Introduction
There are numerous inflammatory lung diseases (ILDs)
that lack effective treatments aimed at the underlying causes,
including adult respiratory distress syndrome, chronic obstructive
pulmonary disease, idiopathic pulmonary fibrosis and asthma. ILDs
are characterized by excessive inflammatory cell infiltration and
the overproduction of inflammatory mediators, including tumor
necrosis factor (TNF)-α, interleukin (IL)-1β and IL-6 (1–3).
ILD can be triggered by lipopolysaccharide (LPS), a
cell wall component unique to gram-negative bacteria. A number of
studies have shown that LPS can promote the development of ILD by
stimulating immunocyte infiltration and inducing the production of
inflammatory mediators (4–6).
Regulation of cytokine production in alveolar
macrophages is mediated by the mitogen-activated protein kinase
(MAPK) signaling pathway. This mechanism is crucial for the defense
against bacterial infection; however, excessive production of these
cytokines can cause lung injury. Therefore, reducing the release of
inflammatory cytokines by inhibiting the MAPK pathway may alleviate
lung injury (7).
In addition to the MAPK pathway, signal transducer
and activator of transcription-3 (STAT3), a member of the
cytoplasmic family of STATs, is also key to the development of
ILDs, such as acute lung injury (ALI) (8). In previous studies, LPS was
demonstrated to induce TNF-α expression in rat lungs, the spleen
and alveolar macrophages from bronchoalveolar lavage fluid, and in
these processes p38 MAPK and STAT3 signaling pathways were found to
be involved (9–11).
In the present study, the interactions between the
MAPK and STAT3 pathways and their roles in LPS-induced TNF-α and
IL-10 production were investigated in cultured mouse alveolar
macrophages (MH-S cells).
Materials and methods
Cells
MH-S cell lines purchased from the American Type
Culture Collection (Manassas, VA, USA) were used for all the
experiments. RPMI-1640 medium was purchased from Gibco Life
Technologies (Grand Island, NY, USA). Fetal bovine serum (FBS),
heat-inactivated and determined to be LPS-free, was purchased from
Atlanta Biologicals, Inc. (Atlanta, GA, USA). LPS was purchased
from Sigma-Aldrich (St Louis, MO, USA) and SB203580 (p38 MAPK
inhibitor) was purchased from Promega Corporation (Madison, WI,
USA). A rabbit polyclonal anti-tyrosine (Tyr 705) phosphorylated
STAT3 (p-STAT3) antibody was used at a 1:500 dilution and was
purchased from Santa Cruz Biotechnology, Inc. (sc-7993-R, Dallas,
TX, USA).
Culture and stimulation of MH-S
cells
MH-S cells were grown in medium in 75-cm2
polystyrene tissue culture flasks. During log-phase growth, the
cells were scraped off the flasks and the cell suspension was added
to 24-well polystyrene tissue culture plates at 5×106
cells/ml culture medium. The study included two experiments.
Experiment 1
After 24 h of growth at 37°C in 5% CO2,
the cells were preincubated with SB203580 (5, 10 or 15 μM) for 20
min in 10% FBS-containing media, followed by the addition of LPS at
a final concentration of 100 ng/ml. A blank control and
LPS-stimulated control were included. Following stimulation for
1.5, 2, 4, 6 or 12 h, the supernatant was collected for subsequent
TNF-α and IL-10 analyses.
Experiment 2
SB203580 (10 μM) was added to the cells 20 min prior
to LPS (100 ng/ml) stimulation for 15 or 30 min. The cells were
then collected and centrifuged (513 × g, 4°C, 10 min). The total
protein extracts were stored at −80°C for examination of STAT3
phosphorylation by western blot analysis. Phosphorylation of STAT3
expression was also examined by immunocytochemistry.
Examination of TNF-α, IL-10 and
STAT3
Levels of TNF-α and IL-10 in LPS-stimulated MH-S
cell lines were measured by enzyme-linked immunosorbent assays
(RayBiotech,Inc.Norcross, GA, USA), with or without the p38
inhibitor, SB203580. Total STAT3 and p-STAT3 expression levels were
examined by western blot analysis and immunocytochemistry. In
brief, the procedure for the western blot analysis was the
following: The cells were stimulated with LPS (100 ng/ml) for 15 or
30 min and the total cellular protein was extracted. To detect the
phosphorylated STAT3, a rabbit polyclonal anti-tyrosine (Tyr 705)
phosphorylated STAT3 antibody (Santa Cruz Biotechnology, Inc.
sc-7993-R) was used as the primary antibody and anti-rabbit IgG
secondary antibody (Beijing DingGuo biotechnology) was used. For
the immunocytochemistry experiments the MH-S cells (5 ×
105/ml) were seeded on cover slips and cultured in a
6-well plate. The following day the cover slips were removed and
fixed with 4 % paraformaldehyde solution at room temperature. In
order to detect the p-STAT3 expression, the same antibodies
described for the western blot analysis were used.
Statistical analysis
Data are expressed as the mean ± standard deviation.
The group differences were analyzed by one-way analysis of variance
using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). If
significant, the data were further analyzed by the
Student-Newman-Keuls test, where P<0.05 was considered to
indicate a statistically significant difference.
Results
Changes in TNF-α and IL-10 levels in the
MH-S cell supernatant
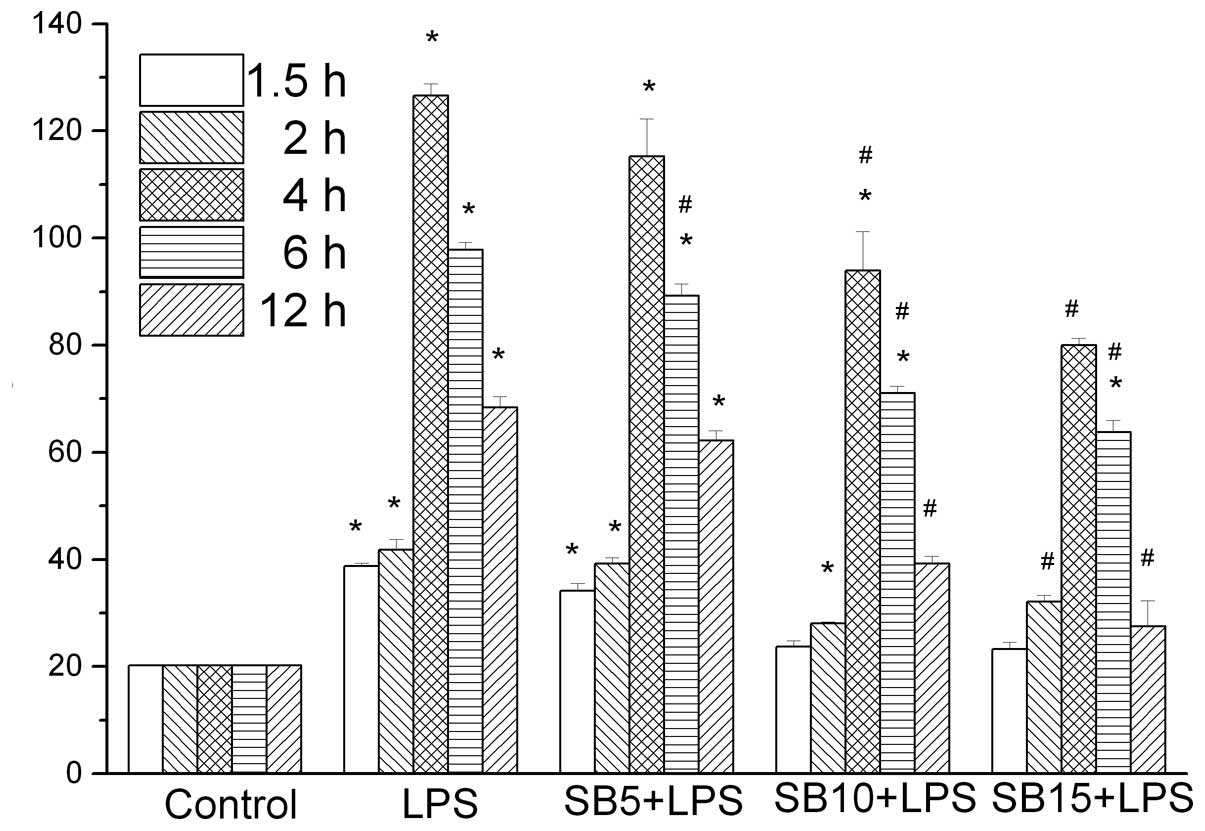
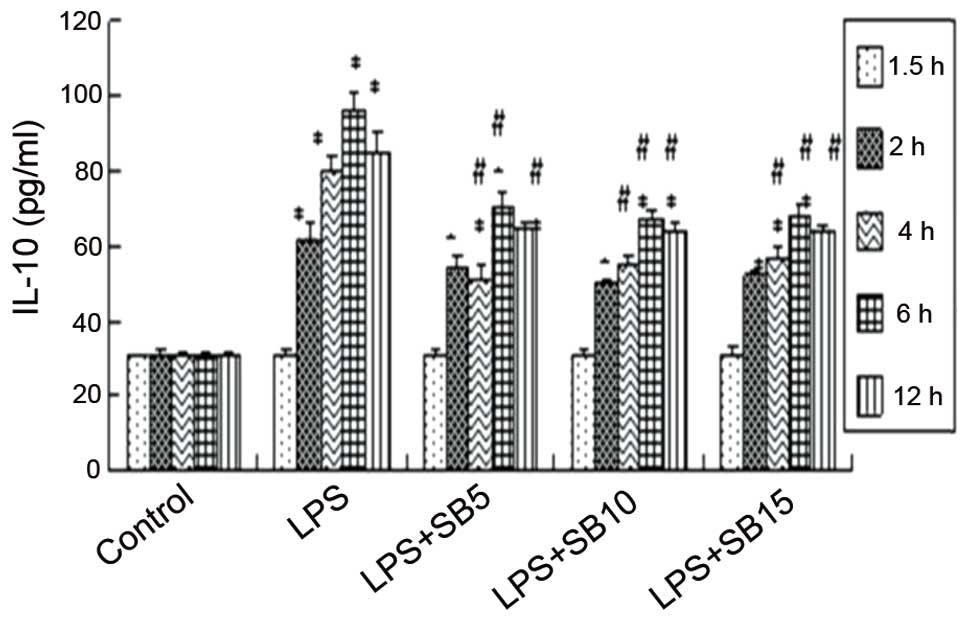
TNF-α and IL-10 levels exhibited an increase at 90
min following LPS stimulation, reaching peak levels at 4 and 6 h,
respectively. This trend was not observed in the control group
(P<0.05). SB203580 treatment was shown to inhibit TNF-α in a
dose-dependent manner (Fig. 1).
Cells treated with 15 μM SB203580 produced significantly less TNF-α
(P<0.05) compared with the SB203580 naive cells (Fig. 1). SB203580 treatment also inhibited
IL-10; however, the dose-dependent inhibition was not significant.
Furthermore, cells treated with 15 μM SB203580 produced
significantly higher levels of TNF-α and IL-10 (Fig. 2) compared with the non-stimulated
cells, indicating that pathways other than p38 may also mediate
TNF-α production.
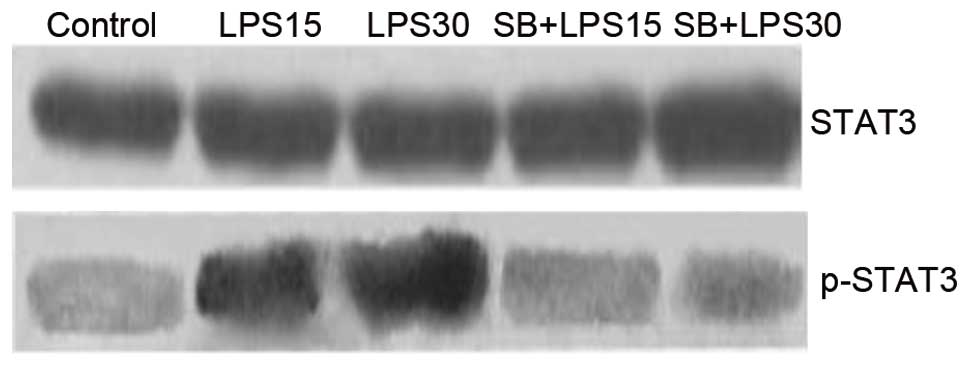
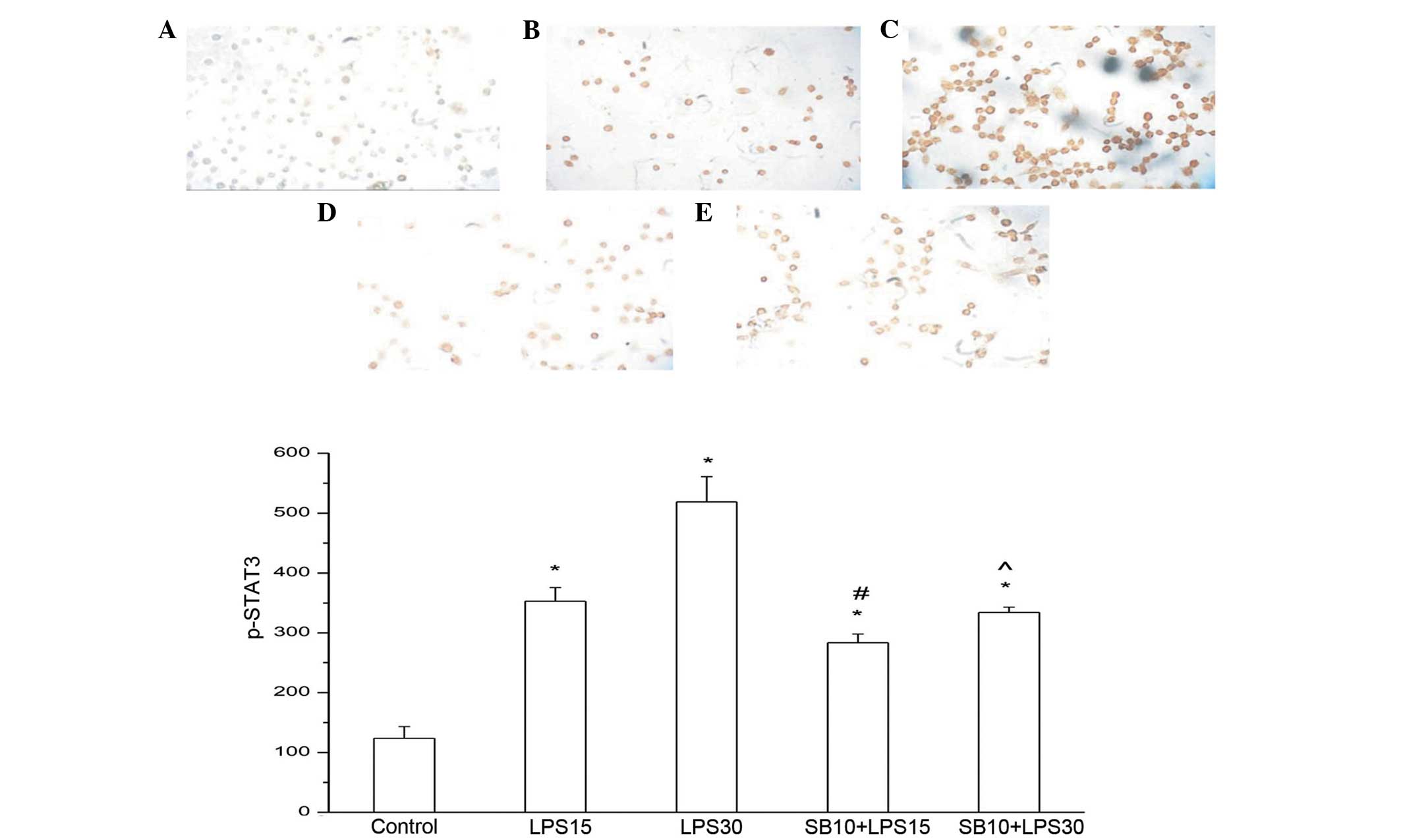
By contrast, the levels of p-STAT3 protein exhibited
a significant increase 15 min after LPS induction (P<0.05), and
peaked 30 min after treatment. SB203580 treatment almost completely
blocked STAT3 phosphorylation, even at a concentration of 10 μM,
indicating that p38 is a key regulator of STAT3 phosphorylation
(Figs. 3 and 4).
Discussion
The results of the present study demonstrated that
in the MH-S cell supernatant, TNF-α levels increased following LPS
stimulation and decreased following the addition of SB203580. The
p-STAT3 expression levels increased in the LPS group, while
SB203580 was found to inhibit the expression of p-STAT3, suggesting
an interaction between p38 MAPK and STAT3.
ILDs are triggered by excessive inflammation, as
exemplified by the overproduction of inflammatory mediators;
however, the underlying mechanism is not fully understood.
LPS-induced inflammation is an appropriate model since LPS
principally acts on alveolar macrophages, causing a massive release
of inflammatory cytokines. The cytokines and the cells that are
involved in LPS-induced inflammation, have been basically defined;
however, the mechanism by which these cytokines are expressed and
regulated remains unclear (12,13).
A previous study demonstrated that macrophage deficiency
significantly attenuated TNF-α production in the lung tissues of
mice (14).
The macrophage MAPK pathway is the key regulator of
inflammation, regulating the gene expression of a variety of
cytokines, and playing an important role in inflammation. Previous
studies revealed that p38 MAPK was the basic signaling pathway in
the regulation of LPS-induced TNF-α synthesis (9–11).
LPS-induced TNF-α and IL-6 secretion, as well as neutrophil
aggregation, protein leakage and bronchiostenosis, in the lungs of
mice was found to be dependent on p38 MAPK signaling (15). Activation of extracellular
signal-regulated kinase (ERK), p38, c-Jun N-terminal kinase (JNK)
and nuclear factor-κB (NF-κB) was shown to be involved in
LPS-induced TNF-α production in human monocytes. p38α is an
important regulator of inflammatory responses, and p38α deficiency
in macrophages has been shown to cause a significant inhibition in
the production of LPS-induced TNF-α, IL-12 and IL-18 (16). However, a p38α deficiency was not
shown to affect the LPS-induced activation of other major signaling
pathways (NF-κB, JNK and ERK), nor the transcriptional activity of
NF-κB (17). p38 MAPK modulates
TNF-α transcription in LPS-activated primary human macrophages,
which is mediated through p38 MAPK regulation of NF-κB. The
regulation of NF-κB by p38 MAPK is cell-type dependent and this may
have consequences for the anti-inflammatory efficacy of inhibitors
of p38 MAPK (18). The present
study clarified the role of p38 MAPK in LPS-induced inflammation
and provided valuable data that may lead to effective treatment
strategies for ILDs.
p38 MAPK, the primary intracellular signaling
pathway that regulates LPS-induced TNF-α biosynthesis (19), can be blocked by a specific p38
MAPK inhibitor, SB203580. Under normal conditions, SB203580 is able
to completely inhibit LPS-induced TNF-α expression by the RAW 264.7
mouse macrophage cell line; however, it lacks this inhibitory
effect under hypoxic conditions (19).
Zhao et al (20) stimulated MH-S cells with LPS for
different time periods and assessed the activation kinetics of ERK
and p38 MAPK. In control alveolar macrophages, TNF-α levels
increased while p38 activity was virtually undetectable. Following
LPS stimulation, p38 was rapidly activated. Similarly, ERK was also
potently activated in response to LPS stimulation, with kinetics
similar to that of p38 activation. In the present study, MH-S cells
were stimulated with LPS for 1.5, 2, 4, 6 and 12 h, and TNF-α
production was shown to increase significantly, which is consistent
with the results reported by Zhao et al (20). Although certain studies have
demonstrated that p38 MAPK is a potential target in regulating
TNF-α, the functions of p38 MAPK in different tissues should be
further investigated (20–23). A previous study revealed that p38
MAPK and NF-κB were involved in LPS-induced TNF-α gene and protein
expression in the rat spleen (11). The present study demonstrated that
TNF-α expression in the supernatant was reduced by SB203580 in a
dose-dependent manner, which indicated that in MH-S cells, SB203580
is able to decrease the secretion of TNF-α by inhibiting p38 MAPK,
attenuating the LPS-induced inflammatory response.
The prognosis of ILD is hypothesized to depend on
the balance between proinflammatory and anti-inflammatory
cytokines. The STAT family is a key regulator of cytokine
production, but there is limited knowledge of its role in mediating
ILDs. STAT3 is an important member of the JAK-STAT pathway, and is
widely expressed in a number of cell and tissue types. The
activation of STAT3 has been implicated in the regulation of cell
proliferation, differentiation, transformation, apoptosis and
inflammation (24,25). STATs may be involved as a pathway
in mediating ALI, regardless of the inciting factors (26). The STAT family has been reported to
be expressed in ALI, while STAT3 has been shown to function as an
anti-inflammatory protein with a protective role in ALI (27). STAT3 activation has been associated
with suppressed inflammatory processes in experimental animals,
murine myeloid cells and macrophage cell lines. Therefore,
manipulation of STAT3 activation may facilitate the development of
new pharmacological interventions in human inflammatory diseases.
However, STAT3 activation is unable to directly regulate LPS
signaling in human monocytes and may only represent part of the
mechanism by which IL-10 suppresses TNF-α production in activated
human monocytes (28). In a murine
model of acute peritonitis, resident macrophages, but not other
cell types, were shown to play a regulatory role in inflammation
through a STAT3 signaling pathway (29). STAT3 appears to function as a
repressor protein in this model of acute inflammation; however,
STAT3 in other cell types may contribute to the production of TNF-α
and macrophage inflammatory protein-2 (27). The present study demonstrated that
p38 was a key regulator of STAT3 phosphorylation in mouse alveolar
macrophages. STAT3 is activated by Tyr 705 and Ser 727
phosphorylation. Depletion of alveolar macrophages markedly reduced
the extent of lung STAT3 activation and decreased the expression of
a number of cytokines in the lungs, including IL-1β, IL-4, IL-6,
IL-10 and TNF-α (30). Macrophages
lacking STAT3 showed an abnormal activation phenotype, such as
increased cytokine production as a result of endotoxin-induced
inflammation.
One possible role of STATs in mediating lung injury
is the upregulation of cytokine gene expression, with a previous
study demonstrating that STAT3 is required for LPS-induced TNF-α
expression in macrophages (31).
The target genes of STATs may promote the pathological development
of ALI, including cytokines, chemical factors, adhesion molecules
and inflammatory regulators. STATs may increase LPS signaling
molecules, including lipopolysaccharide binding protein and MD-2,
indicating that STATs may expand the inflammatory response in the
development of sepsis (32).
Severgnini et al (26)
reported that in LPS-stimulated mice, STAT3 was activated and TNF-α
production increased. STAT3 signaling was reported to play a
crucial role in the downregulation of TNF-α synthesis by human
monocytes in the course of systemic inflammation in vivo,
which suggests that STAT3 may be a potential molecular target for
pharmacological intervention in clinical syndromes characterized by
systemic inflammation (33).
Blocking TNF-α signaling significantly attenuated LPS-mediated ALI,
indicating that TNF-α was a plausible STAT target gene associated
with ALI. LPS-induced MAPK activation, the production of endogenous
IL-10, and STAT3 activation have been reported to play critical
roles in the expression of suppressor of cytokine signaling 3
(SOCS3), which provides for feedback attenuation of
cytokine-induced immune and inflammatory responses in macrophages
(34). The results of the present
study showed that TNF-α levels in the supernatant began to increase
when the MH-S cells were stimulated for 90 min, while STAT3
phosphorylation increased with LPS stimulation for 15 min. These
observations suggested that TNF-α may be regulated by STAT3, which
is consistent with the findings of Severgnini et al
(26).
Zhao et al (20) reported that the LPS-induced
increase in TNF-α in MH-S cells occurred through the p38 MAPK
pathway. The present study showed that the TNF-α levels increased
following stimulation with LPS in MH-S cells, and decreased when
SB203580 was added. However, further studies should investigate
whether the TNF-α changes were dependent on STAT3, and whether
STAT3 functions as a pro- or anti-inflammatory protein. Data on the
activation and function of STAT3 and SOCS3 in the lungs during the
acute inflammatory response are emerging, suggesting that these
molecules may be potential targets for regulating pulmonary
inflammatory responses (3).
Serine phosphorylation on Ser 727 by MAPK has been
identified, and STAT may be the substrate of MAPK (35). In the present study, the rate of
STAT3 Tyr 705 phosphorylation decreased when SB203580 was added to
the MH-S cells, suggesting that STAT3 and p38 MAPK may be relevant
to this process.
In a previous study (36), RAW 264.7 cells preincubated with
the serum of burn rats treated with a topical p38 inhibitor showed
a significantly lower TNF-α expression level following LPS
stimulation when compared with the vehicle-treated group.
Modulating p38 MAPK signaling in burn wounds was shown to reduce
pulmonary microvascular injury and pulmonary edema (36). SB203580 can reduce the secretion of
TNF-α through p38 MAPK; however, the compound is unable to inhibit
the release of TNF-α in endotoxin shock mice and the subsequent
mortality rate (37). Further
study is required to clarify whether SB203580 can decrease the
mortality rate of ALI.
In conclusion, the present study indicated that
understanding the STAT pathway was critical to reveal the
mechanisms of ILDs, as this pathway significantly influences immune
regulation and the production of several important cytokines, and
closely interacts with other signaling pathways. Understanding the
signaling pathways relevant to specific conditions, including ILDs,
may provide useful information for the development of novel
therapeutic approaches.
References
|
1
|
Todd NW, Luzina IG and Atamas SP:
Molecular and cellular mechanisms of pulmonary fibrosis.
Fibrogenesis Tissue Repair. 5:112012. View Article : Google Scholar
|
|
2
|
Barnes PJ: Alveolar macrophages as
orchestrators of COPD. COPD. 1:59–70. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao H and Ward PA: STAT3 and suppressor of
cytokine signaling 3: potential targets in lung inflammatory
responses. Expert Opin Ther Targets. 11:869–880. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jansson AH, Eriksson C and Wang X: Lung
inflammatory responses and hyperinflation induced by an
intratracheal exposure to lipopolysaccharide in rats. Lung.
182:163–171. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kemp MW, Kannan PS, Saito M, et al:
Selective exposure of the fetal lung and skin/amnion (but not
gastro-intestinal tract) to LPS elicits acute systemic inflammation
in fetal sheep. PLoS One. 8:e633552013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li X, Li Z, Zheng Z, Liu Y and Ma X:
Unfractionated heparin ameliorates lipopolysaccharide-induced lung
inflammation by downregulating nuclear factor-κB signaling pathway.
Inflammation. 36:1201–1208. 2013.PubMed/NCBI
|
|
7
|
Jarrar D, Kuebler JF, Rue LW 3rd, et al:
Alveolar macrophage activation after trauma-hemorrhage and sepsis
is dependent on NF-kappaB and MAPK/ERK mechanisms. Am J Physiol
Lung Cell Mol Physiol. 283:L799–L805. 2002.PubMed/NCBI
|
|
8
|
Tang H, Yan C, Cao J, et al: An essential
role for Stat3 in regulating IgG immune complex-induced pulmonary
inflammation. FASEB J. 25:4292–4300. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ling YL, Meng AH, Zhao XY, et al: Effect
of cholecystokinin on cytokines during endotoxic shock in rats.
World J Gastroenterol. 7:667–671. 2001.PubMed/NCBI
|
|
10
|
Meng AH, Ling YL, Zhang XP and Zhang JL:
Anti-inflammatory effect of cholecystokinin and its signal
transduction mechanism in endotoxic shock rat. World J
Gastroenterol. 8:712–717. 2002.PubMed/NCBI
|
|
11
|
Meng AH, Ling YL, Zhang XP, Zhao XY and
Zhang JL: CCK-8 inhibits expression of TNF-α in the spleen of
endotoxic shock rats and signal transduction mechanism of p38 MAPK.
World J Gastroenterol. 8:139–143. 2002.
|
|
12
|
Song KS, Yoon JH, Kim KS and Ahn DW:
c-Ets1 inhibits the interaction of NF-κB and CREB, and
downregulates IL-1β-induced MUC5AC overproductionduring airway
inflammation. Mucosal Immunol. 5:207–215. 2012.PubMed/NCBI
|
|
13
|
Wijagkanalan W, Kawakami S, Higuchi Y, et
al: Intratracheally instilled mannosylated cationic liposome/NFκB
decoy complexes for effective prevention of LPS-induced lung
inflammation. J Control Release. 149:42–50. 2011.PubMed/NCBI
|
|
14
|
Lomas-Neira J, Chung CS, Perl M, et al:
Role of alveolar macrophage and migrating neutrophils in
hemorrhage-induced priming for ALI subsequent to septic challenge.
Am J Physiol Lung Cell Mol Physiol. 290:L51–L58. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schnyder-Candrian S, Quesniaux VF, Di
Padova F, et al: Dual effects of p38 MAPK on TNF-dependent
bronchoconstriction and TNF-independent neutrophil recruitment in
lipopolysaccharide-induced acute respiratory distress syndrome. J
Immunol. 175:262–269. 2005. View Article : Google Scholar
|
|
16
|
Nishiki S, Hato F, Kamata N, et al:
Selective activation of STAT3 in human monocytes stimulated by
G-CSF: implication in inhibition of LPS-induced TNF-alpha
production. Am J Physiol Cell Physiol. 286:C1302–C1311. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang YJ, Chen J, Otsuka M, et al:
Macrophage deletion of p38alpha partially impairs
lipopolysaccharide-induced cellular activation. J Immunol.
180:5075–5082. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Campbell J, Ciesielski CJ, Hunt AE, et al:
A novel mechanism for TNF-alpha regulation by p38 MAPK: involvement
of NF-kappa B with implications for therapy in rheumatoid
arthritis. J Immunol. 173:6928–6937. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu FQ, Liu Y, Lui VC, Lamb JR, Tam PK and
Chen Y: Hypoxia modulates lipopolysaccharide induced TNF-alpha
expression in murine macrophages. Exp Cell Res. 314:1327–1336.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao Q, Shepherd EG, Manson ME, et al: The
role of mitogen-activated protein kinase phosphatase-1 in the
response of alveolar macrophages to lipopolysaccharide: attenuation
of proinflammatory cytokine biosynthesis via feedback control of
p38. J Biol Chem. 280:8101–8108. 2005. View Article : Google Scholar
|
|
21
|
Ehlting C, Lai WS, Schaper F, et al:
Regulation of suppressor of cytokine signaling 3 (SOCS3) mRNA
stability by TNF-alpha involves activation of the MKK6/p38MAPK/MK2
cascade. J Immunol. 178:2813–2826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li W, Huang H, Zhang Y, Fan T, Liu X, Xing
W and Niu X: Anti-inflammatory effect of tetrahydrocoptisine from
Corydalis impatiens is a function of possible inhibition of TNF-α,
IL-6 and NO production in lipopolysaccharide-stimulated peritoneal
macrophages through inhibiting NF-κB activation and MAPK pathway.
Eur J Pharmacol. 715:62–71. 2013.PubMed/NCBI
|
|
23
|
Soromou LW, Chu X, Jiang L, Wei M, Huo M,
Chen N, Guan S, Yang X, Chen C, Feng H and Deng X: In vitro and in
vivo protection provided by pinocembrin against
lipopolysaccharide-induced inflammatory responses. Int
Immunopharmacol. 14:66–74. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Geiser T: Inflammatory cytokines and
chemokines in acute inflammatory disease. Schweiz Med Wochenschr.
129:540–546. 1999.(In German).
|
|
25
|
Sumita N, Bito T, Nakajima K and Nishigori
C: Stat3 activation is required for cell proliferation and
tumorigenesis but not for cell viability in cutaneous squamouscell
carcinoma cell lines. Exp Dermatol. 15:291–299. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Severgnini M, Takahashi S, Rozo LM, et al:
Activation of the STAT pathway in acute lung injury. Am J Physiol
Lung Cell Mol Physiol. 286:L1282–L1292. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
El Kasmi KC, Holst J, Coffre M, et al:
General nature of the STAT3-activated anti-inflammatory response. J
Immunol. 177:7880–7888. 2006.PubMed/NCBI
|
|
28
|
Prêle CM, Keith-Magee AL, Murcha M and
Hart PH: Activated signal transducer and activator of
transcription-3 (STAT3) is a poor regulator of tumour necrosis
factor-alpha production by human monocytes. Clin Exp Immunol.
147:564–572. 2007.PubMed/NCBI
|
|
29
|
Matsukawa A, Kudo S, Maeda T, Numata K,
Watanabe H, Takeda K, Akira S and Ito T: Stat3 in resident
macrophages as a repressor protein of inflammatory response. J
Immunol. 175:3354–3359. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gao H, Guo RF, Speyer CL, et al: STAT3
activation in acute lung injury. J Immunol. 172:7703–7712. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chappell VL, Le LX, LaGrone L and Mileski
WJ: Stat proteins play a role in tumor necrosis factor alpha gene
expression. Shock. 14:400–403. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Abreu MT, Arnold ET, Thomas LS, et al:
TLR4 and MD-2 expression is regulated by immune-mediated signals in
human intestinal epithelial cells. J Biol Chem. 277:20431–20437.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
de Jong PR, Schadenberg AW, van den Broek
T, et al: STAT3 regulates monocyte TNF-alpha production in systemic
inflammation caused by cardiac surgery with cardiopulmonary bypass.
PLoS One. 7:e350702012.PubMed/NCBI
|
|
34
|
Qin H, Roberts KL, Niyongere SA, Cong Y,
Elson CO and Benveniste EN: Molecular mechanism of
lipopolysaccharide-induced SOCS-3 gene expression in macrophages
and microglia. J Immunol. 179:5966–5976. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Park SY, Baik YH, Cho JH, Kim S, Lee KS
and Han JS: Inhibition of lipopolysaccharide-induced nitric oxide
synthesis by nicotine through S6K1-p42/44 MAPK pathwayand STAT3
(Ser 727) phosphorylation in Raw 264.7 cells. Cytokine. 44:126–134.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ipaktchi K, Mattar A, Niederbichler AD, et
al: Attenuating burn wound inflammatory signaling reduces systemic
inflammation and acute lung injury. J Immunol. 177:8065–8071. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen Y, Kam CS, Liu FQ, et al: LPS-induced
up-regulation of TGF-beta receptor 1 is associated with TNF-alpha
expression in human monocyte-derived macrophages. J Leukoc Biol.
83:1165–1173. 2008. View Article : Google Scholar : PubMed/NCBI
|