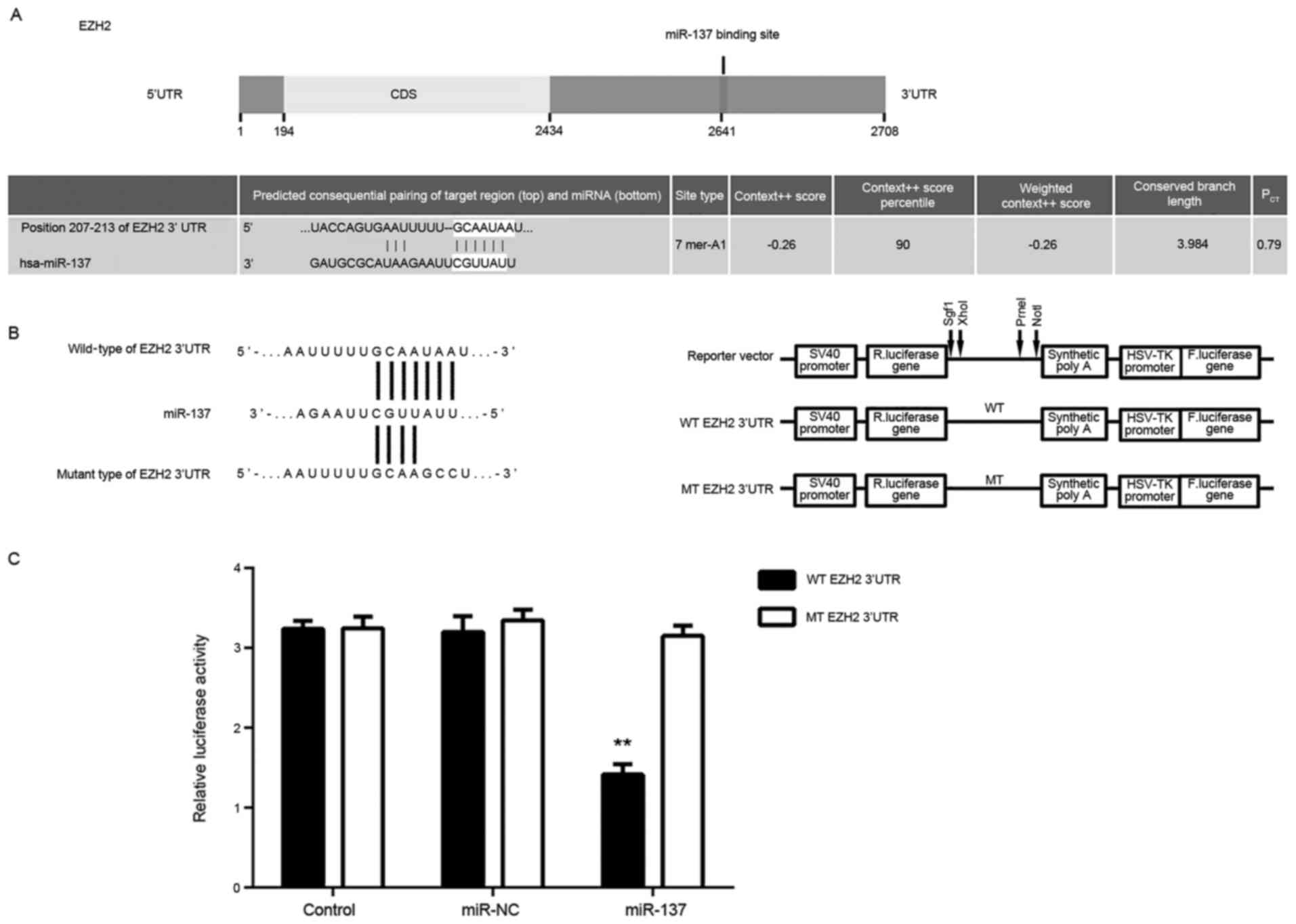
EZH2 is a direct target of miR-137 in
HepG2 cells
Targetscan software was used to predict that EZH2
was a putative target of miR-137 (Fig.
3A). To confirm their targeting relationship, the WT-EZH2-3′UTR
or MT-EZH2-3′UTR luciferase reporter plasmids were generated
(Fig. 3B). HepG2 cells were then
co-transfected with WT- or MT-EZH2-3′UTR luciferase reporter
plasmid, and miR-NC or miR-137 mimic. As demonstrated in Fig. 3C, the luciferase activity was
significantly reduced in cells co-transfected with miR-137 mimic
and WT-EZH2-3′UTR luciferase reporter plasmid when compared with
the control group; the same effect was not observed in cells
co-transfected with miR-137 mimic and MT-EZH2-3′UTR. Accordingly,
these results indicated that miR-137 is a direct target gene of
miR-137 in HepG2 cells.
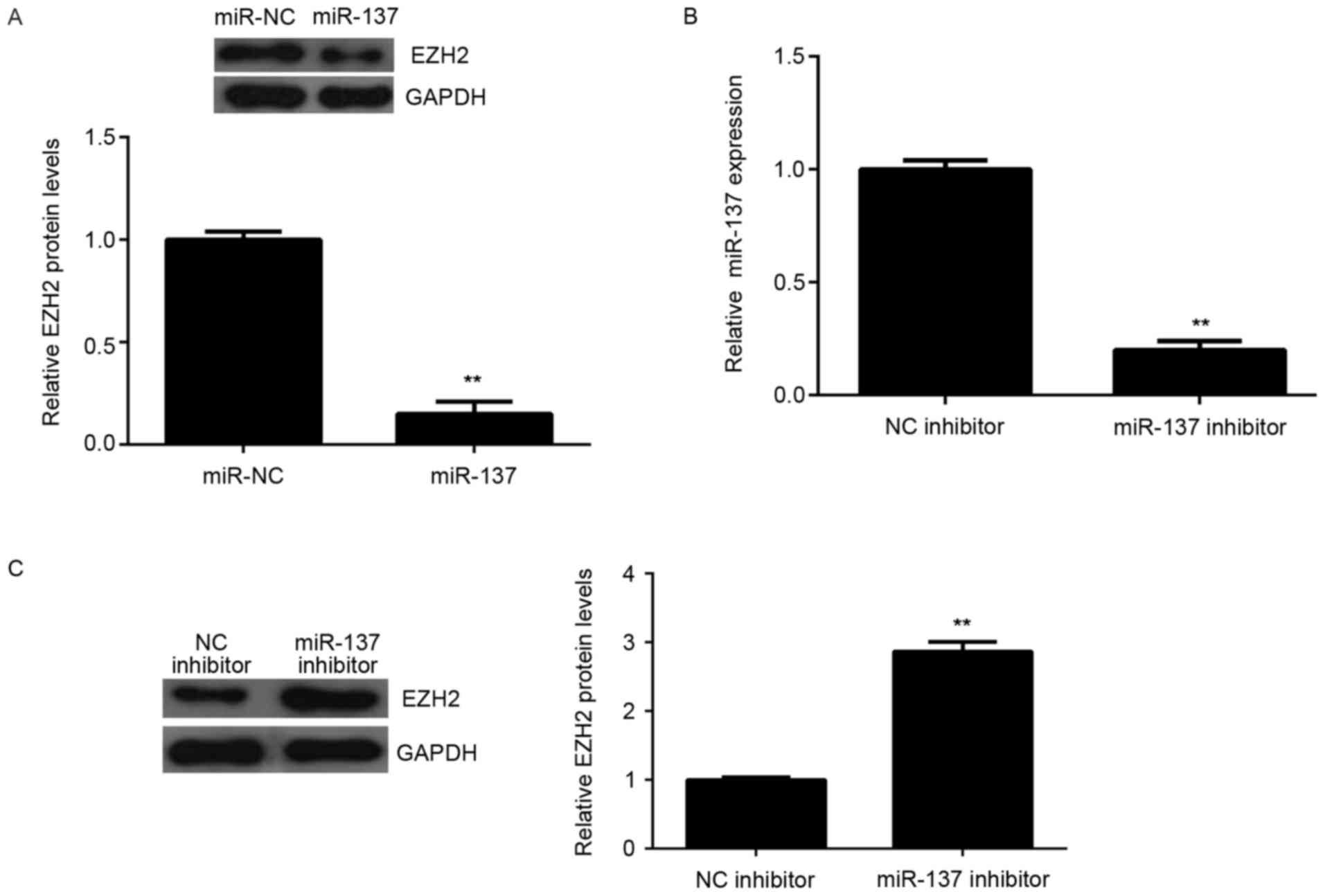
Effects of miR-137 on the protein expression of EZH2
were also determined. The protein levels of EZH2 were significantly
reduced in the miR-137 group compared to miR-NC group, indicating
that overexpression of miR-137 downregulates EZH2 expression
(Fig. 4A). To further confirm
these findings, HepG2 cells were transfected with miR-137
inhibitor, or NC inhibitor as the control group. RT-qPCR data
demonstrated that the miR-137 levels were downregulated in miR-137
inhibitor group compared to NC inhibitor group (Fig. 4B). Additionally, the miR-137
inhibitor significantly increased the protein expression of EZH2 in
HepG2 cells (Fig. 4C).
Accordingly, these results indicated that miR-137 negatively
regulates the protein expression of the target gene, EZH2, in HepG2
cells.
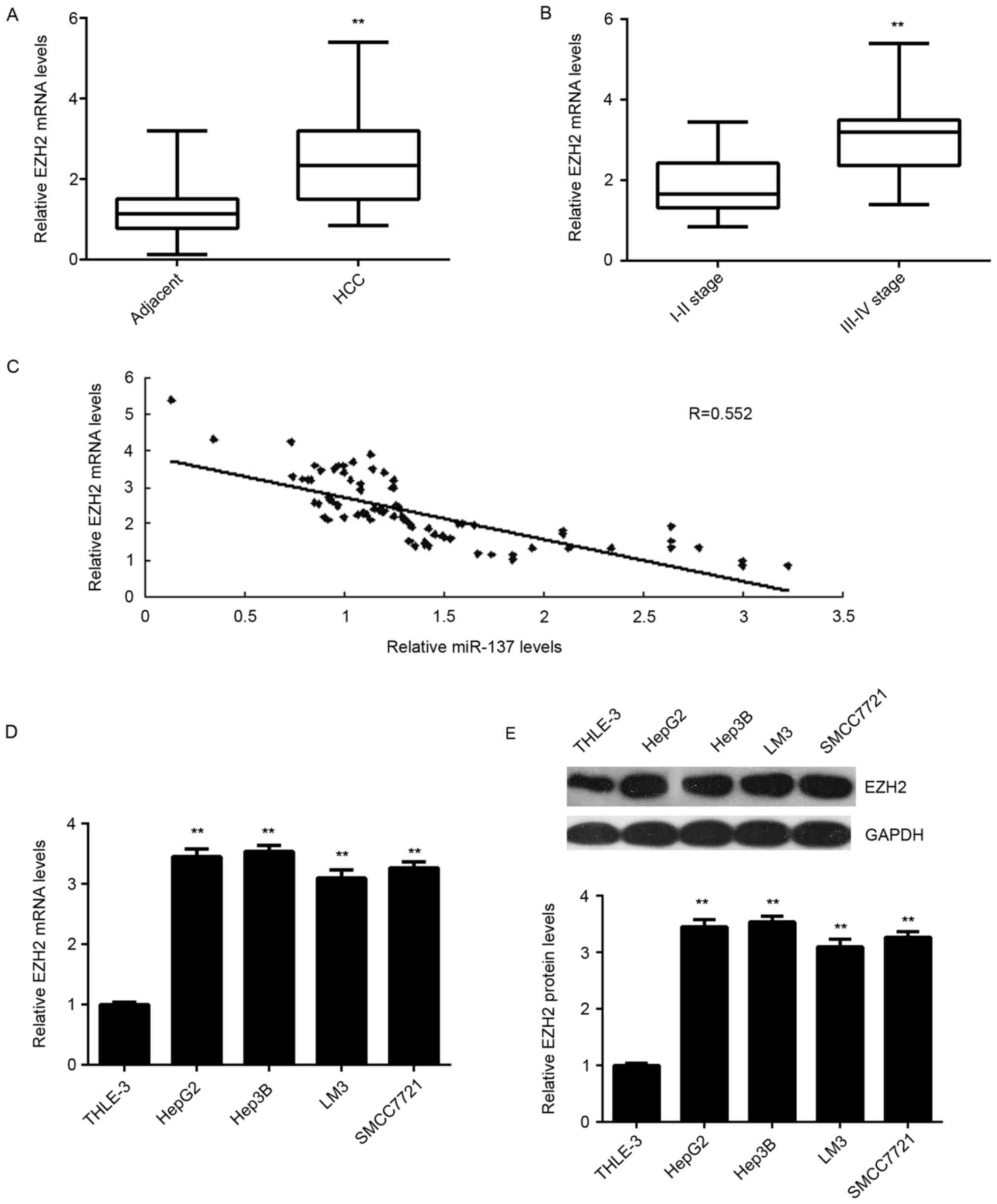
EZH2 is upregulated in HCC and
inversely correlated with the miR-137 levels
To further confirm the targeting relationship
between miR-137 and EZH2, the expression levels of EZH2 in HCC
tissues were examined. As presented in Fig. 5A, RT-qPCR data demonstrated that
the mRNA levels of EZH2 were significantly upregulated in HCC
tissues compared to adjacent non-tumor tissues. Furthermore, its
expression levels were higher in HCC at high stage (III–IV) when
compared with those in HCC at low stage (I–II; Fig. 5B). Further investigation
demonstrated an inverse correlation between the miR-137 and EZH2
mRNA levels in HCC tissues (Fig.
5C). Therefore, these data further support that downregulation
of miR-137 causes the upregulation of its target gene EZH2 in HCC.
In addition, the mRNA and protein levels of EZH2 were also
upregulated in the liver cell lines compared with THLE-3 cells
(Fig. 5D-E).
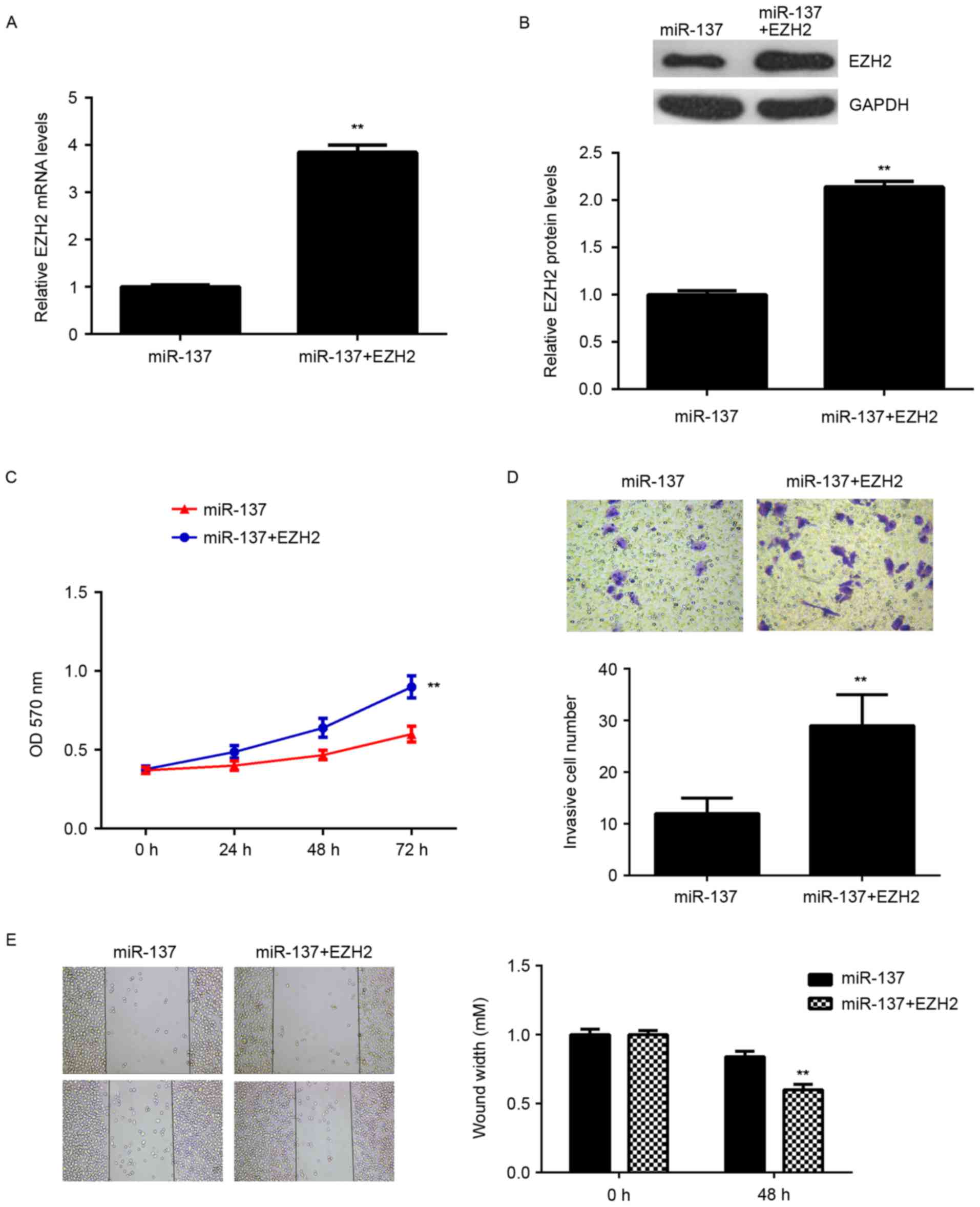
Overexpression of EZH2 eliminates the
inhibitory effects of miR-137 on the malignant phenotypes of HepG2
cells
Subsequently, the role of EZH2 in the
miR-137-mediated malignant phenotypes of HCC cells was
investigated. HepG2 cells were transfected with miR-137 mimic, or
co-transfected with miR-137 mimic and pcDNA3.1-EZH2 ORF plasmid.
RT-qPCR and western blot data indicated that the mRNA and protein
levels of EZH2 were significantly upregulated in cells
co-transfected with miR-137 mimic and pcDNA3.1-EZH2 ORF plasmid,
when compared with those in the miR-137 group (Fig. 6A and B). Cell proliferation,
migration and invasion were also examined. As indicated in Fig. 6C-E, the proliferation, invasion and
migration of HepG2 cells were significantly increased in the
miR-137 + EZH2 group compared with the miR-137 group. Therefore,
overexpression of EZH2 may reverse some of the inhibitory effects
of miR-137 on the malignant phenotypes of HepG2 cells. These
findings suggested that miR-137 inhibits cell proliferation,
migration and invasion, at least partially, by directly targeting
EZH2.
Discussion
The regulatory mechanism of miR-137 underlying HCC
growth and metastasis remains largely unclear. In the present
study, it was demonstrated that miR-137 was significantly
downregulated in HCC tissues and several liver cancer cell lines.
Low expression of miR-137 was significantly associated with vein
invasion, lymph node metastasis, advanced clinical stage and as
poor prognosis. Overexpression of miR-137 led to a significant
reduction in HepG2 cell proliferation, migration and invasion.
EZH2, significantly upregulated in HCC, was further identified as a
direct target gene of miR-137, and the protein expression of EZH2
was negatively regulated by miR-137 in HepG2 cells. Furthermore,
overexpression of EZH2 eliminated the inhibitory effects of miR-137
on the malignant phenotypes of HepG2 cells.
miR-137 has been reported to function as a tumor
suppressor in various common human cancers, including colorectal
cancer (17), glioblastoma
(18), gastric cancer (19), multiple myeloma (20), lung cancer (11) and papillary thyroid carcinoma
(21). Recently, miR-137 was
reported to have a suppressive role in HCC (15). Liu et al (14) reported that miR-137 was
significantly downregulated in HCC, and its decreased expression is
associated with vein invasion, incomplete involucrum, and distant
metastasis. In the present study, miR-137 was significantly
decreased in HCC tissues compared with adjacent non-tumor tissues.
Additionally, the expression of miR-137 was further lower in HCC at
high stage (III–IV) compared with that in HCC at low stage (I–II).
Furthermore, low expression of miR-137 was significantly associated
with vein invasion, lymph node metastasis, and advanced clinical
stage in HCC, but not associated with age, sex, tumor number, grade
or HPV infection. Additionally, Liu et al (14) used multivariate analysis to show
that miR-137 is an independent indicator for poor survival, and
analysis in the current study indicated that patients with HCC with
low expression of miR-137 had shorter survival time compared with
those with high miR-137 levels. Data from the present study and
others suggest that downregulation of miR-137 may contribute to the
malignant progression of HCC.
Furthermore, in the current study miR-137 was
downregulated in liver cancer cell lines compared with normal liver
cells, and overexpression of miR-137 significantly decreased the
proliferation, migration and invasion of HepG2 cells. Liu et
al (14) also reported that
miR-137 overexpression suppressed cell proliferation, migration and
invasion in vitro using HCC cell lines, including SK-Hep1
and QGY-7703. Additionally, Gao et al (15) used HuH7 and MHCC97 L HCC cell
lines, and reported that upregulation of miR-137 inhibited cell
proliferation and metastasis in vitro and tumor growth in
vivo. These findings suggest that miR-137 has an inhibitory
role in HCC growth and metastasis.
EZH2, a histone methyltransferase, is the catalytic
subunit of polycomb repressive complex 2, a highly conserved
protein complex that regulates gene expression by methylating
lysine 27 on histone H3 (22).
EZH2 has been demonstrated to act as an oncogene in certain human
cancers, including HCC (23–25).
Sudo et al (25) reported
that EZH2 was significantly upregulated in HCC tissues compared
with corresponding non-tumor specimens. In the present study, EZH2
was significantly upregulated in HCC tissues and liver cancer cell
lines, and its expression levels were higher in HCC at high stage
(III–IV) compared low stage (I–II). Furthermore, Au et al
(26) reported that the
upregulation of EZH2 was significantly associated with venous
invasion, direct liver invasion and absence of tumor encapsulation,
and that EZH2 epigenetically silenced multiple tumor suppressor
miRs, including miR-139-5p, miR-125b, miR-101, let-7c, and
miR-200b, to promote liver cancer metastasis (26). In the present study, EZH2 was
identified as a direct target gene of miR-137, and the protein
expression of EZH2 was demonstrated to be negatively regulated by
miR-137 in HepG2 cells. Additionally, an inverse correlation
between the miR-137 and EZH2 expression levels in HCC tissues was
observed. Therefore, the upregulation of EZH2 in HCC may be due to
the downregulation of miR-137. Overexpression of EZH2 eliminated
the suppressive effects of miR-137 on the proliferation, migration
and invasion of HepG2 cells, which supports that the suppressive
role of miR-137 in regulating the malignant phenotypes of HCC cells
was via direct targeting of EZH2. In fact, the targeting
association between miR-137 and EZH2 was also identified in several
other cancers, including glioblastoma, neuroblastoma and melanoma
(27–29). Luo et al (29) reported that miR-137 inhibited the
invasion of melanoma cells through downregulation of multiple
oncogenic target genes, including EZH2. Therefore, the results of
the current study expand on the importance of the miR-137/EZH2 in
human cancer.
In summary, the present study demonstrated that
miR-137 is downregulated in HCC, and inhibited HepG2 cell
proliferation, migration and invasion, at least partially, through
directly targeting EZH2 oncogene. Therefore, the findings highlight
the importance of miR-137/EZH2 axis in liver cancer
progression.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moss EG: MicroRNAs: Hidden in the genome.
Curr Biol. 12:R138–R140. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Croce CM and Calin GA: miRNAs, cancer, and
stem cell division. Cell. 122:6–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Meng F, Henson R, Wehbe-Janek H, Ghoshal
K, Jacob ST and Patel T: MicroRNA-21 regulates expression of the
PTEN tumor suppressor gene in human hepatocellular cancer.
Gastroenterology. 133:647–658. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li N, Fu H, Tie Y, Hu Z, Kong W, Wu Y and
Zheng X: miR-34a inhibits migration and invasion by down-regulation
of c-Met expression in human hepatocellular carcinoma cells. Cancer
Lett. 275:44–53. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang R, Deng L, Zhao L, Li X, Zhang F,
Xia Y, Gao Y, Wang X and Sun B: miR-22 promotes HBV-related
hepatocellular carcinoma development in males. Clin Cancer Res.
17:5593–5603. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dou C, Wang Y, Li C, Liu Z, Jia Y, Li Q,
Yang W, Yao Y, Liu Q and Tu K: MicroRNA-212 suppresses tumor growth
of human hepatocellular carcinoma by targeting FOXA1. Oncotarget.
6:13216–13228. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang CY, Zhang JJ, Hua L, Yao KH, Chen JT
and Ren XQ: MicroRNA-98 suppresses cell proliferation, migration
and invasion by targeting collagen triple helix repeat containing 1
in hepatocellular carcinoma. Mol Med Rep. 13:2639–2644. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shen H, Wang L, Ge X, Jiang CF, Shi ZM, Li
DM, Liu WT, Yu X and Shu YQ: MicroRNA-137 inhibits tumor growth and
sensitizes chemosensitivity to paclitaxel and cisplatin in lung
cancer. Oncotarget. 7:20728–20742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang ML, Hsieh TH, Ng KH, Tsai YN, Tsai
CF, Chao ME, Liu DJ, Chu SS, Chen W, Liu YR, et al: Downregulation
of miR-137 and miR-6500-3p promotes cell proliferation in pediatric
high-grade gliomas. Oncotarget. 7:19723–19737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu DC, Zhang MF, Su SG, Fang HY, Wang XH,
He D, Xie YY and Liu XH: HEY2, a target of miR-137, indicates poor
outcomes and promotes cell proliferation and migration in
hepatocellular carcinoma. Oncotarget. 7:38052–38063. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu LL, Lu SX, Li M, Li LZ, Fu J, Hu W,
Yang YZ, Luo RZ, Zhang CZ and Yun JP: FoxD3-regulated microRNA-137
suppresses tumour growth and metastasis in human hepatocellular
carcinoma by targeting AKT2. Oncotarget. 5:5113–5124. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao M, Liu L, Li S, Zhang X, Chang Z and
Zhang M: Inhibition of cell proliferation and metastasis of human
hepatocellular carcinoma by miR-137 is regulated by CDC42. Oncol
Rep. 34:2523–2532. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen DL, Wang DS, Wu WJ, Zeng ZL, Luo HY,
Qiu MZ, Ren C, Zhang DS, Wang ZQ, Wang FH, et al: Overexpression of
paxillin induced by miR-137 suppression promotes tumor progression
and metastasis in colorectal cancer. Carcinogenesis. 34:803–811.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bier A, Giladi N, Kronfeld N, Lee HK,
Cazacu S, Finniss S, Xiang C, Poisson L, deCarvalho AC, Slavin S,
et al: MicroRNA-137 is downregulated in glioblastoma and inhibits
the stemness of glioma stem cells by targeting RTVP-1. Oncotarget.
4:665–676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deng J, Lei W, Xiang X, Zhang L, Lei J,
Gong Y, Song M, Wang Y, Fang Z, Yu F, et al: Cullin 4A (CUL4A), a
direct target of miR-9 and miR-137, promotes gastric cancer
proliferation and invasion by regulating the Hippo signaling
pathway. Oncotarget. 7:10037–10050. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang B, Ma L, Wei J, Hu J, Zhao Z, Wang
Y, Chen Y and Zhao F: miR-137 suppresses the phosphorylation of AKT
and improves the dexamethasone sensitivity in multiple myeloma
cells via targeting MITF. Curr Cancer Drug Targets. 16:807–817.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dong S, Jin M, Li Y, Ren P and Liu J:
miR-137 acts as a tumor suppressor in papillary thyroid carcinoma
by targeting CXCL12. Oncol Rep. 35:2151–2158. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Italiano A: Role of the EZH2 histone
methyltransferase as a therapeutic target in cancer. Pharmacol
Ther. 165:26–31. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhong J, Min L, Huang H, Li L, Li D, Li J,
Ma Z and Dai L: EZH2 regulates the expression of p16 in the
nasopharyngeal cancer cells. Technol Cancer Res Treat. 12:269–274.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang K, Zhang Y, Ren K, Zhao G, Yan K and
Ma B: MicroRNA-101 inhibits the metastasis of osteosarcoma cells by
downregulation of EZH2 expression. Oncol Rep. 32:2143–2149. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sudo T, Utsunomiya T, Mimori K, Nagahara
H, Ogawa K, Inoue H, Wakiyama S, Fujita H, Shirouzu K and Mori M:
Clinicopathological significance of EZH2 mRNA expression in
patients with hepatocellular carcinoma. Br J Cancer. 92:1754–1758.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Au SL, Wong CC, Lee JM, Fan DN, Tsang FH,
Ng IO and Wong CM: Enhancer of zeste homolog 2 epigenetically
silences multiple tumor suppressor microRNAs to promote liver
cancer metastasis. Hepatology. 56:622–631. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sun J, Zheng G, Gu Z and Guo Z: MiR-137
inhibits proliferation and angiogenesis of human glioblastoma cells
by targeting EZH2. J Neurooncol. 122:481–489. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ren X, Bai X, Zhang X, Li Z, Tang L, Zhao
X, Li Z, Ren Y, Wei S, Wang Q, et al: Quantitative nuclear
proteomics identifies that miR-137-mediated EZH2 reduction
regulates resveratrol-induced apoptosis of neuroblastoma cells. Mol
Cell Proteomics. 14:316–328. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luo C, Tetteh PW, Merz PR, Dickes E,
Abukiwan A, Hotz-Wagenblatt A, Holland-Cunz S, Sinnberg T, Schittek
B, Schadendorf D, et al: miR-137 inhibits the invasion of melanoma
cells through downregulation of multiple oncogenic target genes. J
Invest Dermatol. 133:768–775. 2013. View Article : Google Scholar : PubMed/NCBI
|