Introduction
Ischemic stroke refers to cerebral ischemia due to
occlusion of a cerebral blood vessels, and has a high rate of
mortality and morbidity (1). A
cross-sectional study conducted in 2012–2013 demonstrated that the
age-standardized incidence for stroke in China were 247/100,000
(95% CI, 211–283), and ischemic stroke accounted for ~60% among
new-onset stroke patients (2).
According to the ‘China Health and Family Planning Statistical
Yearbook 2016’ (3), in 2015, the
mortality of ischemic stroke was about 46.99 in 100,000 in the
rural areas and 41.82 in 100,000 in the urban areas of China. The
hippocampal injury is aggravated due to the lack of blood flow and
oxygen deprivation, and cerebral ischemia/reperfusion injury (CIRI)
is caused by subsequent restoration of blood flow (4). The mechanisms underlying the
occurrence of CIRI are complicated and involve interactions between
multiple factors. For example, CIRI causes tissue hypoxia, leading
to reduced mitochondrial ATP synthesis, ion channel disorder,
impaired enzyme activity and redox imbalance. These effects in turn
may affect brain tissue oxygen uptake (5). In addition, the pathophysiological
processes of CIRI are complicated, involving oxidative stress,
inflammation, apoptosis and excitotoxicity (6–8).
With the interruption in cerebral blood flow, accumulation of
glutamate (Glu) is observed, and Glu-induced excitotoxicity
involves consumption of intracellular glutathione (9). Along with the depletion of the
endogenous antioxidant glutathione, excessive reactive oxygen
species (ROS) accumulation can occur, resulting in increased
oxidative stress (10). Severe
oxidative stress may induce DNA damage and cell apoptosis, which
affects neurological function (9).
Therefore, the development of novel and effective antioxidants or
therapeutic strategies to maintain the cellular redox homeostasis
is urgently required.
Piceatannol (Pic), a natural stilbene found
primarily in seeds, wines and fruits (11), has been reported to exhibit
multiple protective effects on a variety of diseases, such as
metabolic cardiomyopathy and ischemic cardiomyopathy (12,13).
An increasing number of studies have revealed that Pic serves a
crucial role in regulating a variety of physiological processes due
to its anti-inflammatory capacity and potent antioxidant properties
(14,15). Additionally, Pic has been shown to
participate in a variety of physiological and pathophysiological
processes through the modulation of a range of pathways, which
enables cells to respond to a diverse array of stimuli. Pic
inhibited oxidative stress by activating the PI3K/Akt pathway,
ultimately alleviating macular retinal pigment epithelial cells
degeneration in mice in age-related macular degeneration (11). Moreover, Pic alleviated
inflammation and oxidative stress by regulating the Nrf2/HO-1 and
NF-κB pathways in diabetic cardiomyopathy (13).
The sirtuin 1 (Sirt1)/FoxO1 pathway is a vital
signaling pathway considered to be involved in regulating oxidative
stress, the inflammatory reaction and apoptosis (16). A recent in vitro study
observed that activation of the Sirt1/FoxO1 pathway can alleviate
neuron injury in an oxygen-glucose deprivation/reperfusion model,
which may be due to the antioxidant activity and anti-apoptotic
effects of the downregulation of this pathway (17).
Previous studies have reported that the beneficial
effects of Pic may be due to the action of Sirt1, which contributes
to upregulation of the antioxidative and anti-apoptotic factors
against early-stage nephropathy and hepatic inflammation (18,19).
Although Pic has been demonstrated to exert protective properties
in several cell types and different disease processes, such as
diabetic cardiomyopathy and diabetic cardiomyopathy (12,13),
to the best of our knowledge, no studies have further examined its
effects on CIRI, as well as the potential underlying molecular
mechanisms. Furthermore, neuronal apoptosis, which has also been
proposed as an important mechanism affecting neurologic function,
may serve an important role in the pharmacodynamic processes
(20). In combination with other
evidence, the present study aimed to investigate the antioxidative
stress damage and anti-apoptotic capability of Pic in vitro
and in vivo, as well as to identified the possible
involvement of certain signaling pathways.
Materials and methods
Animals and CIRI model
The animal experiments were approved by the Ethics
Committee of Tangshan Gongren Hospital (approval no.
GRYY-LL-2019-13). For the experiments, male C57BL/6 mice (age, 8
weeks; weight, 20–25 g; 60 mice in total) were obtained from the
Experimental Animal Center, North China University of Science and
Technology. The mice had free access to food and water throughout
the entirety of the trial. The housing conditions were as follows:
20–24°C, 12 h light/dark cycle and 50±10% humidity.
Mice underwent a middle-cerebral artery occlusion
(MCAO) model of CIRI as previously described (21). Briefly, mice were anesthetized via
intraperitoneal injection of sodium pentobarbital (50 mg/kg).
During this stage, the body temperature of mice was maintained at
36.5–37.5°C. Regional cerebral blood flow (rCBF) of each mouse was
monitored using a Laser-Doppler flowmeter (Moor Instruments, Ltd.).
A right side incision in the mouse was performed and the carotid
arteries were separated, including the common carotid artery (CCA),
external carotid artery and internal carotid artery (ICA). After
carefully ligating the right CCA with a microclip, a silk thread
was inserted into the stump of the ICA. The monofilament was then
advanced ~12 mm so that its distal end came to rest across the
origin of the MCA. In order to minimize the differences in CIRI
degree, performance of the MCAO surgery was regarded as successful
if rCBF was decreased abruptly by 70–80%. After obturating blood
flow for 60 sec, the monofilament was then removed to re-establish
carotid blood flow. Apart from Laser Doppler flowmetry, Longa's
neurological scoring (22) was
used to determine whether the MCAO model was successfully
established. The animals with a score of 1–3 after MCAO were
included for analysis. Given the inconsistencies between vascular
occlusion degree and CIRI symptoms, mice with no or slight
neurological deficit were excluded. The Sham group underwent only
dissociation of carotid arteries without inserting a
monofilament.
During the course of establishment of the CIRI
model, mortality occurred in two mice due to disease conditions.
The mortality was lower compared with previous estimates, which
reported mortality rates up to ~10% (21). After the model was successfully
established, MCAO mice were randomly divided into CIRI, low-dose
Pic and high-dose Pic groups using a random-number table, with 14
to 15 mice in each group.
Experimental groups and treatment
C57BL/6 J mice were randomly divided into four
groups: Sham, CIRI, low (L)-dose Pic treatment (10 mg/kg/day) and
high (H)-dose Pic treatment (20 mg/kg/day). Pic was purchased from
Selleck Chemicals (cat. no. S3026) and suspended in 0.3 ml 0.5%
carboxymethyl cellulose solution prior to use. Mice were orally
administered L-and H-doses of Pic 1 h after establishment of CIRI,
and once daily for the next 6 days. Mice in the Sham and CIRI group
were treated with 0.3 ml 0.5% carboxymethyl cellulose solution by
oral administration. After treatment, behavioral performance and
pathological changes in brain hippocampal tissues of mice were
detected using a sequential process.
Ex-527 (cat. no. E7034; Sigma-Aldrich; Merck KGaA),
a Sirt1 inhibitor, was used for subsequent experiments. An
additional 60 mice were randomly assigned to the Sham, CIRI, CIRI +
Pic treatment (20 mg/kg/day) and CIRI + Pic + Ex-527 groups. A
total of 10 µg Ex-527 was dissolved in a 5 µl mixture of DMSO and
normal saline before use. Subsequently, Ex-527 (10 µg) was
administered via intracerebroventricular injection 1 h before MCAO.
Mice were placed in a stereotactic device after anesthetization. In
accordance with the stereotaxic atlas of Paxinos and Watson
(23), injections were made into
the left lateral brain ventricle (lateral, 1.3; dorsoventral, 4.0;
anteroposterior, −0.92 from Bregma and skull) using a
stainless-steel guide cannula. When the guide cannula was 1-cm
above the skull, it was secured using dental acrylic. If no
anomalies were found, a stainless-steel needle, with a diameter of
0.4 mm, was inserted directly into the lateral ventricle along the
guide cannula. Mice in the Sham and CIRI group were treated with
0.3 ml of 0.5% carboxymethyl cellulose solution by oral and 5 µl
mixture of DMSO and normal saline via intracerebroventricular
injection.
Rotarod performance test
In order to measure the effects of the drugs on
vestibulomotor function, an accelerating automated rotarod test
(RotaRod Treadmill; TSE Technical and Scientific Equipment) was
used. Each mouse performed three trials per day. Mice were placed
on a rotating rod that accelerated from 5–40 rpm during the 5-min
experimental trial. Data obtained before the onset of MCAO were
regarded as baseline values. Assessment of motor function was
obtained at days 1–5 after CIRI. As the rotation velocity
increased, the average time until the mice fell twice from the
rotating drum was recorded manually.
Morris water maze (MWM) test
In order to determine the spatial learning and
memory ability of the mice, MWM test was performed 1 week after
completion of Pic treatments. The mice were placed in a round water
pool (diameter=150 cm; height=50 cm) and allowed to swim freely in
the water for 5 min at the start of the positioning navigation
test. A hidden platform was set in the central area of the target
quadrant. At a fixed time of each training day, mice were placed
inside the tank to locate the hidden platform. At a designated
starting point in a different quadrant of the maze, each mouse was
placed randomly into the water. If the mouse succeeded in mounting
the hidden platform during the navigation test, the mouse would be
manually placed on the platform for 15 sec before proceeding to the
next training session.
The spatial exploration test, in which the hidden
platform was removed, was performed on the 5th day of the
experiment. Mice were placed gently into the pool at the same,
randomly chosen training points and the residence time of each
mouse in the target quadrant was recorded as ‘target quadrant
time’. Performance was recorded using a video camera connected to
tracking equipment from HVS Tracking System Ltd.
Neurological score
Modified neurological severity score (mNSS) grades
the composite neurological function of mice on sensory, motor,
reflex and beam balance tests. The mNSS score was assigned as
previously described (24). In the
present study, neurological functional measurement was performed on
mice within 1 week after CIRI establishment using the modified
mNSS. A higher score indicated worse neural defects (least
severe=0; most severe=18).
Hematoxylin and eosin (H&E)
staining
Briefly, after anesthetization and transcardial
perfusions, the mice were rapidly executed via decapitation and
hemispheres were removed. After fixing using 5% neutral buffered
formalin for 7 days at room temperature, the tissues were sliced
into 4-µm sections. The sample slices were stained with hematoxylin
for 2 min and with eosin for 30 sec. An optical microscope (Olympus
Corporation; magnification, ×400) was used to detect the
histological changes in the hippocampal tissues.
Western blotting
Hippocampal tissues were lysed using RIPA lysis
buffer (Thermo Fisher Scientific, Inc.). Protein concentrations
were determined using a BCA assay (OriGene Technologies, Inc.). A
total of 25 µg extracted protein was loaded on a 12% SDS-gel,
resolved using SDS-PAGE and electrophoretically transferred to a
PVDF membrane (Bio-Rad Laboratories, Inc.). The membranes were
blocked using 5% non-fat milk for 1.5 h at room temperature.
Subsequently, the membranes were washed three times with PBS/0.2%
Tween-20 (PBST) for 3 min each time, and incubated overnight at 4°C
with rabbit anti-mouse antibodies. The primary antibodies used
were: Bcl-2 (cat. no. ab196495; 1:1,000; rabbit polyclonal; Abcam),
Bax (cat. no. ab32503; 1:1,000; rabbit monoclonal; Abcam), CC-3
(cat. no. ab49822; 1:1,000; rabbit polyclonal; Abcam), Sirt1 (cat.
no. ab189494; 1:1,000; rabbit monoclonal; Abcam), FoxO1 (cat. no.
ab52857; 1:1,000; rabbit monoclonal; Abcam) and β-actin (cat. no.
ab8227; 1:1,000; rabbit polyclonal; Abcam). After washing three
times for 5 min using PBST, the PVDF membranes were incubated with
IRDye® 800CW goat anti-rabbit IgG secondary antibody
(cat. no. ab216773; 1:10,000; Abcam) at room temperature for 2 h.
Signals were visualized using enhanced chemiluminescent reagent
(Bio-Rad Laboratories, Inc.). Densitometry analysis was performed
using ImageJ (Image Lab 4.1; National Institutes of Health).
Reverse-transcription quantitative PCR
(RT-qPCR)
Cellular RNA extraction was performed using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. A PrimeScript™ RT
reagent kit (cat. no. RR037A; Takara Bio, Inc.) was used for cDNA
generation. The RT reactions were performed at 37°C for 15 min,
85°C for 5 sec, then 4°C for cooling. The RT-qPCR was conducted
with the TB Green® Premix Ex Taq™ II kit (cat. no.
RR820A; Takara Bio, Inc.) in a 20-µl reaction containing 10 µl 2X
TB Green Premix Ex Taq II (Tli RNaseH Plus), 0.8 µl forward primer
(10 µM), 0.8 µl reverse primer (10 µM), 2 µl template DNA and 6.4
µl ddH2O. The thermocycling conditions for RT were 37°C
for 30 min followed by inactivation at 95°C for 1 min. This was
followed by 40 cycles of two-step PCR; 95°C for 18 sec and 60°C for
55 sec, with a final extension step at 72°C for 2.5 min, and
subsequent holding at 4°C. All experiments were repeated ≥3 times.
The sequences of the primers were: β-actin forward,
5′-TGACGTGGACATCCGCAAAG-3′ and reverse, 5′-CTGGAAGGTGGACAGCGAGG-3′;
CC-3 forward, 5′-GAGCACTGGAATGTCATCTCGCTCTG-3′ and reverse,
5′-TACAGGAAGTCAGCCTCCACCGGTATC-3′; Bax forward,
5′-GTTGCCCTCTTCTACTTTGC-3′ and reverse, 5′-ATGGTCACTGTCTGCCATG-3′;
Bcl-2 forward, 5′-GGTCCTCCAGTGGGTATTT-3′ and reverse,
5′-TCCTCCTGAGACTGCCTTAT-3′; Sirt1 forward,
5′-AATCCAGTCATTAAACGGTCTACAA-3′ and reverse,
5′-TAGGACCATTACTGCCAGAGG-3′; and FoxO1 forward,
5′-TTGGATTCCTGCCTGCTGAT-3′ and reverse, 5′-CGCTTCTTGGGTTGATCTAG-3′.
β-actin was used as the internal control. The 2−ΔΔCq
method was used to measure the relative gene expression (25).
ROS, antioxidant enzymes and
non-enzymatic antioxidant detection
The procedures for detecting ROS production were
based on the protocols provided by Applygen Technologies, Inc.
(cat. no. C1300). After the hippocampal tissues were homogenized in
100 mM sodium phosphate buffer (1:20), the homogenate was
centrifuged at 1,000 × g for 10 min at 4°C. A total of 10 µl 1 mM
dichlorodihydrofluorescein diacetate was added to the supernatant.
Fluorescence intensity was quantified after 30 min of incubation at
37°C using a fluorospectrophotometer (excitation, 500 nm; emission,
525 nm).
In the antioxidant enzymes system, superoxide
dismutase (SOD) activity was determined spectrophotometrically
using a SOD assay kit (Cayman Chemical Company) according to the
manufacturer's protocol. At the beginning of the reaction, 20 µl
xanthine oxidase was injected into each well. The cells were
incubated for 20 min at room temperature. According to the
manufacturer's instructions (cat. no. A007-1-1; Nanjing Jiancheng
Bioengineering Institute), catalase (CAT) activity was determined
using the ammonium molybdate spectrophotometric method, which is
based on the decreased absorbance due to consumption of
H2O2 (26).
Glutathione peroxidase (GSH-Px) activity was measured as described
by Hafeman et al (27) in
accordance with the manufacturer's instructions (cat. no. A005-1-2;
Nanjing Jiancheng Bioengineering Institute). GSH-Px degrades
H2O2 in the presence of GSH, which results in
the decline of GSH. The remaining GSH reacts with
5,5′-dithiobis-2-nitrobenzoic acid (28). Absorbance was measured
spectrophotometrically at 440–460 nm for SOD, 405 nm for CAT and
412 nm for GSH-Px at the end of reaction on a microplate reader
(Infinite 200; Tecan Systems, Inc.).
For the non-enzymatic antioxidant system, homogenate
preparation of hippocampal tissue was performed as described
previously by Hissin and Hilf (28), and decreased GSH content was
detected. After mixing with 5% metaphosphoric acid, the supernatant
was centrifuged at 2,000 × g for 2 min at 4°C. Subsequently,
Tris-HCl buffer (0.26 M; pH 7.8), 0.65 N NaOH and O-phthalaldehyde
(1 mg/ml) was added, and the supernatant was left in the dark for
15 min at 4°C. A fluorescent microplate reader (Infinite 200; Tecan
Systems, Inc.) was used to detect the fluorescent intensity with an
excitation wavelength of 320 nm and emission wavelength of 420
nm.
TUNEL staining for detection of
neuronal apoptosis
Detection of neuronal apoptosis was performed as
described previously (29). For
brain tissue specimen collection from the cerebral ischemic area,
mice were transcardially perfused with cold PBS followed by 4%
paraformaldehyde in 0.1 M phosphate buffer. Subsequently, brain
tissues were extracted and fixed with 5% neutral buffered formalin
for 7 days at room temperature. The specimens were dehydrated with
gradient ethyl alcohol solutions and embedded in paraffin for 2 h.
TUNEL staining was performed using a TUNEL Apoptosis Assay Kit
(cat. no. C1088; Beyotime Institute of Biotechnology). After
sectioning, 5-µm sections were deparaffinized and rehydrated.
Following washing twice in 0.1 M PBS (2×5 min), the sections were
incubated with 10 µg/ml Proteinase K working solution (pH 7.5–8.0)
for 15 min at 37°C. Then, tissues were rinsed again in PBS, and the
slices were stained with green fluorescein-labeled dUTP solution
for 10 min at room temperature. Finally, the sections were mounted
with Vectashield® mounting medium containing DAPI for
nuclear labeling for 5 min at room temperature (Vector
Laboratories, Inc.). Using a fluorescence microscope (Olympus
Corporation), TUNEL-positive cells that exhibited green fluorescent
granules were detected. For quantification, five randomly selected
fields of view were selected (magnification, ×400). The percentage
of TUNEL positive cells per mm2 was determined. The
apoptotic ratio is expressed as the percentage of the
TUNEL-positive apoptotic neurons to the total number of DAPI
stained neurons.
Immunofluorescence analysis
Hippocampal tissues were harvested and dehydrated
with 15 and 30% gradient sucrose at 4°C. Tissues were frozen,
embedded in OCT compound (Sakura Finetek Japan, Co., Ltd.), then
cut to 15-µm frozen sections. After processing with 0.4% Triton
X-100 for 10 min, the frozen sections were blocked in 10% normal
donkey serum (cat. no. ab138579; Abcam) for 1 h at room
temperature. The sections were subsequently incubated with a
mixture of primary CC-3 antibody (cat. no. ab49822; 1:1,000; Abcam)
and neuronal nuclei (NeuN) monoclonal antibody (cat. no. ab177487;
1:1,000; Abcam) at 4°C overnight. Then, the cells were incubated
for 1 h at 37°C with Alexa Fluor® 488 goat anti-rabbit
IgG secondary antibodies (cat. no. ab150077; 1:1,000; Abcam) the
following day. Neurons were counterstained with DAPI for 10 min at
room temperature. Confocal fluorescence images were acquired using
an Olympus F1000 laser scanning confocal fluorescence microscope
(magnification, ×400).
Primary hippocampal neuron
culture
The embryos of 17-day pregnant Sprague-Dawley rats
(10 g) were used for primary hippocampal neuron culture studies.
The pregnant rats (3 months old, 280–350 g) were housed in
individual cages and maintained on a 12-h light/12-h dark cycle at
22±2°C and 65% humidity. All rats were allowed to take food and
water ad libitum. The pregnant rats with embryos were
anesthetized with isoflurane inhalation at dose of 4.5% for
induction and 2.5% for maintenance. Pregnant rats were euthanized
via cervical dislocation after the fetuses were removed. The
fetuses were euthanized by CO2 inhalation and rapid
decapitation (30). All efforts
were made to minimize animal suffering. The skull was cut and
opened by cutting along the midline with scissors. Following quick
removal from the skull, hemispheres were separated and hippocampi
were dissected on ice. Then, the hippocampi were cut into pieces
and digested with trypsin (0.25%; Gibco; Thermo Fisher Scientific,
Inc.) at 37°C for 15 min. The digested tissues were filtered and
centrifuged at 1,500 × g for 5 min at 4°C, and the remaining cell
pellet was resuspended with medium containing 92% Neurobasal
medium, 5% FBS (cat. no. 16140063; Gibco; Thermo Fisher Scientific,
Inc.), 2% B27, 1% Glu and 2 µl gentamicin. Cells were seeded at a
density of 1–5×105/ml in 6-well plates for 24 h,
arabinosylcytosin (10 mg/l) was added after 72 h at 37°C to prevent
the growth of non-neuronal cells and the medium was subsequently
half changed every 3 days.
Glu-mediated excitotoxicity model and
Pic treatment groups
After 7 days of culture, primary hippocampal neurons
were washed with Mg2+-free extracellular solution
followed by exposure to 100 mM Glu (Sigma-Aldrich; Merck KGaA) for
5 min at room temperature in HEPES buffer containing 140 mM NaCl,
3.0 mM KCl, 2.0 mM CaCl2, 1.0 mM
NaH2PO4, 4.2 mM NaHCO3, 10.0 mM
glucose and 10.0 mM HEPES. Control group neurons were immersed in
HEPES buffer for the same period. After washing, cultures were
returned to the previous medium. Pic was dissolved in DMSO, such
that the final DMSO concentration was <0.1%. The experimental
groups were: Control, Glu, Pic + Glu and Pic + Glu +
Sirt1-small-interfering (si)RNA. Neurons transfected with control
vector, were considered as the negative control group. An
equivalent volume of DMSO was added at the same time point in the
Control and Glu groups. A Cell Counting Kit-8 (CCK-8) assay
(MedChem Express) was used to observe the optimal concentration and
treatment period of Pic.
Cell viability assay
CCK-8 assays were used to investigate the optimal
drug concentration of Pic for subsequent experiments, based on the
cell viability of primary hippocampal neurons. Cell viability test
was performed using a CCK-8 Kit according to the manufacturer's
instructions (cat. no. CK04; Dojindo Molecular Technologies, Inc.).
Briefly, following Glu treatment, primary hippocampal neurons were
plated at a density of 1×104−105 cells/well
in a 96-well plate in triplicate. Neurons were cultured in a
CO2 incubator at 37°C for 24 h, and treated with
different concentrations of Pic (0.01, 0.1, 1, 10 or 100 mM) at
37°C for 12, 24 or 48 h. Subsequently, 10 µl CCK-8 solution was
added to each well for 2 h. Cell viability was analyzed using
spectrophotometry at 450 nm in a microplate reader.
Transfection of siRNA
Neurons were grown in 6-well plates for 24 h and
then transfected with Sirt1-siRNA (50 nM) for 12 h using
Lipofectamine 2000™ (Invitrogen; Thermo Fisher Scientific, Inc.).
Cells were transfected for 48 h, then used for subsequent
experiments. Sirt1-specific siRNA and negative control (NC) siRNA
were synthesized by Shanghai GenePharma Co., Ltd. Sirt1-siRNA
sequences synthesized for rat neurons were as follows: Sense,
5′-GCGAUGUUAUAAUUAAUGAtt-3′ and antisense,
5′-UCAUUAAUUAUAACAUCGCag-3′. Neurons treated with NC siRNA were
used as the NC group. The NC forward sequence used was
5′-UUCUCCGAACGUGUCACGUTT-3′, and reverse sequence was
5′-ACGUGACACGUUCGGAGAATT-3′. The transfection effect of siRNA was
confirmed western blot. The molecular mass of Sirt1 was determined
by running purified protein on 12% SDS-PAGE with standard molecular
weight protein marker (Bio-Rad).
Statistical analysis
Data are presented as the mean ± SD, and were
analyzed using SPSS version 23.0 (IBM, Corp.). Each experiment was
performed a minimum of three times. Results of the rotarod
performance test, mNSS and oriented navigation trials in MWM test
were analyzed using a two-way mixed model ANOVA with Sidak's post
hoc test. All other data were analyzed using a one-way ANOVA with
Tukey's post hoc test for multiple comparisons. P<0.05
(two-sided) was considered to indicate a statistically significant
difference.
Results
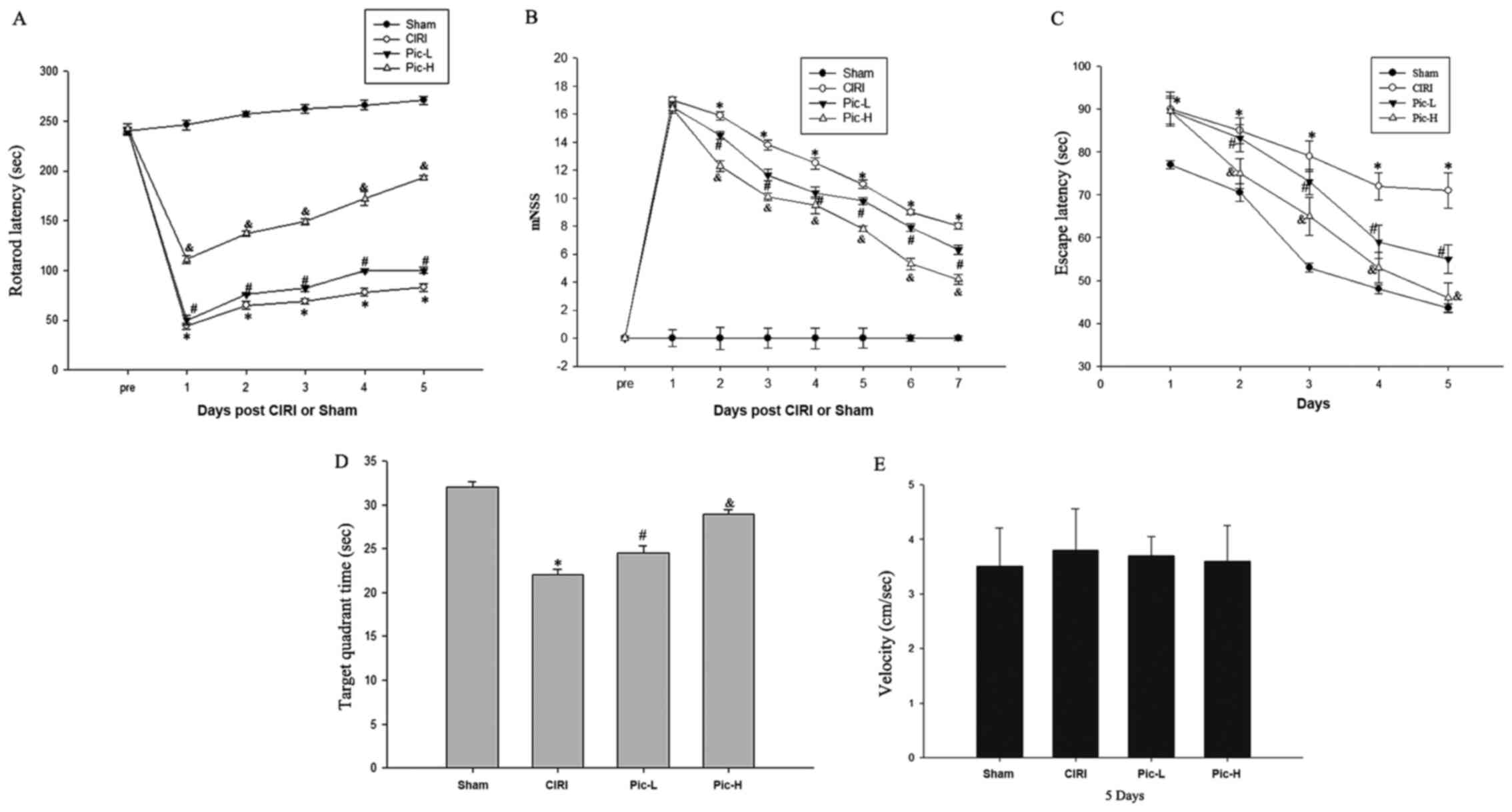
Pic administration ameliorates
CIRI-induced neurological deficits
The rotarod performance test, MWM test and mNSS
provide a more synthetic evaluation of neurological functions
following CIRI (31). The rotarod
latencies of the CIRI mice were significantly shorter compared with
that of the mice in the Sham group (P<0.05; Fig. 1A), whereas a L-dose of Pic
significantly increased rotarod latencies compared with the CIRI
group (P<0.05). There was a significant difference in rotarod
latencies between the Pic-L group and Pic-H group (P<0.05). mNSS
were judged 1 week after CIRI. Mice with CIRI demonstrated impaired
motor function compared with the Sham group (P<0.05; Fig. 1B). By comparison with the CIRI
group, Pic treatment significantly decreased the mNSS and mice
exhibited improved neural function in a dose-dependent manner.
Compared with the Sham group, CIRI led to notable learning and
memory deficits (Fig. 1C);
however, L-dose Pic treatment shortened the escape latency after
the onset of CIRI (P<0.05). Mice in the Pic-H group exhibited a
significantly improved performance in the positioning voyage test
(P<0.05).
After the 4-day training period, the platform was
moved away. As illustrated in Fig.
1D, a significant decline in the target quadrant time was
observed in the CIRI group compared with the Sham group, while an
increase in target quadrant time was observed in the L-dose Pic
treatment groups compared with the CIRI group (P<0.05). The
performance of mice in the L-dose Pic treatment was improved,
compared with the low-dose group (P<0.05). However, there were
no significant differences in swim speed during the space
exploration trials (Fig. 1E).
Pic attenuates pathological injury
caused by CIRI
Structural changes in hippocampal neurons was
observed via H&E staining. Hippocampal tissues in the Sham
group demonstrated normal histology with palely stained neuronal
cells and round nuclei (Fig. 2A).
Compared with the Sham group, various histopathological changes
were observed in the CIRI group, such as nuclear pyknosis, a
decrease in neuronal numbers and neuron atrophy (Fig. 2B). When mice were treated with a
L-dose of Pic, the number of hippocampal neurons increased and
neuronal atrophy was alleviated, although it did not reach the
level of the Sham group (Fig. 2C).
Moreover, hippocampal tissues in the Pic-H treated group had less
pathological impairment (Fig.
2D).
Pic reverses the increase of ROS
levels and decreases the levels of antioxidant molecules
CIRI resulted in significantly increased ROS
generation (P<0.01). By contrast, both L- and H dose Pic
treatment attenuated CIRI-induced ROS increases (Fig. 3A). Moreover, there was a
significant decline in the activity of SOD (P<0.01; Fig. 3B), CAT (P<0.01; Fig. 3C) and GSH-Px (P<0.05; Fig. 3D) in the CIRI group compared with
the Sham group. Of note, there was a rapid increase in cellular
antioxidant enzyme activity in the Pic-L and Pic-H treatment group
in a dose-dependent manner (P<0.05).
 | Figure 3.Pic reverses the increase of ROS and
decrease in antioxidant molecules. (A) ROS production was evaluated
as a marker of oxidative stress. Activities of (B) SOD, (C) CAT and
(D) GSH-Px were detected to measure antioxidant enzymes system. (E)
Content of decreased GSH was determined to evaluate non-enzymatic
antioxidants system. Data are presented as the mean ± SD (n=5 per
group). *P<0.05, **P<0.01 vs. Sham; #P<0.05,
##P<0.01 vs. CIRI; &P<0.05 vs.
Pic-L. CIRI, cerebral ischemia-reperfusion injury; Pic,
piceatannol; ROS, reactive oxygen species; SOD, superoxide
dismutase; CAT-Catalase; GSH-Px, glutathione peroxidase; GSH,
glutathione; L, low; H, high. |
As for the non-enzymatic antioxidants system,
although Pic-L significantly increased GSH content compared with
the CIRI group (P<0.05), no statistically significant difference
was observed between the Pic-L and Pic-H treatment group
(P>0.05; Fig. 3E).
Pic attenuates abnormalities in
apoptosis-related markers caused by CIRI
In order to assess neuronal apoptosis in the
hippocampus, the expression levels of CC-3, Bax and Bcl-2 were
examined using western blotting. It was identified that there was a
significant increase in the expression of CC-3 (P<0.01; Fig. 4A) and in the Bax/Bcl-2 ratio
(P<0.01; Fig. 4B) in the CIRI
group. A dose-dependent decrease in CC-3 expression and Bax/Bcl-2
ratio was observed following treatment with Pic (both doses
P<0.05). The CC-3 mRNA expression level and Bax/Bcl-2 mRNA ratio
also demonstrated the same trend.
Pic attenuates the deficit in the
Sirt1/FoxO1 pathway
Significantly lower gene and protein expression
levels of Sirt1 and FoxO1 were observed in the CIRI group
(P<0.01). Pic treatment significantly reversed this decrease in
Sirt1 and FoxO1 expression levels caused by CIRI. Moreover, the
higher dose of Pic upregulated the Sirt1/FoxO1 signaling pathway to
a greater degree compared with the Pic-L treatment group (all
P<0.05; Fig. 4D and E). The
mRNA expression levels of Sirt1 and FoxO1 also demonstrated the
same trend as the protein expression changes (Fig. 4F).
Pic decreases CIRI-induced apoptosis
via activation of the Sirt1/FoxO1 pathway
The results of the inhibitory effect of Ex-527 on
the Sirt1/FoxO1 pathway are presented in Fig. 5A. CIRI reduced the expression
Sirt1/FoxO1 pathway-related proteins compared with Sham group. Upon
addition of Pic, the Sirt1/FoxO1 pathway is activated. By contrast,
Ex-527 counteracted the activation of Sirt1/FoxO1 pathway by Pic.
The presence of TUNEL-positive cells was significantly increased in
the CIRI group compared with the Sham group (Fig. 5B and C). Compared with CIRI group,
the number of apoptotic hippocampal neurons was significantly
decreased in the CIRI + Pic group (P<0.01; Fig. 5C). Furthermore, the proportion of
apoptotic neurons was significantly increased by addition of Ex-527
(P<0.05), compared with the CIRI + Pic group.
 | Figure 5.Representative images co-stained of
DAPI (blue) and TUNEL (green) demonstrate the effect of Pic on
neuron apoptosis after Sirt1/FoxO1 pathway inhibition. (A) Protein
expression levels of Sirt1 and FoxO1 were inhibited with the
addition of Ex-527. (B) TUNEL-positive cells were barely detected
in the Sham group, but were widely distributed in CIRI group
(magnification, ×400). Pic (20 mg/kg/day) decreased the number of
apoptosis cells, which was blocked by Ex-527. Scale bar, 25 µm. (C)
Compared with the Sham group, apoptosis cells were significantly
increased in CIRI group. Pic decreased the number of TUNEL-positive
cells, whereas Ex-527 combined with Pic reversed this declining
trend. Data are presented as the mean ± SD (n=5 per group).
**P<0.01 vs. Sham; ##P<0.01 vs. CIRI;
&P<0.05, &&P<0.01 vs. CIRI
+ Pic. CIRI, cerebral ischemia-reperfusion injury; Pic,
piceatannol; Sirt1, sirtuin1; L, low; H, high. |
Double immunofluorescent staining of CC-3 and NeuN
indicated that Pic treatment not only decreased the protein
expression of CC-3, but also significantly increased neuronal
survival of the CIRI mice. However, after administration of Ex-527,
apoptosis of hippocampal neurons increased compared with the CIRI +
Pic group (P<0.05; Fig. 6).
Pic alleviates oxidative damage by
modulating the Sirt1/FoxO1 pathway
After inhibition of the Sirt1/FoxO1 pathway, the
generation of ROS significantly increased compared with that in the
CIRI + Pic group (P<0.05; Fig.
7A). Conversely, there was a significant decline in the SOD,
CAT and GSH-Px activities following treatment with Ex-527
(P<0.05; Fig. 7B-D), suggesting
that Pic upregulated cellular antioxidant enzymes activity via the
Sirt1/FoxO1 pathway.
 | Figure 7.Inhibition of the Sirt1/FoxO1 pathway
reverses the decrease of ROS and increase in antioxidant molecules.
(A) Pic treatment caused a decrease of ROS production in the
neurons, whereas inhibition of the Sirt1/FoxO1 pathway could
significantly reverse Pic treatment-induced ROS reduction.
Antioxidant enzymes systems, (B) SOD, (C) CAT and (D) GSH-Px, and
the non-enzymatic antioxidants system, (E) reduced GSH, were also
declined after inhibition of the Sirt1/FoxO1 pathway. Data are
presented as the mean ± SD (n=5 per group). *P<0.05, **P<0.01
vs. Sham; #P<0.05, ##P<0.01 vs. CIRI;
&P<0.05, &&P<0.01 vs. CIRI
+ Pic. CIRI, cerebral ischemia-reperfusion injury; Pic,
piceatannol; ROS, reactive oxygen species; SOD, superoxide
dismutase; CAT, catalase; GSH, Px-Glutathione peroxidase; GSH,
glutathione; Sirt1, Sirtuin1. |
With regards to the non-enzymatic antioxidant
system, the increased GSH content in the CIRI + Pic group was
reversed by inhibition of the Sirt1/FoxO1 pathway using Ex-527
(P<0.05; Fig. 7E).
Inhibiting the activity of the
Sirt1/FoxO1 pathway reverses the effects of Pic on the apoptotic
rate and antioxidant enzymes in primary hippocampal neurons
In the in vitro experiments, in order to
evaluate the functional effects of Pic on neurons treated with or
without Glu, the effect of different doses of Pic (0.01, 0.1, 1, 10
or 100 mM) on cell viability was investigated. Glu significantly
inhibited cell viability at all time points and all concentrations
of Pic. Compared with the control group, 0.01, 0.1, 1 and 10 mM PIC
significantly promoted neuronal viability, irrespective of Glu
treatment (P<0.05; Fig. 8A-B).
However, 100 mM Pic significantly reduced cell viability compared
with cells treated with 10 mM Pic (P<0.05; Fig. 8A and B). Thus, 10 mM Pic was used
for subsequent experiments. Within 24 h, cell viability was
increased in a time-dependent manner, but decreased after 24 h
(P<0.05; Fig. 8C).
 | Figure 8.Effect of Pic on the apoptotic rate
of the primary hippocampal neurons after Sirt1-siRNA treatment.
First, cells were treated with different concentrations (0.01, 0.1,
1, 10 and 100 mM) of Pic under Glu free or Glu treated conditions.
Cell viability treated (A) with and (B) without Glu at 12, 24 and
48 h with different Pic concentrations. *P<0.05 vs. control
group at 12 h; &P<0.05 vs. control group at 24 h;
#P<0.05 vs. control group at 48 h;
@P<0.05 vs. Pic at 10 mM at 12 h;
aP<0.05 vs. Pic at 10 mM at 24 h;
bP<0.05 vs. Pic at 10 mM at 48 h. (C) Cell viability
under the Pic concentration of 10 mM with scheduled duration of
trial treatment. *P<0.05 vs. Glu free condition at 24 h;
&P<0.05 vs. Glu-treated condition at 24 h. (D)
Western blotting results demonstrating (E) the silencing effect of
Sirt1-siRNA. *P<0.05 vs. si-NC. (F) TUNEL-stained green cells
indicated apoptosis-positive cells, DAPI-stained blue as nucleated
cells and Merge stained with TUNEL and DAPI. Magnification, ×200.
(G) Quantification of TUNEL positive cell ratio in different
groups. (H) Inhibiting the activity of the Sirt1/FoxO1 pathway can
reverse the effect of Pic on the activities of SOD and CAT. Data
are presented as the mean ± SD (n=5 per group). *P<0.05,
**P<0.01 vs. Control; #P<0.05,
##P<0.01 vs. Glu; &P<0.05,
&&P<0.01 vs. Pic + Glu. Glu, glutamate; CIRI,
cerebral ischemia-reperfusion injury; Pic, piceatannol; SOD,
superoxide dismutase; CAT, catalase; NC, negative control; siRNA,
small interfering RNA; Sirt1, Sirtuin1. |
The silencing effect of Sirt1-siRNA is presented in
Fig. 8D and E. The rate of
apoptosis was defined as the number of TUNEL-positive cells divided
by the number of DAPI stained cells. The results demonstrated that
knockdown of Sirt1 significantly weakened the protective effects of
Pic on hippocampal neurons from Glu-induced apoptosis (Fig. 8F and G). Compared with the Glu
treated group, the activities of SOD and CAT were increased in the
Pic + Glu group, and the expression of the antioxidant enzymes were
decreased when Sirt1 expression was knocked down (P<0.05;
Fig. 8H).
Discussion
Ischemic stroke is currently a major health concern
that is considered a worldwide health issue. A large-scale survey
in China in 2013 reported that the prevalence of stroke was 1.114%,
and cerebral ischemic stroke accounted for ~70% (2). Cerebral artery occlusion may cause a
sudden interruption of cerebral artery blood supply, leading to
cerebral anoxia and ROS accumulation (6). Upon restoration of blood flow,
cerebral oxidative stress results in further aggravation, leading
to an imbalance between antioxidant defense mechanisms and oxidant
production, thus causing a disturbance of cell-survival mechanisms
and ultimately neurological damage (32,33).
In the present study, the effects of Pic against oxidative stress
and neuronal apoptosis in a murine mouse model of CIRI were
assessed, as well as the possible mechanisms and the involved
signaling pathways.
Pic belongs to the natural polyphenolic compound
family, and has been confirmed to be a naturally occurring
hydroxylated analogue of resveratrol (34). Due to the similarity in the
molecular structure with resveratrol, Pic was hypothesized to exert
similar pharmacological properties. Previous studies reported that
Pic possesses a multitude of biological properties, such as
anti-cancer, antioxidative and hepato- and neuroprotective effects
(35–37). It is now well established that cell
viability increases in a dose-dependent manner when cells are
treated with Pic following injury (38). Furthermore, Pic can exert its
anti-oxidant effects via the upregulation of anti-oxidant enzymes
such as SOD and CAT (36,39). Pic is rapidly metabolized by the
liver and is converted predominantly into a glucuronide conjugate
(40). Thus, it was speculated
that increasing the concentration of Pic appropriately would
increase its antioxidant properties, and this was previously
confirmed by Yokozawa and Kim (41).
The present study demonstrated that mice with CIRI
exhibited an increase in ROS production, accompanied by a decrease
in the expression of antioxidative enzymes. Pic treatment (10
mg/kg/day) decreased the generation of ROS, and this decline was
significantly higher in the Pic-H treatment group. Moreover, SOD,
CAT and GSH-Px activities, which are used as indicators of
antioxidant capability, was determined in the present study. The
activity of SOD, CAT and GSH-Px was markedly upregulated in both
low dose and high dose Pic-treated mice; animal experiments
performed by Wen et al (36) and Zhang et al (37) also presented the same results.
However, it was identified that Pic upregulated GSH content in the
CIRI mice, but this effect was not dose-dependent. Other study also
reported the effects of Pic on non-enzymatic enzyme system, but did
not determine whether these effects were dose-dependent (41). Therefore, further study is required
to determine the potential molecular mechanisms to validate the
current findings.
In addition to alleviating oxidative stress, Pic is
reported to exert anti-apoptotic properties by downregulating CC-3,
as well as maintaining the balance between the apoptosis-related
genes Bax and Bcl-2 (20). The
present results suggested that low dose and high dose of Pic
decreased the expression of CC-3 and Bax/Bcl-2 at the
transcriptional level, but it did not reach the levels found in the
sham group. Despite limited neuroprotection, Pic was able to exert
a small but beneficial effect on neural survival. It has also been
shown that caspases, such as C-9 and C-3, and poly (ADP-ribose)
polymerase are activated by Pic in a dose-dependent manner
(20,42). The present results were consistent
with previous study confirming the reliability of the effects of
Pic (42). However, due to the
different cell types and intervention methods used, there were some
inconsistencies with prior studies. For instance, Song et al
(43) revealed that Pic treatment
was able to increase tumor apoptosis. In a model of transplanted
mammary cancer, Pic treatment was able to increase the expression
levels of Bax and CC-3, but downregulate Bcl-2 expression in tumor
tissues (43). Increased numbers
of tumor cells undergo apoptosis following Pic treatment,
suggesting that Pic inhibits the breast tumor growth via the
induction of apoptosis (43).
Studies determining and targeting the potential mechanisms
regulated by Pic with regards to cell survival are required to
examine the differences between the present and previous
studies.
As for exact mechanism via which Pic exhibits a
positive role, numerous studies have shown that Pic activates
specific signaling pathways to exhibit antioxidant activities and
anti-apoptotic effects, such as the PI3K/Akt and nuclear factor,
erythroid 2 like 2/heme oxygenase 1 signaling pathways (13,44).
Due to the similarity in the structure to resveratrol, Pic was
speculated to exhibit an active role via similar mechanisms. A
previous study revealed that Pic and resveratrol both improved
ischemia and anoxia via upregulating HIF-1α expression, and
downregulating c-Myc, egl-9 family hypoxia inducible factor 1 and
β-catenin expression levels via the activation of Sirt1 (45). The NAD+-dependent
protein deacetylase Sirt1 serves a critical role in the inhibition
of oxidative stress and suppression of apoptosis along with
downstream signaling of the FoxO1 pathway (46). In the present study, Pic enhanced
antioxidant activity and suppressed apoptosis in a dose-response
manner, and the activity of the Sirt1/FoxO1 signaling pathway was
upregulated simultaneously. When cells were treated with Ex-527 or
Sirt1-siRNA, an increase in the number of apoptotic cells and
increased oxidative stress was observed. These results suggested
that the Sirt1/FoxO1 signaling pathway may be associated with the
antioxidant potency and anti-apoptotic abilities of Pic on
hippocampal neurons.
In addition to the aforementioned cytoprotective
effects, the administration of Pic exerts several other effects on
cellular biological processes, such as anti-inflammatory effects
(19), metabolic improvement
(47) and astrocyte
differentiation (48). Several
hypotheses have been suggested as underlying the effects of Pic on
CIRI, such as inhibition of the activation of inflammatory pathway
and innate immune system (49,50).
The presence of numerous hypotheses may be explained by the fact
that the neuroprotective effects of Pic may be the result of
multiple interactions. Therefore, additional studies using further
comprehensive approaches are required to evaluate the underlying
mechanisms that maintained neuronal homeostasis. Moreover,
investigating other natural products with similar chemical
structures may provide a more definitive conclusion. Whether Pic
can also reverse anticoagulant, such as tissue plasminogen
activator, induced toxicity at the subacute stage of ischemic
stroke warrants further investigation. These findings would have an
important clinical impact on the neuroprotection of patients
undergoing intravenous thrombolysis with anticoagulants.
In conclusion, Pic treatment was capable of exerting
neuroprotective effects that were largely due to the maintenance of
the balance of the antioxidative/oxidative system, as well as the
protection of hippocampal neurons from apoptosis. Furthermore, the
Sirt1/FoxO1 signaling pathway was identified to be involved in both
of these processes. The present study provides additional evidence
that highlights the potential use of Pic as a clinically relevant
therapeutic for ischemic stroke.
Acknowledgements
Not applicable.
Funding
The present study was supported by a Project of Key
Medical Science Research Program of Hebei Province (grant no.
20171355).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KJW, WQZ and JZC conceived and designed the study.
JJL and YC were responsible for assessment and accreditation of
laboratory animal care. KJW, WQZ and YC were responsible for data
analysis. JJL and JZC performed histological and behavioral
experiments. KJW, WQZ and JZC wrote the manuscript. JZC reviewed
and edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All research was conducted with the approval of the
Ethics Committee of Tangshan Gongren Hospital (clearance no.
GRYY-LL-2019-13).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zerna C, Thomalla G, Campbell BCV, Rha JH
and Hill MD: Current practice and future directions in the
diagnosis and acute treatment of ischaemic stroke. Lancet.
392:1247–1256. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang W, Jiang B, Sun H, Ru X, Sun D, Wang
L, Wang L, Jiang Y, Li Y, Wang Y, et al: Prevalence, incidence, and
mortality of stroke in China clinical perspective: Results from a
nationwide population-based survey of 480687 adults. Circulation.
135:759–771. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
China's Health and Family Planning
Statistical Yearbook. National Health and Family Planning
Commission of the People's Republic of China. Peking: Peking Union
Medical College Press; 2016
|
|
4
|
Jean WC, Spellman SR, Nussbaum ES and Low
WC: Reperfusion injury after focal cerebral ischemia: The role
inflammation and the the rapeutic horizon. Neurosurgery.
43:1382–1396. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu M, Yiang GT, Liao WT, Tsai AP, Cheng
YL, Cheng PW, Li CY and Li CJ: Current mechanistic concepts in
ischemia and reperfusion injury. Cell Physiol Biochem.
46:1650–1667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang Q, Huang Q, Hu Z and Tang X:
Potential neuroprotective treatment of stroke: Targeting
excitotoxicity, oxidative stress, and inflammation. Front Neurosci.
13:10362019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakka VP, Gusain A, Mehta SL and Raghubir
R: Molecular mechanisms of apoptosis in cerebral ischemia: Multiple
neuroprotective opportunities. Mol Neurobiol. 37:7–38. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Candelario-Jalil E: Injury and repair
mechanisms in ischemic stroke: Considerations for the development
of novel neurotherapeutics. Curr Opin Investig Drugs. 10:644–654.
2009.PubMed/NCBI
|
|
9
|
Huang HL, Fang LW, Lu SP, Chou CK, Luh TY
and Lai MZ: DNA-damaging reagents induce apoptosis through reactive
oxygen species-dependent Fas aggregation. Oncogene. 22:8168–8177.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin TC, Chen YR, Kensicki E, Li AYJ, Kong
M, Li Y, Mohney RP, Shen HM, Stiles B, Mizushima N, et al:
Autophagy: Resetting glutamine-dependent metabolism and oxygen
consumption. Autophagy. 8:1477–1493. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hao Y, Liu J, Wang Z, Yu LL and Wang J:
Piceatannol protects human retinal pigment epithelial cells against
hydrogen peroxide induced oxidative stress and apoptosis through
modulating PI3K/Akt signaling pathway. Nutrients. 11:15152019.
View Article : Google Scholar
|
|
12
|
Boccellino M, Donniacuo M, Bruno F,
Rinaldi B, Quagliuolo L, Ambruosi M, Pace S, De Rosa M, Olgaç A,
Banoglu E, et al: Protective effect of piceatannol and bioactive
stilbene derivatives against hypoxia-induced toxicity in H9c2
cardiomyocytes and structural elucidation as 5-LOX inhibitors. Eur
J Med Chem. 180:637–647. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H, Shi Y, Wang X, Li P, Zhang S, Wu T,
Yan Y, Zhan Y, Ren Y, Rong X, et al: Piceatannol alleviates
inflammation and oxidative stress via modulation of the Nrf2/HO-1
and NF-κB pathways in diabetic cardiomyopathy. Chem Biol Interact.
310:1087542019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang W, Yang R, Yao H, Wu Y, Pan W and Jia
AQ: Inhibiting the formation of advanced glycation end-products by
three stilbenes and the identification of their adducts. Food Chem.
295:10–15. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Moon I, Damodar K, Kim JK, Ryoo S and Jun
JG: Synthesis, anti-inflammatory, and arginase inhibitory activity
of piceatannol and its analogs. Bull Korean Chem Soc. 38:342–349.
2017. View Article : Google Scholar
|
|
16
|
Liu J, Zhou J, Wu Z, Wang X, Liu L and Yao
C: Cyanidin 3-O-β-glucoside ameliorates ethanol-induced acute liver
injury by attenuating oxidative stress and apoptosis: The role of
SIRT 1/FOXO 1 signaling. Alcohol Clin Exp Res. 40:457–466. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yan X, Yu A, Zheng H, Wang S, He Y and
Wang L: Calycosin-7-O-β-D-glucoside attenuates OGD/R-induced damage
by preventing oxidative stress and neuronal apoptosis via the
SIRT1/FOXO1/PGC-1α pathway in HT22 cells. Neural Plast.
2019:87980692019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Llarena M, Andrade F, Hasnaoui M, Portillo
MP, Pérez-Matute P, Arbones-Mainar JM, Hijona E, Villanueva-Millán
MJ, Aguirre L, Carpéné C and Aldámiz-Echevarría L: Potential
renoprotective effects of piceatannol in ameliorating the
early-stage nephropathy associated with obesity in obese Zucker
rats. J Physiol Biochem. 72:555–566. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee HJ, Kang MG, Cha HY, Kim YM, Lim Y and
Yang SJ: Effects of piceatannol and resveratrol on sirtuins and
hepatic inflammation in high-fat diet-fed mice. J Med Food.
22:833–840. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fu Z, Yang J, Wei Y and Li J: Effects of
piceatannol and pterostilbene against β-amyloid-induced apoptosis
on the PI3K/Akt/Bad signaling pathway in PC12 cells. Food Funct.
7:1014–1023. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wong CHY, Jenne CN, Lee WY, Léger C and
Kubes P: Functional innervation of hepatic iNKT cells is
immunosuppressive following stroke. Science. 334:101–105. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Paxinos G and Watson C: The Rat Brain in
Stereotaxic Coordinates (Deluxe Edition). (4th). Rat Brain in
Stereotaxic Coordinates. p2561998.
|
|
24
|
Xiong Y, Qu C, Mahmood A, Liu Z, Ning R,
Li Y, Kaplan DL, Schallert T and Chopp M: Delayed transplantation
of human marrow stromal cell-seeded scaffolds increases
transcallosal neural fiber length, angiogenesis, and hippocampal
neuronal survival and improves functional outcome after traumatic
brain injury in rats. Brain Res. 1263:183–191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aebi H: Catalase in vitro. Methods
Enzymol. 105:121–126. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hafeman DG, Sunde RA and Hoekstra WG:
Effect of dietary selenium on erythrocyte and liver glutathione
peroxidase in the rat. J Nutr. 104:580–587. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hissin PJ and Hilf R: A fluorometric
method for determination of oxidized and reduced glutathione in
tissues. Anal Biochem. 74:214–226. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang W, Cui Y, Gao J, Li R, Jiang X, Tian
Y, Wang K and Cui J: Recombinant osteopontin improves neurological
functional recovery and protects against apoptosis via
PI3K/Akt/GSK-3β pathway following intracerebral hemorrhage. Med Sci
Monit. 24:1588–1596. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kim HJ, Martemyanov KA and Thayer SA:
Human immunodeficiency virus protein Tat induces synapse loss via a
reversible process that is distinct from cell death. J Neurosci.
28:12604–12613. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhi G, Hai-Ping Z, Yu-Min L and Xun-Ming
J: The behavioral testing in mice after cerebral ischemia. Chin J
Comp Med. 22:68–72. 2012.
|
|
32
|
Onwuekwe IO and Ezeala-Adikaibe B:
Ischemic stroke and neuroprotection. Ann Med Health Sci Res.
2:186–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Rodrigo R, Fernández-Gajardo R, Gutiérrez
R, Matamala JM, Carrasco R, Miranda-Merchak A and Feuerhake W:
Oxidative stress and pathophysiology of ischemic stroke: Novel
therapeutic opportunities. CNS Neurol Disord Drug Targets.
12:698–714. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Matsui Y, Sugiyama K, Kamei M, Takahashi
T, Suzuki T, Katagata Y and Ito T: Extract of passion fruit
(Passiflora edulis) seed containing high amounts of
piceatannol inhibits melanogenesis and promotes collagen synthesis.
J Agric Food Chem. 58:11112–11118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Banik K, Ranaware AM, Harsha C, Nitesh T,
Girisa S, Deshpande V, Fan L, Nalawade SP, Sethi G and Kunnumakkara
AB: Piceatannol: A natural stilbene for the prevention and
treatment of cancer. Pharmacol Res. 153:1046352020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wen J, Lin H, Zhao M, Tao L, Yang Y, Xu X,
Jia A, Zhang J and Weng D: Piceatannol attenuates
D-GalN/LPS-induced hepatoxicity in mice: Involvement of ER stress,
inflammation and oxidative stress. Int Immunopharmacol. 64:131–139.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Zhang LH, Chen X, Zhang N and Li
G: Piceatannol attenuates behavioral disorder and neurological
deficits in aging mice via activating the Nrf2 pathway. Food Funct.
9:371–378. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kinoshita Y, Kawakami S, Yanae K, Sano S,
Uchida H, Inagaki H and Ito T: Effect of long-term piceatannol
treatment on eNOS levels in cultured endothelial cells. Biochem
Biophys Res Commun. 430:1164–1168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kiliç V: Piceatannol mediated modulation
of oxidative stress and regeneration in the liver of endotoxemic
mice. J Med Food. 22:594–601. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Piotrowska H, Kucinska M and Murias M:
Biological activity of piceatannol: Leaving the shadow of
resveratrol. Mutat Res. 750:60–82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yokozawa T and Kim YJ: Piceatannol
inhibits melanogenesis by its antioxidative actions. Biol Pharm
Bull. 30:2007–2011. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim HJ, Lee KW and Lee HJ: Protective
effects of piceatannol against beta-amyloid-induced neuronal cell
death. Ann N Y Acad Sci. 1095:473–482. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Song H, Jung JI, Cho HJ, Her S, Kwon SH,
Yu R, Kang YH, Lee KW and Park JHY: Inhibition of tumor progression
by oral piceatannol in mouse 4T1 mammary cancer is associated with
decreased angiogenesis and macrophage infiltration. J Nutr Biochem.
26:1368–1378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wen H, Fu Z, Wei Y, Zhang X, Ma L, Gu L
and Li J: Antioxidant activity and neuroprotective activity of
stilbenoids in rat primary cortex neurons via the PI3K/Akt
signalling pathway. Molecules. 23:23282018. View Article : Google Scholar
|
|
45
|
Hong KS, Park JI, Kim MJ, Kim HB, Lee JW,
Dao TT, Oh WK, Kang CD and Kim SH: Involvement of SIRT1 in hypoxic
down-regulation of c-Myc and β-catenin and hypoxic preconditioning
effect of polyphenols. Toxicol Appl Pharmacol. 259:210–218. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jiang Y, Luo W, Wang B, Wang X, Gong P and
Xiong Y: Resveratrol promotes osteogenesis via activating
SIRT1/FoxO1 pathway in osteoporosis mice. Life Sci. 246:1174222020.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kitada M, Ogura Y, Maruki-Uchida H, Sai M,
Suzuki T, Kanasaki K, Hara Y, Seto H, Kuroshima Y, Monno I and Koya
D: The effect of piceatannol from passion fruit (Passiflora
edulis) seeds on metabolic health in humans. Nutrients.
9:11422017. View Article : Google Scholar
|
|
48
|
Arai D, Kataoka R, Otsuka S, Kawamura M,
Maruki-Uchida H, Sai M, Ito T and Nakao Y: Piceatannol is superior
to resveratrol in promoting neural stem cell differentiation into
astrocytes. Food Funct. 7:4432–4441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Suzuki Y, Nakano Y, Mishiro K, Takagi T,
Tsuruma K, Nakamura M, Yoshimura S, Shimazawa M and Hara H:
Involvement of Mincle and Syk in the changes to innate immunity
after ischemic stroke. Sci Rep. 3:31772013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ye XC, Hao Q, Ma WJ, Zhao QC, Wang WW, Yin
HH, Zhang T, Wang M, Zan K, Yang XX, et al: Dectin-1/Syk signaling
triggers neuroinflammation after ischemic stroke in mice. J
Neuroinflammation. 17:172020. View Article : Google Scholar : PubMed/NCBI
|