Introduction
The neuropathic pain (NP) was newly defined as pain
induced by a disease or lesion of the somatosensory system by
International Association for the Study of Pain (IASP) (1). Due to the unsatisfactory therapeutic
regimens for NP, a multitude of affected patients cannot alleviate
their pain (2). Hence,
understanding of pathogenesis underlying NP and quest for novel
effective drugs for the therapeutics of NP is essential.
Chronic constriction injury (CCI) models have both
peripheral nerve injury and inflammatory components which can mimic
NP in humans and are effective for investing the mechanisms
underlying NP (3,4).
Herein, the effect of a safe and widely used oral
hypoglycemic drug, metformin, was invested on NP. Metformin may
restrain peripheral nerve injury-induced NP (5), and it also may potentially decrease
lumbar radiculopathy pain in humans (6). However, the mechanism of the
inhibitive effects of metformin on NP has not been investigated
in vivo in previous studies. We innovatively explored the
effects of metformin on the spinal cords of rats in a CCI-induced
NP model.
Adenosine monophosphate-activated protein kinase
(AMPK) is a serine/threonine protein kinase that is central to
cellular metabolic processes and energy balance maintenance,
inflammation, cancer, neurodegenerative diseases (7–9).
Studies have also shown that AMPK plays a key role in pain
(10,11). Moreover, metformin is validated to
be an indirect activator of AMPK (12,13).
Therefore, we speculated that AMPK might be involved in the
analgesic effects of metformin.
Metformin reportedly inhibits signal transducer and
activator of transcription 3 (STAT3) in tumor cells (14,15).
In addition, the STAT3 pathway is activated in the spinal dorsal
horn (SDH) in both NP (16–18)
and spinal cord injury models (19). However, whether the STAT3 pathway
is involved in the analgesic effe-cts of metformin on NP remained
unclear. Here, we evaluated the analgesic effects of metformin and
its potential effects on the STAT3 signaling pathway in the SDH in
rodent models of NP.
The activation of spinal glial cells plays an
crucial role in NP (20,21). Further, AMPK and STAT3 are
implicated in the regulation of the activation of glial cells
(22,23). We also investigated the effect of
metformin on microglia and astrocytes in the CCI models.
Collectively, our data reveal a potential mechanism underlying the
analgesic effect of metformin and may shed new light on the
therapeutics for NP.
Materials and methods
Animals
Male Sprague-Dawley rats (weighing 200–230 g, aged
6–8 weeks) were obtained from the experimental animal center of
Xuzhou Medical University (Xuzhou, China). The animals were housed
under the following controlled conditions: A 12 h light-dark cycle
(lights on from 6 a.m. to 6 p.m.) at a steady temperature of 25°C ±
1°C with free access to water and food. All protocols were approved
by the Institutional Animal Care and Use Committee at Xuzhou
Medical University. The experimental procedures were conducted
according to the ethical guidelines for the care and use of
laboratory animals of the National Institutes of Health (NIH
Publications, no. 8023, revised 1978) and the IASP. Effort was made
to decrease the number of animals used and to reduce animal
suffering.
CCI of the sciatic nerve
The CCI rat model was established complied to a
previously described method (24).
The rats were anesthetized using 10% chloral hydrate (0.3 ml/100 g,
i.p.). The left sciatic nerve was exposed using a small incision at
the mid-thigh level. A constriction injury was induced proximal to
the trifurcation of the sciatic nerve by making loose ligations
with four 4-0 silk threads (1 mm apart). In the sham-operated
group, the nerve was exposed without ligations. The wound was then
treated using antibiotics (penicillin) and closed
layer-by-layer.
Experimental groups
A total of 112 rats were used across all
experiments. Behavioral assessment and fasting blood glucose test
consists of two parts. Part I: The rats were randomly divided into
the 4 groups (n=8): Sham + normal saline (sham + NS) group, sham +
metformin group, CCI + NS group, and CCI + metformin group. The
rats received i.p. injections of 5% metformin in NS (200 mg/kg,
Sigma, USA) or an equal dose of NS once per day for 6 consecutive
days beginning on the 5th day after surgery. The dose of metformin
was selected according to a previous reference (5). Behavioral assessment was performed on
days-1 (the day before surgery), and 3, 5, 7, 10, 14 after surgery.
The data of-1th day were detected as baseline. On the 5th day after
surgery, the behavioral assessment was performed before metformin
and saline administered, whereas on the 7th and 10th days, the
behavioral assessment was performed two h after the injections.
Fasting blood glucose levels were measured after the behavioral
assessment on days −1, 3, 5, 7, 10, 14. The blood was taken from
the tail vein. Part II: The rats were randomly divided into 5
groups (n=8): CCI group, CCI + NS group, CCI + metformin group, CCI
+ compound C (AMPK inhibitor) group, and CCI + metformin + compound
C group. The rats received intraperitoneal injections of metformin
or an equal dose of NS from 5th day to 10th day according to the
same method mentioned above. Compound C (30 ug) was intrathecal
administration on 10th day in CCI + compound C group and CCI +
metformin + compound C group. Behavioral assessment was performed
on days −1, 3, 5, 7, 10, 14 after surgery.
Western blot analysis and immunofluorescence
staining. The rats were randomly divided into sham + NS, sham +
metformin, CCI + NS, and CCI + metformin groups (n=10). In each
group, 4 rats were used for western blot and 6 rats for
immunofluorescence staining. On the 10th day, the rats were deeply
anesthetized, then sacrificed, and the L4-L6 spinal cord segments
were collected for either immunofluorescence staining or western
blot analysis.
Reagents
The major reagents which were employed in this study
were metformin (Sigma-Aldrich, USA), compound C (Sigma-Aldrich,
USA), rabbit polyclonal anti-phosphorylated (p)-AMPK (Cell
Signaling Technology; Bioworld Technology Inc., St. Louis Park, MN,
USA), rabbit polyclonal anti-p-STAT3 (Cell Signaling Technology;
Bioworld Technology Inc.); rabbit polyclonal anti-AMPK, rabbit
polyclonal anti-STAT3, rabbit polyclonal anti-GAPDH (all Bioworld
Technology Inc.), goat polyclonal anti-ionized calcium binding
adaptor molecule 1 (Iba-1; Abcam, Cambridge, UK), mouse polyclonal
anti-glial fiber acidic protein (GFAP; Cell Signaling Technology,
Inc., Danvers, MA, USA), Alexa 488 donkey anti-rabbit IgG, Alexa
546 donkey anti-goat IgG, Alexa 594 donkey anti-mouse IgG (all
Invitrogen, Carlsbad, CA, USA), and HRP-conjugated goat anti-rabbit
secondary antibodies (Beyotime Institute of Biotechnology,
Shanghai, China).
Behavioral assessment
Thermal hyperalgesia was evaluated by measuring paw
withdrawal latency (PWL) in response to heat stimulation. A plantar
heat analgesia meter model 390 (IITC Life Science Inc., Woodland
Hills, CA, USA) was used according to the protocols described by
Hargreaves et al to provide a heat source (25). Each rat was placed in a transparent
acrylic chamber that was placed on a glass plate. Following a
one-hour adaptation period, the middle of the plantar surface of
the left hind paw was exposed to a radiant heat source using a beam
of light. The length of time between the start of the light beam
and the foot lift was recorded as the PWL. To avoid damaging the
tissue, an automatic 25 sec cutoff was set. This thermal stimulus
was repeated five times for each paw at an interval of 5 min. The
test was conducted in a noise-free and temperature-controlled
(25–26°C) room. The intensity of the thermal stimulation was
maintained at the same level throughout the entire study.
Intrathecal injection procedure
Rats were anesthetized with inhalation of isoflurane
and placed in a prone position on the operating table. Midpoint
between the tips of the iliac crest was labeled as the fifth to
sixth lumbar intervertebral space (L5-L6). Intrathecal injection
was performed by a micro syringe, which was inserted into the
subarachnoid space of the spinal cord between the L5 and L6 spinous
processes. Intrathecal injection was confirmed by observation of an
obvious tail flick.
Western blot analysis
The rats were anesthetized with 10% chloral hydrate
(0.3 ml/100 g, i.p.) and killed by decapitation. The dissected
L4-L6 spinal dorsal horn samples were rapidly isolated and frozen
in liquid nitrogen and thereafter placed in long-term storage at
−80°C till used. The tissue samples were homogenized in RIPA buffer
containing protease inhibitors and phosphatase inhibitors and
centrifuged for 15 min (12,000 g, at 4°C), with the supernatant
collected. The protein concentration of the supernatant was
determined by a Bicinchoninic Acid Protein Assay kit (Beyotime
Institute of Biotechnology) according to the protocol described in
the previous study (26). Equal
amount of protein samples were separated by 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis and transferred to
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). After thrice rinsed, the membranes were blocked in 3% bovine
serum albumin for 2 h at room temperature (r/t) and incubated with
the following primary antibodies: Rabbit anti-AMPK (1:300; Bioworld
Technology Inc.), rabbit anti-p-AMPK (1:600; Cell Signaling
Technology), rabbit anti-STAT3 (1:500; Bioworld Technology Inc.),
rabbit anti-p-STAT3 (1:300, Cell Signaling Technology), and rabbit
anti-GAPDH (1:2,000; Bioworld Technology Inc.) for overnight at
4°C. After three washes, the membranes were incubated with
HRP-conjugated goat anti-rabbit secondary antibodies for 1 h
(1:1,000; Beyotime Institute of Biotechnology) at r/t. The bands
were visualized with the use of ECL and X-film. The densities of
the protein band were quantified using Image J software (NIH,
Bethesda, MD, USA).
Immunofluorescence histochemistry
The rats were deeply anesthetized using 10% chloral
hydrate (0.3 ml/100 g, i.p.) and intracardially perfused with 300
ml of NS followed by 300 ml of 4% paraformaldehyde (PFA) in
phosphate buffer (PB; 0.1 M; pH 7.2–7.4; 4°C). The spinal cords
were quickly isolated and post-fixed in 4% PFA for 2 h at 4°C.
Subsequently, the L4-L6 spinal cord samples were equilibrated in a
30% sucrose solution at 4°C until the sample blocks dropped to the
bottom. The tissues were embedded in tissue freezing medium and
sliced at thickness of 35 µm by a cryostat (Leica CM1800;
Heidelberg, Germany). After washing thrice with 1X PBS, selected
sections were blocked in 1X PBS containing 3% Triton-X-100 (PBST)
and 10% donkey serum for 2 h at r/t and incubated with the
following primary antibodies in PBST: rabbit anti-p-AMPK (1:200;
Bioworld Technology Inc.), rabbit anti-p-STAT3 (1:50; Bioworld
Technology Inc.), mouse anti-GFAP (1:400; Cell Signaling
Technology), or goat anti-Iba-1 (1:300; Abcam) for 48 h at 4°C. The
slices were then washed 3 times (5 min each) with PBS and incubated
with Alexa 488 donkey anti-rabbit IgG, Alexa 546 donkey anti-goat
IgG or Alexa 594 donkey anti-mouse IgG (all 1:200; Invitrogen) for
2 h at r/t. Images were captured using a confocal laser microscope
(FV1000; Olympus, Corp., Tokyo, Japan) and analyzed by Image
Pro-Plus 6.0 software (Media Cybernetics, Silver Spring, MD, USA).
Five slices for each spinal cord sample were randomly selected for
statistical analysis.
Statistical analyses
The data were analyzed using GraphPad Prism 5.0
Software (GraphPad Software, La Jolla, CA, United States). All data
are shown as the mean ± SD. The normal distribution and variance
homogeneity were tested. One-way ANOVA was used to analyze the
western blot and immunofluorescence results across the four groups.
Two-way ANOVA was used for the PWLs and fasting blood glucose
values, which included two factors of time and group. If a
significant difference was found, a post hoc Bonferroni
multiple test was used to detect differences within the groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Behavioral outcomes and blood glucose
values
To investigate the influence of metformin on CCI
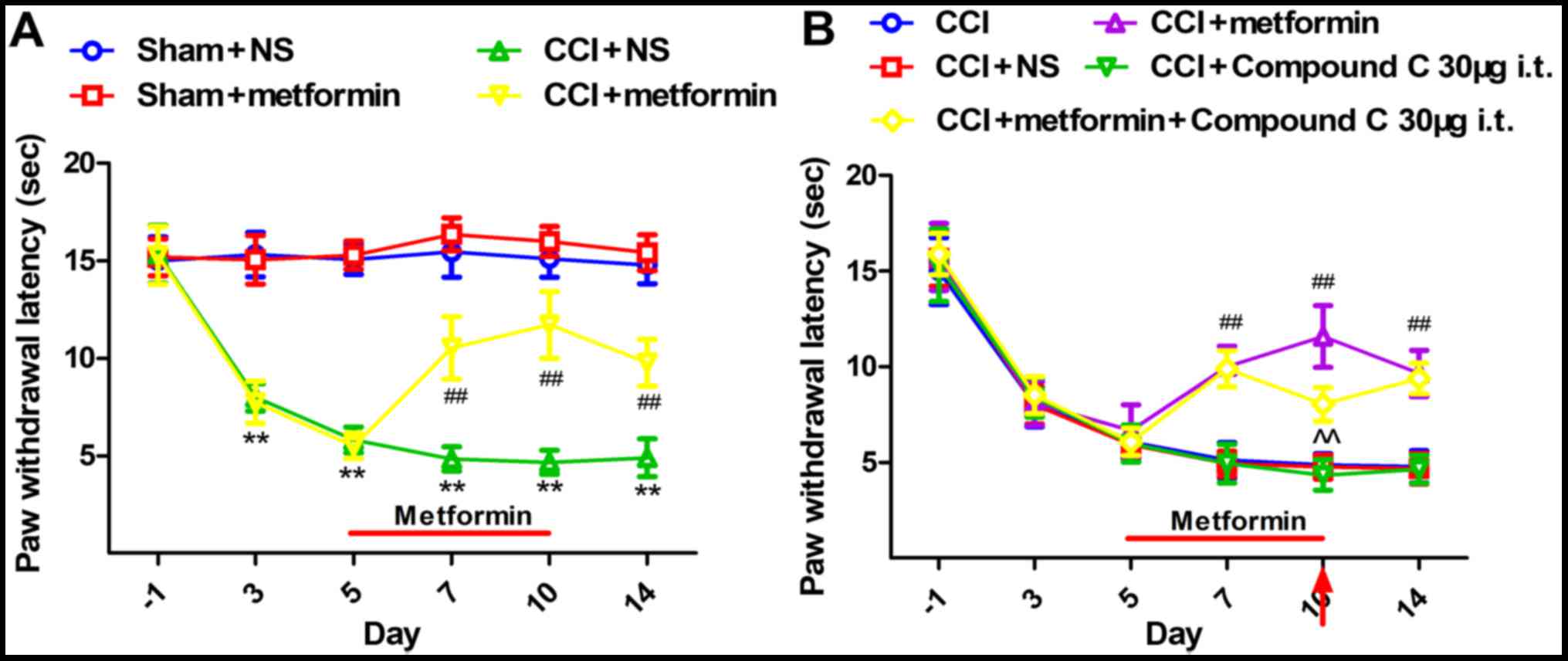
rats, we measured PWLs on days 1, 3, 5, 7, 10, and 14 after surgery
(Fig. 1). There were no
differences in the PWLs between the sham + NS and sham + metformin
groups. On days 3–14, the PWLs in the rats in the CCI + NS group
were significantly lower than those in the sham + NS group.
Meanwhile, the PWLs of the CCI rats that were injected with
metformin (200 mg/kg, once a day in days 5–10) were markedly longer
than those in the CCI + NS group (P<0.01), but shorter than
those in the sham + NS and sham + metformin groups. Even after the
therapy was stopped for 4 days on day 14, the PWLs in the CCI +
metformin group were still considerably higher than those in the
CCI + NS group (P<0.01) (Fig.
1A). These data revealed that the analgesic ability of
metformin was sustained.
Fasting blood glucose values were collected after
the same fasting time in the 4 groups following the behavioral
assessment on days 1, 3, 5, 7, 10 and 14 post-surgery. There was no
difference in blood glucose values among the four groups. metformin
did not affect blood sugar values in the sham-operated rats and CCI
rats (Fig. 2).
We further studied the relationship of metformin and
the NP by AMPK inhibitor (compound C). On the 10th day, the PWLs in
the rats in the CCI + metformin + compound C group were
significantly lower than those in the CCI + metformin group
(P<0.01) (Fig. 1B). We
demonstrated that compound C could reverse the analgesic effect of
metformin.
Activation and distribution of AMPK in
spinal cords after CCI and metformin administration
To further explore how metformin alleviates NP in
CCI rats, we determined p-AMPK and AMPK levels in the spinal dorsal
horns of rats on day 10 after the final metformin or NS treatment
administered. The results of western blot analysis showed that the
expression level of p-AMPK was lower in the CCI + NS group than
that in the sham + NS group (P<0.01) and the expression level of
p-AMPK was significantly higher in the CCI + metformin group than
that in the CCI + NS group (P<0.01). However, the expression
level of AMPK in the SDH was not significantly different among the
four groups of rats (Fig. 3).
Next, we investigated the distribution of p-AMPK in
the SDH by immunofluorescent staining. We found that p-AMPK was
widely distributed in the SDH but mainly concentrated in lamina I
and II. The mean fluorescence intensity and the number of cells
that were positively stained for p-AMPK were significantly lower in
the CCI + NS group than those in the sham + NS group (P<0.01)
and significantly higher in the CCI + metformin group than those in
the CCI + NS group (P<0.01) (Fig.
4).
These data showed that p-AMPK was widely distributed
in the SDH of rats. In the NP model of CCI rats, the expression of
p-AMPK was suppressed, and metformin activated AMPK in the SDH via
phosphorylation, thereby inhibiting the development of NP.
Activation and distribution of STAT3
in SDH after CCI and metformin administration
We further investigated whether metformin inhibited
the activation of STAT3 in the spinal cords of CCI rats by
exploring the expression of p-STAT3 and STAT3 in the dorsal horn.
The expression level of p-STAT3 in the spinal cord was
significantly increased in the CCI + NS group than that in the sham
+ NS group (P<0.01), and the expression level of p-STAT3 was
significantly decresed in the CCI + metformin group than that in
the CCI + NS group (P<0.01), with no apparent difference in the
level of STAT3 in four groups of rats (Fig. 5).
The results of immunofluorescence histochemistry
showed that p-STAT3 was widely distributed, mainly in laminas I and
II of the SDH. The p-STAT3 expression was rarely detected in the
sham + NS rats. However, the number of positively stained cells and
the mean fluorescence intensity for p-STAT3 in the dorsal horn were
higher in the CCI + NS rats than those in the sham + NS rats
(P<0.01). In addition, the values were lower in the CCI +
metformin group than those in the CCI + NS group (P<0.01)
(Fig. 6).
Our results in this NP model indicated that the
STAT3 signaling pathway was activated which demonstrated by the
increased expression of p-STAT3, and this might be one of the
causes of NP. Metformin prevented spinal dorsal horn STAT3
phosphorylation, which might account for its ability to alleviate
NP.
Metformin injection suppressed the
activation of microglia and astrocytes in the SDH
Glial cell activation plays a key role in the
occurrence and maintenance of pain (27). In this study, we further explored
whether metformin affects microglia and astrocytes.
Immunofluorescence staining was performed to detect the expression
of the microglia marker Iba-1 and the astrocyte marker GFAP.
Consistent with previous findings, our results showed that Iba-1
and GFAP are widely distributed in the SDH. Compared with the sham
+ NS group, the mean fluorescence intensity for Iba-1 was higher
and the number of positively stained cells for Iba-1 increased in
the SDH in the CCI + NS group (P<0.01) (Fig. 7A). Similar findings have been found
in astrocytes (Fig. 7B). Moreover,
microglia and astrocytes were morphologically activated after CCI.
And Compared with the CCI + NS group, the activation of astrocytes
and microglia was effectively suppressed by metformin in CCI +
metformin group (P<0.01) (Fig.
7).
Discussion
In the present study, the PWLs of the CCI rats
dropped to a low level from day 3 to day 14. The expression of
p-AMPK was downregulated in the SDH in rats with chronic
constriction injuries, while the expression of p-STAT3 was
upregulated. On day 10 post-surgery, astrocytes and microglia were
widely activated in these rats. However, administration of
metformin (200 mg/kg) over 6 days effectively alleviated the
thermal hyperalgesia and reversed the expression of p-AMPK and
p-STAT3, with the suppression of the activated microglia and
astrocytes simultaneously.
This is the first study to explore the mechanism by
which metformin inhibits NP in the SDH. The SDH is a complex relay
station that transmits nociceptive information and occupies an
important position in NP (28,29).
There are multiple pathways that transmit pain in the SDH,
including the AMPK and STAT3 signaling pathways (11,16–18).
Metformin, an oral anti-diabetic drug, has been
safely applied to treat type 2 diabetes for decades (30) and serves as an indirect agonist of
AMPK. The increase of the cellular AMP/ATP ratio contributes to the
activation of AMPK (9).
Additionally, metformin promotes a shift from aerobic to anaerobic
glycolysis by inhibiting complex I of the mitochondrial respiratory
chain to increase the cellular AMP/ATP ratio (12,13).
In general, the chronic neuroinflammation in the nervous system
contributes to the development of NP (31,32).
AMPK is a negative regulator of the mitogen-activated protein
kinases and the mammalian target of rapamycin (mTOR) pathways,
which are associated with the occurrence of chronic
neuroinflammation (33–36). Further, AMPK can interfere with the
synthesis of proteins by these pathways, and this has been linked
to the development of chronic pain and the reduced expression of
inflammatory factors. Thus, AMPK might be an ideal target for
reducing or preventing hyperalgesia (9). In spinal nerve ligation (SNL) and
spared nerve injury (SNI) models, metformin inhibited hyperalgesia
and activated AMPK in vitro (5). Metformin also reduced diabetic NP and
chemotherapy-induced NP in animal models (37,38).
In present study, we evidenced that metformin attenuates pain in
CCI rats and suppresses the expression of p-AMPK, which may explain
how metformin eases NP. The incidence of diabetic peripheral
neuropathy (DPN) reportedly decreased by long-term oral
administration of metformin, whereas the insulin treatment did not
show this effect. In our study, there was insignificant difference
of the blood glucose levels in rats with NP with or without the
application of metformin, indicating that the analgesic effect of
metformin is not associated with blood sugar levels, which is
supported by the prior report (39).
STAT3 is an important component of the Janus kinase
2 (JAK2)-STAT3 signaling pathway. The JAK2-STAT3 signal pathway is
widely involved in cell proliferation, differentiation, immune
regulation and information transfer, which is involved in the
formation of central sensitization in the spinal cord following
peripheral nerve injury (40).
Subsequent to the peripheral nerve damage, the inflammatory factor
IL-6 is released and combines with the soluble IL-6 receptor
(sIL-6R) in the SDH, thus activating gpl30 on the cell surface and
the JAK2-STAT3 signal transduction pathway, which is followed by
the activation and proliferation of glial cells as well as the
production and release of inflammatory cytokines (40,41).
These events result in a series of immune inflammatory reactions
that enhance neuronal excitability and lead to NP. Recently,
metformin is evidenced to repress the progression of cancers by the
inhibition of the STAT3 signaling pathway (14,15).
There was a paucity of data regarding the effect of metformin on
the STAT3 signaling pathway, with the underlying mechanisms remains
elusive. AMPK is considered as a mediator between metformin and the
STAT3 pathway (42,43). Moreover, the activation of the
STAT3 pathway depends on the activation of mTOR (44). Additionally, the activation of AMPK
inhibited inflammation and suppressed the mTOR signaling pathway in
a pain model (5,11). In our experiments, mTOR may be the
link between AMPK and the STAT3 pathway, and this may be the
mechanism by which metformin inhibits STAT3 activation. We have
therefore provided the first evidence showing that metformin
decreases the activation of STAT3 in the SDH, which plays an
important role in reducing NP.
The central nervous system consists of two types of
cells-glia and neurons, the latter of which has been a heightened
interest in the field of NP. Recently, emerging evidence focuses on
the ion channels and neurotransmitter receptors on the membranes of
glial cells. Glial cells can produce and release cytokines, which
promote the development of NP (20,21).
Glia in the spinal cord are mainly composed of astrocytes,
microglia and oligodendrocytes, with astrocytes being the most
ones. Spinal astrocytes and microglia have small cell bodies with
branched processes and are quiescent cells that monitor the local
environment in the central nervous system under the normal
conditions. In the event of an injury, spinal astrocytes and
microglial are activated (45,46),
and synthesize and release the pro-inflammatory cytokines, which
promote the neuroinflammation and NP. In our experiment, the glial
cells and astrocytes were activated after CCI, which were
effectively suppressed by metformin. The findings imply the
involvement of glial cells and astrocytes in the pathogenesis of NP
and the regulation of metformin for NP. In addition, STAT3 pathway
was implicated in modulation of spinal glial cell proliferation and
NP maintenance in rats (17,23,40).
In addition, AMPK is also involved in the functional regulation of
astrocytes and microglia (11,22,47).
Hence, metformin activated AMPK and inhibited the activation of the
STAT3 signaling pathway, both of which might further affect the
functionality of astrocytes and microglia.
Previous studies have shown that the activation of
microglia plays a role in the early stage of NP, while astrocyte
activation takes effect in the maintenance phase of NP (48). We determined the distribution of
astrocytes and microglia in the lumbar SDH on day 10 after sciatic
nerve ligation, the time point of which is considered as a stable
stage for CCI-induced NP. The L4-L6 spinal cord is the reference
point for sciatic nerve lesion. Iba-1 and GFAP immunofluorescence
staining results suggested that microglia and astrocytes were
activated in the L4-L6 SDH. It indicated that the activation of
microglia did not decrease and astrocytes have already been
activated on day 10 after surgery. Our results confirm the previous
claim that the activation of microglia and astrocytes may be an
important step during the initiation and maintenance of NP after a
peripheral nerve injury (20,21,45).
Our data show that metformin effectively inhibited the activation
of spinal microglia and astrocytes, which might be an important
mechanism by which metformin relieves NP.
In conclusion, intraperitoneal injections of
metformin activated AMPK and inhibited the activation of the STAT3
signaling pathway in the SDH of CCI rats. Metformin could inhibit
the activation of astrocytes and microglia, which might further
inhibit the release of inflammatory cytokines, such as interleukin
(IL-6, IL-1β) and tumor necrosis factor α (TNF-α), to alleviate NP.
However, the limitation of our present study is that the
colocalization of AMPK or STAT3 with glial cells was not presented.
We will further study their relationship in the following
studies.
The data in these experiments show that metformin is
an effective and safe drug for NP. Given its affordability and easy
accessibility, it is worthwhile to explore its clinical application
in NP. Following further explorations of the characteristics of
this drug and the mechanisms underlying NP, we believe that it is
reasonable that this drug can be served as a potential therapeutic
agent for chronic pain.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (grant no.
81200861).
References
|
1
|
Jensen TS, Baron R, Haanpää M, Kalso E,
Loeser JD, Rice AS and Treede RD: A new definition of neuropathic
pain. Pain. 152:2204–2205. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Finnerup NB, Sindrup SH and Jensen TS: The
evidence for pharmacological treatment of neuropathic pain. Pain.
150:573–581. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Vry J, Kuhl E, Franken-Kunkel P and
Eckel G: Pharmacological characterization of the chronic
constriction injury model of neuropathic pain. Eur J Pharmacol.
491:137–148. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Austin PJ, Wu A and Moalem-Taylor G:
Chronic constriction of the sciatic nerve and pain hypersensitivity
testing in rats. J Vis Exp:. 13:33932012.
|
|
5
|
Melemedjian OK, Asiedu MN, Tillu DV,
Sanoja R, Yan J, Lark A, Khoutorsky A, Johnson J, Peebles KA, Lepow
T, et al: Targeting adenosine monophosphate-activated protein
kinase (AMPK) in preclinical models reveals a potential mechanism
for the treatment of neuropathic pain. Mol Pain. 7:702011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taylor A, Westveld AH, Szkudlinska M,
Guruguri P, Annabi E, Patwardhan A, Price TJ and Yassine HN: The
use of metformin is associated with decreased lumbar radiculopathy
pain. J Pain Res. 6:755–763. 2013.PubMed/NCBI
|
|
7
|
Kim J, Yang G, Kim Y, Kim J and Ha J: AMPK
activators: Mechanisms of action and physiological activities. Exp
Mol Med. 48:e2242016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dasgupta B and Chhipa RR: Evolving Lessons
on the complex role of AMPK in normal physiology and cancer. Trends
Pharmacol Sci. 37:192–206. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Price TJ and Dussor G: AMPK: An emerging
target for modification of injury-induced pain plasticity. Neurosci
Lett. 557:9–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bullón P, Alcocer-Gómez E, Carrión AM,
Marín-Aguilar F, Garrido-Maraver J, Román-Malo L, Ruiz-Cabello J,
Culic O, Ryffel B, Apetoh L, et al: AMPK phosphorylation modulates
pain by activation of NLRP3 inflammasome. Antioxid Redox Signal.
24:157–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song H, Han Y, Pan C, Deng X, Dai W, Hu L,
Jiang C, Yang Y, Cheng Z, Li F, et al: Activation of adenosine
monophosphate-activated protein kinase suppresses neuroinflammation
and ameliorates bone cancer pain. Anesthesiology. 123:1170–1185.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hawley SA, Ross FA, Chevtzoff C, Green KA,
Evans A, Fogarty S, Towler MC, Brown LJ, Ogunbayo OA, Evans AM and
Hardie DG: Use of cells expressing gamma subunit variants to
identify diverse mechanisms of AMPK activation. Cell Metab.
11:554–565. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Krishan S, Richardson DR and Sahni S:
Adenosine monophosphate-activated kinase and its key role in
catabolism: structure, regulation, biological activity and
pharmacological activation. Mol Pharmacol. 87:363–377. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Q, Yuan W, Tong D, Liu G, Lan W, Zhang
D, Xiao H, Zhang Y, Huang Z, Yang J, et al: Metformin represses
bladder cancer progression by inhibiting stem cell repopulation via
COX2/PGE2/STAT3 axis. Oncotarget. 7:282352016.PubMed/NCBI
|
|
15
|
Park J, Kim WG, Zhao L, Enomoto K,
Willingham M and Cheng SY: Metformin blocks progression of
obesity-activated thyroid cancer in a mouse model. Oncotarget.
7:348322016.PubMed/NCBI
|
|
16
|
Liu S, Li Q, Zhang MT, Mao-Ying QL, Hu LY,
Wu GC, Mi WL and Wang YQ: Curcumin ameliorates neuropathic pain by
down-regulating spinal IL-1β via suppressing astroglial NALP1
inflammasome and JAK2-STAT3 signalling. Sci Rep. 6:289562016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsuda M, Kohro Y, Yano T, Tsujikawa T,
Kitano J, Tozaki-Saitoh H, Koyanagi S, Ohdo S, Ji RR, Salter MW and
Inoue K: JAK-STAT3 pathway regulates spinal astrocyte proliferation
and neuropathic pain maintenance in rats. Brain. 134:1127–1139.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang ZF, Li Q, Liu SB, Mi WL, Hu S, Zhao
J, Tian Y, Mao-Ying QL, Jiang JW, Ma HJ, et al: Aspirin-triggered
Lipoxin A4 attenuates mechanical allodynia in association with
inhibiting spinal JAK2/STAT3 signaling in neuropathic pain in rats.
Neuroscience. 273:65–78. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Park KW, Lin CY and Lee YS: Expression of
suppressor of cytokine signaling-3 (SOCS3) and its role in neuronal
death after complete spinal cord injury. Exp Neurol. 261:65–75.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alfonso Romero-Sandoval E and Sweitzer S:
Nonneuronal central mechanisms of pain: Glia and immune response.
Prog Mol Biol Transl Sci. 131:325–358. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Walters ET: Neuroinflammatory
contributions to pain after SCI: Roles for central glial mechanisms
and nociceptor-mediated host defense. Exp Neurol. 258:48–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gui Y, Li A, Chen F, Zhou H, Tang Y, Chen
L, Chen S and Duan S: Involvement of AMPK/SIRT1 pathway in
anti-allodynic effect of troxerutin in CCI-induced neuropathic
pain. Eur J Pharmacol. 769:234–241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Molet J, Mauborgne A, Diallo M, Armand V,
Geny D, Villanueva L, Boucher Y and Pohl M: Microglial Janus
kinase/signal transduction and activator of transcription 3 pathway
activity directly impacts astrocyte and spinal neuron
characteristics. J Neurochem. 136:133–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gerard E, Spengler RN, Bonoiu AC, Mahajan
SD, Davidson BA, Ding H, Kumar R, Prasad PN, Knight PR and
Ignatowski TA: Chronic constriction injury-induced nociception is
relieved by nanomedicine-mediated decrease of rat hippocampal tumor
necrosis factor. Pain. 156:1320–1333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hargreaves K, Dubner R, Brown F, Flores C
and Joris J: A new and sensitive method for measuring thermal
nociception in cutaneous hyperalgesia. Pain. 32:77–88. 1988.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang T, Sun K, Shen W, Qi L, Yin W and
Wang LW: SOCS1 regulates neuropathic pain by inhibiting neuronal
sensitization and glial activation in mouse spinal cord. Brain Res
Bull. 124:231–237. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cao H and Zhang YQ: Spinal glial
activation contributes to pathological pain states. Neurosci
Biobehav Rev. 32:972–983. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Benarroch EE: Dorsal horn circuitry
Complexity and implications for mechanisms of neuropathic pain.
Neurology. 86:1060–1069. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
West SJ, Bannister K, Dickenson AH and
Bennett DL: Circuitry and plasticity of the dorsal horn-Toward a
better understanding of neuropathic pain. Neuroscience.
300:254–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Martin M and Marais R: Metformin: A
diabetes drug for cancer, oral cancer drug for diabetics? J Clin
Oncol. 30:2698–2700. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wan W, Cao L, Khanabdali R, Kalionis B,
Tai X and Xia S: The emerging role of HMGB1 in neuropathic pain: A
Potential therapeutic target for neuroinflammation. J Immunol Res.
2016:64304232016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou YQ, Liu Z, Liu ZH, Chen SP, Li M,
Shahveranov A, Ye DW and Tian YK: Interleukin-6: An emerging
regulator of pathological pain. J Neuroinflammation. 13:1412016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Edelmayer RM, Brederson JD, Jarvis MF and
Bitner RS: Biochemical and pharmacological assessment of MAP-kinase
signaling along pain pathways in experimental rodent models: A
potential tool for the discovery of novel antinociceptive
therapeutics. Biochem Pharmacol. 87:390–398. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin X, Wang M, Zhang J and Xu R: p38 MAPK:
A potential target of chronic pain. Curr Med Chem. 21:4405–4418.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lisi L, Aceto P, Navarra P and Dello Russo
C: MTOR kinase: A possible pharmacological target in the management
of chronic pain. Biomed Res Int. 2015:3942572015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sanna MD, Stark H, Lucarini L, Ghelardini
C, Masini E and Galeotti N: Histamine H4 receptor activation
alleviates neuropathic pain through differential regulation of ERK,
JNK and P38 MAPK. phosphorylation. 156:1–2504. 2015.
|
|
37
|
Ma J, Yu H, Liu J, Chen Y, Wang Q and
Xiang L: Metformin attenuates hyperalgesia and allodynia in rats
with painful diabetic neuropathy induced by streptozotocin. Eur J
Pharmacol. 764:599–606. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mao-Ying QL, Kavelaars A, Krukowski K, Huo
XJ, Zhou W, Price TJ, Cleeland C and Heijnen CJ: The anti-diabetic
drug metformin protects against chemotherapy-induced peripheral
neuropathy in a mouse model. PLoS One. 9:e1007012014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pop-Busui R, Lu J, Lopes N and Jones TL;
BARI 2D Investigators, : Prevalence of diabetic peripheral
neuropathy (DPN) and relation to glycemic control strategies at
baseline in the BARI 2D cohort. J Peripher Nerv Syst. 14:1–13.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dominguez E, Rivat C, Pommier B, Mauborgne
A and Pohl M: JAK/STAT3 pathway is activated in spinal cord
microglia after peripheral nerve injury and contributes to
neuropathic pain development in rat. J Neurochem. 107:50–60. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dubový P, Klusáková I, Svízenská I and
Brázda V: Satellite glial cells express IL-6 and corresponding
signal-transducing receptors in the dorsal root ganglia of rat
neuropathic pain model. Neuron Glia Biol. 6:73–83. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yue W, Zheng X, Lin Y, Yang CS, Xu Q,
Carpizo D, Huang H, DiPaola RS and Tan XL: Metformin combined with
aspirin significantly inhibit pancreatic cancer cell growth in
vitro and in vivo by suppressing anti-apoptotic proteins Mcl-1 and
Bcl-2. Oncotarget. 6:21208–21224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hattori Y, Hattori K and Hayashi T:
Pleiotropic benefits of metformin: Macrophage targeting its
anti-inflammatory mechanisms. Diabetes. 64:1907–1909. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yokogami K, Wakisaka S, Avruch J and
Reeves SA: Serine phosphorylation and maximal activation of STAT3
during CNTF signaling is mediated by the rapamycin target mTOR.
Curr Biol. 10:47–50. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tsuda M: Microglia in the spinal cord and
neuropathic pain. J Diabetes Investig. 7:17–26. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nimmerjahn A, Kirchhoff F and Helmchen F:
Resting microglial cells are highly dynamic surveillants of brain
parenchyma in vivo. Science. 308:1314–1318. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Maixner DW, Yan X, Gao M, Yadav R and Weng
HR: Adenosine Monophosphate-activated protein kinase regulates
interleukin-1β expression and glial glutamate transporter function
in rodents with neuropathic pain. Anesthesiology. 122:1401–1413.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hald A, Nedergaard S, Hansen RR, Ding M
and Heegaard AM: Differential activation of spinal cord glial cells
in murine models of neuropathic and cancer pain. Eur J Pain.
13:138–145. 2009. View Article : Google Scholar : PubMed/NCBI
|