Introduction
Taxol (paclitaxel) is a valuable cancer
chemotherapeutic agent used for the treatment of numerous types of
cancer, including ovary, breast, oral and lung carcinomas (1–4). The
primary cellular targets of Taxol are the microtubules of cancer
cells, which are vital for mitotic activity, cellular motility and
proliferative capacity (3). Taxol
is also known to block cell replication, arrest cells in the
G2/M phase of the cell cycle and induce apoptosis
(5,6). Despite significant clinical responses
initially, the majority of patients eventually develop resistance
to Taxol. Currently, mechanisms accounting for Taxol resistance
include alterations to the tubulin structure (7–9),
changes in the drug-binding affinity of the microtubules (10) and cell cycle deregulation (11). However, the detailed molecular
mechanisms that may contribute to the Taxol resistance of cancer
cells are not fully understood.
Cisplatin (CDDP) is a DNA-damaging agent that
induces cytotoxicity through the production of DNA damage caused by
the formation of CDDP-DNA adducts (12), which leads to irreparable DNA damage
and ultimately, cell death. However, similar to Taxol, the
development of CDDP resistance in cancer cells is a major
impediment in clinical treatment (13). Currently, the mechanisms of CDDP
resistance remain unclear. It has been reported that CDDP and other
anticancer agents induce the activation of the epidermal growth
factor receptor in multiple cancer cells that overexpress the
receptor (14). In addition,
another study described the fact that the uptake of various
compounds, including nutrients such as glucose, was reduced in
CDDP-resistant hepatoma cells compared with CDDP-sensitive cells
(15). Moreover, a recent study
reported that the knockdown of GLUT-1, which is a glucose
transporter on the cell membrane, facilitated CDDP treatment,
resulting in increased rates of apoptosis in oral cancer cells
under hypoxic conditions (16),
indicating that the glucose metabolism pathway is involved in CDDP
resistance.
Lactate dehydrogenase-A (LDHA) is one of the
principle isoforms of LDH that is expressed in breast tissue. LDHA
controls the conversion of pyruvate to lactate and plays a
significant role in glucose metabolism (17). A study has also shown that LDHA is
important in Taxol resistant breast cancer cells (18). This study detected increased LDHA
expression and activity in Taxol-resistant cells, and identified
that the sensitivity of Taxol-resistant cells to Taxol was
significantly increased by the downregulation of LDHA by small
interfering (si)RNA, indicating that LDHA may be a therapeutic
target for overcoming Taxol resistance.
As previous studies have shown that CDDP treatment
inhibits LDHA expression in lung cancer cells (15), in the present study investigated
whether LDHA is downregulated by CDDP treatments in oral squamous
cancer cells. In addition, the role of LDHA in Taxol resistance was
investigated. Whether synergistic cytotoxicity of cisplatin and
Taxol on Taxol resistant oral cancer cells occurs via the
inhibition of LDHA was also explored. This study will provide a
theoretical explanation to support the combined treatment of Taxol
and CDDP to develop clinical chemotherapeutic strategies for oral
cancer patients.
Materials and methods
Cell culture and conditions
The human oral squamous cell carcinoma (OSCC) cell
lines, OECM-1 and H-1, were purchased from the American Type
Culture Collection (Manassas, VA, USA). The cultivation conditions
were used as described previously (19). Briefly, the cells were routinely
cultured in Dulbecco’s modified Eagle’s medium (DMEM; Gibco BRL,
Paisley, UK) containing 10% fetal bovine serum (FBS; HyClone,
Logan, UT, USA), at 37°C in a humid atmosphere with 5%
CO2.
Cell viability assay
The cancer cells were treated with Taxol,
5-fluorouracil (FU) or CDDP with the indicated concentrations for
24 h. The cells were seeded in a 96-well plate, at a density of
3,000 cells/well in 0.2 ml DMEM containing 10% FBS. Following
overnight incubation under the same cultivating conditions, each
well was refreshed with 0.2 ml serum-free medium (SFM) for another
day. The cells were then treated with 0.2 ml SFM containing various
concentrations of Taxol, CDDP, 5-FU, Taxol/CDDP or Taxol/5-FU. The
drug-containing SFM was refreshed after 2 days and incubated under
the same conditions for another 2 days. Finally, cell viability was
accessed with an MTT reagent (Sigma-Aldrich, Inc., St. Louis, MO,
USA), and by measuring the absorbance at 590 nm with a plate
reader. Relative viability was obtained from the absorbance at 590
nm of the drug-treated OECM-1 cells divided by the absorbance at
590 nm of the untreated OECM-1 cells. The same experiment was
repeated three times.
Western blotting and antibodies
The cells were harvested and lysed in a buffer
containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 2 mM EDTA, 1%
Triton, 1 mM PMSF and a protease inhibitor cocktail (Sigma-Aldrich)
for 20 min on ice. Lysates were separated by centrifugation at
16,000 × g, at 4°C for 10 min. Supernatants were collected and
protein concentrations were determined by the Bradford assay
(Bio-Rad, Hercules, CA, USA). The proteins were then separated with
an SDS/polyacrylamide gel and transferred to a nitrocellulose
membrane (Bio-Rad). Subsequent to being blocked in
phosphate-buffered saline (PBS) with 5% skimmed dry milk for 1 h,
the membranes were incubated overnight at 4–8°C with the primary
antibodies in PBS with 5% skimmed dry milk. The following
antibodies were utilized: Anti-LDHA rabbit antibody (1:1,000; Cell
Signaling Technology, Inc., Danvers, MA, USA) and anti-β-actin
monoclonal antibody (1:2,000; Sigma-Aldrich). Membranes were
extensively washed with PBS and incubated with horseradish
peroxidase conjugated secondary anti-mouse antibody or anti-rabbit
antibody (1:2,000; Bio-Rad). Subsequent to additional washes with
PBS, antigen-antibody complexes were visualized with an enhanced
chemiluminescence kit (Pierce Biotechnology, Inc., Rockford, IL,
USA).
Generation of Taxol- and CDDP-resistant
cell lines
The Taxol- and CDDP-resistant cell lines were
generated according to previously described methods (18,19).
Briefly, OECM-1 Taxol-resistant (Tax R) or (Cis R) CDDP-resistant
cells were developed from parental OECM-1 cells by treating them
with gradually increasing concentrations of Taxol or CDDP in
regular cell culture medium. Tax R or Cis R single or pooled clones
were identified and cultured. All resistant cells were verified by
the treatments every four weeks.
siRNA and plasmid DNA transfection
siRNA oligonucleotides for LDHA were purchased from
Sigma-Aldrich, with a scrambled siRNA (Sigma-Aldrich) used as a
control. A vector containing wild-type LDHA was purchased from
Origene (RC209378; Rockville, MD, USA). Transfection was performed
using the Oligofectamine Transfection Reagent (Invitrogen Life
Technologies, Carlsbad, CA, USA) according to the manufacturer’s
protocol. At 48 h post-transfection, whole-cell lysates were
prepared for further analysis.
Statistical analysis
The unpaired Student’s t-test was used for the data
analysis. All data are presented as the mean ± standard error.
P<0.05 was considered to indicate a statistically significant
difference.
Results
LDHA is downregulated in response to CDDP
treatment
The present study investigated which signaling
pathway may be involved in CDDP resistance, according to LDHA
expression. Following screening, which involved the investigation
of mitochondrial oxidation consumption, the AKT and mTOR pathway
and microRNAs which may target glycolysis and mitochondrial
apoptosis pathways, including cytochrome c and Bcl-2, the
expression of LDHA was found to be significantly decreased
subsequent to CDDP treatment at varying doses in two oral cancer
cell lines (P<0.05; Fig. 1A).
Next, a CDDP-resistance cell line was generated. The OECM-1 cells,
which have been widely used as a model of oral cancer in previous
studies, were treated with gradually increasing concentrations of
CDDP in cell culture medium for the selection of resistant cells.
Following successive treatments for 3 months, several resistant
cell clones were developed (Fig.
1B). More of the Cis R cells remained alive following the
addition of 50 and 100 μM CDDP, while the OECM-1-sensitive (Cis S)
cells exhibited significant cell death following the 50- and 100-μM
treatments (P<0.05 and P<0.01, respectively). We hypothesized
that the expression of LDHA should be altered in the Cis R cells,
and the data in Fig. 1C confirmed
this by showing that the level of LDHA expression in the Cis R
cells was decreased. Taken together, these results revealed a tight
correlation between the expression of LDHA and CDDP resistance.
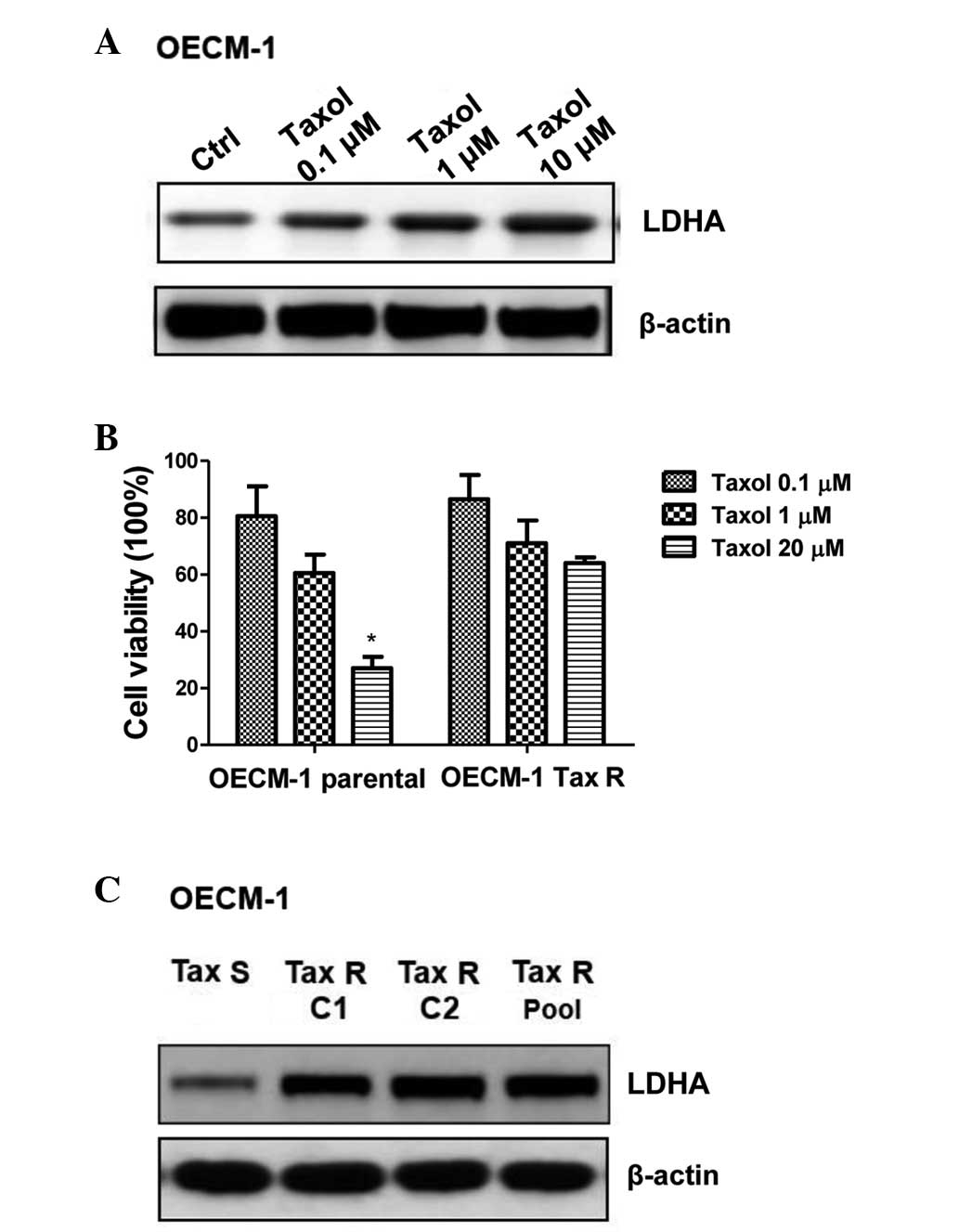
Tax R cells display upregulated LDHA
expression
It has been reported that LDHA has a significant
role in Taxol-resistant breast cancer cells (18). The present study next checked the
LDHA expression levels in response to Taxol treatment in the oral
cancer cells. Fig. 2A shows that
Taxol treatment at varying concentrations induced LDHA expression.
Similar to the generation of CDDP resistance cell line, a
Taxol-resistant cell line was generated for OECM-1. More of the
OECM-1 Taxol-resistant (Tax R) cells remained alive following the
addition of 1 and 20 μM CDDP, while the OECM-1 Taxol-sensitive (Tax
S) cells exhibited significant cell death following the 1- and
20-μM treatments (Fig. 2B;
P<0.05). As expected, the LDHA expression level was upregulated
in the Tax R cells, indicating that LDHA is an important glycolytic
enzyme involved in Taxol-resistance (Fig. 2C).
Downregulation of LDHA re-sensitizes oral
cancer cells to Taxol
To further support the results, LDHA-knockdown was
performed using siRNA specific to LDHA in the OECM-1 cells,
followed by the measurement of the cell sensitivities to the Taxol
and CDDP treatments. Fig. 3A shows
the efficient knockdown of LDHA, which rendered the oral cells
insensitive to CDDP treatment, but sensitive to Taxol
treatment.
 | Figure 3Lactate dehydrogenase-A (LDHA) plays
reverse roles in response to Taxol and cisplatin (CDDP) treatments.
(A) Knockdown of LHDA sensitized the OECM-1 cells to Taxol
treatment, but desensitized the OECM-1 cells to the CDDP treatment.
The OECM-1 cells were transfected with small interfering (si)RNA
for 48 h, then treated with CDDP at 10, 50 and 100 μM or Taxol at
0.1, 0.5 and 1 μM, followed by the measurement of cell viability.
(B) Overexpression of LDHA sensitized the OECM-1 cells to CDDP
treatment, but desensitized the OECM-1 cells to Taxol treatment.
The OECM-1 cells were transfected with overexpression vector
containing wild-type LDHA (V) for 48 h, then treated with CDDP at
5, 10 and 50 μM or Taxol at 0.1, 5 and 20 μM, followed by the
measurement of cell viability. The columns represent the mean of
three independent experiments, and the bars represent the standard
error. *P<0.05 and **P<0.01 vs.
control. |
Upregulation of LDHA re-sensitizes oral
cancer cells to CDDP
The results were then verified by the overexpression
of LDHA by transient transfection of a vector containing wild-type
LDHA. The OECM-1 cells consistently exhibited a higher level of
LDHA and an acquired resistance to Taxol, but became vulnerable to
CDDP (Fig. 3B). Taken together,
these data showed that LDHA plays reverse roles in the response to
Taxol and CDDP.
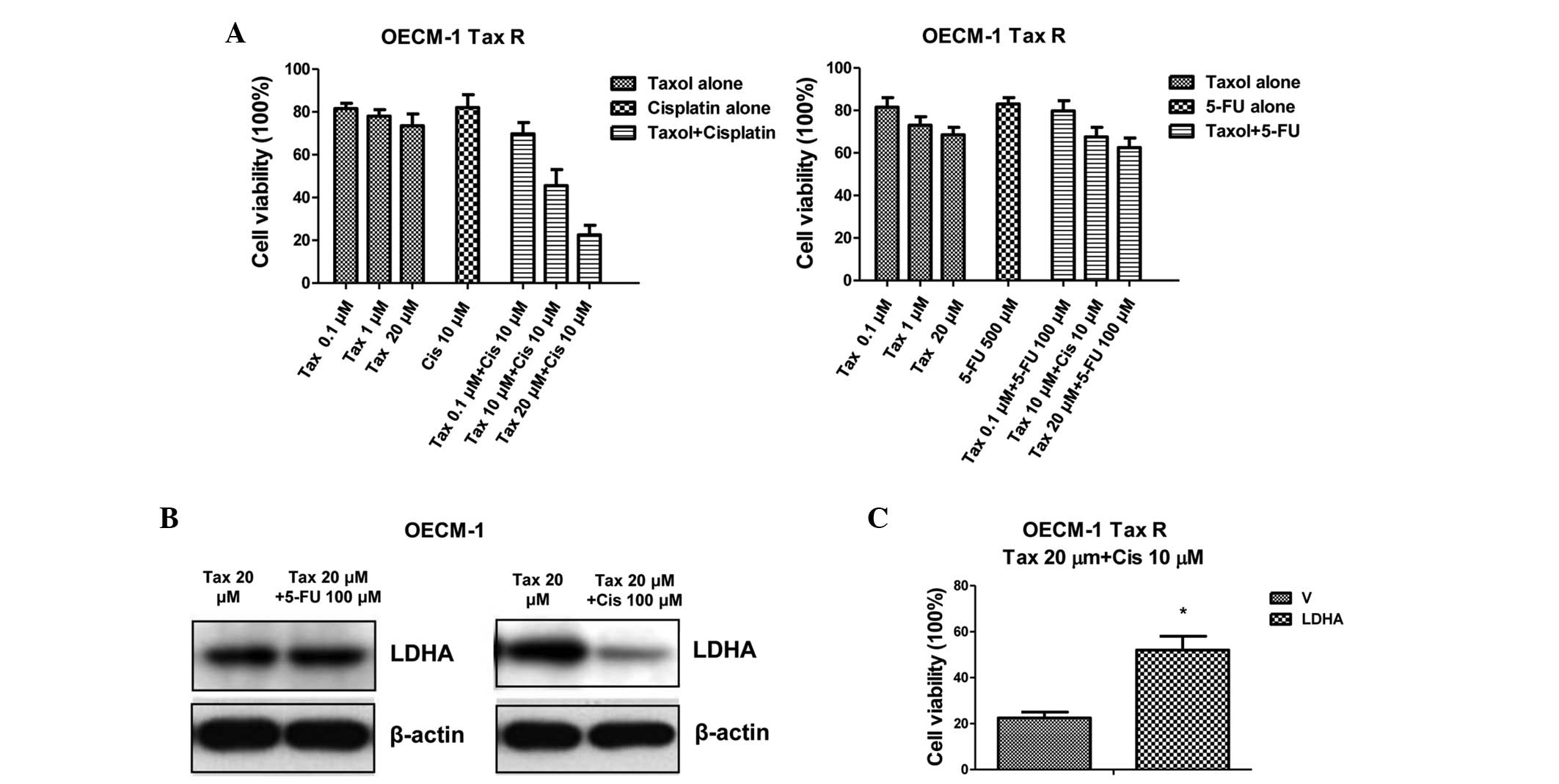
Combination of Taxol and CDDP shows a
synergistic effect on Tax R cells by blocking LDHA expression
Since it has been reported that the combination of
Taxol with CDDP treatment shows synergistic cytotoxicity in OECM-1
cells (19). In the present study,
experiments were designed to examine whether treating
Taxol-resistant oral cancer cells with CDDP would result in
synergistically therapeutic effects. Fig. 4A shows that treatment with Taxol or
CDDP alone in the Tax R cells exhibited no significant inhibitory
effects, but that combining Taxol and CDDP together resulted in
significant inhibition of cell viability (Fig. 4A; P<0.05). Since it has been
reported that 5-FU-resistant cancer cells exhibit upregulated
glucose metabolism (20), the
present study then treated the Tax R cells with a combination of
Taxol and 5-FU. The results showed that the combination of Taxol
and 5-FU did not generate better chemotherapeutic effects (Fig. 4A), indicating that the synergistic
inhibitory effects of Taxol plus CDDP acted through the inhibition
of the expression of LDHA by CDDP in the Tax R cells. To support
this conclusion, the LDHA expression was compared for the combined
treatments; the LDHA expression level was significantly
downregulated by Taxol and CDDP (P<0.05), but no change was
evident following treatment with Taxol and 5-FU (Fig. 4B). When LDHA was exogenously
overexpressed, the OECM-1 parental cells obtained resistance to the
combined treatments (Fig. 4C),
indicating that the downregulation of LDHA specifically accounted
for the mechanisms of the synergistic effects of the combination of
Taxol and CDDP. In summary, these results suggested that CDDP plays
essential roles in overcoming Taxol resistance in oral cancer
cells.
Discussion
CDDP is employed for the treatment of a wide array
of solid malignancies via multiple mechanisms, such as those that
involve the steps prior to CDDP binding to DNA (pre-target
resistance), those directly associating DNA-CDDP adducts (on-target
resistance), those associated with the lethal signaling pathway(s)
induced by CDDP-mediated DNA damage (post-target resistance) and
those that affect molecular circuitry without clear links to
CDDP-induced signals (off-target resistance) (13). LDH-A is one of the main isoforms of
LDH expressed in breast tissue, catalyzing the conversion of
pyruvate to lactate, which is a key step in glucose metabolism. It
has previously been shown that LDH-A plays a vital role in
glycolysis, growth properties and tumor maintenance, as well as in
the chemoresistance of breast cancer cells (18). To date, few studies have focused on
the association between glucose metabolism and CDDP resistance in
oral cancer cells. However, it has been shown that CDDP-resistant
cancer cells commonly exhibit slower proliferation rates and
reduced uptake levels of various compounds, including nutrients
(15). Another study has reported
the function of GLUT-1 in CDDP resistance. This study reported the
silencing of GLUT-1-sensitized oral cancer cells to CDDP during
hypoxia (16), indicating that
CDDP-resistant cells had an impaired glucose metabolism
pathway.
Taxol is a widely used chemotherapeutic agent for
the treatment of several types of cancer, including oral cancer.
Taxol resistance may result in the subsequent recurrence and
metastasis of cancer. The specific mechanisms involved remain
poorly understood, although extensive investigations have been
conducted. It has been reported that treatment with the combination
of Taxol and CDDP in human OSCC shows a synergistic effect
(19), although the mechanisms
remain unclear. In the present study, it was found that CDDP
treatment resulted in decreased LDHA expression levels in the oral
cancer cells, while Taxol treatment showed the reverse results,
with an increased level of LDHA expression. In addition,
Taxol-resistant cells showed increased LDHA expression and the
CDDP-resistant cells showed decreased LDHA expression.
The downregulation of LDHA by LDHA siRNA increased
the sensitivity of the cells to Taxol, but increased their
resistance to CDDP. This indicated that Taxol treatment triggers
the glucose metabolism pathway to ensure cancer cell survival, most
likely through promoting cell glycolysis. A previous study has
shown that cancer cells inhibit cytochrome c-mediated
apoptosis by a mechanism of deregulated glucose metabolism
(21). Thus, the Taxol-induced high
expression levels and activity of LDHA detected in Taxol-resistant
cells could be an adaptation of these cells to Taxol treatment and
may be used to modulate glucose metabolism and glycolysis to avoid
the apoptosis induced by Taxol. As aforementioned, CDDP showed a
high efficiency in the inhibition of LDHA, which may result in the
further examination of the synergistic effects of the combination
of these two drugs. As expected, CDDP re-sensitized the Tax R cells
through the inhibition of LDHA, and this effect was reversed
following overexpression of LDHA in the parental oral cancer cells.
Our future studies will continue to investigate the therapeutic
effects on the recovery of chemoresistance through the alteration
of glucose metabolism by specific glycolysis inhibitors. Taken
together, the results of this study indicate that LDH may
potentially serve as a target for overcoming Taxol resistance in
human breast cancer patients.
Acknowledgements
The authors would like to thank the staff and
faculty of the Oral Medical Research Center of the Chinese People’s
Liberation Army General Hospital (Beijing, China), and also Dr Yu
Wan Dong for editorial assistance. The authors are grateful for the
support provided by the Chinese National Postdoctoral Fellowship
(Chinese Academy of Science; grant no. 2013-52-134523). This paper
was also supported by the China Postdoctoral Science Foundation
(grant no. 2014M552645).
References
|
1
|
Stordal B, Pavlakis N and Davey R: A
systematic review of platinum and taxane resistance from bench to
clinic: an inverse relationship. Cancer Treat Rev. 33:688–703.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lai D, Ho KC, Hao Y and Yang X: Taxol
resistance in breast cancer cells is mediated by the hippo pathway
component TAZ and its downstream transcriptional targets Cyr61 and
CTGF. Cancer Res. 71:2728–2738. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gonçalves A, Braguer D, Kamath K, Martello
L, Briand C, Horwitz S, Wilson L and Jordan MA: Resistance to Taxol
in lung cancer cells associated with increased microtubule
dynamics. Proc Natl Acad Sci USA. 98:11737–11742. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cheung CH, Wu SY, Lee TR, et al: Cancer
cells acquire mitotic drug resistance properties through beta
I-tubulin mutations and alterations in the expression of
beta-tubulin isotypes. PLoS One. 5:e125642010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen LP, Cai SM, Fan JX and Li ZT: PEBA
regimen (cisplatin, etoposide, bleomycin, and adriamycin) in the
treatment of drug-resistant choriocarcinoma. Gynecol Oncol.
56:231–234. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Donnenberg VS and Donnenberg AD: Multiple
drug resistance in cancer revisited: the cancer stem cell
hypothesis. J Clin Pharmacol. 45:872–877. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kavallaris M, Kuo DY, Burkhart CA, et al:
Taxol-resistant epithelial ovarian tumors are associated with
altered expression of specific beta-tubulin isotypes. J Clin
Invest. 100:1282–1293. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Panda D, Miller HP, Banerjee A, Ludueña RF
and Wilson L: Microtubule dynamics in vitro are regulated by the
tubulin isotype composition. Proc Natl Acad Sci USA.
91:11358–11362. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Martello LA, Verdier-Pinard P, Shen HJ, et
al: Elevated levels of microtubule destabilizing factors in a
Taxol-resistant/dependent A549 cell line with an alpha-tubulin
mutation. Cancer Res. 63:1207–1213. 2003.PubMed/NCBI
|
|
10
|
Orr GA, Verdier-Pinard P, McDaid H and
Horwitz SB: Mechanisms of Taxol resistance related to microtubules.
Oncogene. 22:7280–7295. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tan M, Jing T, Lan KH, et al:
Phosphorylation on tyrosine-15 of p34(Cdc2) by ErbB2 inhibits
p34(Cdc2) activation and is involved in resistance to taxol-induced
apoptosis. Mol Cell. 9:993–1004. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cepeda V, Fuertes MA, Castilla J, et al:
Biochemical mechanisms of cisplatin cytotoxicity. Anticancer Agents
Med Chem. 7:3–18. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Galluzzi L, Senovilla L, Vitale I, et al:
Molecular mechanisms of cisplatin resistance. Oncogene.
31:1869–1883. 2012. View Article : Google Scholar
|
|
14
|
Hiraishi Y, Wada T, Nakatani K, et al:
EGFR inhibitor enhances cisplatin sensitivity of oral squamous cell
carcinoma cell lines. Pathol Oncol Res. 14:39–43. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang XJ, Finkel T, Shen DW, et al: SIRT1
contributes in part to cisplatin resistance in cancer cells by
altering mitochondrial metabolism. Mol Cancer Res. 6:1499–1506.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shimanishi M, Ogi K, Sogabe Y, et al:
Silencing of GLUT-1 inhibits sensitization of oral cancer cells to
cisplatin during hypoxia. J Oral Pathol Med. 42:382–388. 2013.
View Article : Google Scholar
|
|
17
|
Semenza GL: Tumor metabolism: cancer cells
give and take lactate. J Clin Invest. 118:3835–3837.
2008.PubMed/NCBI
|
|
18
|
Zhou M, Zhao Y, Ding Y, et al: Warburg
effect in chemosensitivity: targeting lactate dehydrogenase-A
re-sensitizes taxol-resistant cancer cells to taxol. Mol Cancer.
9:332010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang GC, Liu SY, Lin MH, Kuo YY and Liu
YC: The synergistic cytotoxicity of cisplatin and taxol in killing
oral squamous cell carcinoma. Jpn J Clin Oncol. 34:499–504. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shin YK, Yoo BC, Hong YS, et al:
Upregulation of glycolytic enzymes in proteins secreted from human
colon cancer cells with 5-fluorouracil resistance. Electrophoresis.
30:2182–2192. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vaughn AE and Deshmukh M: Glucose
metabolism inhibits apoptosis in neurons and cancer cells by redox
inactivation of cytochrome c. Nat Cell Biol. 10:1477–1483. 2008.
View Article : Google Scholar : PubMed/NCBI
|