Introduction
Pancreatic ductal adenocarcinoma (PDAC) is one of
the most aggressive malignant tumors worldwide; it has a poor
prognosis and a 5-year survival rate of ~8% in the USA (1). The median survival time for patients
with advanced PDAC is <7 months, even with active treatment
(2). Statistics from the National
Cancer Institute Surveillance, Epidemiology and End Results
database (https://seer.cancer.gov) indicate that
PDAC is the fourth leading cause of cancer-related mortality,
accounting for ~7% of all cancer deaths in the United States. The
mortality rate of men with pancreatic cancer increased slightly
from 2005 to 2014, at a rate of 0.3% per year (1). Based on this trend, Hezel et
al predicted that pancreatic cancer would become the second
most common cause of cancer-related death by 2030 (3). Resection of pancreatic tumor tissue
during the early stage is currently the only curative treatment
method. However, as symptoms and biomarkers for early diagnosis are
lacking, >80% of patients with PDAC are not diagnosed until they
are in the advanced stage, thereby missing the best timeframe for
curative treatment (4).
Additionally, the lack of effective and credible interventions is
another key cause of the high mortality rate in patients with PDAC
(3). Thus, new targets for PDAC
diagnosis and better therapeutics for PDAC treatment are urgently
needed.
Pancreatic cancer has many risk factors, including
individual, lifestyle-related, disease and drug-related factors,
with PDAC being primarily caused by somatically acquired mutations
(5). Jones et al
investigated gene mutations involved in pancreatic cancer and found
an average of 63 mutated genes (6). A number of previous studies have
examined gene mutations in pancreatic cancer. For example, Nagata
et al reported and validated that mucins, such as mucin
(MUC) 1, MUC5 and MUC6, were overexpressed in
invasive ductal carcinoma (IDC) and in pancreatic intraepithelial
neoplasia, which develops into IDC (7). Kirsten rat sarcoma viral oncogene
homolog (KRAS) has been confirmed as the driver gene in PDAC
(6–8). By establishing a genetically
engineered mouse PDAC model, Rozenblum et al demonstrated
that SMAD4 is inactivated in most pancreatic cancers, which
leads to inactivation of the tumor suppressor gene cyclin-dependent
kinase inhibitor 2A and activation of KRAS (9,10).
p53, another tumor suppressor gene, has been found to be mutated in
>50% of patients with PDAC (3,9).
Fong and Winter systematically summarized the presence of various
biomarkers in cancer cells and body fluids in patients with PDAC
based on previous studies; these biomarkers included the genes
KRAS, Fanconi anemia complementation group I (FANCI)
and MUC1, and the proteins CA-19-9, CA-125 and CEA, as well
as microRNA (miR)-21 and miR-210 (11). Unexpectedly, although >2,000
studies have been conducted on biomarkers for pancreatic cancer,
the CA-19-9 serum level is the only widely used Food and Drug
Administration-approved tumor marker, albeit not for diagnosis. To
date, no biomarkers are sufficiently accurate for widespread
diagnostic use (12).
Since the emergence of high-throughput genomic
technologies, including microarrays several decades ago, these
techniques have been applied to study many diseases. For example,
these technologies are used to identify disease subtypes (13), explore gene expression profiles of
disease (14) and to identify
potential novel pathogenic genes and diseases (15). Additionally, molecular signatures
discovered by microarrays have become predictive biomarkers for
certain diseases (16). An
increasing number of studies have applied this technique to
identify differentially expressed genes (DEGs) between tumor
tissues and normal tissues (17).
Although microarray technology has enabled the discovery of
biomarkers, study results vary owing to inaccuracies or quality
problems in microarray analyses and verification, and small sample
sizes exacerbate this issue (18).
A meta-analysis, as a large-sample study, has an advantage in
addressing this limitation due to its enhanced statistical power
(19). Prior meta-analyses have
been applied to study tumors to confirm DEGs between tumor tissue
and normal tissue in glioma (20,21),
lung cancer (22), bladder cancer
(23), breast cancer (24), osteosarcoma (25), liver cancer (26) and pancreatic cancer (27). In the present study, integrative
meta-analysis of expression data (INMEX) was used to conduct a
meta-analysis based on 11 qualified microarray datasets, with the
aim to identify crucial DEGs between PDAC samples and normal
pancreatic samples that may serve as biomarkers for PDAC treatment
and prognosis.
Materials and methods
Patients and tissue collection
The mean age of patients was 56.3 years (range,
39–73). Paired normal pancreatic tissues and PDAC tissues were
collected from 15 patients (female to male ratio, 4:11) with PDAC
who underwent surgical procedures at the Union Hospital of Tongji
Medical College, Huazhong University of Science and Technology
(Wuhan, China) between July 2018 and September 2018. Human study
protocols were approved by the ethics committee of Tongji Medical
College, Huazhong University of Science and Technology (Wuhan,
China), and all patients in the study provided written informed
consent. The pathological diagnoses of the specimens were confirmed
by two clinicopathological experts at the Department of Pathology,
Union Hospital of Tongji Medical College, Huazhong University of
Science and Technology prior to RNA extraction. Specimens were snap
frozen in liquid nitrogen and stored at −80°C. According to the
American Joint Commission on Cancer Staging System for patients
with pancreatic adenocarcinoma (28), patients were divided into three
groups: i) Stage I (n=7); ii) stage II (n=7); and stage III
(n=1).
Identification of PDAC microarray
datasets and validation in The Cancer Genome Atlas (TCGA)
The keywords ‘pancreatic cancer’ were used to search
for gene chips to study in the Gene Expression Omnibus (GEO)
datasets of the National Center for Biotechnology Information
database (http://www.ncbi.nlm.nih.gov/geo). The inclusion
criteria for the qualified chips were as follows: i) The chip was a
gene expression chip; ii) the genes were from normal pancreatic
and/or PDAC tissues; iii) the GEO series (GSE) dataset included
>3 samples. Gene chips that met any of the following criteria
were excluded: i) Chips containing pancreatic cancer cell lines;
ii) methylated gene chips; iii) genes originating from animals
other than humans; iv) non-microarray gene chips. Following the
determination of the microarray datasets, the GSE number,
information regarding the expression platform and sample number,
source literature and the number of normal and pancreatic cancer
samples were collected from the GEO database. To validate the
results in TCGA (https://tcga-data.nci.Nih.gov/tcga/), DNA expression
levels in pancreatic cancer and normal tissues and patients'
survival time were obtained from the database.
Elimination of batch differences and
individual data analysis
INMEX (http://www.inmex.ca) is an online tool that supports
multiple microarray platforms and commonly used gene IDs; it has
comprehensive processing capabilities for meta-analysis of multiple
gene expression sets (29). The
ComBat option using the empirical Bayes method in INMEX was applied
to eliminate the differences between dataset batches on different
platforms and on the same platform and to ensure that different
microarray experiments were directly comparable to each other to
avoid inaccuracies resulting from differences unrelated to the
disease. Extremely high or low expression ratios for additional
genes were robustly stabilized by contracting differences by the
empirical Bayes method (30).
Meta-analysis of microarray
datasets
All selected microarray datasets were
log2 transformed and each constructed relative
expression value table was set to match INMEX's upload format: The
rows contained gene information and the columns contained sample
information; each table was then uploaded. The data were annotated,
function IDs were matched, and outlier samples were determined from
boxplots and principal component analysis plots to ensure
normalized quantile data. The correctness and integrity of all
datasets were checked prior to running the ComBat adjust batch
effect function, and the meta-analysis was performed. Considering
the characteristics of each statistical method, the Cochran Q test
was performed on the uploaded INMEX data, which revealed notable
heterogeneity among different studies. Therefore, a random-effects
model combined with the moderated effect size (ES) and metaMA
packages were selected for the DEG screening (31,32).
The gplots package in R software (version 3.5.0; http://www.r-project.org) was used to perform
hierarchical cluster analysis of the DEGs.
Functional enrichment analysis of the
DEGs
To determine the potential functions of the screened
DEGs, Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and
Genomes (KEGG) pathway analyses were performed in the Database for
Annotation, Visualization, and Integrated Discovery (DAVID;
http://david.ncifcrf.gov) and the top 10 most
enriched terms with P<0.05 were identified.
Network-based meta-analysis
To better understand DEG expression, protein-protein
interaction (PPI) networks were constructed using NetworkAnalyst
with an extensive and high-quality PPI database based in InnateDB
(33). The Hub Explorer tool was
used to retrieve information on node levels, betweenness centrality
and expression levels. The types of data obtained were the degree
of the node (number of connections to other nodes) and betweenness
centrality (number of shortest paths through the node). The
expression was defined as the log fold change value of the
corresponding node. Nodes with the highest degree or betweenness
values were considered crucial hub nodes.
Verification of gene expression and
Kaplan-Meier analysis
To verify the expression of the top ten upregulated
and downregulated genes in PDAC tissues compared with normal
tissues, scatter plots of TCGA data were produced using GraphPad
Prism 6 (GraphPad Software, Inc.). To explore the effects of DEGs
on survival time, Kaplan-Meier curves were constructed using
GraphPad Prism 6 based on DNA expression profiles in the PDAC
tissue samples and the corresponding survival times extracted from
TCGA. The median expression level of each DEG was set as the cutoff
value to split TCGA cohorts into high- and low-expression groups.
The log-rank tests were used to assess the differences between
groups of patients exhibiting low or high expression of the
selected DEGs. P<0.05 was considered to indicate a statistically
significant difference.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
To confirm S100A6, KRT19 and GNG7
expression, PDAC tissues and paired normal pancreatic tissues from
15 patients with pathologically confirmed PDAC were collected.
Total RNA was extracted from tissues using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. First-strand cDNA was synthesized using
the RevertAid First Strand cDNA Synthesis kit (Thermo Fisher
Scientific, Inc.) following the manufacturer's instructions.
RT-qPCR was performed using the FastStart Universal SYBR Green
master mix (Roche Diagnostics). The thermocycling conditions were
as follows: Initial denaturation at 95°C for 10 min, followed by 40
cycles of denaturation at 95°C for 15 sec and annealing/elongation
at 60°C for 60 sec. The 2−∆∆Cq quantification method was
used for analyzing the qPCR data (34). The primers used are listed in
Table I. The gene expression
levels in each sample were normalized to GAPDH.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Gene name | Primer sequence
(5′-3′) |
|---|
| GAPDH | F:
ACTTTGGTATCGTGGAAGGACTCAT |
|
| R:
GTTTTTCTAGACGGCAGGTCAGG |
| S100A6 | F:
CCATCTTCCACAAGTACTCCGG |
|
| R:
GCAGCTTCGAGCCAATGGT |
| GNG7 | F:
CGCATAGAAGCCGGGATTGA |
|
| R:
TTGTCCTTAAAGGGGTTCTCCG |
| KRT19 | F:
AGAATTGAACCGGGAGGTCG |
|
| R:
CCTGATTCTGCCGCTCACTA |
Statistical analysis
The ES combined random-effects model was used to
perform the meta-analysis, and the threshold to screen for DEGs was
set at P<0.05. The Benjamini-Hochberg false discovery rate was
used to correct the P-value to obtain more precise outcomes. The
Mann-Whitney U test was used to conduct validation of gene
expression in TCGA. S100A6, KRT19 and GNG7 expression
results for the paired PDAC and normal samples were compared using
paired Student's t-test or Wilcoxon test. P<0.05 was considered
to indicate a statistically significant difference.
Results
Meta-analysis
Raw data from the following 11 microarray gene
expression profile datasets were downloaded from the GEO database:
GSE15471 (35), GSE16515 (36), GSE22780, GSE28735 (37,38),
GSE32676 (39), GSE43288-GPL96
(40), GSE43288-GPL97 (40), GSE43795 (41), GSE46234, GSE55643 (42) and GSE62165 (43). In total, 485 samples were analyzed
(334 tumor samples and 151 non-tumor samples); the two sample types
were subjected to meta-analysis to identify DEGs. Specific
information for each dataset included the first author, country,
publication date, platform and numbers of PDAC and normal
pancreatic tissue samples. The complete information and reference
numbers of each dataset are presented in Table II. The detailed screening and
processing workflow is depicted in Fig. 1.
| Table II.Characteristics of datasets used in
the meta-analysis of pancreatic ductal adenocarcinoma vs. Normal
tissue. |
Table II.
Characteristics of datasets used in
the meta-analysis of pancreatic ductal adenocarcinoma vs. Normal
tissue.
|
|
|
|
| Numbers |
|
|---|
|
|
|
|
|
|
|
|---|
| Author, year | Source
accession | Country | Platform | PDAC | Normal | (Refs.) |
|---|
| Badea et al,
2009 | GSE15471 | Romania | GPL570 | 39 | 39 | (35) |
| Pei et al,
2009 | GSE16515 | USA | GPL570 | 36 | 16 | (36) |
| Killary, 2011 | GSE22780 | USA | GPL570 | 8 | 8 | N/A |
| Hussain et
al, 2012 | GSE28735 | USA | GPL6244 | 45 | 45 | (37,38) |
| Donahue et
al, 2011 | GSE32676 | USA | GPL570 | 25 | 7 | (39) |
| Crnogorac-Jurcevic
et al, 2013 | GSE43288 | UK | GPL96 | 8 | 6 | (40) |
| Crnogorac-Jurcevic
et al, 2013 | GSE43288 | UK | GPL97 | 8 | 6 | (40) |
| Kim et al,
2013 | GSE43795 | South Korea | GPL10558 | 6 | 5 | (41) |
| Raeder, 2017 | GSE46234 | Norway | GPL570 | 4 | 4 | N/A |
| Jamieson et
al, 2014 | GSE55643 | UK | GPL6480 | 45 | 8 | (42) |
| Janky et al,
2016 | GSE62165 | Belgium | GPL13667 | 118 | 13 | (43) |
Eliminating differences between
batches
As differences between platforms lead to
inaccuracies, biases in these datasets must be eliminated to
increase the precision and reliability of the results. Therefore,
prior to the meta-analysis, ComBat in INMEX was used to correct for
differences between batches online. A principal component analysis
plot demonstrated modest separation among the 11 datasets (data not
shown).
Meta-analysis of gene expression in
PDAC
DEGs with low but continuous expression levels in
all profile datasets were defined as gained genes, and DEGs that
disappeared in the meta-analysis but were expressed in individual
analyses or in the meta-analysis resulting from experimental errors
in the platforms were defined as lost genes (20,44,45).
In total, one gained gene, unkempt family zinc finger-like
(UNKL), which was weakly expressed in all datasets (ES,
−0.25395), and 1,278 lost genes were identified. Through the
meta-analysis, 1,587 DEGs were identified, of which 1,004 were
upregulated and 583 were downregulated (data not shown).
Unsupervised hierarchical clustering of the DEGs was performed, and
the results demonstrated that PDAC and normal samples were
partitioned into two major groups (Fig. 2). Among all DEGs, the top 10
upregulated genes were hematological and neurological expressed 1
(HN1), integrin subunit α2 (ITGA2), S100
calcium-binding protein A6 (S100A6), inhibin βA subunit
(INHBA), keratin-19 (KRT19), membrane-bound
O-acyltransferase domain-containing 2 (MBOAT2), collagen
type I α2 chain (COL1A2), collagen type III α1 chain
(COL3A1), monoglyceride lipase (MGLL) and β-actin
(ACTB) (Table III). The
top 10 downregulated genes were kinesin family member 1A
(KIF1A), Dymeclin (DYM), β-secretase 1
(BACE1), nucleobindin-2 (NUCB2), phosphoserine
aminotransferase 1 (PSAT1), LIF receptor α (LIFR), BR
serine/threonine kinase 2 (BRSK2), potassium two pore domain
channel subfamily K member 3 (KCNK3), family with sequence
similarity 46 member C (FAM46C) and G protein subunit γ7
(GNG7) (Table III).
| Table III.Top 20 differentially expressed genes
identified in the meta-analysis of PDAC vs. normal tissues. |
Table III.
Top 20 differentially expressed genes
identified in the meta-analysis of PDAC vs. normal tissues.
| A, Top 10
upregulated genes in PDAC vs. Normal tissue |
|---|
|
|---|
| Entrez ID | Gene | Gene name | Combined ES | Adjusted
P-value |
|---|
| 51155 | HN1 | Hematological and
neurological expressed 1 | 2.4036 |
7.5536×10−10 |
| 3673 | ITGA2 | Integrin subunit
alpha 2 | 2.3434 |
5.0162×10−7 |
| 6277 | S100A6 | S100 calcium
binding protein A6 | 2.1598 |
1.1665×10−6 |
| 3624 | INHBA | Inhibin beta A
subunit | 2.1409 |
2.4647×10−5 |
| 3880 | KRT19 | Keratin 19 | 2.0556 |
5.215×10−5 |
| 129642 | MBOAT2 | Membrane bound
O-acyltransferase domain containing 2 | 2.046 | <0.001 |
| 1278 | COL1A2 | Collagen type I
alpha 2 chain | 2.0415 |
4.4806×10−5 |
| 1281 | COL3A1 | Collagen type III
alpha 1 chain | 1.993 |
1.0244×10−4 |
| 11343 | MGLL | Monoglyceride
lipase | 1.8727 |
1.1349×10−12 |
| 60 | ACTB | β actin | 1.8532 |
1.0806×10−4 |
|
| B, Top 10
downregulated genes in PDAC vs. Normal tissue |
|
| Entrez
ID | Gene | Gene
name | Combined
ES | Adjusted
P-value |
|
| 547 | KIF1A | Kinesin family
member 1A | −1.9848 |
7.0263×10−7 |
| 54808 | DYM | Dymeclin | −1.8453 |
3.6218×10−7 |
| 23621 | BACE1 | β-secretase 1 | −1.8387 |
1.302×10−4 |
| 4925 | NUCB2 | Nucleobindin 2 | −1.8176 |
5.8813×10−7 |
| 29968 | PSAT1 | Phosphoserine
aminotransferase 1 | −1.809 |
6.8582×10−7 |
| 3977 | LIFR | LIF receptor
alpha | −1.7681 |
1.2712×10−10 |
| 9024 | BRSK2 | BR serine/threonine
kinase 2 | −1.7642 |
6.1567×10−4 |
| 3777 | KCNK3 | Potassium two pore
domain channel subfamily K member 3 | −1.7365 |
1.1409×10−6 |
| 54855 | FAM46C | Family with
sequence similarity 46 member C | −1.7315 |
4.01×10−11 |
| 2788 | GNG7 | G protein subunit
gamma 7 | −1.6543 |
1.6178×10−6 |
Identification of hub genes using
network-based meta-analysis
Hub genes serve key roles in interrelationships
among DEGs. To identify hub genes among the PDAC DEGs, a PPI
network analysis was conducted using NetworkAnalyst, which included
comprehensive data curated from the literature by InnateDB. One
large subnetwork comprising 8,527 nodes and 37,579 edges, and a
smaller subnetwork containing 3 nodes and 2 edges were obtained,
and the top 10 hub genes in PDAC vs. normal tissue were identified
(Table IV). To display the
results more clearly, a zero-edge network analysis was performed,
which comprised 965 nodes and 2,806 edges (Fig. 3). Finally, hub genes in the network
were ranked by degree. Growth factor receptor-bound protein 2
(GRB2), with a combined ES of 0.76104 and an adjusted
P-value of 0.0053254, had the highest degree (90) and betweenness
(53,725) among upregulated DEGs. Histone deacetylase 5
(HDAC5), with a combined ES of −0.37624 and an adjusted
P-value of 0.0018425, had the highest degree (45) and betweenness (17,157) among
downregulated DEGs.
 | Figure 3.Identification of hub genes using
network-based meta-analysis. ‘Zero order’ interaction network for
DEGs by meta-analysis of PDAC in comparison with normal tissue in a
forced atlas layout. CAND1, cullin-associated and
neddylation-dissociated 1; CBL, Cbl proto-oncogene;
CREBBP, CREB binding protein; CTNNB1, catenin β1;
CUL1, cullin 1; GRB2, growth factor receptor-bound
protein 2; HDAC5, histone deacetylase 5; SMURF1,
SMAD-specific E3 ubiquitin protein ligase 1; SP1, Sp1
transcription factor; UBE2I, ubiquitin-conjugating enzyme E2
I. |
| Table IV.The top 10 hub genes in PDAC vs.
Normal tissue. |
Table IV.
The top 10 hub genes in PDAC vs.
Normal tissue.
| A, Upregulated
genes |
|---|
|
|---|
| ID | Gene | Degree | Betweenness |
|---|
| 2885 | GRB2 | 90 | 53,725 |
| Q86VP6 | CAND1 | 83 | 40,684 |
| Q13616 | CUL1 | 82 | 43,269 |
| P08047 | SP1 | 74 | 47,977 |
| P35222 | CTNNB1 | 56 | 27,401 |
| P63279 | UBE2I | 45 | 24,406 |
| Q9UQL6 | HDAC5 | 45 | 17,157 |
| Q9HCE7 | SMURF1 | 41 | 16,525 |
| P22681 | CBL | 40 | 11,550 |
| Q92793 | CREBBP | 39 | 14,026 |
|
| B, Downregulated
genes |
|
| ID | Gene | Degree |
Betweenness |
|
| Q9UQL6 | HDAC5 | 45 | 17,157 |
| Q92793 | CREBBP | 39 | 14,026 |
Functional annotation and pathway
enrichment analyses
To examine the functions of the identified DEGs, the
DEGs were mapped in DAVID to perform GO term and KEGG pathway
analyses. The following three aspects were included in the GO
analyses: Biological process (BP), cellular component (CC) and
molecular function (MF). In the GO analyses, DEGs identified
between normal and PDAC tissues were notably enriched in ‘cell-cell
adhesion’ (BP), ‘cytosol’ (CC) and ‘protein binding’ (MF) (Fig. 4A-C). In total, 27 KEGG pathways
were significantly enriched (P<0.05) between normal and PDAC
tissues. The top two enriched pathways with the lowest P value were
‘ubiquitin-mediated proteolysis’ (P=7.40×10−9) and
‘pathways in cancer’ (P=6.23×10−5) (Fig. 4D), which demonstrated that these
DEGs may be strongly associated with pancreatic cancer
occurrence.
TCGA validation
The top 10 upregulated and downregulated DEGs
obtained by meta-analysis were validated in TCGA using 147 PDAC
samples and four normal samples. The results revealed that compared
with the expression levels in normal samples, S100A6 and
KRT19 were strongly upregulated in the PDAC samples
(Fig. 5A and B). Compared with the
expression levels in normal samples, LIFR and GNG7
were significantly downregulated in the PDAC samples (Fig. 5C and D).
Kaplan-Meier analysis
The TCGA dataset comprised 157 pancreatic cancer
cases with detailed clinical and prognostic information and gene
expression data. To determine the effects of the identified DEGs on
the survival time of patients with PDAC, Kaplan-Meier analysis was
used for the top 10 upregulated DEGs and the top 10 downregulated
DEGs among the PDAC samples in the TCGA dataset. The results
revealed that patients with high ITGA2 expression levels had
shorter survival times compared with patients with lower
ITGA2 expression levels (Fig.
6A). KRT19, MBOAT2 and MGLL exhibited similar
results to ITGA2 (Fig.
6C-E). By contrast, patients with high FAM46C expression
had longer survival times compared with patients with low
FAM46C expression (Fig.
6G). These findings further demonstrated that high expression
levels of upregulated DEGs, such as ITGA2, KRT19, MBOAT2 and
MGLL, may represent a risk factor for poor survival in PDAC.
By contrast, high expression levels of downregulated DEGs, such as
FAM46C, may positively influence survival times of patients
with PDAC.
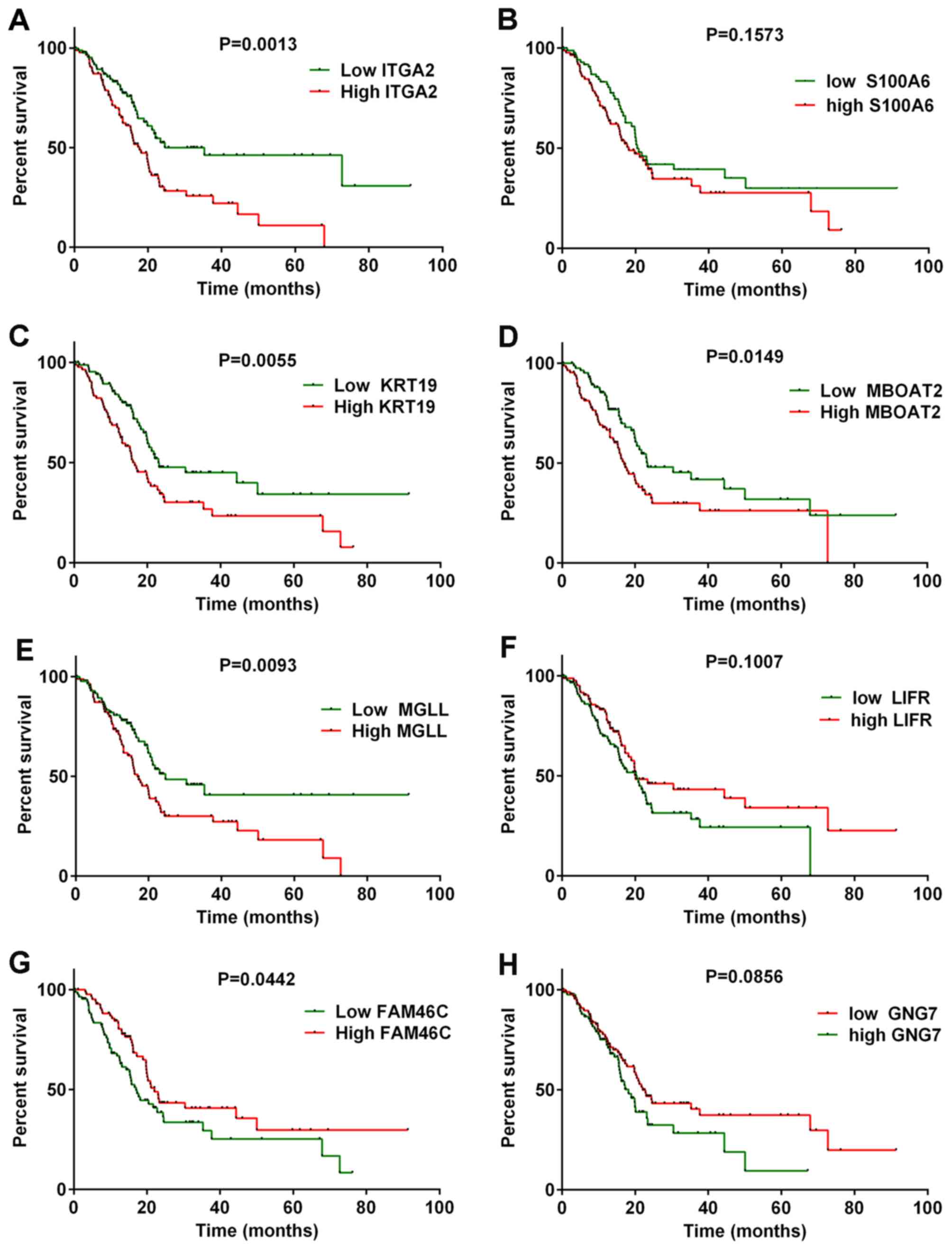 | Figure 6.Kaplan-Meier survival curve analysis.
Kaplan-Meier analysis of overall survival for patients with PDAC
with high expression levels of upregulated and downregulated genes.
(A) ITGA2, (B) S100A6, (C) KRT19, (D) MBOAT2 and (E) MGLL were
upregulated genes. (F) LIFR, (G) FAM46C and (H) GNG7 were
downregulated genes. Red lines represent high expression of DEGs
and green lines represent low expression of DEGs. The censored
subjects are marked with ticks on the Kaplan-Meier survival curves.
DEG, differentially expressed gene; FAM46C, family with
sequence similarity 46 member C; GNG7, G protein subunit 7;
ITGA2, integrin subunit α2; KRT19, keratin-19;
LIFR, LIF receptor; MBOAT2, membrane-bound
O-acyltransferase domain-containing 2; MGLL, monoglyceride
lipase; PDAC, pancreatic ductal adenocarcinoma; S100A6, S100
calcium-binding protein. |
Validation of genes with significantly
different expression in TCGA by RT-qPCR
To validate S100A6, KRT19 and GNG7
expression, RT-qPCR was performed using paired normal pancreatic
tissues and PDAC tissues, which were collected from 15 patients.
Among the three genes, S100A6 was significantly upregulated
in PDAC tissues compared with normal pancreatic tissues (Fig. 7A). By contrast, the expression of
KRT19 and GNG7 (Fig. 7B
and C) PDAC did not significantly differ from normal tissues,
although enhanced/low expression levels of these genes were
detected in some PDAC tissues.
Discussion
With the rapid development of high-throughput
genomic technology, researchers have explored gene mutations in
pancreatic cancer over the past two decades, including oncogenes
such as KRAS, SMAD4, proto-oncogene c-Myc and RAD51
recombinase (46), as well as
tumor suppressor genes such as cyclin-dependent kinase inhibitor
(CDKN2A) and p53 (6). Although these studies initially
identified several pancreatic cancer-related gene mutations, DEGs
derived from individual studies may be unreliable due to the
limited quality and quantity of microarray chips. Identifying
specific genes that can be used as biomarkers to diagnose
pancreatic cancer or for targeted therapies is the key to reducing
pancreatic cancer-related death and improving pancreatic cancer
prognosis. Tang et al identified 205 pancreatic
cancer-related genes (142 upregulated and 63 downregulated) by
screening and merging DEGs from four transcriptome microarray
datasets (226 PDAC samples and 65 normal pancreatic tissue samples)
in the GEO database (27). Through
functional analysis, including GO, KEGG pathway, Kaplan-Meier
survival and PPI network analyses, the authors identified
DKK1 and HMGA2 as candidate genes in PDAC. Notably,
these two genes were not identified as DEGs in the present study.
In the present study, a meta-analysis was performed using 11
high-quality microarray datasets (334 tumor samples and 151 normal
samples) from the GEO database to identify reliable PDAC-related
DEGs; 1,587 DEGs were identified, including 1,004 upregulated and
583 downregulated genes. One gained gene, UNKL, was
identified with a combined ES value of −0.25395. This gained gene
was previously unreported. Additionally, based on the PPI network,
GRB2 and HDAC5 were identified as pivotal hub genes
in PDAC. In addition to the aforementioned functional analysis by
Tang et al (27), the
expression of several top genes was verified by TCGA analysis and
RT-qPCR.
Among the upregulated genes, HN1 had the
highest combined ES value of 2.4036. The present study was
consistent with a previous study by Tang et al (27). In addition, many studies have
confirmed that HN1 mutations are related to cancer
occurrence. For example, Lu et al reported that HN1
is mutated in ovarian carcinoma epithelial tissue and can be used
as a biomarker for ovarian cancer (47). Laughlin et al established a
mouse glioma model and confirmed that murine HN1 depletion
led to a significant reduction in glioma volume (48). They also demonstrated that
HN1 was associated with the melanoma phenotype and regulated
cell proliferation and melanogenesis (48). HN1 is overexpressed in
breast cancer and can promote invasion and metastasis in breast
cancer (49) and prostate cancer
cells (50). These studies
illustrate that HN1 gene expression is related to the
proliferation, invasion and metastasis of tumor cells. The results
of the bioinformatics analysis in the present study demonstrated
that HN1 was upregulated in PDAC samples, which indicated
that PDAC occurrence may be related to HN1 overexpression.
This result is consistent with the previously studied mechanism of
action of HN1 (49).
Conversely, RT-qPCR analysis of HN1 did not demonstrate a
significant difference between PDAC samples and normal pancreatic
tissue. Despite the lack of a statistical difference between PDAC
patients with high levels and low HN1 expression levels in
the TCGA dataset, since the present study suggested that HN1
expression is associated with PDAC, further exploration of the
association between HN1 gene expression, PDAC progression
and patient survival is important.
ITGA2 is a collagen receptor expressed on
cell membranes that mediates cell-cell and cell-matrix adhesions. A
number of previous studies have indicated that ITGA2
expression is upregulated in normal epithelial cells, and its
expression changes when tumorigenesis occurs (51–53).
Ramirez et al revealed a loss of ITG α2β1 in breast and
prostate cancer (51). Previous
studies have also reported that loss of ITGA2 serves a
critical role in colon cancer metastasis (52) and that ITGA2 is strongly
expressed in PDAC (53). The
results of the present study are consistent with previous studies
on pancreatic cancer in that ITGA2 expression was
significantly higher in PDAC tissue compared with normal pancreatic
tissue.
S100A6, or its product calcyclin, has been
identified to be associated with various malignancies, such as
osteosarcoma (54), gastric cancer
(55) and malignant melanoma
(56). Upregulation of
S100A6 expression in tumor tissue is associated with cell
proliferation (57). In addition,
Ohuchida et al quantitatively analyzed S100A6 and
demonstrated that the S100A6 expression level was markedly
higher in pancreatic cancer tissues compared with normal pancreatic
tissue (58). S100A6 is one
of the 158 PDAC-related DEGs identified by Logsdon et al
using microarray datasets (59).
In the present study, S100A6 expression was upregulated in
PDAC samples compared with normal tissue, which was verified by
TCGA and RT-qPCR; the findings were consistent with previous
studies. Unfortunately, no association was observed between
S100A6 gene expression and patient survival time in the TCGA
dataset.
In the present study, KIF1A was identified as
the most strongly downregulated gene, with a combined ES of
−1.9848. The protein encoded by KIF1A is
microtubule-dependent molecular motor involved in important
intracellular functions such as organelle transport and cell
division (60). De et al
(61) found that KIF1A is highly
expressed in breast cancer and its upregulation is associated with
breast cancer resistance to docetaxel. By contrast, Hattori et
al demonstrated that KIF1A expression was upregulated in
human and murine adrenal tumors compared with normal adrenal tissue
(62). However, few studies have
shown that the PDAC occurrence is related to the downregulation of
KIF1A, although mutation of KIF1A is strongly
associated with cancer (63). In
the present study, KIF1A was the most strongly downregulated
DEG in PDAC, further indicating that KIF1A may be related to
the disease.
DYM encodes a Golgi-related protein
transported within cells, and previous studies have shown that
DYM loss of function is closely related to microcephaly in
Dyggve-Melchior-Clausen syndrome (64,65).
In the present study, DYM was the second most downregulated
gene, with a combined ES value of −1.8453, which suggested that it
may have potential protective functions against PDAC. KIF1A
and DYM expression levels were validated by TCGA, although
no significant differences between PDAC and normal tissues were
observed. In addition, RT-qPCR analysis of KIF1A and
DYM was performed multiple times. The expression of these
two genes could not be detected in some normal or PDAC tissues,
potentially owing to sub-threshold expression levels of these two
genes, or high sensitivity of the genes to pancreatin, leading to a
certain extent of degradation. Additional specimens need to be
collected for further study.
Overexpression of BACE1, which hydrolyzes
transmembrane amyloid precursor protein into amyloid-b protein, has
been demonstrated to serve an important role in Alzheimer's disease
(66). Excessive amyloid-b protein
aggregation damages neurons (67).
Chen et al reported that the long non-coding RNA BACE1
antisense RNA (BACE1-AS) may be used as a novel target of
anisomycin to inhibit the proliferation and invasion of ovarian
cancer stem cells; the authors also demonstrated that the increased
BACE1 levels caused by silencing of BACE1-AS resulted in
poor cancer suppression by anisomycin (68). Therefore, BACE1 may be a
tumor suppressor. In the present study, BACE1 was
downregulated in PDAC compared with normal pancreatic tissues.
Nonetheless, the relationship between BACE1 and cancer,
especially in PDAC, requires further study.
The top 10 upregulated and downregulated genes were
validated by performing a meta-analysis on a dataset from TCGA. The
results demonstrated that high expression levels of ITGA2,
KRT19, MBOAT2, and MGLL were associated with shorter
survival times, whereas high FAM46C expression was
associated with longer survival times. Dong et al (69) reported that ITGA2 is associated
with the tumor, node and metastasis classification system and
staging of gastric cancer, and that the expression level of ITGA2
was increased in metastatic lymph nodes and distant metastases.
They demonstrated that increased ITGA2 levels were associated with
reduced overall survival rates in patients with gastric cancer.
Previous studies have shown that KRT19 is a marker associated with
metastasis and poor prognosis of pancreatic ductal adenocarcinoma
(70,71). Badea et al (35) combined gene expression analysis of
whole-tissue and microdissected PDAC, and found that upregulation
of MBOAT2 is inversely correlated with patient survival.
Transcriptomic analyses identified that MGLL overexpression is an
unfavorable prognostic marker in primary gastrointestinal stromal
tumors (72). The results of Caba
et al (73) suggested that
FAM46C was downregulated in patients with PDAC and could be used as
prognostic indicator in these patients. The present results are
consistent with previous studies and suggested that these genes may
be novel targets for diagnosing and treating PDAC.
Using NetworkAnalyst for PPI network analysis,
GRB2, with a degree of 90, and HDAC5, with a degree
of 45, were identified as two hub genes among the upregulated and
downregulated DEGs, respectively. The GRB2 protein contains one Src
homology 2 domain and is a linker protein in the tyrosine kinase
receptor signaling pathway (74).
GRB2 expression directly affects cell proliferation and
differentiation (75). Liang et
al found that GRB2 expression was higher in
hepatocellular carcinoma compared with normal liver tissue and that
high GRB2 expression levels were associated with shorter
survival times (76). This is
consistent with the GRB2 expression profile in PDAC
described in the present study, although the expression level of
GRB2 did not appear to be significantly associated with overall
survival rates in the present study. In addition, a previous study
has confirmed that Grb2 may be used as a therapeutic target for
pancreatic cancer (77). Wang
et al demonstrated that GRB2 is a target of miR-329
through the GRB2/pERK pathway, which may lead to
novel therapies for pancreatic cancer (78). The results of these studies verify
the accuracy and credibility of the results of the present study
and further demonstrate that GRB2 is a potential target for
treating pancreatic cancer.
Histone deacetylases are epigenetic regulators that
suppress transcription by acting on gene promoters. HDAC1,
HDAC2, HDAC3 and HDAC8 are targets of multiple
epigenetic inhibitors, such as the REST corepressor 1, the
nucleosome remodeling deacetylase and the SIN3 histone deacetylase
complexes (79). A number of
studies have reported high HDAC5 expression in various
tumors, including medulloblastomas (GSE15471, GSE16515, GSE22780,
GSE28735, GSE32676, GSE43288-GPL96, GSE43288-GPL97, GSE43795,
GSE46234, GSE55643 and GSE62165.) (80), breast cancer (81) and pancreatic neuroendocrine tumors
(82). In addition, HDAC5
overexpression is associated with tumor cell proliferation and
invasion, as well as poor prognosis (80–82).
He et al demonstrated that HDAC5 is overexpressed in
and promotes the proliferation of colorectal cancer cells by
upregulating delta-like canonical Notch ligand 4 expression
(83). However, Özdağ et al
reported the opposite finding that, with the exception of high
HDAC5 expression levels in two samples of rectal cancer,
HDAC5 expression in all other rectal cancers was markedly
downregulated and that HDAC5 expression could be used to
distinguish between rectal tumors and normal tissues (84). These results are similar to the
results of the present study, which demonstrated that HDAC5
was downregulated in PDAC tissues compared with normal tissues.
Overall, findings to date demonstrate that HDAC5 may serve a
key role in tumorigenesis, but its role in PDAC requires further
study.
In conclusion, 1,587 DEGs were identified in the
present meta-analysis. HN1, ITGA2 and S100A6 were the
top upregulated genes and may be promising potential targets for
diagnosing and treating PDAC. HN1, which had the highest
combined ES, has rarely been reported as being important in PDAC;
thus, this gene should be further investigated. KIF1A, DYM
and BACE1 were the most markedly downregulated genes in this
study, providing new perspectives regarding PDAC mechanisms and
targeted treatment. GRB2 and HDAC5 were identified as
hub genes, serving the most pivotal roles in PDAC. However, a lack
of adequate validation in vitro or in vivo is a
limitation of this study. As the number of PDAC cases in the TCGA
database is limited, the present results need to be confirmed by
further experiments. Therefore, future research will include
experimental verification of the meta-analysis results using
immunohistochemistry and cell proliferation assays, and
identification of genes associated with PDAC progression by
screening differentially expressed genes at different stages.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81371661 and
81873895).
Availability of data and materials
The datasets used and analyzed during the present
study are available from GEO GSE15471, GSE16515, GSE22780,
GSE28735, GSE32676, GSE43288-GPL96, GSE43288-GPL97, GSE43795,
GSE46234, GSE55643 and GSE62165 (http://www.ncbi.nlm.nih.gov/geo) and TCGA (https://tcga-data.nci.Nih.gov/tcga/).
Authors' contributions
LL analyzed the results. LL and SW wrote the
manuscript. PH and SW designed the study. CC performed the
experiments. SP, YC and XL conceived the study and perform
statistical analyses. ND, QL and LM collected and analyzed the
data. All authors approved the manuscript for publication and agree
to be accountable for all aspects of the research.
Ethics approval and consent to
participate
Human study protocols were approved by the ethics
committee of Tongji Medical College, Huazhong University of Science
and Technology (Wuhan, China), in accordance with the Declaration
of Helsinki (reference no. S298). All patients in the study
provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Crane CH, Varadhachary GR, Yordy JS,
Staerkel GA, Javle MM, Safran H, Haque W, Hobbs BD, Krishnan S,
Fleming JB, et al: Phase II trial of cetuximab, gemcitabine, and
oxaliplatin followed by chemoradiation with cetuximab for locally
advanced (T4) pancreatic adenocarcinoma: Correlation of Smad4(Dpc4)
immunostaining with pattern of disease progression. J Clin Oncol.
29:3037–3043. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hezel AF, Kimmelman AC, Stanger BZ,
Bardeesy N and Depinho RA: Genetics and biology of pancreatic
ductal adenocarcinoma. Genes Dev. 20:1218–1249. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gillen S, Schuster T, Meyer Zum
Büschenfelde C, Friess H and Kleeff J: Preoperative/neoadjuvant
therapy in pancreatic cancer: A systematic review and meta-analysis
of response and resection percentages. PLoS Med. 7:e10002672010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stratton MR, Campbell PJ and Futreal PA:
The cancer genome. Nature. 458:719–724. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jones S, Zhang X, Parsons DW, Lin JC,
Leary RJ, Angenendt P, Mankoo P, Carter H, Kamiyama H, Jimeno A, et
al: Core signaling pathways in human pancreatic cancers revealed by
global genomic analyses. Science. 321:1801–1806. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nagata K, Horinouchi M, Saitou M, Higashi
M, Nomoto M, Goto M and Yonezawa S: Mucin expression profile in
pancreatic cancer and the precursor lesions. J Hepatobiliary
Pancreat Surg. 14:243–254. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sausen M, Phallen J, Adleff V, Jones S,
Leary RJ, Barrett MT, Anagnostou V, Parpart-Li S, Murphy D, Kay Li
Q, et al: Clinical implications of genomic alterations in the
tumour and circulation of pancreatic cancer patients. Nat Commun.
6:76862015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rozenblum E, Schutte M, Goggins M, Hahn
SA, Panzer S, Zahurak M, Goodman SN, Sohn TA, Hruban RH, Yeo CJ and
Kern SE: Tumor-suppressive pathways in pancreatic carcinoma. Cancer
Res. 57:1731–1734. 1997.PubMed/NCBI
|
|
10
|
Bardeesy N, Cheng KH, Berger JH, Chu GC,
Pahler J, Olson P, Hezel AF, Horner J, Lauwers GY, Hanahan D and
DePinho RA: Smad4 is dispensable for normal pancreas development
yet critical in progression and tumor biology of pancreas cancer.
Genes Dev. 20:3130–3146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fong ZV and Winter JM: Biomarkers in
pancreatic cancer: Diagnostic, prognostic, and predictive. Cancer
J. 18:530–538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Poruk KE, Gay DZ, Brown K, Mulvihill JD,
Boucher KM, Scaife CL, Firpo MA and Mulvihill SJ: The clinical
utility of CA 19-9 in pancreatic adenocarcinoma: Diagnostic and
prognostic updates. Curr Mol Med. 13:340–351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim SM, Kwon IJ, Myoung H, Lee JH and Lee
SK: Identification of human papillomavirus (HPV) subtype in oral
cancer patients through microarray technology. Eur Arch
Otorhinolaryngol. 275:535–543. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gunel T, Hosseini MK, Gumusoglu E,
Kisakesen HI, Benian A and Aydinli K: Expression profiling of
maternal plasma and placenta microRNAs in preeclamptic pregnancies
by microarray technology. Placenta. 52:77–85. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Qiao X, Wang H, Wang X, Zhao B and Liu J:
Microarray technology reveals potentially novel genes and pathways
involved in non-functioning pituitary adenomas. Balkan J Med Genet.
19:5–16. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li G, Li X, Yang M, Xu L, Deng S and Ran
L: Prediction of biomarkers of oral squamous cell carcinoma using
microarray technology. Sci Rep. 7:421052017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J, Chen Z, Tian L, Zhou C, He MY, Gao
Y, Wang S, Zhou F, Shi S, Feng X, et al: LncRNA profile study
reveals a three-lncRNA signature associated with the survival of
patients with oesophageal squamous cell carcinoma. Gut.
63:1700–1710. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ramasamy A, Mondry A, Holmes CC and Altman
DG: Key issues in conducting a meta-analysis of gene expression
microarray datasets. PLoS Med. 5:e1842008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rung J and Brazma A: Reuse of public
genome-wide gene expression data. Nat Rev Genet. 14:89–99. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang S, Jin F, Fan W, Liu F, Zou Y, Hu X,
Xu H and Han P: Gene expression meta-analysis in diffuse low-grade
glioma and the corresponding histological subtypes. Sci Rep.
7:117412017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Y, Xia Q and Lin J: Identification
of the potential oncogenes in glioblastoma based on bioinformatic
analysis and elucidation of the underlying mechanisms. Oncol Rep.
40:715–725. 2018.PubMed/NCBI
|
|
22
|
Xiao W, Pacyna-Gengelbach M, Schluns K, An
Q, Gao Y, Cheng S and Petersen I: Differentially expressed genes
associated with human lung cancer. Oncol Rep. 14:229–234.
2005.PubMed/NCBI
|
|
23
|
Tang F, He Z, Lei H, Chen Y, Lu Z, Zeng G
and Wang H: Identification of differentially expressed genes and
biological pathways in bladder cancer. Mol Med Rep. 17:6425–6434.
2018.PubMed/NCBI
|
|
24
|
Peng C, Ma W, Xia W and Zheng W:
Integrated analysis of differentially expressed genes and pathways
in triplenegative breast cancer. Mol Med Rep. 15:1087–1094. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun W, Ma X, Shen J, Yin F, Wang C and Cai
Z: Bioinformatics analysis of differentially expressed pathways
related to the metastatic characteristics of osteosarcoma. Int J
Mol Med. 38:466–474. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hoshida Y, Nijman SM, Kobayashi M, Chan
JA, Brunet JP, Chiang DY, Villanueva A, Newell P, Ikeda K,
Hashimoto M, et al: Integrative transcriptome analysis reveals
common molecular subclasses of human hepatocellular carcinoma.
Cancer Res. 69:7385–7392. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tang Y, Zhang Z, Tang Y, Chen X and Zhou
J: Identification of potential target genes in pancreatic ductal
adenocarcinoma by bioinformatics analysis. Oncol Lett.
16:2453–2461. 2018.PubMed/NCBI
|
|
28
|
Amin MB, Edge S, Greene F, Byrd DR,
Brookland RK, Washington MK, Gershenwald JE, Compton CC, Hess KR,
Sullivan DC, et al: AJCC Cancer Staging Manual. 8th. Springer
International Publishing; New York, NY: 2017, View Article : Google Scholar
|
|
29
|
Xia J, Fjell CD, Mayer ML, Pena OM,
Wishart DS and Hancock RE: INMEX-a web-based tool for integrative
meta-analysis of expression data. Nucleic Acids Res. 41:W63–W70.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Johnson WE, Li C and Rabinovic A:
Adjusting batch effects in microarray expression data using
empirical bayes methods. Biostatistics. 8:118–127. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cochran WG: The combination of estimates
from different experiments. Biometrics. 10:101–129. 1954.
View Article : Google Scholar
|
|
32
|
Marot G, Foulley JL, Mayer CD and
Jaffrézic F: Moderated effect size and P-value combinations for
microarray meta-analyses. Bioinformatics. 25:2692–2699. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Breuer K, Foroushani AK, Laird MR, Chen C,
Sribnaia A, Lo R, Winsor GL, Hancock RE, Brinkman FS and Lynn DJ:
InnateDB: Systems biology of innate immunity and beyond-recent
updates and continuing curation. Nucleic Acids Res. 41:D1228–D1233.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Badea L, Herlea V, Dima SO, Dumitrascu T
and Popescu I: Combined gene expression analysis of whole-tissue
and microdissected pancreatic ductal adenocarcinoma identifies
genes specifically overexpressed in tumor epithelia.
Hepatogastroenterology. 55:2016–2027. 2008.PubMed/NCBI
|
|
36
|
Pei H, Li L, Fridley BL, Jenkins GD,
Kalari KR, Lingle W, Petersen G, Lou Z and Wang L: FKBP51 affects
cancer cell response to chemotherapy by negatively regulating Akt.
Cancer Cell. 16:259–266. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang G, Schetter A, He P, Funamizu N,
Gaedcke J, Ghadimi BM, Ried T, Hassan R, Yfantis HG, Lee DH, et al:
DPEP1 inhibits tumor cell invasiveness, enhances chemosensitivity
and predicts clinical outcome in pancreatic ductal adenocarcinoma.
PLos One. 7:e315072012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang G, He P, Tan H, Budhu A, Gaedcke J,
Ghadimi BM, Ried T, Yfantis HG, Lee DH, Maitra A, et al:
Integration of metabolomics and transcriptomics revealed a fatty
acid network exerting growth inhibitory effects in human pancreatic
cancer. Clin Cancer Res. 19:4983–4993. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Donahue TR, Tran LM, Hill R, Li Y,
Kovochich A, Calvopina JH, Patel SG, Wu N, Hindoyan A, Farrell JJ,
et al: Integrative survival-based molecular profiling of human
pancreatic cancer. Clin Cancer Res. 18:1352–1363. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Crnogorac-Jurcevic T, Chelala C, Barry S,
Harada T, Bhakta V, Lattimore S, Jurcevic S, Bronner M, Lemoine NR
and Brentnall TA: Molecular analysis of precursor lesions in
familial pancreatic cancer. PLos One. 8:e548302013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Park M, Kim M, Hwang D, Park M, Kim WK,
Kim SK, Shin J, Park ES, Kang CM, Paik YK and Kim H:
Characterization of gene expression and activated signaling
pathways in solid-pseudopapillary neoplasm of pancreas. Mod Pathol.
27:580–593. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lunardi S, Jamieson NB, Lim SY, Griffiths
KL, Carvalho-Gaspar M, Al-Assar O, Yameen S, Carter RC, McKay CJ,
Spoletini G, et al: IP-10/CXCL10 induction in human pancreatic
cancer stroma influences lymphocytes recruitment and correlates
with poor survival. Oncotarget. 5:11064–11080. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Janky R, Binda MM, Allemeersch J, Van den
Broeck A, Govaere O, Swinnen JV, Roskams T, Aerts S and Topal B:
Prognostic relevance of molecular subtypes and master regulators in
pancreatic ductal adenocarcinoma. BMC Cancer. 16:6322016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jha PK, Vijay A, Sahu A and Ashraf MZ:
Comprehensive gene expression meta-analysis and integrated
bioinformatic approaches reveal shared signatures between
thrombosis and myeloproliferative disorders. Sci Rep. 6:370992016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang X, Ning Y and Guo X: Integrative
meta-analysis of differentially expressed genes in osteoarthritis
using microarray technology. Mol Med Rep. 12:3439–3445. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Han H, Bearss DJ, Browne LW, Calaluce R,
Nagle RB and Von Hoff DD: Identification of differentially
expressed genes in pancreatic cancer cells using cDNA microarray.
Cancer Res. 62:2890–2896. 2002.PubMed/NCBI
|
|
47
|
Lu KH, Patterson AP, Wang L, Marquez RT,
Atkinson EN, Baggerly KA, Ramoth LR, Rosen DG, Liu J, Hellstrom I,
et al: Selection of potential markers for epithelial ovarian cancer
with gene expression arrays and recursive descent partition
analysis. Clin Cancer Res. 10:3291–3300. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Laughlin KM, Luo D, Liu C, Shaw G,
Warrington KH Jr, Qiu J, Yachnis AT and Harrison JK: Hematopoietic-
and neurologic-expressed sequence 1 expression in the murine GL261
and high-grade human gliomas. Pathol Oncol Res. 15:437–444. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang C, Xu B, Lu S, Zhao Y and Liu P: HN1
contributes to migration, invasion, and tumorigenesis of breast
cancer by enhancing MYC activity. Mol Cancer. 16:902017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Varisli L, Ozturk BE, Akyuz GK and Korkmaz
KS: HN1 negatively influences the β-catenin/E-cadherin interaction,
and contributes to migration in prostate cells. J Cell Biochem.
116:170–178. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ramirez NE, Zhang Z, Madamanchi A, Boyd
KL, O'Rear LD, Nashabi A, Li Z, Dupont WD, Zijlstra A and Zutter
MM: The α2β1 integrin is a metastasis suppressor in mouse models
and human cancer. J Clin Invest. 121:226–237. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Robertson JH, Yang SY, Winslet MC and
Seifalian AM: Functional blocking of specific integrins inhibit
colonic cancer migration. Clin Exp Metastasis. 26:769–780. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shimoyama S, Gansauge F, Gansauge S,
Oohara T and Beger HG: Altered expression of extracellular matrix
molecules and their receptors in chronic pancreatitis and
pancreatic adenocarcinoma in comparison with normal pancreas. Int J
Pancreatol. 18:227–234. 1995.PubMed/NCBI
|
|
54
|
Li Y, Wagner ER, Yan Z, Wang Z, Luther G,
Jiang W, Ye J, Wei Q, Wang J, Zhao L, et al: The calcium-binding
protein S100A6 accelerates human osteosarcoma growth by promoting
cell proliferation and inhibiting osteogenic differentiation. Cell
Physiol Biochem. 37:2375–2392. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wang XH, Zhang LH, Zhong XY, Xing XF, Liu
YQ, Niu ZJ, Peng Y, Du H, Zhang GG, Hu Y, et al: S100A6
overexpression is associated with poor prognosis and is
epigenetically up-regulated in gastric cancer. Am J Pathol.
177:586–597. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Maelandsmo GM, Florenes VA, Mellingsaeter
T, Hovig E, Kerbel RS and Fodstad O: Differential expression
patterns of S100A2, S100A4 and S100A6 during progression of human
malignant melanoma. Int J Cancer. 74:464–469. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
He X, Xu X, Khan AQ and Ling W: High
expression of S100A6 predicts unfavorable prognosis of lung
squamous cell cancer. Med Sci Monit. 23:5011–5017. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Ohuchida K, Mizumoto K, Ishikawa N, Fujii
K, Konomi H, Nagai E, Yamaguchi K, Tsuneyoshi M and Tanaka M: The
role of S100A6 in pancreatic cancer development and its clinical
implication as a diagnostic marker and therapeutic target. Clin
Cancer Res. 11:7785–7793. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Logsdon CD, Simeone DM, Binkley C,
Arumugam T, Greenson JK, Giordano TJ, Misek DE, Kuick R and Hanash
S: Molecular profiling of pancreatic adenocarcinoma and chronic
pancreatitis identifies multiple genes differentially regulated in
pancreatic cancer. Cancer Res. 63:2649–2657. 2003.PubMed/NCBI
|
|
60
|
Okada Y, Yamazaki H, Sekine-Aizawa Y and
Hirokawa N: The neuron-specific kinesin superfamily protein KIF1A
is a unique monomeric motor for anterograde axonal transport of
synaptic vesicle precursors. Cell. 81:769–780. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
De S, Cipriano R, Jackson MW and Stark GR:
Overexpression of kinesins mediates docetaxel resistance in breast
cancer cells. Cancer Res. 69:8035–8042. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hattori Y, Kanamoto N, Kawano K, Iwakura
H, Sone M, Miura M, Yasoda A, Tamura N, Arai H, Akamizu T, et al:
Molecular characterization of tumors from a transgenic mouse
adrenal tumor model: Comparison with human pheochromocytoma. Int J
Oncol. 37:695–705. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Brait M, Loyo M, Rosenbaum E, Ostrow KL,
Markova A, Papagerakis S, Zahurak M, Goodman SM, Zeiger M,
Sidransky D, et al: Correlation between BRAF mutation and promoter
methylation of TIMP3, RARbeta2 and RASSF1A in thyroid cancer.
Epigenetics. 7:710–719. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Osipovich AB, Jennings JL, Lin Q, Link AJ
and Ruley HE: Dyggve-Melchior-Clausen syndrome: Chondrodysplasia
resulting from defects in intracellular vesicle traffic. Proc Natl
Acad Sci U S A. 105:16171–16176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Dimitrov A, Paupe V, Gueudry C, Sibarita
JB, Raposo G, Vielemeyer O, Gilbert T, Csaba Z, Attie-Bitach T,
Cormier-Daire V, et al: The gene responsible for
Dyggve-Melchior-Clausen syndrome encodes a novel peripheral
membrane protein dynamically associated with the Golgi apparatus.
Hum Mol Genet. 18:440–453. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Shi Z, Hong Y, Zhang K, Wang J, Zheng L,
Zhang Z, Hu Z, Han X, Han Y, Chen T, et al: BAG-1M co-activates
BACE1 transcription through NF-κB and accelerates Aβ production and
memory deficit in Alzheimer's disease mouse model. Biochim Biophys
Acta Mol Basis Dis. 1863:2398–2407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yi X, Hao Y, Xia N, Wang J, Quintero M, Li
D and Zhou F: Sensitive and continuous screening of inhibitors of
β-site amyloid precursor protein cleaving enzyme 1 (BACE1) at
single SPR chips. Anal Chem. 85:3660–3666. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen Q, Liu X, Xu L, Wang Y, Wang S, Li Q,
Huang Y and Liu T: Long non-coding RNA BACE1-AS is a novel target
for anisomycin-mediated suppression of ovarian cancer stem cell
proliferation and invasion. Oncol Rep. 35:1916–1924. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Dong J, Wang R, Ren G, Li X, Wang J, Sun
Y, Liang J, Nie Y, Wu K, Feng B, et al: HMGA2-FOXL2 axis regulates
metastases and epithelial-to-mesenchymal transition of
chemoresistant gastric cancer. Clin Cancer Res. 23:3461–3473. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yao H, Yang Z, Liu Z, Miao X, Yang L, Li
D, Zou Q and Yuan Y: Glypican-3 and KRT19 are markers associating
with metastasis and poor prognosis of pancreatic ductal
adenocarcinoma. Cancer Biomark. 17:397–404. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Dugnani E, Sordi V, Pellegrini S,
Chimienti R, Marzinotto I, Pasquale V, Liberati D, Balzano G,
Doglioni C, Reni M, et al: Gene expression analysis of embryonic
pancreas development master regulators and terminal cell fate
markers in resected pancreatic cancer: A correlation with clinical
outcome. Pancreatology. 18:945–953. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Li CF, Chuang IC, Liu TT, Chen KC, Chen
YY, Fang FM, Li SH, Chen TJ, Yu SC, Lan J and Huang HY:
Transcriptomic reappraisal identifies MGLL overexpression as an
unfavorable prognosticator in primary gastrointestinal stromal
tumors. Oncotarget. 7:49986–49997. 2016.PubMed/NCBI
|
|
73
|
Caba O, Irigoyen A, Jimenez-Luna C,
Benavides M, Ortuño FM, Gallego J, Rojas I, Guillen-Ponce C, Torres
C, Aranda E and Prados J: Identification of gene expression
profiling associated with erlotinib-related skin toxicity in
pancreatic adenocarcinoma patients. Toxicol Appl Pharmacol.
311:113–116. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wintgens JP, Wichert SP, Popovic L,
Rossner MJ and Wehr MC: Monitoring activities of receptor tyrosine
kinases using a universal adapter in genetically encoded split TEV
assays. Cell Mol Life Sci. 76:1185–1199. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Dharmawardana PG, Peruzzi B, Giubellino A,
Burke TR Jr and Bottaro DP: Molecular targeting of growth factor
receptor-bound 2 (Grb2) as an anti-cancer strategy. Anticancer
Drugs. 17:13–20. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liang C, Xu Y, Ge H, Xing B, Li G, Li G
and Wu J: miR-564 inhibits hepatocellular carcinoma cell
proliferation and invasion by targeting the GRB2-ERK1/2-AKT axis.
Oncotarget. 8:107543–107557. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Birchmeier C, Birchmeier W, Gherardi E and
Vande Woude GF: Met, metastasis, motility and more. Nat Rev Mol
Cell Biol. 4:915–925. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Wang X, Lu X, Zhang T, Wen C, Shi M, Tang
X, Chen H, Peng C, Li H, Fang Y, et al: mir-329 restricts tumor
growth by targeting grb2 in pancreatic cancer. Oncotarget.
7:21441–21453. 2016.PubMed/NCBI
|
|
79
|
Witt O, Deubzer HE, Milde T and Oehme I:
HDAC family: What are the cancer relevant targets? Cancer Lett.
277:8–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Milde T, Oehme I, Korshunov A,
Kopp-Schneider A, Remke M, Northcott P, Deubzer HE, Lodrini M,
Taylor MD, von Deimling A, et al: HDAC5 and HDAC9 in
medulloblastoma: Novel markers for risk stratification and role in
tumor cell growth. Clin Cancer Res. 16:3240–3252. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Li A, Liu Z, Li M, Zhou S, Xu Y, Xiao Y
and Yang W: HDAC5, a potential therapeutic target and prognostic
biomarker, promotes proliferation, invasion and migration in human
breast cancer. Oncotarget. 7:37966–37978. 2016.PubMed/NCBI
|
|
82
|
Klieser E, Urbas R, Stättner S, Primavesi
F, Jäger T, Dinnewitzer A, Mayr C, Kiesslich T, Holzmann K, Di
Fazio P, et al: Comprehensive immunohistochemical analysis of
histone deacetylases in pancreatic neuroendocrine tumors: HDAC5 as
a predictor of poor clinical outcome. Hum Pathol. 65:41–52. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
He P, Liang J, Shao T, Guo Y, Hou Y and Li
Y: HDAC5 promotes colorectal cancer cell proliferation by
up-regulating DLL4 expression. Int J Clin Exp Med. 8:6510–6516.
2015.PubMed/NCBI
|
|
84
|
Ozdağ H, Teschendorff AE, Ahmed AA, Hyland
SJ, Blenkiron C, Bobrow L, Veerakumarasivam A, Burtt G,
Subkhankulova T, Arends MJ, et al: Differential expression of
selected histone modifier genes in human solid cancers. BMC
Genomics. 7:902006. View Article : Google Scholar : PubMed/NCBI
|



















