Introduction
The identification of risk factors for colon cancer
has been an ongoing investigation in present research (1). With the exception of familial polyposis,
the majority of risk factors have indicated controversial effects
(1). However, ~90% of colon cancer
cases in the US and other developed countries have been reported to
be attributed to environmental factors (1). Among the postulated dietary factors,
vegetable and fruit consumption has been reported as the most
consistent to exert an anti-carcinogenic effect (2,3). It has
been reported that vegetables and fruits have numerous putative
anti-carcinogenic substances, providing biological support to the
hypothesis that vegetable and fruit consumption may aid in the
prevention of the disease (4). Su
Yang Decoction (SYD) comprises two brassicaceous vegetables,
broccoli and green cabbage, which have been reported to exert
anticancer effects in vivo and in vitro (5,6). In
addition, the sulforaphane constituents in vegetables, including
broccoli and green cabbage, have been reported to inhibit the
proliferation of pancreatic cancer (7) and gastric cancer cells (6) and induce cancer cell apoptosis.
Therefore, it has been reported that broccoli and green cabbage are
considered to have anticancer properties and are extensively
consumed in China (2). However, the
expected therapeutic effects of SYD as a compound formula require
further investigation.
Apoptosis is a form of programmed cell death that is
responsible for tissue homeostasis in cancer cells and is induced
by numerous cancer treatments (8). It
has been indicated that apoptosis involves two major pathways: The
intrinsic (mitochondrial-mediated) pathway, which involves the
activation of caspase-9 (CASP9) and caspase-10, and the extrinsic
[death receptor (DR)-mediated] pathway (3). In the extrinsic pathway, the binding of
extracellular death ligands to their cell-surface DRs has been
reported to induce caspase-8 (CASP8) activation (4). In contrast, the intrinsic pathway has
been reported to be activated by the release of proapoptotic
factors, including cytochrome c from the mitochondria to the
cytosol and the activation of CASP9, in addition to being amplified
by the CASP8-mediated cleavage of BH3 interacting domain death
agonist (9). The extrinsic apoptosis
pathway is initiated by the binding of death receptor ligands,
including tumor necrosis factor (TNF)-related apoptosis-inducing
ligand (TRAIL) or cluster of differentiation 95 ligand, to their
cognate death receptors at the cell membrane (10). Active caspase-8 activates caspase-3,
resulting in apoptosis (11). As an
anti-apoptotic protein, cellular FADD-like IL-1β-converting
enzyme-inhibitory protein-inhibitory protein (c-FLIP) can block
death-receptor signaling by interfering with caspase-8 activation
at the DISC (10). Therefore, the
present study aimed to investigate the anticancer activity of SYD
on colon cancer HT-29 cells, in addition to examining the SYD
anticancer underlying mechanism.
Materials and methods
Materials
High performance liquid chromatography (HPLC)-grade
methanol was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt,
Germany). Ultrapure water was prepared using a Millipore SAS 67120
system (Merck KGaA). The sulforaphane (98% purity), as the
reference standard substance, was purchased from Shanghai Yuanye
Biotechnology Co., Ltd., (Shanghai, China). Fetal bovine serum
(FBS), penicillin G, streptomycin and amphotericin B were obtained
from Gibco (Thermo Fisher Scientific, Inc., Waltham, USA). Dimethyl
sulfoxide, ribonuclease (RNase), propidium iodide (PI) and
RPMI-1640 were purchased from Sigma-Aldrich (Merck KGaA). Broccoli
and green cabbage material were obtained from Infinitus Company
Ltd. (Guangzhou, China) and were placed on dry ice and freeze-dried
immediately to preserve their freshness.
HPLC-ultraviolet (UV) analysis
A Shimadzu LC-20AT HPLC system with an UV detector
was used (Shimadzu Corporation, Kyoto, Japan) for quantitative
determination. A Phenomenex Luna C18 column (4.6×250 mm, 5 µm;
Guangzhou FLM Scientific Instrument Co., Ltd., Guangzhou, China)
was used, according to the manufacturer's protocols, and the mobile
phase composed of methanol:water (20:80% v/v) at a flow rate of 1.0
ml/min. Furthermore, the detection wavelength was 225 nm and the
temperature of the column was set to 30°C. The injection volume was
20 ml. The limit of detection was 0.2 µg/ml. Data acquisition was
performed using the LabSolutions CS software version 2.53 (Shimadzu
Corporation). There are 6 points in the Standard curve (6 standard
samples); additionally, a representative standard sample and tested
sample were used. All experiment were repeated twice each one in
duplicate.
Preparation of SYD
SYD was prepared according to the procedure
described by Rose et al (7).
In brief, 100 mg of freeze-dried material, including broccoli and
green cabbage at a weight ratio of 1:1, was weighed into a 50-ml
polypropylene tube and subjected to two 30 min cycles of ultrasonic
disruption (59 kHz) in 70% ethanol (3 ml) at 70°C. The mixture was
cooled to room temperature and centrifuged at 3,000 × g for 5 min.
Following centrifugation, 1-ml aliquots were removed and condensed
in a vacuum to 200 µl. The resulting concentrates were filtered
through sterile non-pyrogenic filters (٠.٢ µm; Merck KGaA) and
stored at 70°C prior to testing. The extracts for each sample
yielded an equivalent concentration of 50 mg/ml. The sulforaphane
composition in the extracts was analyzed with a Shimadzu LC-20AT
HPLC system with an UV detector (Shimadzu Corporation, Kyoto,
Japan).
Cell culture
Following ethical approval by the Institutional
Animal Care Committee and the Local Veterinary Office and Ethics
Committee of Guangzhou Medical University (Guangzhou, China), the
human colon cancer HT-29, LS-174-T and CRL-1790 cell lines were
obtained from the Clinic Research Center of Guangzhou Medical
University (Guangzhou, China). They were used for the subsequent
proliferation, apoptosis, cell cycle and western-blotting assays.
The human colon epithelial CRL-1790 cell line was obtained from
American Type Culture Collection (Manassas, VA, USA). HT-29,
LS-174-T and CRL-1790 cells were grown in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) with
10% FBS and 1% Penicillin G-Streptomycin and maintained at 37°C
with 5% CO2 atmosphere. All cell studies were approved
by the Institutional Animal Care Committee and the Local Veterinary
Office and Ethics Committee of Guangzhou Medical University
(Guangzhou, China).
Proliferation assay
Proliferation was assessed through an MTS assay kit
(Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocols. HT-29, LS-174-T and CRL-1790 cells were
inoculated in 96-well plates at the density of 5×104
cells/well. At 0, 24, 48 and 72 h of incubation at 37°C, 20 µl
MTS/PMS reagent was added for another 4 h of incubation at room
temperature. Dimethyl sulfoxide was used to dissolve the purple
formazan. The absorbance at 490 nm was detected by a microplate
reader. HT-29 cells were treated by 0, 10, 50, 100 or 200 µg/ml SYD
for 48 h at 37°C, and then lactate dehydrogenase (LDH) release was
assessed with a LDH activity kit (cat. no. 03002209122; Roche
Diagnostics, Indianapolis, IN, USA) and expressed as a percentage
[(sample absorbance/lysed cell absorbance-control absorbance)
×100]. Each individual experiment was performed in triplicate.
A total of 5×104−1×105 control
small interfering RNA (siRNA) or c-FLIP siRNA HT-29 cells/well were
seeded in 6-well plates. After 24 h at 37°C, the DMEM with 10% FBS
was replaced with fresh DMEM with 10% FBS and 0, 100 or 200 µg/ml
SYD at 37°C and incubated for 7 days. Cell viability was determined
by Typan Blue staining at room temperature for 30 sec and direct
cell counting using a hematocytometer (Thorlabs, Inc., Newton, NJ,
USA).
Flow cytometry analysis of the cell
cycle distribution
The flow cytometric analysis was performed as
described previously (12).
Subconfluent cultures of HT-29 cells (1.0×105
cells/well) were treated with either PBS or 0, 100, 200 and 400
µg/ml SYD. Subsequent to 24, 48 and 72 h of treatment at room
temperature, the cells were harvested by brief trypsinization and
centrifugation at room temperature at 3,000 × g for 5 min. The cell
pellets were washed twice with ice-cold PBS and 0.5×106
cells were suspended in 500 ml of saponin/PI solution [0.3% (w/v)
saponin; 25 µg/ml PI; 0.1 mmol/L EDTA and 10 µg/ml RNase A in PBS]
and incubated at ٤°C for 24 h in the dark. Apoptosis was quantified
using an annexin V-fluorescein isothiocyanate (FITC) kit (R&D
Systems, Inc., Minneapolis, MN, USA), according to the
manufacturer's protocols. The stained cells were analyzed using a
FACS flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA) and
FCS Express V3.0 software (De Novo Software, Glendale, CA, USA).
ModFit LT software v3.3 (Verity Software House, Inc., Topsham, ME,
USA) was used to analyze the results of the cell cycle.
Colorimetric caspase activation
assay
HT-29 cells (1×106) were seeded into 6 cm
dishes and treated with 0, 100, 200 and 400 µg/ml SYD for the
indicated times, the activities of caspase-3 (CASP3), CASP8 and
CASP9 were measured by cleavage of a 5 µl IETD-ρNA substrate
(caspase-8) or DEVD-ρNA substrate (caspase-3, 9). CAS activity was
measured using the Colorimetric CaspACE assay system, and the
activities of CASP8 (cat. no. AAH-APO-١-٢) and CASP9 (cat. no.
68FL-Casp9-S100) were determined with CASP8 and CASP9 Colorimetric
assay kits, according to the manufacturer's protocols (RayBiotech
Life, Norcross, GA, USA). To inhibit the tested caspases, the cells
were preincubated at 37°C for 2 h with 50 mM Z-DEVD-FMK (cat. no.
A1920; CASP3 inhibitor), Z-IETD-FMK (cat. no. B3232; CASP8
inhibitor), Z-LEHD-FMK (cat. no. B3233; CASP9 inhibitor) and
Z-VAD-FMK (cat. no. A1902; Pan-caspase inhibitor) (all from
ApexBio, Shanghai, China).
Measurement of the release of cytochrome c. The
release of cytochrome c from the mitochondria of HT-29 cells
into the cytosol was measured using a streptavidin-peroxidase
immunohistochemical kit (cat. no. SP0041; OriGene Technologies,
Inc., Beijing, China) as described previously (13). Briefly, following incubation at 37°C
for 24 h, HT-29 cells (1×106) were seeded into 6 cm
dishes and were exposed to either 100 µg/ml or 200 µg/ml SYD or
left untreated (control group). Subsequent to an additional ١٢ h
incubation at 37°C, the cells were sequentially treated with 3%
H2O2, blocking buffer (provided in the kit),
incubated overnight at 37°C with the primary antibody (1:100;
anti-cytochrome c; cat. no. KG22230; Nanjing KeyGen Biotech
Co., Ltd., Nanjing, China). All sections were stained using the
streptavidin-horseradish peroxidase complex method with goat
anti-rabbit antibodies (1:1,000; provided in the kit) as the
secondary antibody at 37°C for 30 min, and then 4°C overnight.
Brown-yellow granules in the cytoplasm represented a positive
staining.
The HT-29 cells incubated with different does of SYD
were dyed with rhodamine 123 for 1 h at room temperature. The
primary antibody (1:100; anti-cytochrome c; cat. no.
KG22230) was incubated for 1h and then incubated with goat
anti-rabbit secondary antibodies with APC fluorescence (cat. no.
A-11034; Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 30
min. Finally, DAPI (cat. no. 40727ES10; Nanjing KeyGen Biotech Co.,
Ltd.) was directly nucleated for 30 min at room temperature. The
release of cytochrome c was observed under a light
microscope (Olympus CX31-LV320 Olympus (China) Co., Ltd., Shanghai,
China) subsequent to color development (×400).
Western blot analysis
Western blot analyses were performed as described
previously (13) using the following
antibodies: Major histocompatibility complex, Class II, DR Beta 4
(DR4; cat. no., 42533S; 1:1,000), major histocompatibility complex,
Class II, DR Beta 5 (DR5; cat. no., 3696S; 1:1,000), Fas cell
surface death receptor (Fas; cat. no., 8023S; 1:1,000), CASP8 (cat.
no., 4927S; 1:1,000), CASP9 (cat. no., 9502S; 1:1,000), CASP3 (cat.
no., 9664S; 1:1,000) poly (ADP-ribose) polymerase (PARP; cat. no.,
9532S; 1:1,000), β-actin (cat. no., 4970S; 1:1,000), TNF receptor
associated factor 2 (TRAF2; cat. no., 4724S; 1:1,000), TRAIL (cat.
no., 3219S; 1:1,000), TNF-α (cat. no., 34; 1:500), X-linked
inhibitor of apoptosis (XIAP; cat. no., 14334S; 1:1,000), BCL2,
apoptosis regulator (Bcl-2; cat. no., 15071; 1:1,000), BCL2 like 1
(Bcl-xl; cat. no., 2762S; 1:500), MCL1, BCL2 family apoptosis
regulator (Mcl-1; cat. no., 94296S; 1:1,000), BCL2 associated X,
apoptosis regulator (Bax; cat. no., 2774S; 1:1,000) and truncated
(t)-BH3 interacting domain death agonist (BID; cat. no., 8762S;
1:500). All aforementioned antibodies were purchased from Cell
Signaling Technology, Inc., (Danvers, MA, USA). In addition,
cellular-FLICE-like inhibitory protein (c-FLIP; cat. no., sc-8346,
1:500) was purchased from Santa Cruz Biotechnology, Inc., (Dallas,
TX Santa Cruz Biotechnology). Total proteins were extracted from
cells using lysis buffer containing phenylmethyl sulfonylfluoride
(Beyotime Institute of Biotechnology, Haimen, China) at 25°C for 30
min and protein concentration was determined with a BCA Protein
Assay kit (Beyotime Institute of Biotechnology). A total of 20 µg
protein was separated by SDS-PAGE (١٠٪ gel) and transferred onto a
polyvinylidene fluoride membrane. Membranes were blocked with 5%
non-fat milk at room temperature for 2 h and incubated with primary
antibodies aforementioned at 4°C for 12 h. Anti-β-actin (cat. no.,
4970S; 1:1,000; Cell Signaling Technology, Inc.) was used as a
loading control. Membranes were then washed and incubated with
horseradish peroxidase-conjugated secondary antibody (cat. nos.
A21020 and A٢١٠١٠; ١:٨,٠٠٠; Abbkine Scientific Co., Ltd., Wuhan,
China) at room temperature for 2 h. Immunoreactive bands were
visualized using a chemiluminescence solution with an enhanced
chemiluminescent kit (Beyotime Institute of Biotechnology).
Quantity One (v4.6.8; Bio-Rad Laboratories, Inc., Hercules, CA,
USA) was used as the software for quantification.
RNA interference
For the transient knockdown of c-FLIP, the HT-29
cells were transfected with 150 pmol Stealth RNAi siRNA directed
against c-FLIP (si-c-FLIP) or non-targeting control siRNA (Scr)
(both from Invitrogen; Thermo Fisher Scientific, Inc.) using the
TransMessenger transfection reagent kit (Qiagen GmbH, Hilden,
Germany). Cells were incubated with the transfection complexes for
4 h under their normal growth conditions. Subsequently, the
complexes were removed from the cells, the cells were washed once
with PBS, and then 500 µl fresh medium containing FBS and
antibiotics (1% penicillin G-streptomycin; cat. no. 15140122;
Thermo Fisher Scientific, Inc.) was added to the cells, according
to the manufacturer's protocols. c-FLIP, sense,
5′-CGGACTATAGAGTGCTGATGG-3′ and antisense
5′-GATTATCAGGCAGATTCCTAG-3′.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
For the measurement of mRNA expression levels in
HT-29 cells with SDY treatment, the mRNA expression levels of DR3,
DR4, DR5, Fas and TNF-α receptor were detected by RT-qPCR following
treatment with 0, 10, 50, 100, 150 or 200 µg/ml SYD for 24 h at
room temperature. The housekeeper gene GAPDH was used as a control.
The RNA of cells was extracted using TRIzol® reagent
(cat. no. 15596018; Invitrogen; Thermo Fisher Scientific, Inc.).
cDNA templates from HT-29 cell lines were prepared using a
TIANScript RT kit (Tiangen Biotech, Co., Ltd., Beijing, China),
according to the manufacture's protocol. RT-qPCR cycling was
performed in 96-well plates on a LightCycler 480 Real-Time PCR
system (Roche Applied Science, Penzberg, Germany). The reaction was
performed in a 20 µl total volume containing ١٠ µl SYBR®
Select Master mix (cat. no. 4472908; Thermo Fisher Scientific,
Inc.), 1 µl each primer (١٠ µM) and ٢ µl template cDNA. The primer
sequences used for PCR are presented in Table I. The amplification protocol consisted
of an initial denaturation step at 95°C for 5 min, followed by
two-step PCR for 40 cycles at 95°C for 30 sec and 60°C for 30 sec.
The mRNA expression levels of each target were determined based on
the cycle threshold (Cq) value for the reference and each target
and calculated as 2-∆Cq (5). Three
independent experiments were performed.
 | Table I.Primer sequences used for PCR. |
Table I.
Primer sequences used for PCR.
| Gene | Forward | Reverse |
|---|
| DR3 |
5′-GAGAAGTCCCTGCACCACGA-3′ |
5′-TGGGTTCTTTGAGGCTGCTG-3′ |
| DR4 |
5′-GCCCCACAACAAAAGAGGTC-3′ |
5′-GGAGGTCATTCCAGTGAGTG-3′ |
| DR5 |
5′-GCTGGGCATCTGGACCCTCCTACCT-3′ |
5′-CAGTCACTTGGGCATTAACACTT-3′ |
| Fas |
5′-CCTGTGAGGAGGACGAAC-3′ |
5′-CCTGTGAGGAGGACGAAC-3′ |
| TNF-αR |
5′-GTCTTCACCACCATGGAG-3′ |
5′-CCACCCTGTTGCTGTAGC-3′ |
| GAPDH |
5′-TGACTTGATCCGACACATGG-3′ |
5′-TCCCATCCATCCAAAAACAT-3′ |
Statistical analysis
Data were analyzed using SPSS software (version
19.0; IBM Corp., Armonk, NY, USA). The data are presented as the
mean ± standard deviation. The data were analyzed using a
two-tailed Student's t-test. For multiple comparisons, a one-way
analysis of variance was used with Scheffe as the post-hoc test.
P<0.05 or P<0.001 was considered to indicate a statistically
significant difference. The calibration was assessed using
Hosmer-Lemeshow goodness-of-fit test and Pearson's correlation
coefficient was used to calculate the coefficient of
correlation.
Results
Quality control of SYD
For the quality control of SYD, a HPLC analysis was
applied in the present study. The chromatogram of a reference
standard and a typical chromatogram obtained from the analyses of
SYD are indicated Fig. 1. The
sulforaphane peak (9.6 min) in the chromatogram of SYD was
identified by comparing the retention time with that of its
reference compound (sulforaphane). The linearity of the calibration
curve for sulforaphane was assessed at six concentration levels
ranging from 0.0281 to 1.4060 mg/ml, and triplicate injections were
applied for each concentration. A calibration curve was constructed
by plotting the integrated chromatographic peak areas (Y) vs. the
corresponding concentration of the injected standard solutions (X).
A least squares regression analysis was employed and a regression
equation (Y=2,623,732.13X+7,853.33), and coefficient of correlation
(r2 was 0.99990) were obtained over relatively wide
concentration ranges for all the analyses. The sulforaphane
concentration in the sample was indicated to equal 0.0624
mg/ml.
Effects of SYD on colon cancer cell
viability
The anti-proliferative effect of SYD in human
colorectal cancer cells was tested and it was revealed that SYD
inhibited the viability of colon cancer cells in a dose- and
time-dependent manner. No significant difference was revealed at
the 24 h time point, but significant differences were noted between
0, 10, 50, 100, 200 and 400 µg/ml SYD following 48 and 72 h of
treatment (P=0.001). The viability of HT-29 and LS-174-T cells
decreased with increasing doses of SYD at 48 and 72 h, respectively
(Fig. 2A and B). At a concentration
of 400 µg/ml, SYD inhibited the viability of HT-29 and LS-174-T
cells. A concentration of 400 µg/ml SYD yielded ~88/80 and 95/92%
inhibition of HT-29/LS-174-T cell viability compared with the
control group (no treatment) at 48 and 72 h, respectively, and the
corresponding half maximal inhibitory concentration
(IC50) values were 103.89/132.05 and 94.23/93.15 µg/ml.
After 48 h of treatment with 0–200 µg/ml SYD, LDH increased in a
dose-dependent manner in the HT29 cell line (Fig. 2C). As a control, the indicated dose of
SYD did not significantly decrease the viability of CRL-1790
(normal human colon epithelial) cells, indicating that SYD exhibits
selective cytotoxicity in colon cancer cells (Fig. 2D).
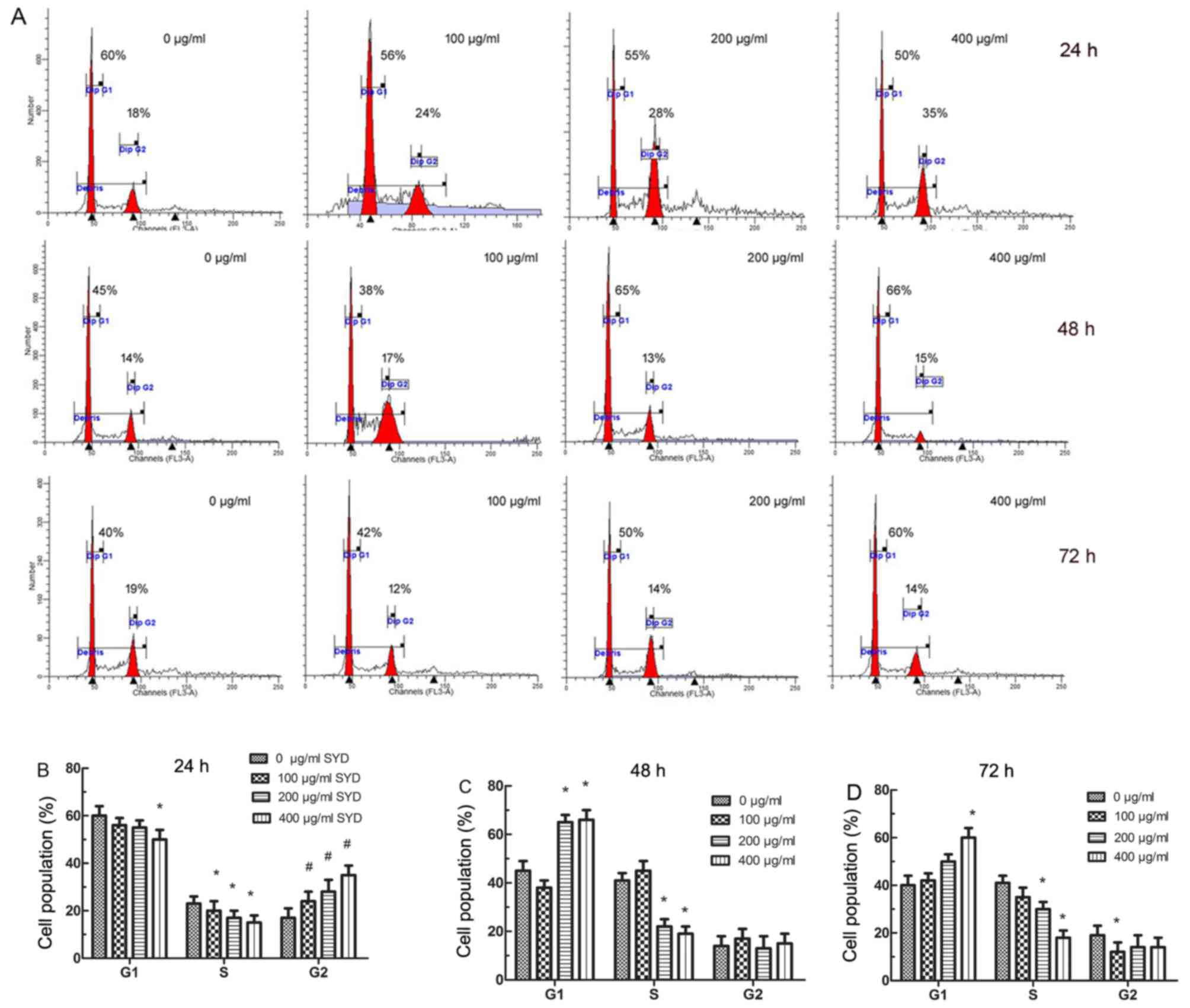
SYD-induced cell cycle changes and
decrease of S-phase cells in HT-29 cells
The effects of SYD on the cell cycle progression of
HT-29 cells are indicated in Fig. 3.
SYD treatment resulted in a significant dose-dependent accumulation
of cells in the G2-M phase at 24 h (P<0.001, compared with the
control; Fig. 3A and B). The
population of G2 cells was increased by 6.2, 8.4 and 18.9%
following treatment with 100, 200 and 400 µg/ml SYD, respectively,
compared with the control. However, marked G1 arrest was observed
with 200 and 400 µg/ml SYD following 48 and 72 h treatment
(Fig. 3A, C and D). The greatest
differences of 41 and 18% were obtained at 48 and 72 h,
respectively, between the 0 and 400 µg/ml SYD groups, indicating
that SYD decreased the population of cells in the S phase.
SYD triggers dose-dependent,
caspase-mediated apoptosis
SYD induced dose-dependent apoptosis of HT-29 and
LS-174-T cells. The apoptosis significantly increased in the 10,
50, 100, 150 and 200 µg/ml SYD groups, compared with the 0 µg/ml
SYD group (P<0.05; Fig. 4A). The
activation of caspases by SYD treatment was subsequently analyzed.
A western blot analysis was performed to verify the rapid caspase
activation observed with the FACScan assay. The results revealed an
accumulation of cleaved CASP8 and CASP3 expression in addition to
an increase in the expression level of cleaved CASP9 (P<0.001,
compared with the 0 µg/ml SYD group; Fig.
4B). SYD-induced apoptosis was significantly decreased by
inhibition of CASP8 (IETD-FMK) compared with control (P<0.05),
whereas the inhibition of CASP9 (LEHD-FMK) had a minimal effect on
SYD-triggered HT-29 cell apoptosis (Fig.
4C). Furthermore, the treatment of HT-29 cells with a
pan-caspase inhibitor (Z-VAD-FMK) significantly delayed SYD-induced
apoptosis compared with control (P<0.05; Fig. 4C).
 | Figure 4.SYD induces apoptosis in a
dose-dependent manner. (A) HT-29 and LS-174-T cells were treated
with various concentrations of SYD (0, 10, 50, 100, 150, and 200
µg/ml) for 24 h. *P<0.05 and #P<0.001 compared with 0 µg/ml.
(B) SYD activates caspases as HT-29 cells were treated with
different doses of SYD for 24 h and incubated with either
Caspase-8, Caspase-3, Caspase-9 or PARP antibodies. #P<0.001
compared with 0 µg/ml SYD (control). (C) HT-29 cells were treated
with 100 µg/ml or 200 µg/ml SYD for 24 h in the presence or absence
of biochemical inhibitors of Caspase-3, Caspase-8, Caspase-9 and
Pan-caspase. *P<0.05 and **P<0.05 compared with control group
(HT-29 cells with no inhibitor treatment). PARP, poly (ADP-ribose)
polymerase; SYD, Su Yang Decoction. |
SYD regulates the expression of
apoptosis-associated genes in colon cancer cells
To determine which apoptosis pathway serves a role
in SYD-induced apoptosis, apoptosis-associated genes were first
assessed by western blot analysis, which revealed that DR (DR3, DR4
and DR5), tumor necrosis factor-α receptor and the corresponding
Fas exhibited minimal alterations in SYD-treated HT-29 cells (data
not shown). Furthermore, a RT-qPCR analysis revealed that the mRNA
levels were unchanged (data not shown). In contrast, as the
concentration of SYD increased, the expression levels of TRAIL and
the Fas ligand (FasLG) also increased. Importantly, the c-FLIP
expression levels significantly decreased in a
concentration-dependent manner (P<0.05 for 10–100 µg/ml SYD and
P<0.001 for 150–200 µg/ml SYD, compared with 0 µg/ml SYD;
Fig. 5A).
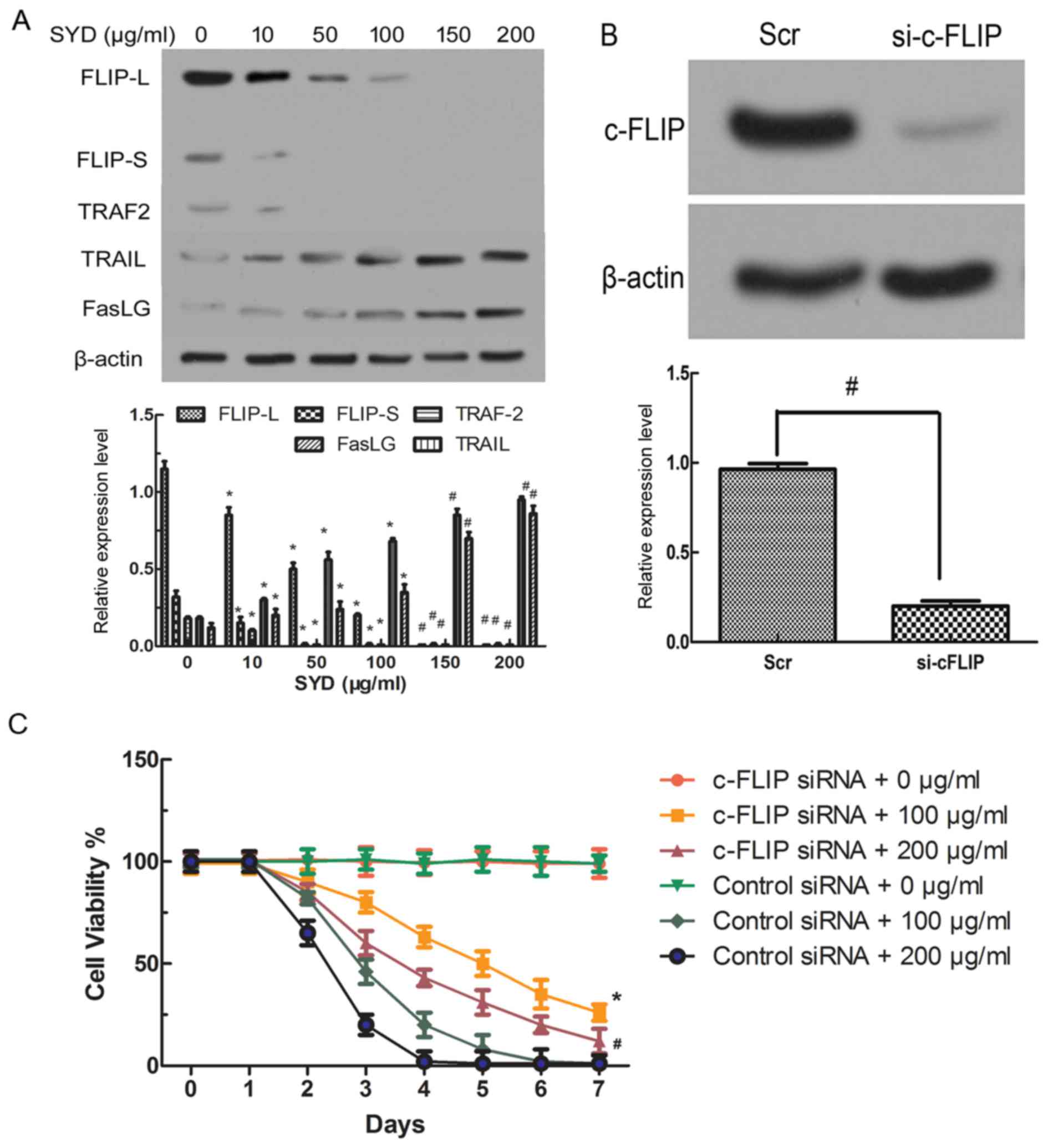 | Figure 5.c-FLIP knockout decreases the
threshold. (A) Expression levels of FLIP-L, FLIP-S, TRAF2, FasLG,
and TRAIL in HT-29 cells treated with different concentrations of
SYD for 48 h. *P<0.05 and #P<0.001 compared with 0 µg/ml SYD.
(B) Expression level of c-FLIP in HT-29 cells transfected with
either control siRNA or siRNA targeting c-FLIP. β-Actin served as
the loading control. *P<0.05 for Scr compared with si-c-FLIP.
(C) The HT-29 cells were seeded in the presence of SYD (100 µg/ml
or 200 µg/ml), and the viability was monitored for 7 days based on
the cell count obtained using a hemocytometer with trypan blue
staining. The results are expressed as the ratio of viable cells to
the total number of cells. *P<0.05 and #P<0.001 compared with
cells transfected with control siRNA and treated with 100 µg/ml or
200 µg/ml SYD, cells transfected with c-FLIP siRNA and not treated
or cells transfected with control siRNA and not treated. TRAIL, TNF
superfamily member 10; FasLG, Fas ligand; TRAF2, TNF receptor
associated factor 2; SYD, Su Yang Decoction; FLICE, FADD-like
interleukin-1β-converting enzyme; FLIP, FLICE-like inhibitory
protein; FLIP-L, long-FLIP; FLIP-S, short FLIP; si/siRNA, small
interfering RNA; Scr, scrambled oligonucleotide; c-, cellular. |
The activation of CASP8 by SYD encouraged the focus
on nuclear factor κβ (NF-κβ)-regulated proteins that might regulate
CASP8 or DR-mediated apoptosis. c-FLIP modulates CASP8 activity by
competing with CASP8 for binding at the death domain, which cleaves
pro-CASP8 (14). c-FLIP has two
alternatively spliced isoforms, short (c-FLIP-S) and long
(c-FLIP-L). Although TRAF2 was activated by different doses of SYD,
its intensity was weaker than that of CASP8 in HT-29 cells
(P<0.05, compared with the 0 µg/ml SYD group; Fig. 5A). The CASP8-processed N-terminal
fragment of c-FLIPL is more efficient than c-FLIPL at recruiting
TRAF2 and RIP1 (15), as was
confirmed by the results of the present study.
c-FLIP decreases the threshold of
SYD-mediated growth inhibition
To verify the potential significance of c-FLIP
downregulation in SYD cytotoxicity, c-FLIP siRNA was transfected
into HT-29 colon cancer cells, which decreased the c-FLIP protein
expression levels compared with Src-transfected cells (P<0.05,
compared with si-c-FLIP; Fig. 5B).
The viability of HT-29 cells was examined with and without c-FLIP
knockdown and it was indicated that the knockdown of c-FLIP in
HT-29 cells decreased sensitivity to SYD, increased viability and
prolonged survival compared with transfection with control siRNA 0,
100 and 200 µg/ml (P<0.05, compared with cells transfected with
control siRNA and treated with 100 µg/ml or 200 µg/ml SYD; Fig. 5C). Comparative proliferation assays of
cells with and without c-FLIP knockdown were performed and no
significant differences were indicated, suggesting that the
observed changes in viability are not an artifact of the altered
proliferation rate (Fig. 5C and
D).
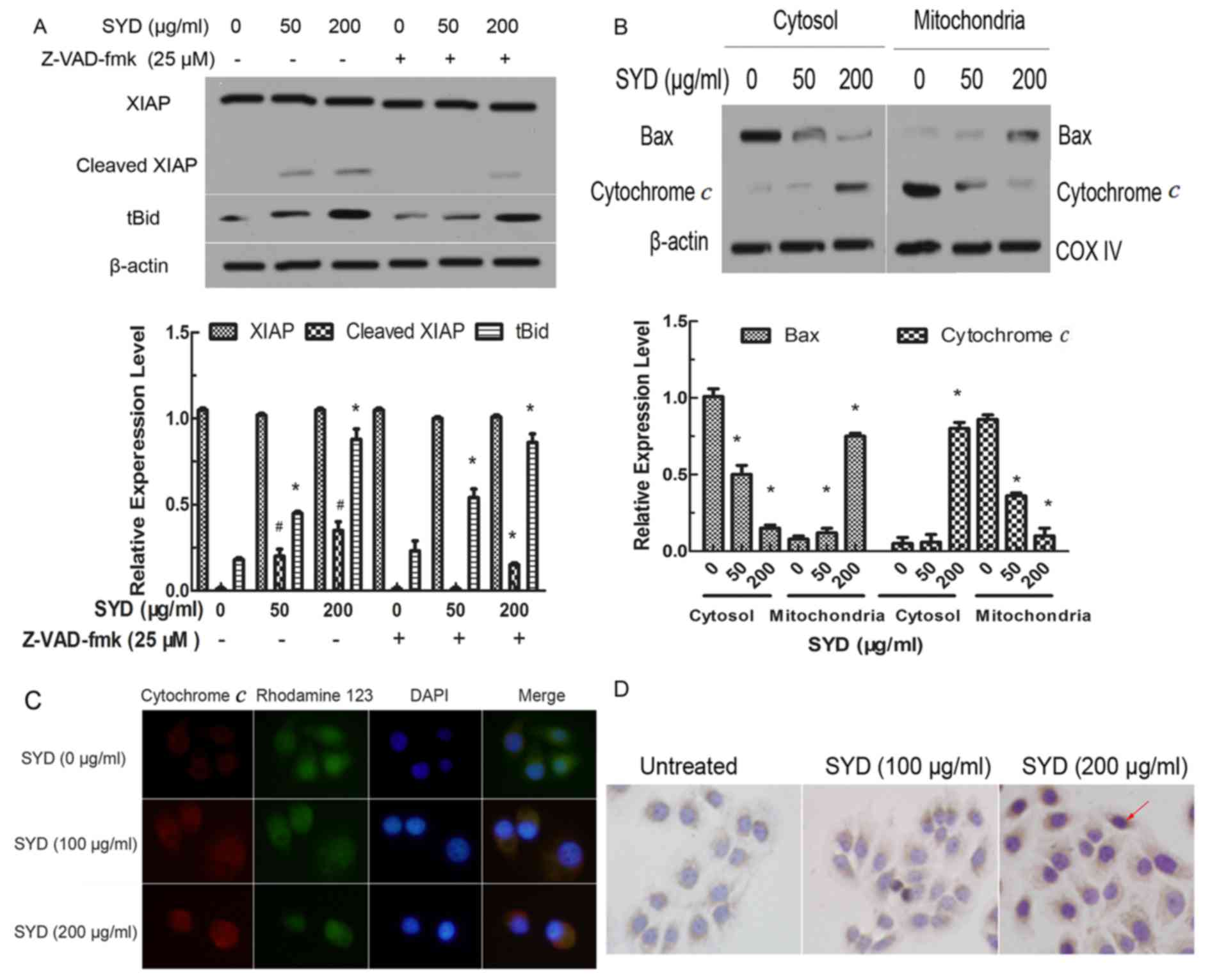
SYD induced XIAP cleavage, the
mitochondrial translocation of Bax and tBid and cytochrome
discharge
To further confirm the SYD-induced apoptosis
pathway, the expression levels of other NF-iβ-regulated proteins
were examined. The results indicated that SYD did not affect the
total expression level of the anti-apoptotic Bcl-2 family members,
including Bcl-2, Bcl-xl, Mcl-1 and Bax (data not shown). XIAP was
indicated to serve as a potent inhibitor of the downstream
effectors CASP3, caspase-7 and CASP9, and XIAP cleavage increased
in a concentration-dependent manner. The Pan-caspase inhibitor
Z-VAD-FMK partially blocked XIAP cleavage, indicating that the
inhibition of viability by SYD in colon cancer is partly due to
caspase-dependent apoptosis induction (P<0.05, compared with the
0, 50 or 100 µg/ml SYD groups; Fig.
6A).
Although the total expression level of pro-apoptotic
Bax was not altered, the cytosolic expression level of Bax was
decreased and the mitochondrial levels were significantly increased
following treatment with increasing concentrations of SYD
(P<0.05). Inversely, increasing concentrations of SYD resulted
in significant increases in the levels of cytochrome c in
the cytosol and therefore significant decreases in the
mitochondrial levels, as determined by western blotting and a
cytochrome c assay (P<0.05, compared with the 0 µg/ml SYD
group; Fig. 6B and C). Cytochrome
c increased with higher doses of SYD (Fig. 6D). Subsequently, a
concentration-dependent accumulation of tBid was further revealed.
(Fig. 6A). Z-VAD-FMK had no effect on
Bid cleavage, indicating a potential caspase-independent underlying
mechanism.
Discussion
Natural products, used as a complementary therapy,
have been reported to serve an important role in the treatment of
patients with advanced cancer (6).
SYD was developed by Infinitus Company Ltd. for the prevention and
treatment of colon cancer. SYD has been indicated to improve
quality of life (data not shown). However, its multiple constituent
compounds and the molecular mechanisms underlying its anticancer
activity require further investigation.
SYD is similar in composition to other prescriptions
of Chinese herbal compounds. For example, Songyou Yin has been
reported to inhibit tumor growth and prolong survival in nude mice
bearing a human hepatocellular carcinoma xenograft with high
metastatic potential (16,17), however its composition is unknown. It
has been reported that SYD comprises two cruciferous vegetable
species with some bioactive constituents that exhibit antitumor
properties (4,18). It has been demonstrated that
sulforaphane, allyl isothiocyanate and phenethyl isothiocyanate in
broccoli prevent liver cancer, lung cancer, prostate cancer, breast
cancer, rectal cancer and stomach cancer (7). However, it has been indicated that
sulforaphane is the best constituent to inhibit the aforementioned
cancer types and therefore, sulforaphane constitutes the optimal
choice for quality control (8). The
present experiment was repeated and it was indicated that the peak
at 9.6 min in the SYD sample corresponds with that in the standard
sample. Mass spectrum data further demonstrated that molecular
weight 177.2 from sulforaphane standards was in accordance with
that in SYD (data not shown), which was hydrolyzed by glucoraphanin
in SYD.
Previous epidemiological evidence has indicated that
a diet high in cruciferous vegetables may reduce the risk of breast
cancer in females (19). The
aforementioned data are further supported by data from experimental
rodent models treated with the main bioactive components of
cruciferous vegetables (data not shown). It has been demonstrated
that cell cycle arrest is induced by sulforaphane, which promotes
breast cancer cell apoptosis in vitro (20). Sulforaphane has also been indicated to
inhibit chemically induced breast cancer in female rats (21), suggesting that sulforaphane may have
anticancer activities. Although there are many unknown constituents
in SYD, sulforaphane, as the key ingredient in SYD, was tested by
HPLC for the quality control of SYD.
The main objective of the present study was to
evaluate the anticancer efficacy and associated underlying
mechanisms of SYD in human colorectal cancer cells in vitro.
It was indicated that SYD inhibited the viability and colony
formation of HT-29 and LS-174-T cells. Furthermore, the
aforementioned result was also demonstrated by an LDH assay in
SYD-treated HT-29 cells (Fig. 2D). In
contrast, normal CRL-1790 cells were minimally affected by SYD,
even at concentrations that were highly toxic to colon cancer
cells. The aforementioned data highlight the importance of broccoli
and green cabbage consumption, as natural products, for the
prevention and treatment of cancer. In subsequent experiments, SYD
was reported to inhibit cell proliferation by inducing cell cycle
arrest, specifically in the S phase, and apoptosis in human
colorectal cancer cells in vitro.
Mechanistic experiments from the present study
indicated that the anti-colon cancer activity of SYD was associated
with the activation of CSPs and c-FLIP. Notably, SYD has been
reported to trigger minimal CASP9 cleavage, however robust CASP8
cleavage was observed. The aforementioned data are consistent with
the results of the present study, which indicated that biochemical
inhibitors of CASP8 had a greater impact on the anti-colon cancer
activity of SYD, suggesting that CASP9 serves a minor role in
inducing colon cancer apoptosis. Western blot analysis and RT-qPCR
assay demonstrated that although the expression levels of DRs,
including as DR4, DR5 and Fas, remained unchanged following SYD
treatment, SYD considerably increased the levels of their ligands,
including TRAIL and FasL (Fig.
5B).
It has been reported that c-FLIP is structurally
similar to CASP8 and the expression level of c-FLIP isoforms is
increased in various types of cancer, including colon cancer
(22,23). Furthermore, it has been demonstrated
that the direct siRNA-mediated silencing of c-FLIP increases CASP8
recruitment to the death-enhancing domain and apoptosis induction
in some cancer models (24). c-FLIP
represents a critical target for therapeutic intervention, which
inhibits its transcription and posttranscriptional modification
(25). Therefore, the present study
suggests that the main toxic effect of SYD results from the
downregulation of c-FLIP mediated by SYD. In other words, c-FLIP
siRNA decreased the SYD dose needed for apoptosis, suggesting that
c-FLIP downregulation is important for SYD cytotoxicity.
Although the expression levels of Bcl-2 and Bax in
HT-29 cells were unaffected by SYD, a concentration-dependent
increase in the mitochondrial levels of Bax was revealed subsequent
to SYD treatment, along with a decrease in the cytoplasmic Bax
expression levels. The data of the present study further
demonstrated that SYD treatment yielded a dose-dependent decrease
in the release of cytochrome c from the mitochondria
accompanied by an increase in the cytosol cytochrome c
level. It has been reported that because the release of cytochrome
c requires mitochondrial membrane insertion and the
oligomerization of Bax, the translocation of Bax proteins from the
cytosol to the mitochondria represents a key event in the
activation of the intrinsic pathway (25). tBid rapidly binds to membranes and
interacts with Bax, causing the insertion of Bax into the membrane
and oligomerization, resulting in membrane permeabilization
(26). Furthermore, it has been
demonstrated that active CASP8 additionally mediates the
proteolytic cleavage of Bid to tBid, which is translocated to the
mitochondria and amplifies the intrinsic apoptotic pathway
(27). Therefore, the data of the
present study indicate that SYD induces Bid truncation and Bax
translocation to the mitochondria from the cytosol, leading to the
release of cytochrome c, enhanced mitochondrial dysfunction
and the induction of apoptosis in HT-29 cells.
The results of the present study indicated that SYD
has anticancer activity in HT-29 and LS-174-T cells and that this
formula activates apoptosis and modulates cell cycle regulators by
the activation of caspases. Future studies are required to further
investigate the exact components of SYD affecting cancer cells in
addition to examining the ability of SYD to prevent and treat
cancer, the associated underlying mechanism of action and the
food-drug interactions of SYD with conventional cancer
preventatives and therapeutics. Further testing of control cell
lines is required along with in vivo experiments.
Acknowledgements
The authors would like to thank Dr He Wang and Mr
Lei Ma (Cancer treatment center, the Affiliated Second Hospital of
Guangzhou Medical University, Guangzhou, China) for their technical
assistance.
Funding
The present study was financially supported by the
School Enterprise Cooperation Fund (grant no., 1562005; Guangzhou,
China) and the Guangdong Province Medical Science and Technology
Research Fund (grant no., A2015530; Guangdong, China).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ designed the study and interpreted the results.
YG, QG, FY, XZ and YL collected the test data and drafted the
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Animal Care Committee and the Local Veterinary Office and Ethics
Committee of Guangzhou Medical University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tei M, Otsuka M, Suzuki Y, Kishi K,
Tanemura M and Akamatsu H: Safety and feasibility of single-port
laparoscopic multivisceral resection for locally advanced left
colon cancer. Oncol Lett. 15:10091–10097. 2018.PubMed/NCBI
|
|
2
|
Jin P, Yao D, Xu F, Wang H and Zheng Y:
Effect of light on quality and bioactive compounds in postharvest
broccoli florets. Food Chem. 172:705–709. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang K, Zhang C, Yu B, Chen B, Liu Z, Hou
C, Wang F, Shen H and Chen Z: Autophagic degradation of FOXO3a
represses the expression of PUMA to block cell apoptosis in
cisplatin-resistant osteosarcoma cells. Am J Cancer Res.
7:1407–1422. 2017.PubMed/NCBI
|
|
4
|
Atay K, Canbakan B, Koroglu E, Hatemi I,
Canbakan M, Kepil N, Tuncer M and Senturk H: Apoptosis and disease
severity is associated with insulin resistance in non-alcoholic
fatty liver disease. Acta Gastroenterol Belg. 80:271–277.
2017.PubMed/NCBI
|
|
5
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang D, Zhang Y, Lu J, Wang Y, Wang J,
Meng Q, Lee RJ, Wang D and Teng L: Cordycepin, a natural
antineoplastic agent, induces apoptosis of breast cancer cells via
caspase-dependent pathways. Nat Prod Commun. 11:63–68.
2016.PubMed/NCBI
|
|
7
|
Rose P, Huang Q, Ong CN and Whiteman M:
Broccoli and watercress suppress matrix metalloproteinase-9
activity and invasiveness of human MDA-MB-231 breast cancer cells.
Toxicol Appl Pharmacol. 209:105–113. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang TT, Schoene NW, Milner JA and Kim YS:
Broccoli-derived phytochemicals indole-3-carbinol and
3,3′-diindolylmethane exerts concentration-dependent pleiotropic
effects on prostate cancer cells: Comparison with other cancer
preventive phytochemicals. Mol Carcinog. 51:244–256. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hensley P, Mishra M and Kyprianou N:
Targeting caspases in cancer therapeutics. Biol Chem. 394:831–843.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu X, Yue P, Schonthal AH, Khuri FR and
Sun SY: Cellular FLICE-inhibitory protein down-regulation
contributes to celecoxib-induced apoptosis in human lung cancer
cells. Cancer Res. 66:11115–11119. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mitupatum T, Aree K, Kittisenachai S,
Roytrakul S, Puthong S, Kangsadalampai S and Rojpibulstit P: Hep88
mAb-mediated paraptosis-like apoptosis in HepG2 cells via
downstream upregulation and activation of caspase-3, caspase-8 and
caspase-9. Asian Pac J Cancer Prev. 16:1771–1779. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fang S, Zhu W, Zhang Y, Shu Y and Liu P:
Paeoniflorin modulates multidrug resistance of a human gastric
cancer cell line via the inhibition of NF-kappaB activation. Mol
Med Rep. 5:351–356. 2012.PubMed/NCBI
|
|
13
|
Wang H, Yin H, Yan F, Sun M, Du L, Peng W,
Li Q, Feng Y and Zhou Y: Folate-mediated mitochondrial targeting
with doxorubicin-polyrotaxane nanoparticles overcomes multidrug
resistance. Oncotarget. 6:2827–2842. 2015.PubMed/NCBI
|
|
14
|
Pop C, Oberst A, Drag M, Van Raam BJ,
Riedl SJ, Green DR and Salvesen GS: FLIP(L) induces caspase 8
activity in the absence of interdomain caspase 8 cleavage and
alters substrate specificity. Biochem J. 433:447–457. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dohrman A, Kataoka T, Cuenin S, Russell
JQ, Tschopp J and Budd RC: Cellular FLIP (long form) regulates CD8+
T cell activation through caspase-8-dependent NF-kappa B
activation. J Immunol. 174:5270–5278. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang XY, Huang ZL, Wang L, Xu YH, Huang
XY, Ai KX, Zheng Q and Tang ZY: Herbal compound ‘Songyou Yin’
reinforced the ability of interferon-alfa to inhibit the enhanced
metastatic potential induced by palliative resection of
hepatocellular carcinoma in nude mice. BMC Cancer. 10:5802010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng S, Jia Q, Shen H, Xu X, Ling J, Jing
C and Zhang B: Treatment with the herbal formula Songyou Yin
inhibits epithelial-mesenchymal transition in hepatocellular
carcinoma through downregulation of TGF-β1 expression and
inhibition of the SMAD2/3 signaling pathway. Oncol Lett.
13:2309–2315. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kapiszewska M: A vegetable to meat
consumption ratio as a relevant factor determining cancer
preventive diet. The Mediterranean versus other European countries.
Forum Nutr. 59:130–153. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ambrosone CB, McCann SE, Freudenheim JL,
Marshall JR, Zhang Y and Shields PG: Breast cancer risk in
premenopausal women is inversely associated with consumption of
broccoli, a source of isothiocyanates, but is not modified by GST
genotype. J Nutr. 134:1134–1138. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tseng E, Scott-Ramsay EA and Morris ME:
Dietary organic isothiocyanates are cytotoxic in human breast
cancer MCF-7 and mammary epithelial MCF-12A cell lines. Exp Biol
Med (Maywood). 229:835–842. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pajak B, Turowska A, Orzechowski A and
Gajkowska B: Bisindolylmaleimide IX facilitates extrinsic and
initiates intrinsic apoptosis in TNF-alpha-resistant human colon
adenocarcinoma COLO 205 cells. Apoptosis. 13:509–522. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li TW, Zhang Q, Oh P, Xia M, Chen H,
Bemanian S, Lastra N, Circ M, Moyer MP, Mato JM, et al:
S-Adenosylmethionine and methylthioadenosine inhibit cellular FLICE
inhibitory protein expression and induce apoptosis in colon cancer
cells. Mol Pharmacol. 76:192–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sung B, Prasad S, Ravindran J, Yadav VR
and Aggarwal BB: Capsazepine, a TRPV1 antagonist, sensitizes
colorectal cancer cells to apoptosis by TRAIL through
ROS-JNK-CHOP-mediated upregulation of death receptors. Free Radic
Biol Med. 53:1977–1987. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moriwaki K, Shinzaki S and Miyoshi E:
GDP-mannose-4,6-dehydratase (GMDS) deficiency renders colon cancer
cells resistant to tumor necrosis factor-related apoptosis-inducing
ligand (TRAIL) receptor- and CD95-mediated apoptosis by inhibiting
complex II formation. J Biol Chem. 286:43123–43133. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kadenbach B, Arnold S, Lee I and Huttemann
M: The possible role of cytochrome c oxidase in
stress-induced apoptosis and degenerative diseases. Biochim Biophys
Acta. 1655:400–408. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lovell JF, Billen LP, Bindner S,
Shamas-Din A, Fradin C, Leber B and Andrews DW: Membrane binding by
tBid initiates an ordered series of events culminating in membrane
permeabilization by Bax. Cell. 135:1074–1084. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu Z, Tang K, Wang M, Rao Q, Liu B and
Wang J: A new caspase-8 isoform caspase-8s increased sensitivity to
apoptosis in Jurkat cells. J Biomed Biotechno. 2009:9304622009.
|