Introduction
Lung cancer is the leading cause of
cancer-associated mortality in the United States, with 157,000
cases of lung cancer-associated mortality in 2010 and 160,000
people in 2013, accounting for 26 and 28% of all female and male
cancer-associated deaths, respectively (1,2). Lung
cancer is classified into two main histological types: Non-small
cell lung cancer (NSCLC) and small cell lung cancer (SCLC),
accounting for 87% and 13% of all lung cancer cases, respectively
(3). The predominant histological
subtypes of NSCLC are adenocarcinoma (50–60%), squamous cell
carcinoma (30–35%) and large-cell carcinoma (5–10%) (4). At present despite improvements in the
diagnosis and treatment of lung cancer, factors such as
postoperative recurrence and metastatic infiltration mean the
prognosis of patients with lung cancer is still poor. However, the
precise mechanisms of cancer recurrence and metastasis remain
unclear.
The family of fibroblastic growth factors (FGF) has
23 identified members, which bind with 4 FGF receptor (FGFR)
ligands, consisting of an extracellular portion, a transmembrane
region and an intracellular domain. FGF family members are involved
in cell growth, differentiation, morphogenesis, tissue repair,
inflammation, angiogenesis, tumor growth and numerous developmental
processes including embryonic and skeletal development (5–9). As
such, the role of the FGF family has been widely studied during
tumor growth and metastasis and has been shown to increase the
proliferation, motility and invasiveness of a range of cell types
(10,11). FGF18 has been demonstrated to serve
an important role in skeletal growth and limb development,
potentially through the modulation of osteoblasts, chondrocytes,
and osteoclasts (12,13). Furthermore, FGF18 expression was
upregulated in colon cancer and ovarian cancer, and increased
expression of FGF18 mRNA and protein is associated with tumor
progression and poor overall survival in patients (14–16).
Mitogen activated protein kinase (MAPK) is an
intracellular signaling pathway, with physiological functions
including cell proliferation, apoptosis and differentiation. The
extracellular signal-regulated kinase (ERK) signaling pathway,
which is one of the MAPKs, plays the role of proliferation,
migration, differentiation (17).
FGF activated FGFRs then activate a number of downstream signaling
pathways, including ERK and p38, phospholipase Cγ, protein kinase C
and phosphatidylinositol 3-kinase (11,17,18).
The majority of these signaling pathways are involved in the growth
and metastasis of cancer cells. In the present study, we aimed to
investigate the effect of FGF18 and short interfering RNA
(siRNA)-FGF18 on the proliferation and migration of NSCLC cells, in
addition to the underlying mechanisms.
Materials and methods
Cell culture
The H460 human NSCLC line was obtained from Chemical
Biology Research Center (Wenzhou Medical University, Wenzhou,
China). The cells were cultured in Roswell Park Memorial Institute
(RPMI)-1640 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) containing 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.), and 1% antibiotic-antimycotic (Gibco) at 37°C
and 10% CO2.
Cell proliferation assay
A 3-(4,5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide (MTT) assay (Beyotime Institute of
Biotechnology, Haimen, China) was used to determine the
proliferation of H460 cells. The cells (4,000 cells/well) were
seeded and cultured for 24 h in 96-well plates, and serum-free
medium was added for 4 h, following which cells were stimulated
with (0, 5, 10, 50, 100 and 200 ng/ml rhFGF18 (Bioreactor, Wenzhou,
China) (19), 5 µmol ERK inhibitor
(FR180204, Sigma-Aldrich), 5 µmol p38 inhibitor (SB203580,
Sigma-Aldrich) and further incubated for 0, 24, 48 and 72 h.
Subsequently, 20 µl of 5 mg/ml MTT (BioSharp, Hefei, China) was
added to each well, the plate was incubated for 4 h and the
absorbance at 490 nm (SpectraMax M2, Molecular Devices, Sunnyvale,
CA, USA) was subsequently detected. In each group, three wells were
measured for cell proliferation; the data are shown as the mean ±
standard deviation (SD).
Cell cycle analysis
Cell cycle distribution was analyzed by propidium
iodide (PI; BD Bioscience, San Jose, CA, USA) staining and flow
cytometry. The H460 cells (20,000 cells/well) were seeded in 6-well
plates and exposed to (0, 5, 10 and 50 ng/ml) FGF18 for 48 h. The
cells were then harvested, fixed with 70% ice-cold ethanol, and
stored at −20°C until analysis. After fixation, the cells were
washed twice with cold phosphate-buffered saline (PBS) and
centrifuged, following which the supernatants were removed. The
pellet was resuspended and stained with PBS containing 50 mg/ml PI
and 100 mg/ml RNaseA for 20 min in the dark. The DNA content was
analyzed by flow cytometry using a FACSCalibur instrument and
CellQuest software (BD Bioscience). The cell cycle data were
analyzed using FlowJo 7.6 software, version (FlowJo, LLC, Ashland,
OR, USA) and the data are shown as the mean ± SD.
Would healing assay
H460 cells were seeded into 6-well plates and were
cultured at 37°C until they reached 100% confluence. The monolayers
were scratched with a pipette tip and cultured under normal
conditions or (0, 5, 10 and 50 ng/ml) FGF18 after the cell
fragments were removed by washing with PBS. Images (Olympus IX51,
magnification: ×40, Olympus Corporation, Tokyo, Japan) were
captured at 0, 24, 48 and 72 h, and the data are shown as the
migration distance between the two edges. The migration distance
was assessed using ToupView software. The data are shown as the
mean ± SD.
Cell transfection
Chemically synthesized FGF18 siRNA (Guangzhou
Ribobio Co., Ltd., Guangzhou, China) were used for transfection. To
make the transfection mixture, riboFECT™ CP buffer and siRNA were
first prepared in 1.5 ml micro-centrifuge tubes. Subsequently, they
were mixed with riboFECT CP reagent and allowed to incubate at room
temperature for 15 min. Cells in 6-well plates were washed with PBS
twice and the transfection mixture was added to the cells. The
plates were gently rocked following which the medium was added to
the cells. Subsequently, the H460 cells were transfected with FGF18
siRNA for 48 h.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from the cells using a TRIzol
plus kit (Takara Bio, Inc., Otsu, Japan) according to the
manufacturer's protocol. RNA was reverse transcribed using
PrimeScript™ RT Master Mix (Takara Bio, Inc.) according to the
manufacturer's instructions. The PCR amplifications were performed
for 40 cycles of 94°C for 30 sec, 60°C for 30 sec, and 72°C for 30
sec, using a Applied CFX96™ Real-Time PCR (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) with 1.0 µl of cDNA and SYBR Green
Real-time PCR Master Mix (Takara Bio, Inc.). Data were collected
and analyzed by Bio-Rad CFX Manager software. The expression level
of each sample was internally normalized against that of the
glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The relative
quantitative value was calculated using the 2−∆∆Ct
method (20). Each experiment was
performed in triplicate. The primers used in real-time PCR were as
follow: Matrix metalloproteinase 26 (MMP26), forward
5′-GGCCAGGTGGTATCTTAGGC-3′ and reverse
5′-AGCTGACCAGTGTTCATTCTTG-3′; FGF18 forward sequence
5′-GGACATGTGCAGGCTGGGCTA-3′ and reverse
5′-GTAGAATTCCGTCTCCTTGCCCTT-3′; and GAPDH forward
5′-ACAACAGCCTCAAGATCATCAG-3′ and reverse
5′-GGTCCACCACTGACACGTTG-3′. The primers were designed and
chemically synthesized in Genewiz, Inc. (South Plainfield, NJ,
USA).
Western blotting
Cell samples were digested in lysis buffer (Beyotime
Institute of Biotechnology), and the protein concentrations were
measured using the bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). Total protein (40 µg) was resolved by
12% odium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred onto 0.22 µm polyvinylidene fluoride membranes and
probed with the following primary antibodies, overnight at −4°C:
Anti-human FGF18 and anti-MMP26 (Abcam, Cambridge, MA, USA),
anti-phosphorylated (p)-ERK1/2, anti-ERK1/2, anti-p-p38, anti-p38
and anti-β-actin (Cell Signaling Technology, Inc., Danvers, MA,
USA). Following three washes, the membranes were incubated with
horseradish peroxidase conjugated anti-rabbit IgG secondary
antibodies (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 1
h at room temperature. The detection of specific proteins was
carried out using an enhanced chemiluminscence western blotting kit
(Santa Cruz Biotechnology, Inc.).
Statistical analysis
For each experiment, three independent replicates
were performed. All data are expressed as the mean ± SD.
Statistical evaluation was conducted using Student's test. The
intergroup differences were compared using one-way analysis of
variance followed by Dunnett's test. P<0.05 was considered to
indicate a statistically significant difference. *P<0.05,
**P<0.0l, and ***P<0.001 vs. control.
Results
FGF18 promotes the proliferation of
H460 cells
A cell proliferation assay was used to investigate
the effect of FGF18 on H460 cells. Cells were treated with 0, 5,
10, 50, 100 and 200 ng/ml of FGF18 for 0, 24, 48 or 72 h. As shown
in Fig. 1, stimulation with FGF18
significantly increased H460 cell proliferation in a time and
concentration-dependent manner. At concentrations of 5, 10 and 50
ng/ml, FGF18 had a remarkable effect on H460 cell proliferation.
When time reached 72 h, however, FGF18 was unable to affect the
growth as a consequence of the too long culture. The results
suggest that FGF18 has a role in promoting the proliferation of
lung cancer cells.
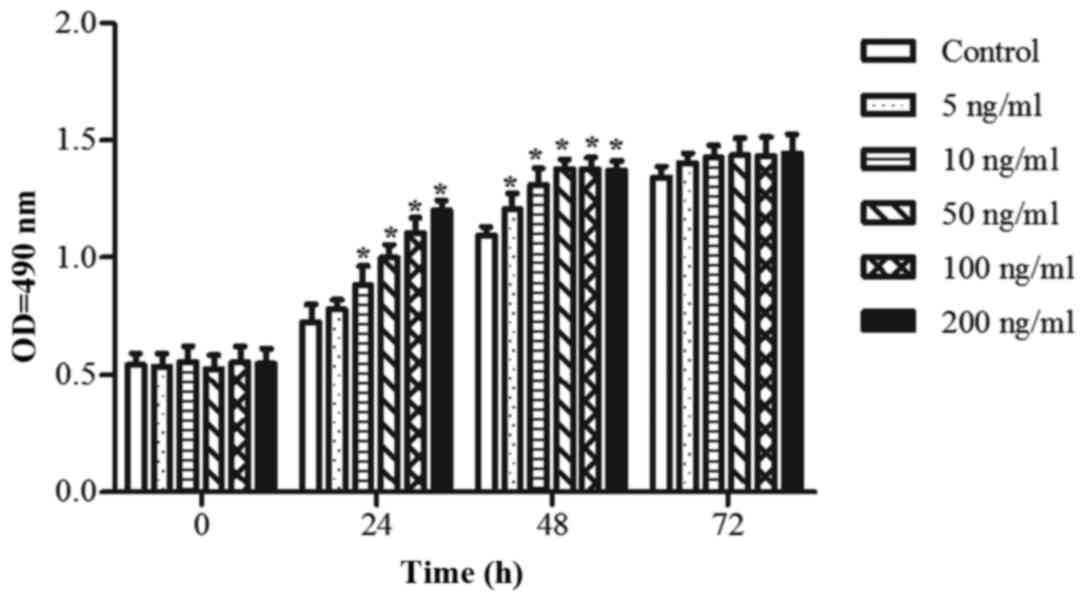 | Figure 1.Effect of FGF18 on the proliferation
of H460 cells. H460 cells were cultured in 96-well plates for 24 h
and treated with different concentrations (0, 5, 10, 50, 100 and
200 ng/ml) of FGF18 for 0, 24, 48 or 72 h. Subsequently, cell
proliferation was assessed by the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay.
Values are presented as the mean ± SD. (n=3). *P<0.05 vs.
control group. FGF18, fibroblast growth factor 18; OD, optical
density. |
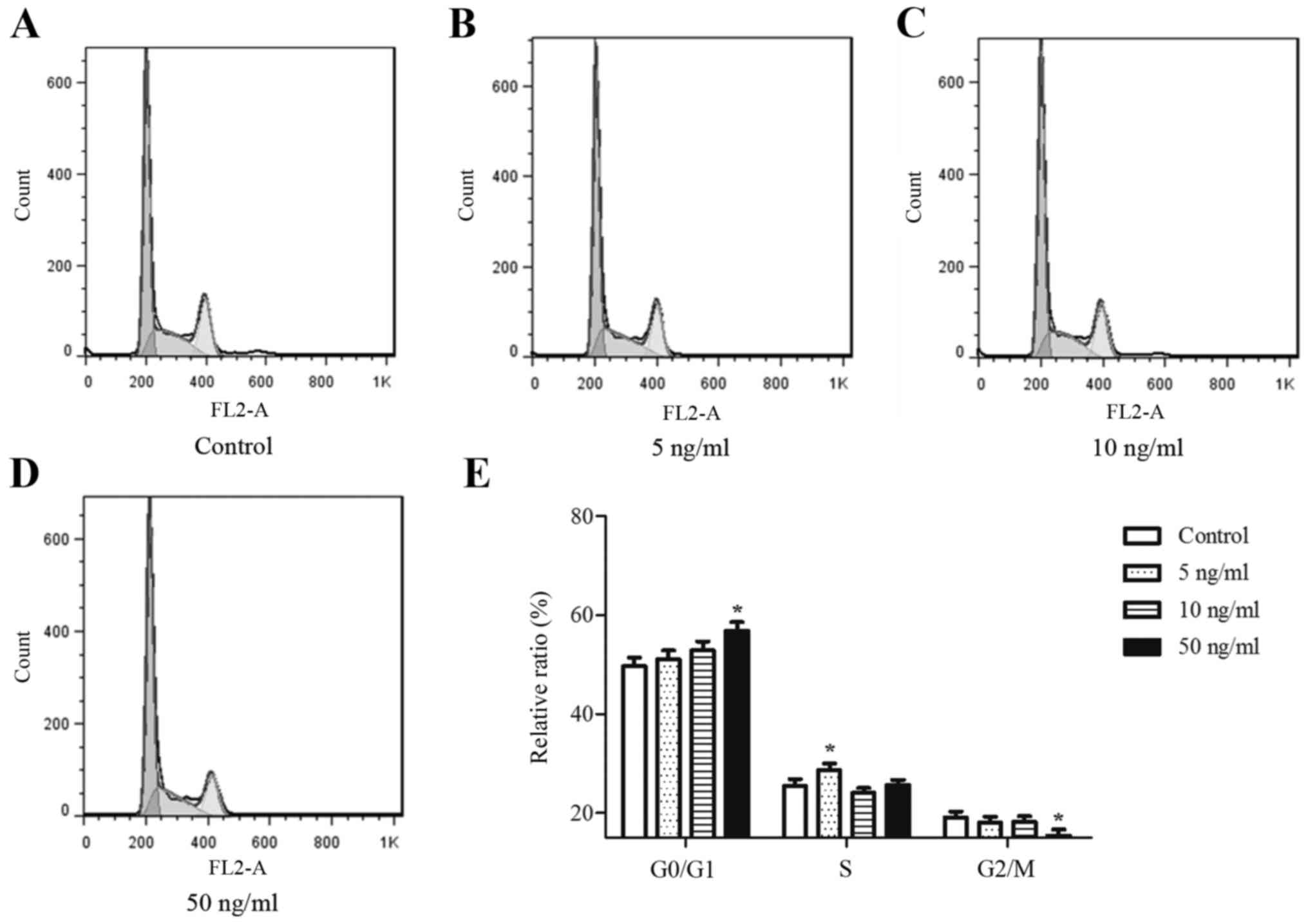
FGF18 promotes cell cycle
progression
Different concentrations of FGF18 promoted
proliferation of H460 cells, indicating that FGF18 may alter cell
cycle-related events in H460 cells. Therefore, the effects of FGF18
on cell cycle kinetics were investigated to understand how FGF18
may regulate the cell cycle. As shown in Fig. 2A-E, stimulation with FGF18 increased
the proportion of cells in the G0/G1 phase and reduced the
proportion of cells in the S or G2/M phases compared with untreated
H460 cells. These results suggest that the FGF18-mediated increase
in H460 cell proliferation occurred via increasing the proportion
of cells in G0/G1 phase and S phase.
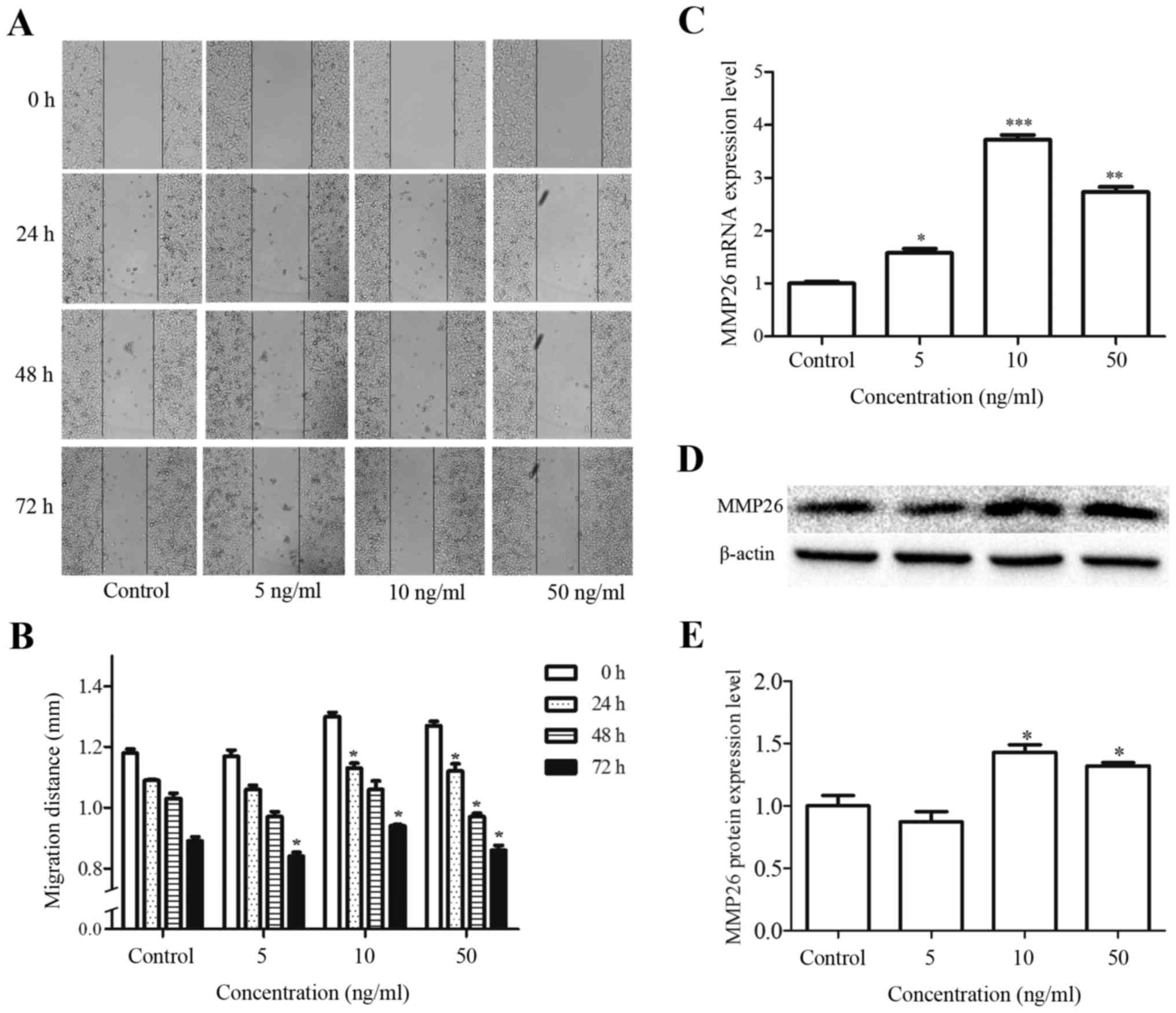
FGF18 promotes the migration of H460
cells and modulates the migration-related factor
A wound healing assay was used to investigate the
effects of FGF18 on cell migration. Following treatment with 5, 10
and 50 ng/ml FGF18 for 24, 48 or 72 h, the migration distance was
increased in H460 cells compared with the control untreated cells
(Fig. 3A and B). Additionally, the
effect of FGF18 on the expression of the migration-related factor
MMP26, was investigated in H460 cells. As shown in the results of
the western blot and RT-qPCR assays in Fig. 3C and E, FGF18 increased the mRNA and
protein expression levels of MMP26 in a dose-dependent manner,
particularly at 10 and 50 ng/ml concentrations. These results
suggest that FGF18 promotes the migration of H460 cells and affects
the migration-related factor MMP26.
ERK and p38 signaling are involved in
FGF18 mediated promotion of proliferation and migration in H460
cells
The above findings indicate that FGF18 significantly
promotes the proliferation and migration of H460 cells. Moreover,
the expression of MMP26 was promoted by FGF18 in H460 cells.
However, the underlying mechanisms responsible for the effect of
FGF18 are unclear. Hence, the effect of FGF18 on the signal
transduction of MAPKs was further assessed by western blot
analysis. H460 cells were treated with 5, 10 and 50 ng/ml FGF18 for
48 h, and the total protein lysates collected and subjected to
western blotting with p-ERK1/2, ERK1/2, p38 MAPK and p-p38 MAPK
antibodies. As shown in Fig. 4A and
B, FGF18 increased the levels of p-ERK1/2 and p-p38. To address
which of these pathways are regulating these effects, we used the
specific inhibitors, FR180204 (ERK inhibitor) and SB203580 (p38
inhibitor). Results showed that the effect of FGF18 stimulation on
proliferation and migration was inhibited by application of 5
µmol/l FR180204 and 5 µmol/l SB203580 (Fig. 4C-E). However, MMP26 protein did not
change by the inhibitors (Fig. 4F and
G). These data suggest that the effects of FGF18 in
proliferation and migration of H460 cells is potentially mediated
via modulations of the ERK1/2 pathway and p38 MAPK pathways.
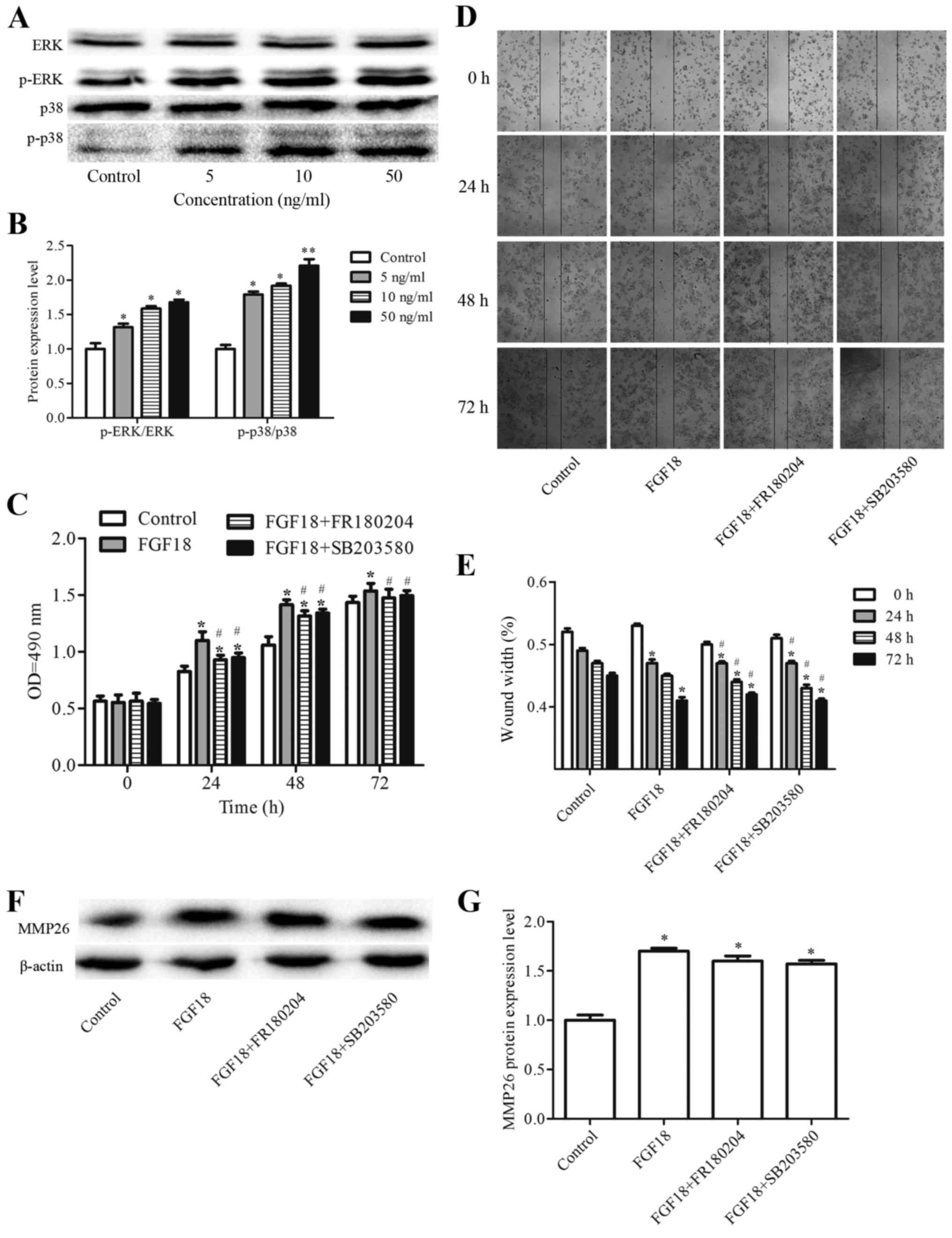 | Figure 4.Effect of FGF18 on ERK, p-ERK, p38 and
p-p38 expression levels in H460 cells. (A) H460 cells were treated
with various concentrations of FGF18 (0, 5, 10 or 50 ng/ml) for 48
h, then cells were collected and subjected to western blot
analysis. (B) Quantification of the protein expression levels. (C)
H460 cells were cultured on 96-well plates for 24 h and treated
with FGF18 (50 ng/ml), FGF18+FR180204 (ERK inhibitor, 5 µmol),
FGF18+SB203580 (p38 inhibitor, 5 µmol) for 0, 24, 48 or 72 h.
Subsequently, cell proliferation was assessed by
3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide
assay. (D and E) Effects of FGF18, FGF18+FR180204, and
FGF18+SB203580 on the migration of H460 cells. (F and G) Effect of
FGF18, FGF18+FR180204, and FGF18+SB203580 on MMP26 expression
levels in H460 cells. Values are presented as the mean ± SD (n=3).
*P<0.05, **P<0.01 vs. control. #P<0.05 vs.
FGF18. FGF18, fibroblast growth factor 18; ERK, extracellular
signal-regulated kinase; p-ERK, phosphorylated ERK; MMP, matrix
metalloproteinase. |
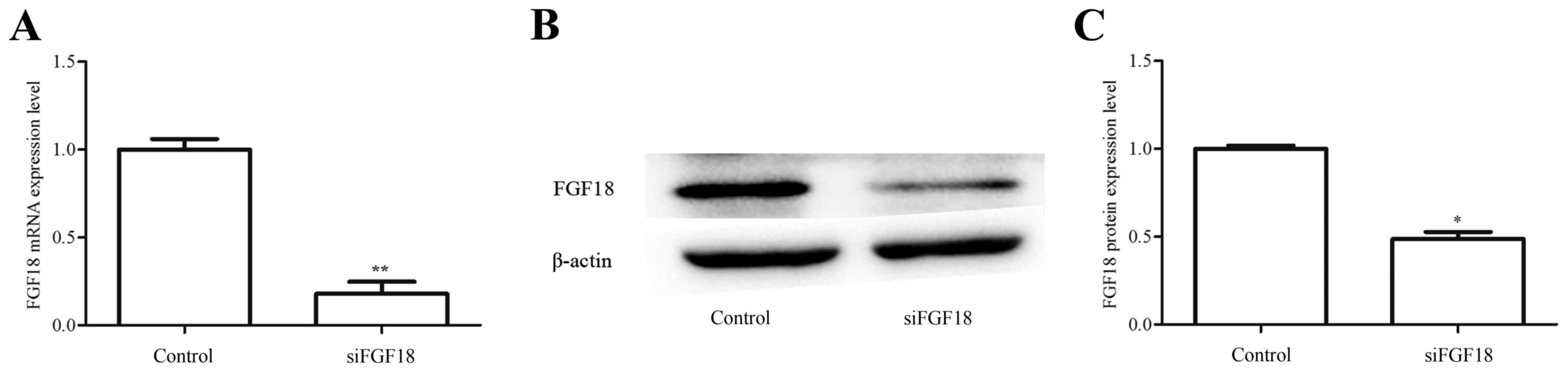
FGF18 expression in H460 cells is
downregulated by FGF18-siRNA
The RT-qPCR results indicated that the mRNA
expression of FGF18 in H460 cells transfected with siFGF18 was
reduced compared with control group (Fig. 5A). Additionally, FGF18 protein
expression was examined using western blot analyses (Fig. 5B and C), with the results consistent
with the mRNA data. These results demonstrate the successful
knockdown of FGF18 using siRNA.
FGF18 siRNA inhibits cell
proliferation and migration in the ERK and p38 signaling pathways
in H460 cells
The proliferation and migration of cancer cells are
key steps in the progression of cancer. Following cell
transfection, the effect of FGF18 siRNA on cell proliferation was
evaluated in H460. The OD values were significantly reduced in the
siFGF18 group compared with the control group (Fig. 6A), indicating that FGF18 siRNA
inhibited cell proliferation in H460 cells. However, as shown in
Fig. 6B and C, the siFGF18 group
did not exhibit alterations in the cell cycle of H460 cells. In
addition, the migration distance of siFGF18-transfected H460 cells
was reduced compared with the control groups (Fig. 6D and E). Expression of MMP26 gene
and protein was additionally significantly reduced following
siFGF18, with a 57% gene inhibition rate and 62% protein inhibition
rate, respectively (Fig. 6F-H). The
levels of p-ERK and p-p38 protein were significantly lower in the
siFGF18 group compared with the cells in the control group
(Fig. 6I and J). Taken together,
these results suggest that FGF18 siRNA is able to repress cell
proliferation and migration, and this effect is potentially
mediated through the ERK and pP38 signaling pathways in H460
cells.
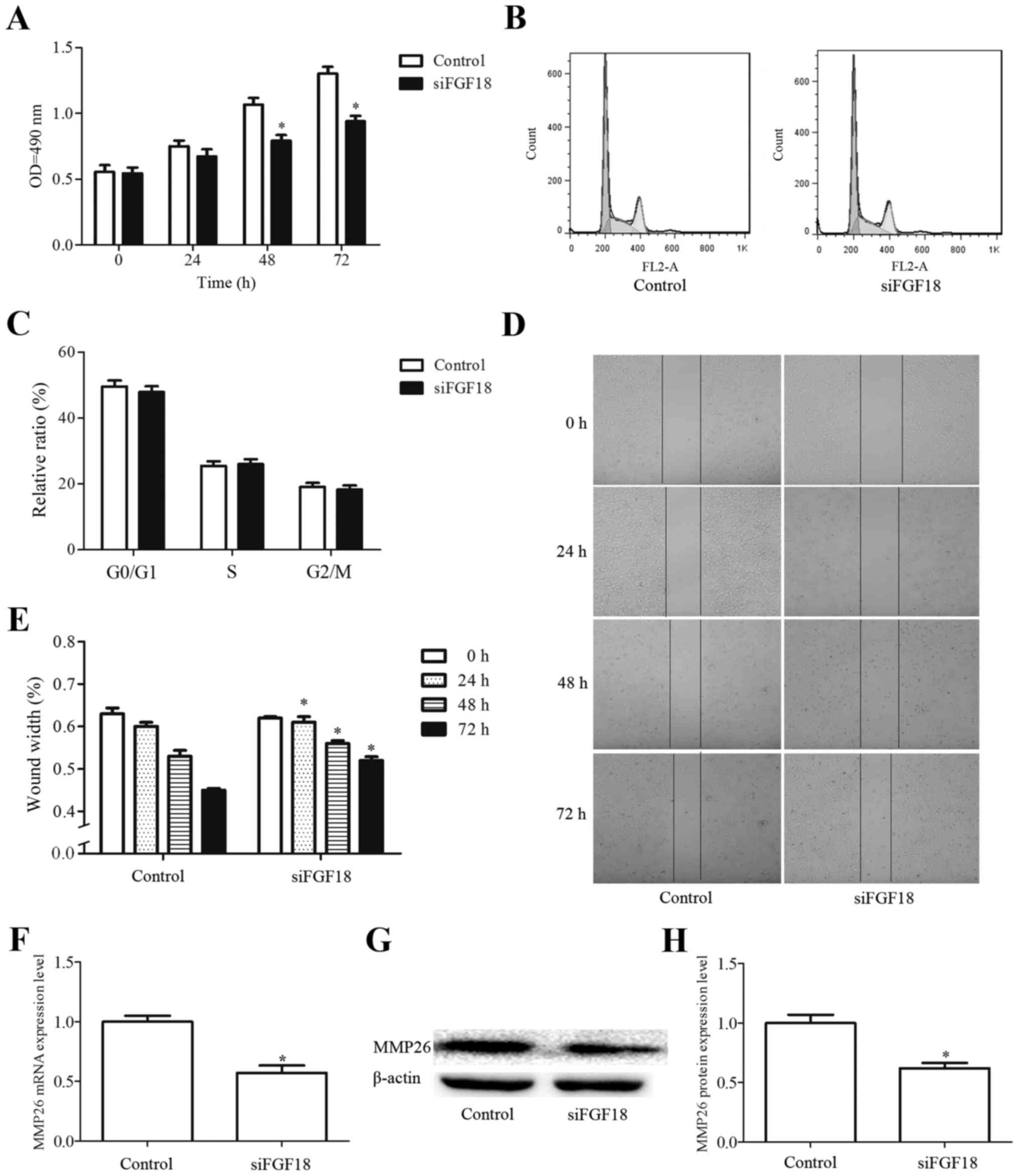 | Figure 6.Effect of siFGF18 on H460 cells. (A)
H460 cells were cultured on 96-well plates for 24 h and treated
with siFGF18 for 0, 24, 48 or 72 h. Subsequently, cell
proliferation was assessed by 3-(4,5-dimethylthiazol-2-yl)-2,
5-diphenyltetrazolium bromide assay. (B and C) Effect of siFGF18 on
the cell cycle of H460 cells. (D and E) Effects of siFGF18 on the
migration of H460 cells. (F-H) RNA and protein levels of MMP26 in
H460 cell. MMP, matrix metalloproteinase; OD, optical density.
*P<0.05 vs. control. Effect of siFGF18 on H460 cells. (I) Effect
of siFGF18 on ERK, p-ERK, p38 and p-p38 expression levels in H460
cells. (J) Quantification of protein expression levels. Values are
presented as the mean ± SD (n=3). *P<0.05, **P<0.01 vs.
control. siFGF18, short interfering RNA against fibroblast growth
factor 18; ERK, extracellular signal-regulated kinase; p-ERK,
phosphorylated ERK. |
Discussion
In NSCLC, the FGF and FGFR family has been
demonstrated to be associated with its progression. FGF1 expression
has been identified in the cytoplasm, or both cytoplasm and nucleus
of NSCLC cells, and high expression levels of FGF1 in cancer cells
were significantly correlated with larger primary tumor size and
vascular invasion (21). FGF9-FGFR3
signaling has a complex role in the initiation, growth and
propagation of lung cancer (22).
Nevertheless, the role of FGF18 has not been previously studied in
the context of lung cancer. Previous studies have indicated that
elevated expression of FGF18 promotes the growth of colon cancer
cells in culture (23), and
demonstrated the pronounced oncogenic effect of FGF18 on ovarian
tumor growth and metastasis (16).
These observations are in agreement with our conclusion that FGF18
promoted the proliferation and migration of H460 cells. The present
study, to the best of our knowledge, is the first report of the
effects of FGF18 on NSCLC cells.
A number of studies have shown that MAPKs (JNK1/2,
ERK1/2, and p38) are involved in the growth, migration, and
alterations in MMPs activity (24,25) in
various cancers including lung, prostate, colorectal and ovarian
cancer (26–30). Furthermore, the ERK signaling
pathway in addition to the p38 signaling pathway may regulate
numerous factors that are associated with cancer progression and
poor prognosis in NSCLC (31–33).
The MMP family comprises 24 zinc-dependent endopeptidases, and
serves important roles in tumor metastasis and MMP26 is associated
with the invasion and metastasis of non-small cell lung cancer
(34–36). It has been reported that the
expression of MMPs is regulated by MAPK pathways (37,38).
To confirm whether ERK and p38 signaling pathway is involved in
this situation, we applied specific inhibitors for ERK and p38,
respectively, to the FGF18-stimulated H460 cells. We found that the
inhibition of ERK and p38 decreased the proliferation and migration
in response to FGF18 stimulation, while the activation of MMP26 did
not change, suggesting that the activation of ERK and p38 signaling
may cause an increase in proliferation and migration of H460 cells
without MMP26 protein. However, using either ERK inhibitor or p38
inhibitor could not completely inhibit the H460 proliferation and
migration, which may be due to the presence and activation of
another signal transduction cascade. The present study suggests
that the stimulation with FGF18 in NSCLC cells activated the ERK
and p38 signaling pathways, and additionally upregulated the level
of MMP26. This suggests that the underlying molecular mechanism may
involve activated ERK and p38 signaling pathways in NSCLC cells
inducing proliferative and migratory signals.
The present study additionally demonstrated that the
transfection of cancer cells with siRNA targeted against the FGF18
gene can effectively reduce FGF18 gene expression and suppress the
effects on proliferation, and migration in H460 cells. However, the
effect of siFGF18 on proliferation was not accompanied by
alterations in the cell cycle. Therefore further studies are
required to explore the mechanisms of FGF18 on proliferation.
Western blot analysis was used to investigate the expression of
ERK, p38 and MMP26 in human H460 cells following siRNA-FGF18
treatment for 48 h, which demonstrated marked inhibition of the ERK
and p38 signaling pathways. These results imply that the use of
siRNA holds potential in treating lung cancer cells, and that the
FGF18 gene may be a potential therapeutic target in NSCLC.
In conclusion, the present study suggests that FGF18
serves a key role in the proliferation and migration of NSCLC cells
by regulating the ERK, p38 signaling pathways and MMP26 protein
expression levels, indicating that FGF18 may represent a potential
molecular drug target in NSCLC. Nevertheless, further studies using
NSCLC cell lines and investigating the in vivo physiological
role of FGF18 are required.
Acknowledgements
This work was supported by grants of the National
Key New Drug Foundation of China (no. 2013ZX09103-003-005), Wenzhou
Scientific Innovation Grant to Dr Chao Jiang, and Scientific
Research Fund of Wenzhou Medical University, project number:
QTJ16003.
Glossary
Abbreviations
Abbreviations:
|
FGF18
|
fibroblast growth factor 18
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
MMP26
|
matrix metalloproteinase 26
|
|
NSCLC
|
non-small cell lung cancer
|
|
SCLC
|
small cell lung cancer
|
|
FGF
|
fibroblastic growth factors
|
|
FGFR
|
fibroblastic growth factor
receptor
|
|
MAPK
|
mitogen activated protein kinase
|
|
siRNA
|
short interfering RNA
|
|
PI
|
propidium iodide
|
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Travis WD, Brambilla E, Nicholson AG,
Yatabe Y, Austin JH, Beasley MB, Chirieac LR, Dacic S, Duhig E,
Flieder DB, et al: The 2015 World Health Organization
Classification of Lung Tumors: Impact of genetic, clinical and
radiologic advances since the 2004 classification. J Thorac Oncol.
10:1243–1260. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ettinger DS, Akerley W, Bepler G, Blum MG,
Chang A, Cheney RT, Chirieac LR, D'Amico TA, Demmy TL, Ganti AK, et
al: Non-small cell lung cancer. J Natl Compr Canc Netw. 8:740–801.
2010.PubMed/NCBI
|
|
5
|
Antoine M, Wirz W, Tag CG, Mavituna M,
Emans N, Korff T, Stoldt V, Gressner AM and Kiefer P: Expression
pattern of fibroblast growth factors (FGFs), their receptors and
antagonists in primary endothelial cells and vascular smooth muscle
cells. Growth Factors. 23:87–95. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hu MC, Qiu WR, Wang YP, Hill D, Ring BD,
Scully S, Bolon B, DeRose M, Luethy R, Simonet WS, et al: FGF-18, a
novel member of the fibroblast growth factor family, stimulates
hepatic and intestinal proliferation. Mol Cell Biol. 18:6063–6074.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Canalis E, McCarthy TL and Centrella M:
Growth factors and cytokines in bone cell metabolism. Annu Rev Med.
42:17–24. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu Z, Xu J, Colvin JS and Ornitz DM:
Coordination of chondrogenesis and osteogenesis by fibroblast
growth factor 18. Genes Dev. 16:859–869. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dailey L, Ambrosetti D, Mansukhani A and
Basilico C: Mechanisms underlying differential responses to FGF
signaling. Cytokine Growth Factor Rev. 16:233–247. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Turner N and Grose R: Fibroblast growth
factor signalling: From development to cancer. Nat Rev Cancer.
10:116–129. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Beenken A and Mohammadi M: The FGF family:
Biology, pathophysiology and therapy. Nat Rev Drug Discov.
8:235–253. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Marie PJ: Fibroblast growth factor
signaling controlling osteoblast differentiation. Gene. 316:23–32.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Moore EE, Bendele AM, Thompson DL, Littau
A, Waggie KS, Reardon B and Ellsworth JL: Fibroblast growth
factor-18 stimulates chondrogenesis and cartilage repair in a rat
model of injury-induced osteoarthritis. Osteoarthritis Cartilage.
13:623–631. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sonvilla G, Allerstorfer S, Stättner S,
Karner J, Klimpfinger M, Fischer H, Grasl-Kraupp B, Holzmann K,
Berger W, Wrba F, et al: FGF18 in colorectal tumour cells:
Autocrine and paracrine effects. Carcinogenesis. 29:15–24. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Koneczny I, Schulenburg A, Hudec X,
Knöfler M, Holzmann K, Piazza G, Reynolds R, Valent P and Marian B:
Autocrine fibroblast growth factor 18 signaling mediates
Wnt-dependent stimulation of CD44-positive human colorectal adenoma
cells. Mol Carcinog. 54:789–799. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei W, Mok SC, Oliva E, Kim SH, Mohapatra
G and Birrer MJ: FGF18 as a prognostic and therapeutic biomarker in
ovarian cancer. J Clin Invest. 123:4435–4448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rubinfeld H and Seger R: The ERK cascade:
A prototype of MAPK signaling. Mol Biotechnol. 31:151–174. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Krejci P, Prochazkova J, Bryja V, Kozubik
A and Wilcox WR: Molecular pathology of the fibroblast growth
factor family. Hum Mutat. 30:1245–1255. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Song L, Huang Z, Chen Y, Li H, Jiang C and
Li X: High-efficiency production of bioactive recombinant human
fibroblast growth factor 18 in Escherichia coli and its effects on
hair follicle growth. Appl Microbiol Biotechnol. 98:695–704. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) methods. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Wei Z, Li H, Dang Q, Zhang Z, Wang
L, Gao W, Zhang P, Yang D, Liu J, et al: Clinicopathological
significance of fibroblast growth factor 1 in non-small cell lung
cancer. Hum Pathol. 46:1821–1828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Arai D, Hegab AE, Soejima K, Kuroda A,
Ishioka K, Yasuda H, Naoki K, Kagawa S, Hamamoto J, Yin Y, et al:
Characterization of the cell of origin and propagation potential of
the fibroblast growth factor 9-induced mouse model of lung
adenocarcinoma. J Pathol. 235:593–605. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shimokawa T, Furukawa Y, Sakai M, Li M,
Miwa N, Lin YM and Nakamura Y: Involvement of the FGF18 gene in
colorectal carcinogenesis, as a novel downstream target of the
beta-catenin/T-cell factor complex. Cancer Res. 63:6116–6120.
2003.PubMed/NCBI
|
|
24
|
Tang SW, Yang TC, Lin WC, Chang WH, Wang
CC, Lai MK and Lin JY: Nicotinamide N-methyltransferase induces
cellular invasion through activating matrix metalloproteinase-2
expression in clear cell renal cell carcinoma cells.
Carcinogenesis. 32:138–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McCubrey JA, Steelman LS, Chappell WH,
Abrams SL, Wong EW, Chang F, Lehmann B, Terrian DM, Milella M,
Tafuri A, et al: Roles of the Raf/MEK/ERK pathway in cell growth,
malignant transformation and drug resistance. Biochim Biophys Acta.
1773:1263–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun L, Zhang Q, Li Y, Tang N and Qiu X:
CCL21/CCR7 up-regulate vascular endothelial growth factor-D
expression via ERK pathway in human non-small cell lung cancer
cells. Int J Clin Exp Pathol. 8:15729–15738. 2015.PubMed/NCBI
|
|
27
|
Han Y, Luo Y, Wang Y, Chen Y, Li M and
Jiang Y: Hepatocyte growth factor increases the invasive potential
of PC-3 human prostate cancer cells via an ERK/MAPK and Zeb-1
signaling pathway. Oncol Lett. 11:753–759. 2016.PubMed/NCBI
|
|
28
|
Li L, Duan T, Wang X, Zhang RH, Zhang M,
Wang S, Wang F, Wu Y, Huang H and Kang T: KCTD12 Regulates
colorectal cancer cell stemness through the ERK pathway. Sci Rep.
6:204602016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xia ZX, Li ZX, Zhang M, Sun LM, Zhang QF
and Qiu XS: CARMA3 regulates the invasion, migration, and apoptosis
of non-small cell lung cancer cells by activating NF-κB and
suppressing the P38 MAPK signaling pathway. Exp Mol Pathol.
100:353–360. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bai RX, Wang WP, Zhao PW and Li CB:
Ghrelin attenuates the growth of HO-8910 ovarian cancer cells
through the ERK pathway. Braz J Med Biol Res. 49(pii):
S0100-879X2016000300602. 2016.
|
|
31
|
Kim JH, Cho EB, Lee J, Jung O, Ryu BJ, Kim
SH, Cho JY, Ryou C and Lee SY: Corrigendum to <Emetine inhibits
migration and invasion of human non-small-cell lung cancer cells
via regulation of ERK and p38 signaling pathways>. Chem Biol
Interact. Nov 2–2015.(Epub ahead of print). doi:
10.1016/j.cbi.2015.10.023.
|
|
32
|
Zhang Y, Zhao J, Qiu L, Zhang P, Li J,
Yang D, Wei X, Han Y, Nie S and Sun Y: Co-expression of ILT4/HLA-G
in human non-small cell lung cancer correlates with poor prognosis
and ILT4-HLA-G interaction activates ERK signaling. Tumour Biol.
37:11187–11198. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang C, Shi J, Mao SY, Xu YS, Zhang D,
Feng LY, Zhang B, Yan YY, Wang SC, Pan JP, et al: Role of p38 MAPK
in enhanced human cancer cells killing by the combination of
aspirin and ABT-737. J Cell Mol Med. 19:408–417. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Zhao H, Wang Y, Lin Y, Tan Y,
Fang X and Zheng L: Non-small cell lung cancer invasion and
metastasis promoted by MMP-26. Mol Med Rep. 4:1201–1209.
2011.PubMed/NCBI
|
|
35
|
Ming SH, Sun TY, Xiao W and Xu XM: Matrix
metalloproteinases-2, −9 and tissue inhibitor of
metallo-proteinase-1 in lung cancer invasion and metastasis. Chin
Med J (Engl). 118:69–72. 2005.PubMed/NCBI
|
|
36
|
Zhao D, Lu Y, Yang C, Zhou X and Xu Z:
Activation of FGF receptor signaling promotes invasion of
non-small-cell lung cancer. Tumour Biol. 36:3637–3642. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu X, Yang L, Zheng Z, Li Z, Shi J, Li Y,
Han S, Gao J, Tang C, Su L, et al: Src promotes cutaneous wound
healing by regulating MMP-2 through the ERK pathway. Int J Mol Med.
37:639–648. 2016.PubMed/NCBI
|
|
38
|
Xia Y, Lian S, Khoi PN, Yoon HJ, Joo YE,
Chay KO, Kim KK and Do Jung Y: Chrysin inhibits tumor
promoter-induced MMP-9 expression by blocking AP-1 via suppression
of ERK and JNK pathways in gastric cancer cells. PLoS One.
10:e01240072015. View Article : Google Scholar : PubMed/NCBI
|