Introduction
Pancreatic cancer (PC) has one of poorest prognosis
rates of all tumors and no successful cure has yet been identified.
Despite impressive improvements in surgical and chemotherapeutic
approaches (1), the 5-year
survival rate remains <5% (2).
Thus, it is urgently necessary to search for novel highly specific
and sensitive treatments in order to improve the prognosis of
patients with PC.
Ezrin-radixin-moesin (ERM)-binding phosphoprotein 50
(EBP50), also termed Na+/H+ exchanger
regulatory factor 1 (NHERF1), belongs to the NHERF protein family.
This family contains important molecular scaffold proteins that
coordinate a diverse range of regulatory processes for ion
transport and second messenger cascades (3–5).
EBP50 comprises two tandem PSD-95/discs large/ZO-1 (PDZ) domains
and a carboxyl (C)-terminal EB region (3,6,7),
which is a 50-kD microvillar scaffolding protein. EBP50 binds with
the majority of protein receptors, including the parathyroid
hormone type 1, β2-adrenergic, κ-opioid, parathyroid hormone type 1
and G protein-coupled receptors, through the first PDZ domain
(8–10). Growth factor tyrosine kinase
receptors, including the platelet-derived and epidermal growth
factor receptors, are also able to interact with EBP50 (11–13).
The second PDZ domain is able to bind with only a few proteins,
such as β-catenin, sodium-hydrogen exchanger 3 (NHE3) and
yes-associated protein 65 (Yap 65) (14,15).
A previous study revealed that the downregulation of
EBP50 promoted PC cell proliferation, increased the colony-forming
ability of cells and accelerated G1-to-S phase progression
(16). However, to the best of our
knowledge, no study on the effect of upregulation of EBP50 on PC
cell lines has been published. To examine whether EBP50
overexpression was efficient at treating PC, the present study used
the Pbk-CMV-HA-EBP50 plasmid to upregulate EBP50 expression in PC
cells and investigate its effect on SW1990 and PANC-1 PC cells.
Furthermore, the antitumor efficacy of Pbk-CMV-HA-EBP50 was
detected in vivo, using two mouse tumor xenograft models,
individually originating from PANC-1 and SW1990 cells.
Materials and methods
Cell culture and transfection
Human pancreatic cancer cell lines PANC-1 and SW1990
were purchased from the Cell Bank of the Shanghai Institutes for
Biological Sciences (Shanghai, China). They were maintained in a
laboratory and cultured in HyClone™ RPMI-1640 medium (Gibco-BRL,
Grand Island, NY, USA) containing 10% fetal calf serum, 100 U/ml
penicillin and 100 μg/ml streptomycin at 37°C and 5% CO2
in a Forma™ incubator ( Hera Cell, Thermo Scientific, Waltham, MA,
USA). The Pbk-CMV-HA-EBP50 plasmid was kindly provided by Dr. Randy
Hall from Emory University (Atlanta, GA, USA) and Pbk-CMV-HA was
obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Cells were seeded on a six-well plate at 2×105/ml and
transfected with equal amounts of Pbk-CMV-HA-EBP50 to generate
EBP50-PANC-1 and EBP50-SW1990 cells, or Pbk-CMV-HA to create
HA-PANC-1 and HA-SW1990 cells. The transfection was conducted using
Lipofectamine®2000 (Invitrogen Life Technologies,
Carlsbad, CA, USA) in accordance with the manufacturers’
instructions. The cells were collected after 24 or 48 h.
Transfection with Pbk-CMV-HA was used as the negative control
(17). The stably transfected cell
lines were constructed using a previously described method
(16). Cells were trypsinized and
reseeded into a 12-well plate. Stably transfected cell clones were
selected using G418 solution (Gibco-BRL). Once the single-cell
clones were isolated, the clones were expanded. Western blot
analysis was carried out to determine the transfection efficiency
of each cell clone.
Western blot analysis
The cultured cells were collected and lysed using
radioimmunoprecipitation (RIPA, Thermo Scientific) lysis buffer.
The protein concentration was measured using a bicinchoninic acid
(BCA) Protein Assay kit (Pierce Biotechnology, Inc., Rockford, IL,
USA). Equal amounts of proteins were separated on sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels and
electro-transferred to nitrocellulose membranes at 4°C for 2 h. The
primary antibody was rabbit polyclonal anti-human EBP50 (Novus,
Saint Charles, MO, USA), which was added to the proteins at 1:800
dilution and incubated overnight at 4°C. Another antibody was
anti-Bcl-2 (1:1,000; Abcam, Cambridge, MA, USA), which was also
incubated overnight at 4°C. The proteins were then incubated with
horseradish peroxidase-conjugated anti-rabbit antibodies (Sigma,
St. Louis, MO, USA) for 1 h at 4°C. The proteins were subsequently
detected by enhanced chemiluminescence (ECL; Amersham Pharmacia
Biotech, Piscataway, NJ, USA) according to the manufacturer’s
instructions and quantified by densitometry using UN-SCAN-IT
software (Silk Scientific Corp., Orem, UT, USA) (18) with β-actin used as an internal
control.
Cell proliferation assay
A cell counting kit-8 (CCK-8; Dojindo, Kumamoto,
Japan) colorimetric assay was used to determine cell proliferation
and viability. Cells were washed twice with ice-cold
phosphate-buffered saline (PBS), harvested by trypsinization,
counted and plated at a final density of 5×103
cells/well in a 96-well plate. Cell viability was assessed once
daily for seven consecutive days using the CCK-8. The absorbance
was detected in a microplate reader model 450 (Bio-Rad
Laboratories, Hercules, CA, USA) at 450 nm wavelength. Flow
cytometry (FACSCalibur, Becton-Dickinson, Franklin Lanes, NJ, USA)
was used to analyze the cell cycle of the stained cells (data not
shown) and the cell population in each phase was calculated by
computer model fitting (Verity Software House, Topsham, ME,
USA).
Cell cycle analysis by Annexin
V-fluorescein isothiocyanate (FITC) staining
The treated cells were plated at a density of
1×106 cells/ml in six-well plates. The culture medium
was substituted with fresh medium. The cells were collected, washed
with cold PBS and suspended in 1× binding buffer at
105–106 cells/ml. Annexin V and propidium
iodide (PI) were added to the prepared cell suspension. The cell
suspension was kept on ice and incubated for 10 min in the dark. It
was subsequently diluted with 1× binding buffer. Flow cytometry was
used to analyze the cell cycle of the stained cells (19).
Hoechst 33258 staining
Stably transfected cells were fixed, stained with
Hoechst 33258 and observed using a fluorescence microscope (model
IX71, Olympus, Tokyo, Japan). The stained cells were identified as
apoptotic if they were highly condensed with brightly stained
nuclei, or non-apoptotic if they were stained pale blue (20).
Tumor xenografts
A total of 30 female BALB/c nude mice aged 6–8 weeks
were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing,
China) and were kept under specific pathogen-free conditions. The
mice were randomly divided into six groups (n=5) and subcutaneously
injected with cells of one of the following types: i) PANC-1; ii)
HA-PANC-1; iii) EBP50-PANC-1; iv) SW1990; v) HA-SW1990; and vi)
EBP50-SW1990. After 36 days, the mice were sacrificed and the tumor
tissues were analyzed by hematoxylin and eosin (H&E) staining,
immunohistochemical analysis and a terminal deoxynucleotidyl
transferase dUTP nick end labeling (TUNEL) assay (18,21).
The tissues were harvested and fixed in 4%
paraformaldehyde at 4°C. Samples were embedded in paraffin,
sectioned at 5-μm thickness with a microtome and stained with
H&E staining for light microscopic examination (18).
Immunohistochemistry staining was performed on
paraffin sections with anti-EBP50 polyclonal rabbit antibody (1:800
dilution, Novus, Saint Charles, MO, USA) according to the
manufacturer’s protocol and included the following brief steps:
Deparaffinization followed by antigen retrieval, initial blocking
followed by incubation with the primary antibody, and further
washing and blocking followed by incubation with the biotinylated
secondary antibody. Finally, the sections were washed, blocked
again, washed, mounted and observed. Negative controls were
achieved by substituting the primary antibody with an
isotype-matched irrelevant antibody (21).
The TUNEL assay was performed on the
paraffin-embedded sections using a standard TUNEL assay kit (Roche,
Mannheim, Germany). Using the same method as the
immunohistochemical analysis, the sections were treated (n=5, in
each group) in order to prepare them at 24 h after reperfusion. The
staining was performed according to the manufacturer’s
instructions. The nucleus of the cells that was stained brown was
an indicator of the presence of TUNEL-positive cells.
The animal experiments were approved by the
Institutional Animal Care and Use Committee of Wuhan University
(Wuhan, Hubei).
Statistical analyses
Data are presented as mean ± standard deviation.
Analysis of variance (ANOVA) was used to perform statistical
comparisons of in vitro and in vivo data. P<0.05
was considered to indicate a statistically significant
difference.
Results
Overexpression of EBP50 inhibits the
proliferation of human PC cells
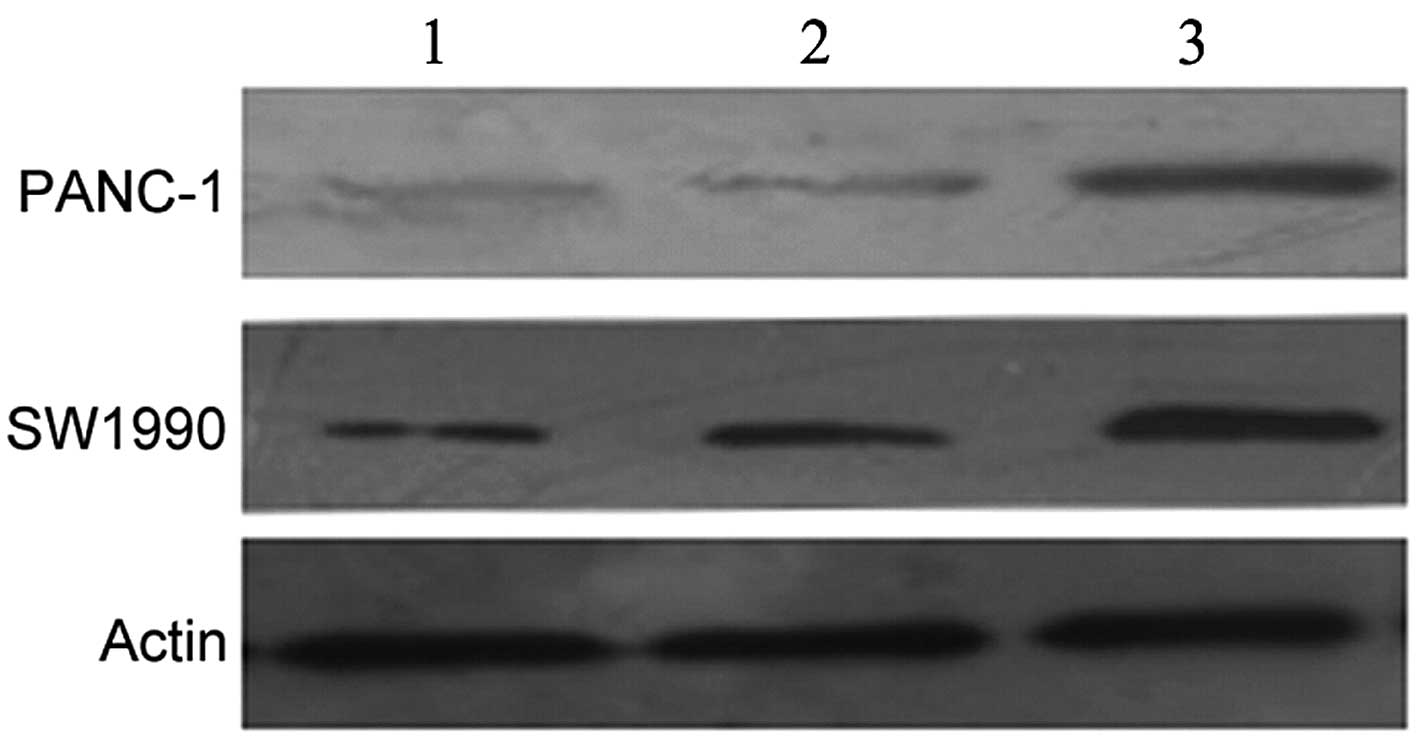
The Pbk-CMV-HA-EBP50 plasmid was stably transfected
into PANC-1 and SW1990 cells. Following selection with G418
solution, western blot analysis was carried out to identify the
stable clones. The data revealed that the expression levels of the
EBP50 protein were significantly higher in EBP50-PANC-1 and
EBP50-SW1990 cells than those in the PANC-1/HA-PANC-1 and
SW1990/HA-SW1990 cells, respectively (Fig. 1), thus indicating that the EBP50
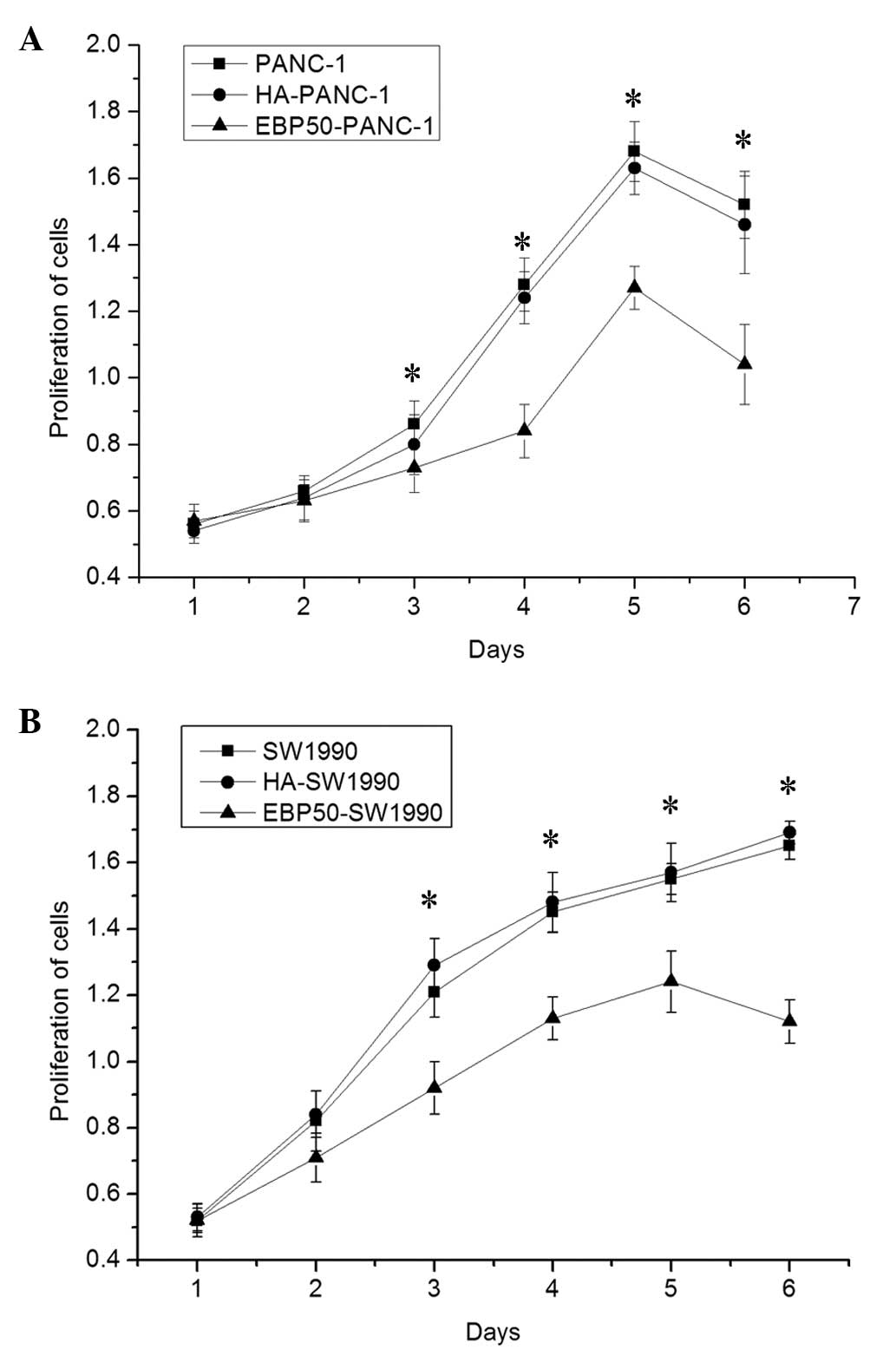
protein was upregulated in the two PC cell lines. Subsequently, to
test the effect of EBP50 overexpression on the suppression of PC
cell proliferation, a CCK-8 assay was performed to detect the cell
viability of two treated human PC cell lines. There was significant
suppression of PC cell proliferation in the EBP5-PANC-1 and
EBP5-SW1990 cells; however, there was no significant difference in
proliferation between the untreated and negative control groups in
the two PC cell lines (Fig.
2).
EBP50 overexpression induces G1-to-S
phase cell cycle arrest and cell apoptosis in human PC cells
To examine the effect of EBP50 overexpression on the
cell cycle, flow cytometry was carried out to detect the cell cycle
stages of two PC cell lines following stable transfection. All
lines transfected with Pbk-CMV-HA exhibited a pattern of phase
distribution similar to that of the corresponding untransfected
cell lines. However, the phase distribution of the two PC cell
lines transfected with Pbk-CMV-HA-EBP50 was markedly different from
that of the control and untreated groups. There was a significant
increase in the percentage of G0/G1 phase cells and a marked
reduction in the percentage of S phase cells in the
Pbk-CMV-HA-EBP50 groups compared with the corresponding control
groups (Fig. 3). Thus, EBP50
overexpression may promote cell apoptosis through arresting cell
cycle progression of the two PC cell lines between the G1 and S
phases.
To further investigate the role of EBP50
overexpression in the regulation of cell apoptosis, a Hoechst 33258
stain was used to detect the levels of cell apoptosis in the two PC
cell lines following stable transfection. As shown in Fig. 4, the apoptosis rates of cells that
were stably transfected with Pbk-CMV-HA-EBP50 were markedly higher
compared with those of the cells in the corresponding untreated and
control groups. These data further support the hypothesis that
EBP50 overexpression promotes apoptosis in PC cells.
EBP50 overexpression promotes the
apoptosis rate of PC cells by altering the expression level of the
Bcl-2 protein
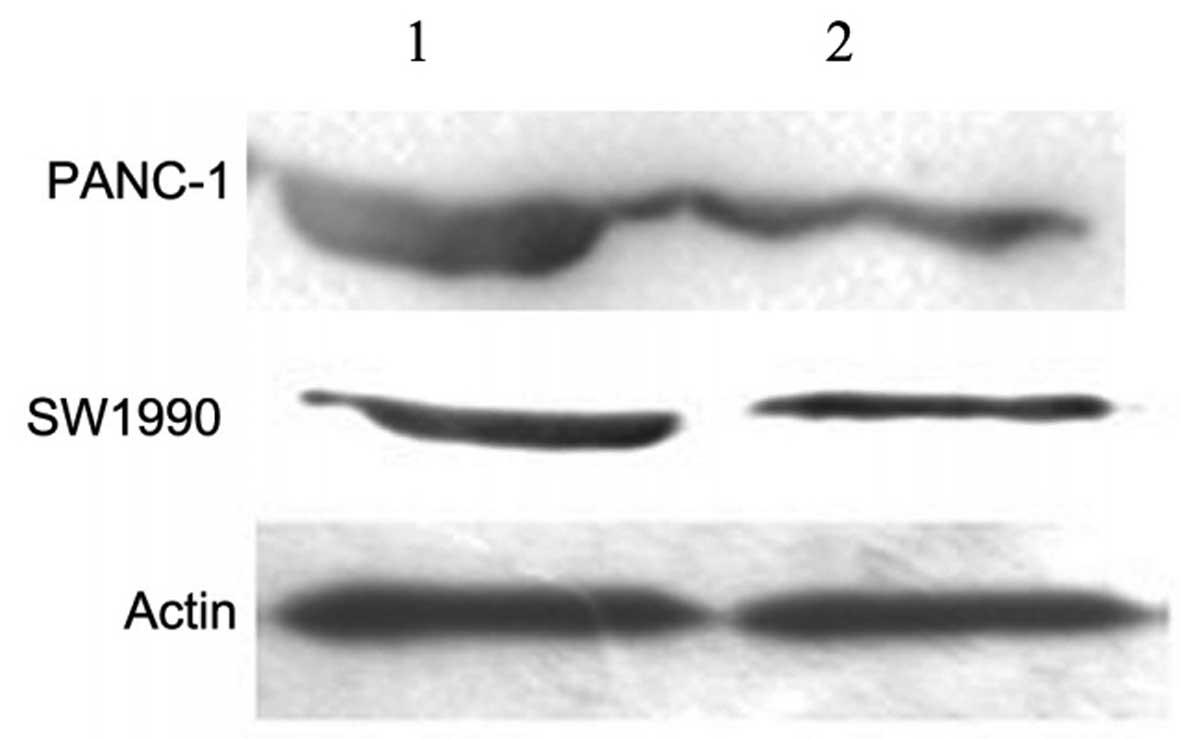
To explore the molecular mechanisms underlying the
anticancer effect of Pbk-CMV-HA-EBP50, western blot analysis was
carried out to determine the expression levels of Bcl-2, an
anti-apoptosis protein, in the PC cells. The results revealed that
Bcl-2 was markedly suppressed in the two PC cell lines when EBP50
was overexpressed (Fig. 5). These
data suggest that the anticancer effects observed during EBP50
overexpression occur due to the suppression of Bcl-2
expression.
EBP50 overexpression inhibits the growth
of PC cells and promotes cell apoptosis in vivo
To detect the therapeutic efficacy of EBP50
overexpression in vivo, mice were randomly divided into six
groups. Six tumor xenograft nude mice models were established by
subcutaneously injecting six types of PC cells. After 36 days, the
tumors in the EBP50-PANC-1-treated mice were significantly smaller
compared with those in the HA-PANC-1-treated mice and
PANC-1-treated mice (all P<0.01); however, the difference in
tumor sizes between the HA-PANC-1 and PANC-1 groups was not
statistically significant (Fig.
6). Similarly, the tumor sizes of the EBP50-SW1990-treated mice
were smaller than the tumors in the corresponding control and
untreated groups. The tumor sizes of mice in the SW-1990 untreated
and control groups were similar. The in vivo results of the
present study indicate that EBP50 overexpression was able to
inhibit the growth of PC tumors.
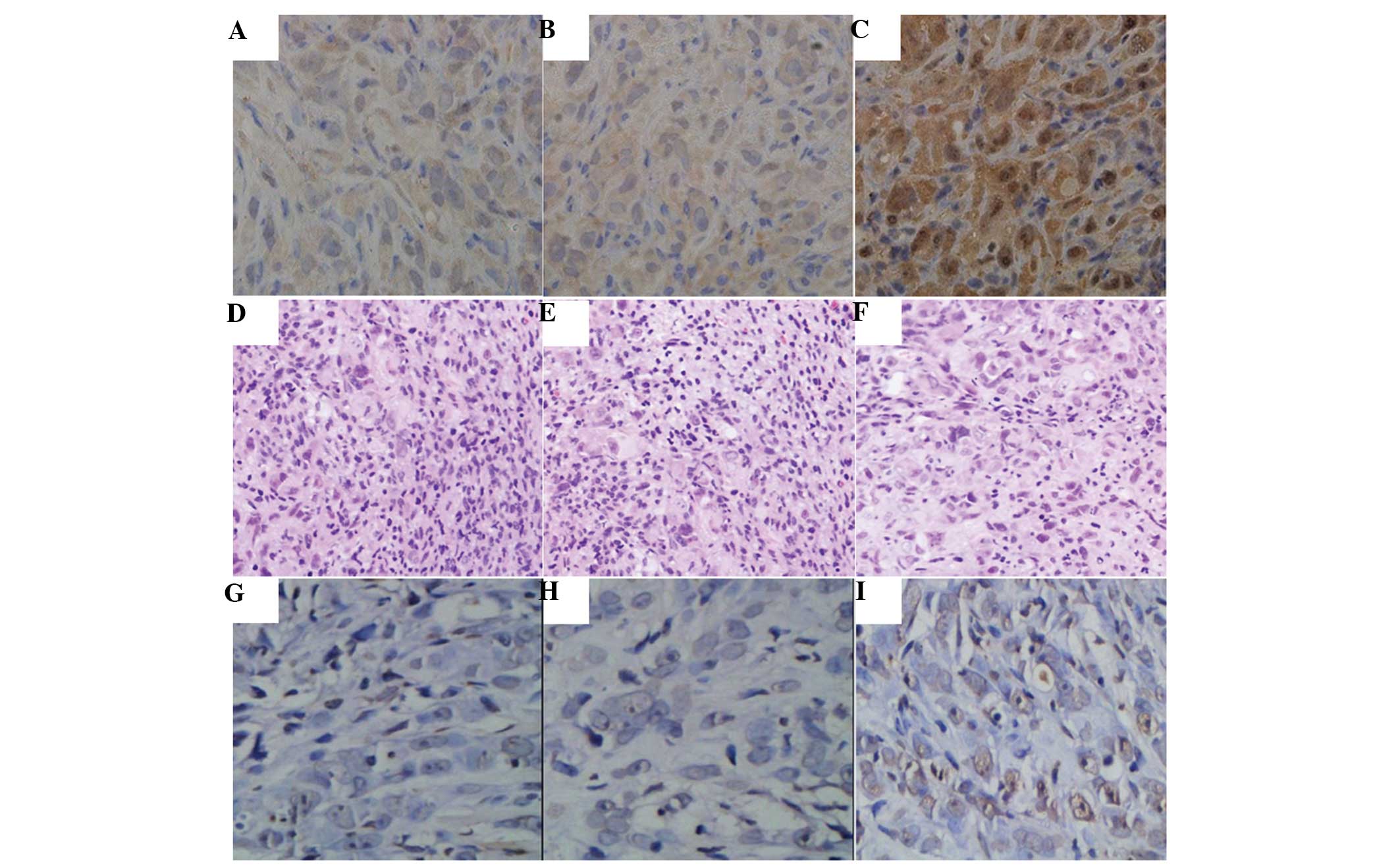
Immunohistochemical analysis revealed that the
expression levels of EBP50 were significantly higher in the
EBP50-PANC-1 cell tumors compared with those in the tumors treated
with negative control (HA-PANC-1) or untreated cells (PANC-1)
(Fig. 7). H&E staining results
demonstrated that there was a compact mass of epithelial cells in
the untreated and control group tumors (Fig. 7). However, the EBP50-PANC-1 cell
tumors appeared as loose epithelial cells with scattered apoptotic
cells characterized by dark shrunken cytoplasms and purple pyknotic
nuclei. The TUNEL assay data revealed that tumors in which EBP50
was overexpressed had a significantly higher apoptosis rate
compared with that of tumors that were treated with the negative
control or untreated cells (Fig.
7), indicating that EBP50 overexpression promoted apoptosis
in vivo.
Discussion
The current study revealed that EBP50 exhibits
significant antitumor effects in PC cells in vitro and in
vivo. The data demonstrated that EBP50 inhibited the
proliferation of PC cells and promoted cell apoptosis, altered cell
cycle progression, suppressed the growth of mouse xenograft tumors
and promoted their apoptosis. In tumors with EBP50 overexpression,
the expression levels of Bcl-2 were markedly reduced, consistent
with the presumed mechanism of action of EBP50. The current study
provides the first evidence, to the best of our knowledge, to
suggest that EBP50 overexpression may suppress the tumorigenicity
of PC in vivo and in vitro by decreasing the
expression levels of Bcl-2.
A previous study reported that the downregulation of
EBP50 promoted PC cell proliferation, increased the colony-forming
ability of cells and accelerated G1-to-S phase progression
(16). In the present study, EBP50
overexpression significantly induced growth inhibition, G1-to-S
cell cycle arrest and cell apoptosis in two PC cell lines.
Furthermore, it was revealed that the expression levels of Bcl-2
were significantly reduced in the cells with EBP50 overexpression.
The marked effect of EBP50 overexpression in vitro prompted
further study to detect its effect in vivo. It was
demonstrated that the overexpression of EBP50 exhibited a notable
effect on growth inhibition and apoptosis in EBP50-PANC-1 and
EBP50-SW1990 tumors through a reduction in the expression levels of
Bcl-2.
The expression of EBP50 (3,6,8) has
been reported in a number of human tumors, including breast and
liver cancers. The majority of these studies have demonstrated that
EBP50 may have an anticancer effect in human cancers. One study
revealed that EBP50 was able to interact with phosphatase and
tensin homolog (PTEN) to exert an inhibitory effect on the
phosphoinositide-3 kinase (PI3K)/Akt pathway (22). Another study demonstrated that
overexpression of EBP50 decreased the colony-formation ability of
cells and inhibited cell proliferation through the suppression of
extracellular-signal-regulated kinase (ERK) activity (23). The binding of EBP50 to the
epidermal growth factor receptor (EGFR) and neurofibromin 2 (NF2)
may also result in tumor suppression (24). EBP50 may also form ternary
complexes with platelet-derived growth factor (PDGF) and NF2 to
generate anticancer effects (25).
The results of the current study are consistent with published data
on the role of EBP50 in breast tumors (6).
The present study indicated that high expression
levels of EBP50 are associated with a lower malignant potential of
PC tumors. Thus, it is proposed that EBP50 expression may be a
valid method of combating PC.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Mackenzie GG, Huang L, Alston N, et al:
Targeting mitochondrial STAT3 with the novel phospho-valproic acid
(MDC-1112) inhibits pancreatic cancer growth in mice. PLoS One.
8:e615322013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Georgescu MM, Morales FC, Molina JR and
Hayashi Y: Roles of NHERF1/EBP50 in cancer. Curr Mol Med.
8:459–468. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ghaneh P, Costello E and Neoptolemos JP:
Biology and management of pancreatic cancer. Postgrad Med J.
84:478–497. 2008. View Article : Google Scholar
|
|
5
|
Clapéron A, Guedj N, Mergey M, et al: Loss
of EBP50 stimulates EGFR activity to induce EMT phenotypic features
in biliary cancer cells. Oncogene. 31:1376–1388. 2012.PubMed/NCBI
|
|
6
|
Sun C, Zheng J, Cheng S, Feng D and He J:
EBP50 phosphorylation by Cdc2/Cyclin B kinase affects actin
cytoskeleton reorganization and regulates functions of human breast
cancer cell line MDA-MB-231. Mol Cells. 36:47–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kreimann EL, Morales FC, de Orbeta-Cruz J,
et al: Cortical stabilization of beta-catenin contributes to
NHERF1/EBP50 tumor suppressor function. Oncogene. 26:5290–5299.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shibata T, Chuma M, Kokubu A, Sakamoto M
and Hirohashi S: EBP50, a beta-catenin-associating protein,
enhances Wnt signaling and is over-expressed in hepatocellular
carcinoma. Hepatology. 38:178–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee YJ, Choi IK, Sheen YY, Park SN and
Kwon HJ: Identification of EBP50 as a specific biomarker for
carcinogens via the analysis of mouse lymphoma cellular proteome.
Mol Cells. 33:309–316. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lv XG, Ji MY, Dong WG, et al: EBP50 gene
transfection promotes 5-fluorouracil-induced apoptosis in gastric
cancer cells through Bax- and Bcl-2-triggered mitochondrial
pathways. Mol Med Rep. 5:1220–1226. 2012.
|
|
11
|
Molina JR, Morales FC, Hayashi Y, Aldape
KD and Georgescu MM: Loss of PTEN binding adapter protein NHERF1
from plasma membrane in glioblastoma contributes to PTEN
inactivation. Cancer Res. 70:6697–6703. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fraenzer JT, Pan H, Minimo L Jr, Smith GM,
Knauer D and Hung G: Overexpression of the NF2 gene inhibits
schwannoma cell proliferation through promoting PDGFR degradation.
Int J Oncol. 23:1493–1500. 2003.PubMed/NCBI
|
|
13
|
Morales FC, Takahashi Y, Kreimann EL and
Georgescu MM: Ezrin-radixin-moesin (ERM)-binding phosphoprotein 50
organizes ERM proteins at the apical membrane of polarized
epithelia. Proc Natl Acad Sci USA. 101:17705–17710. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin YY, Hsu YH, Huang HY, et al: Aberrant
nuclear localization of EBP50 promotes colorectal carcinogenesis in
xenotransplanted mice by modulating TCF-1 and β-catenin
interactions. J Clin Invest. 122:1881–1894. 2012.PubMed/NCBI
|
|
15
|
Morales FC, Hayashi Y, van Pelt CS and
Georgescu MM: NHERF1/EBP50 controls lactation by establishing basal
membrane polarity complexes with prolactin receptor. Cell Death
Dis. 3:e3912012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ji MY, Fan DK, Lv XG, Peng XL, Lei XF and
Dong WG: The detection of EBP50 expression using quantum dot
immunohistochemistry in pancreatic cancer tissue and down-regulated
EBP50 effect on PC-2 cells. J Mol Histol. 43:517–526. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pan Y, Wang L and Dai JL: Suppression of
breast cancer cell growth by Na+/H+ exchanger
regulatory factor 1 (NHERF1). Breast Cancer Res. 8:R632006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo J, Gao J, Li Z, et al: Adenovirus
vector-mediated Gli1 siRNA induces growth inhibition and apoptosis
in human pancreatic cancer with Smo-dependent or Smo-independent Hh
pathway activation in vitro and in vivo. Cancer Lett.
339:185–194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu WJ and Liu HL: Induction of pancreatic
cancer cell apoptosis, invasion, migration, and enhancement of
chemotherapy sensitivity of gemcitabine, 5-FU, and oxaliplatin by
hnRNP A2/B1 siRNA. Anticancer Drugs. 24:566–576. 2013.PubMed/NCBI
|
|
20
|
Li X, Yan J, Wang L, et al: Beclin1
inhibition promotes autophagy and decreases gemcitabine-induced
apoptosis in Miapaca2 pancreatic cancer cells. Cancer Cell Int.
13:262013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boyer C, Teo J, Phillips P, et al:
Effective delivery of siRNA into cancer cells and tumors using
well-defined biodegradable cationic star polymers. Mol Pharm.
10:2435–2444. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Morales FC, Takahashi Y, Momin S, Adams H,
Chen X and Georgescu MM: NHERF1/EBP50 head-to-tail intramolecular
interaction masks association with PDZ domain ligands. Mol Cell
Biol. 27:2527–2537. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hayashi Y, Molina JR, Hamilton SR and
Georgescu MM: NHERF1/EBP50 is a new marker in colorectal cancer.
Neoplasia. 12:1013–1022. 2010.PubMed/NCBI
|
|
24
|
Li X, Xu WM, Yin TL, Zhao QH, Peng LY and
Yang J: Temporal and spatial regulation of
ezrin-radixin-moesin-binding phosphoprotein-50-kDa (EBP50) during
embryo implantation in mouse uterus. Int J Mol Sci. 13:16418–16429.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yao W, Feng D, Bian W, et al: EBP50
inhibits EGF-induced breast cancer cell proliferation by blocking
EGFR phosphorylation. Amino Acids. 43:2027–2035. 2012. View Article : Google Scholar : PubMed/NCBI
|